RETRACTED: Low Iron Diet Increases Susceptibility to Noise-Induced Hearing Loss in Young Rats

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Iron Status
2.3. Hearing Measurements
2.4. Noise Exposure
2.5. Cochlear Tissue Processing and HE Staining
2.6. Electron Microscopy
2.7. Immunofluorescence
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. ID without Anemia Leads to Growth and Iron Status of Young Rat Effects
3.2. ID without Anemia Reduces ABR Wave I Peak Amplitude of Young Rats
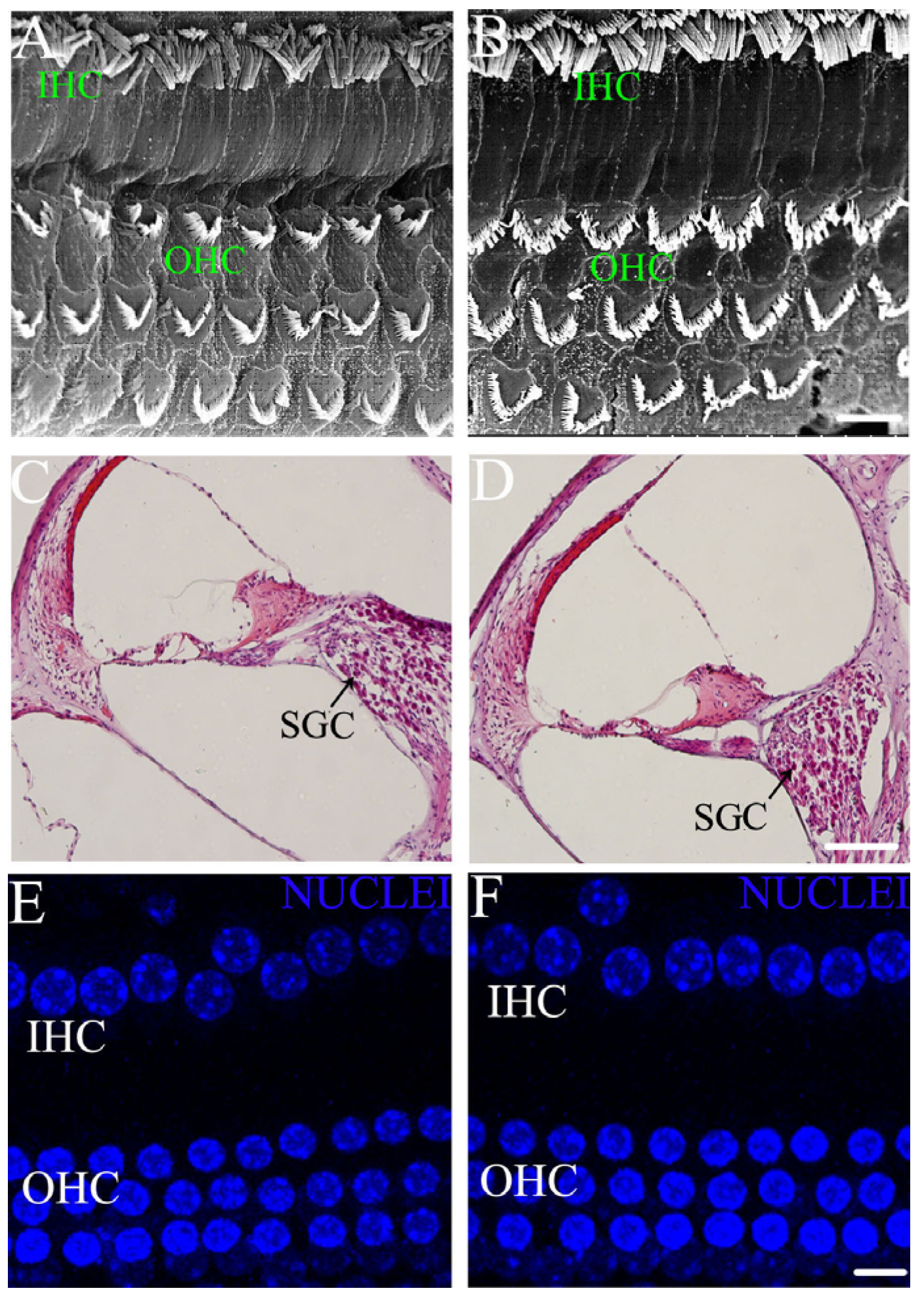
3.3. ID without Anemia Lacks Significant Disruption in Hair Cells and SGCs Morphology of Young Rats
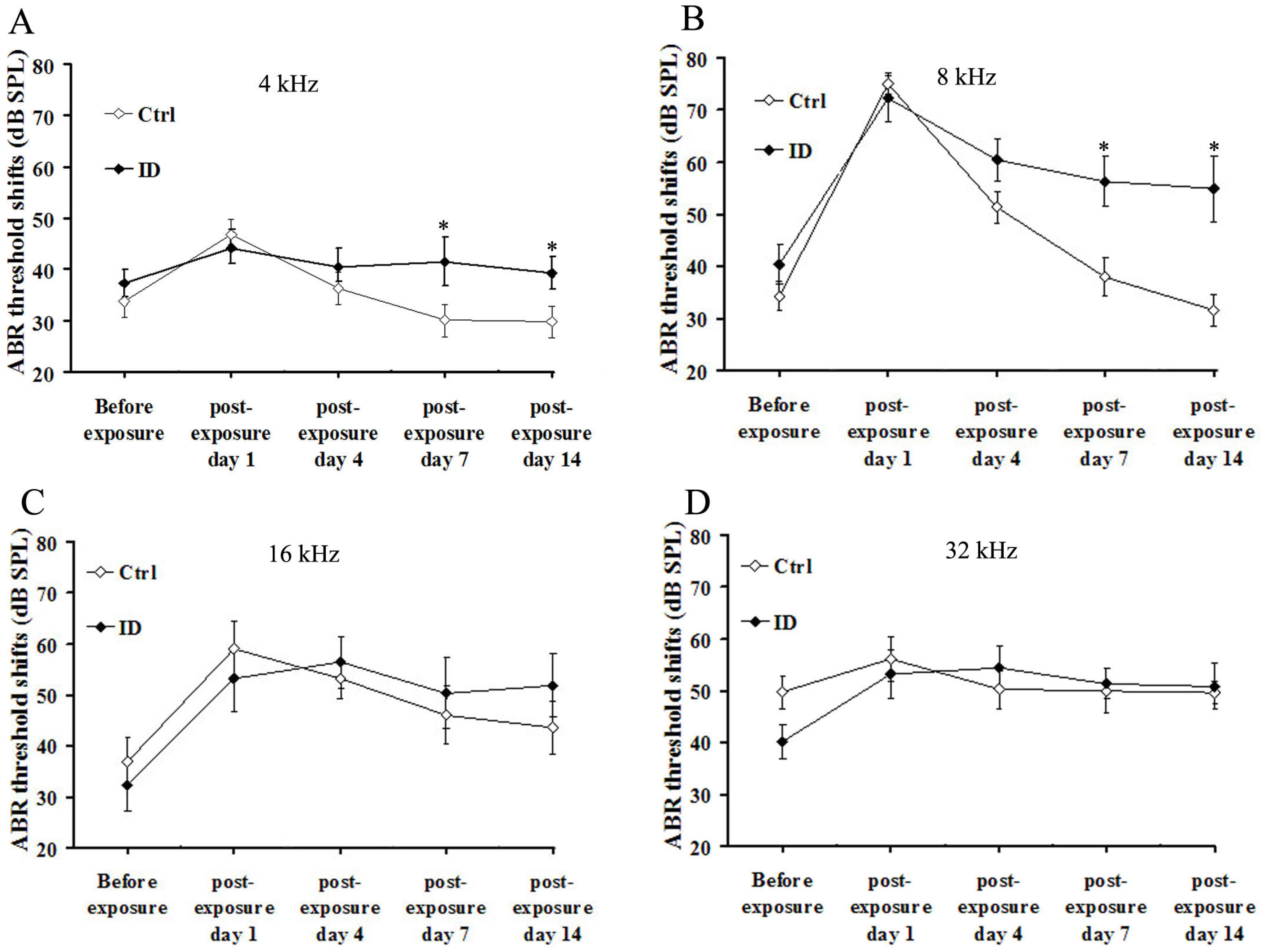
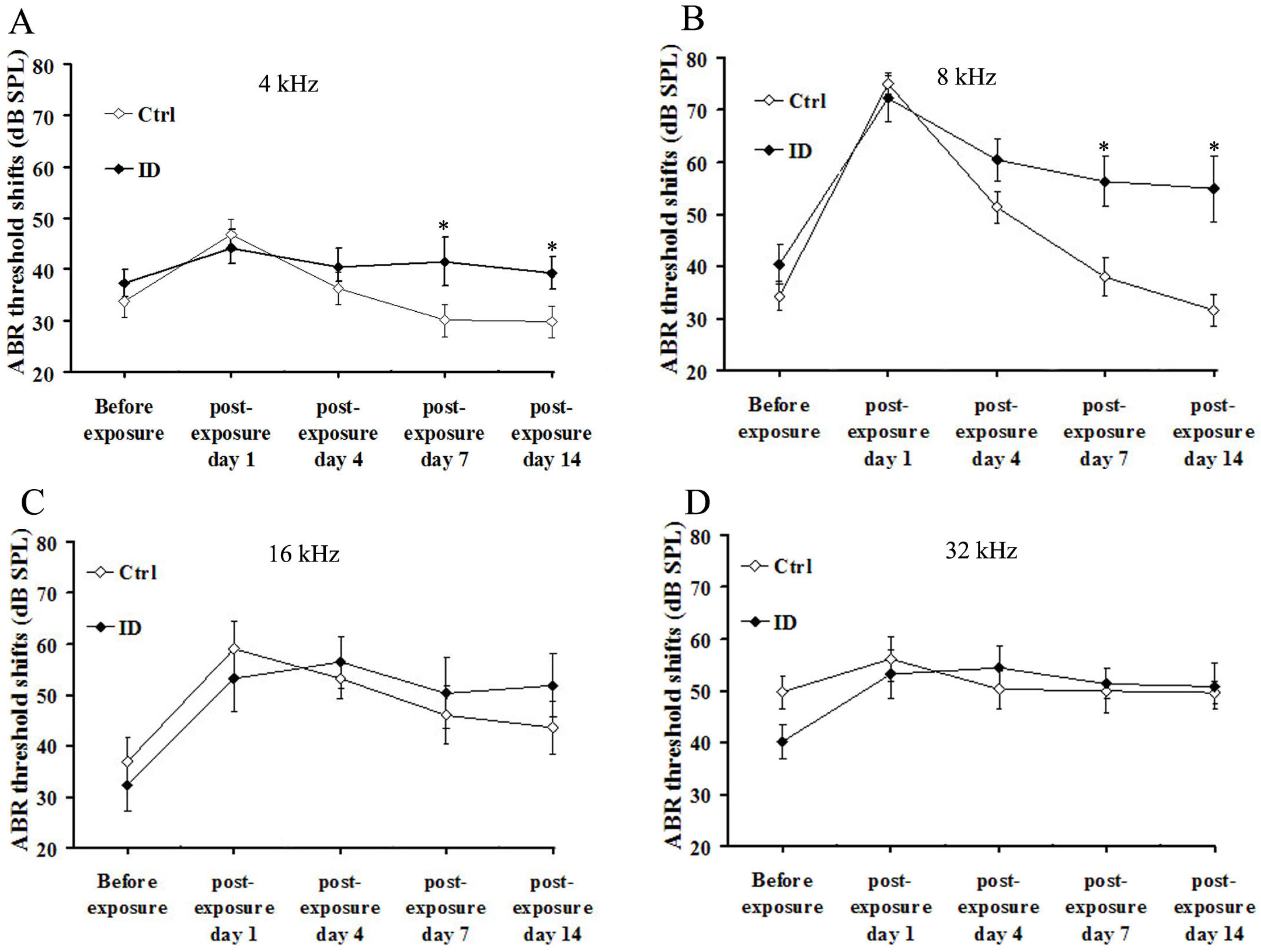
3.4. ID without Anemia Delays the Recovery from Noise-Induced Hearing Loss in Young Rats
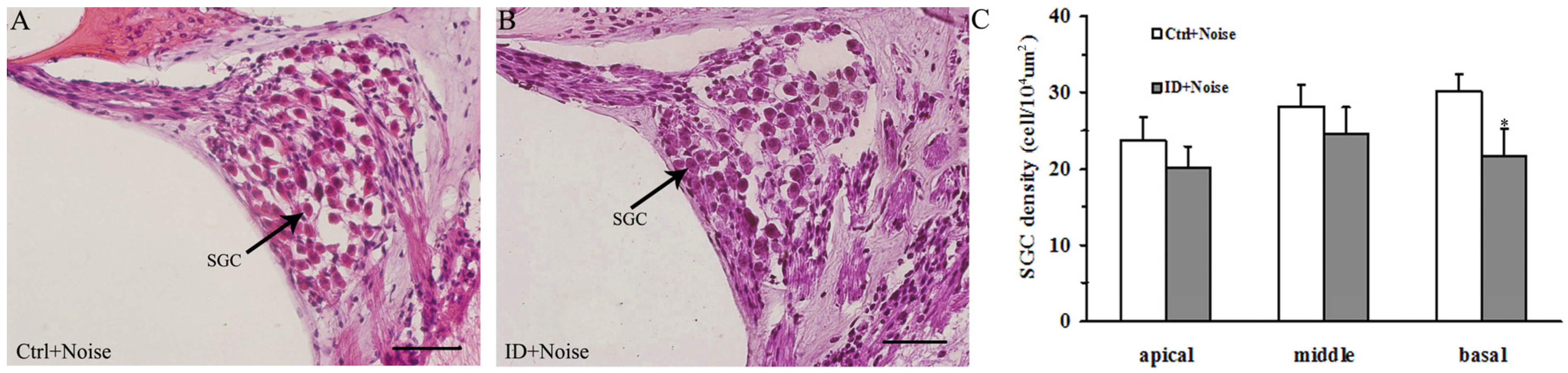
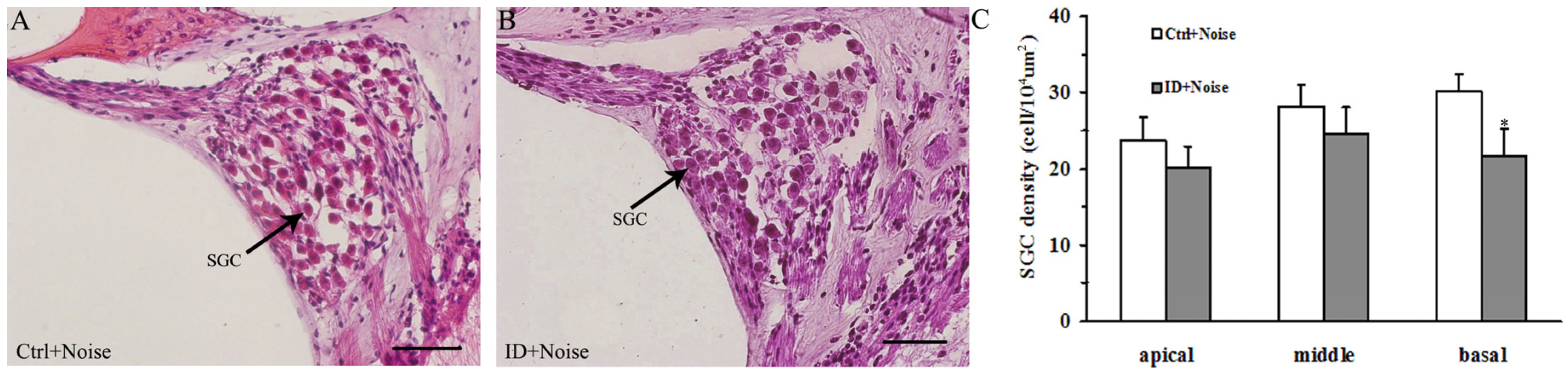
3.5. ID without Anemia Increases SGCs Loss in the Basal Turn of the Cochleae of Young Rats after Noise Exposure
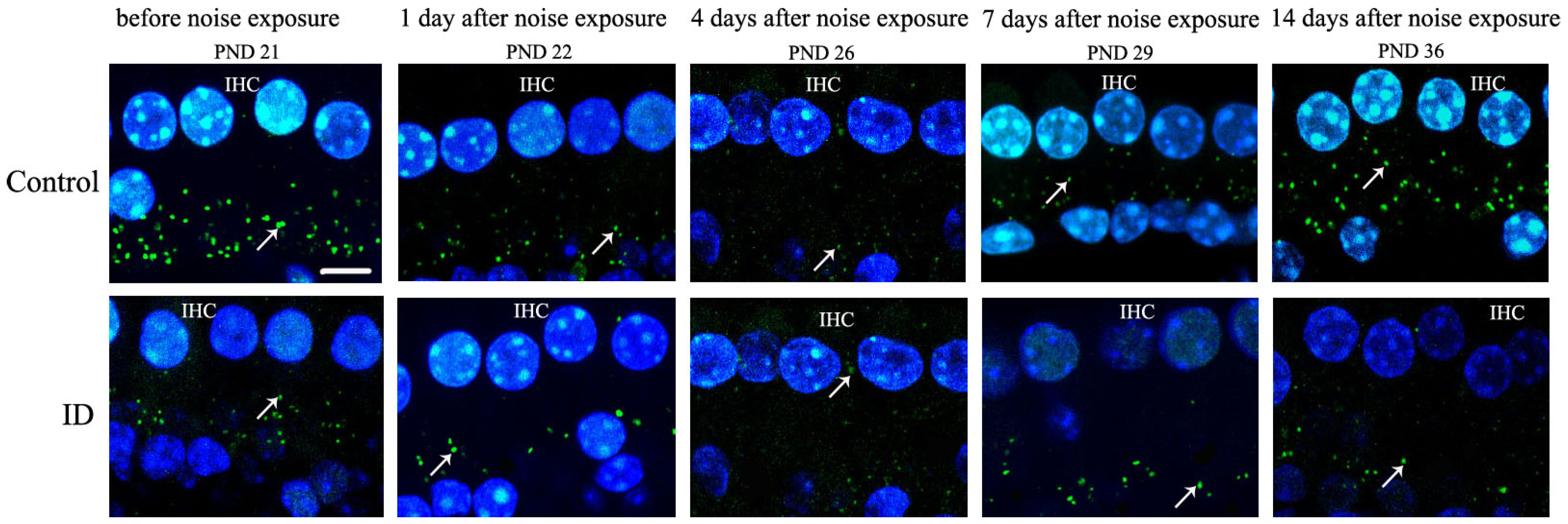
3.6. ID without Anemia Delays the Recovery from IHC Ribbon Synapses Damage in Young Rats after Noise Exposure
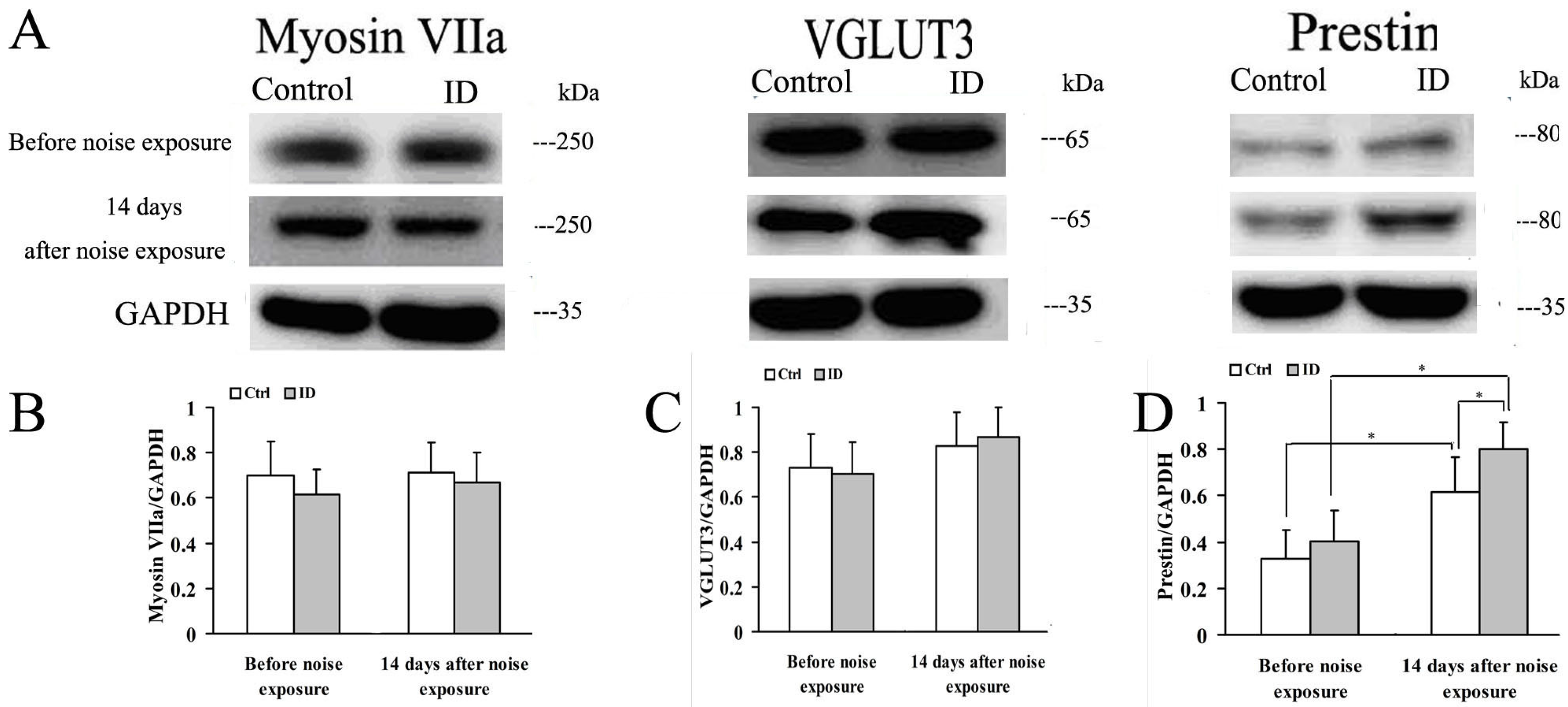
3.7. ID without Anemia Does Not Regulate Myosin VIIa, VGLUT3 and Prestin in Young Rat Cochleae of before and after Noise Exposure, but Upregulates Prestin in the Cochleae after Noise Exposure
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABR | auditory brainstem response |
| CtBP2 | carboxyterminal binding protein 2 |
| DPOAE | distortion product otoacoustic emission |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GD | gestational day |
| Hb | hemoglobin |
| Hct | hematocrit |
| HE staining | hematoxylin–eosin staining |
| ID | iron deficiency |
| IDA | iron deficiency anemia |
| IHC | inner hair cell |
| OHC | outer hair cell |
| PND | postnatal day |
| SEM | scanning electron microscopy |
| SI | serum level of iron |
| SF | serum ferritin |
| SGC | spiral ganglion cell |
| SPL | sound pressure level |
| VGLUT | vesicular glutamate transporter |
References
- Scholl, T.O. Iron status during pregnancy: Setting the stage for mother and infant. Am. J. Clin. Nutr. 2005, 81, 1218S–1222S. [Google Scholar] [PubMed]
- Zimmermann, M.B.; Hurrell, R.F. Nutritional iron deficiency. Lancet 2007, 370, 511–520. [Google Scholar] [CrossRef]
- McLean, E.; Cogswell, M.; Egli, I.; Wojdyla, D.; de Benoist, B. Worldwide prevalence of anaemia, who vitamin and mineral nutritionin formationsystem, 1993–2005. Public Health Nutr. 2009, 12, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Radlowski, E.C.; Johnson, R.W. Perinatal iron deficiency and neurocognitive development. Front. Hum. Neurosci. 2013, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Cameron, B.M.; Neufeld, L.M. Estimating the prevalence of iron deficiency in the first two years of life: Technical and measurement issues. Nutr. Rev. 2011, 69, S49–S56. [Google Scholar] [CrossRef] [PubMed]
- Kürekçi, A.E.; Sarici, S.U.; Karaoglu, A.; Ulaş, U.H.; Atay, A.A.; Serdar, M.A.; Akin, R.; Ozcan, O. Effects of iron deficiency versus iron deficiency anemia on brainstem auditory evoked potentials in infancy. Turk. J. Pediatr. 2006, 48, 334–339. [Google Scholar] [PubMed]
- Amin, S.B.; Orlando, M.; Eddins, A.; MacDonald, M.; Monczynski, C.; Wang, H. In utero iron status and auditory neural maturation in premature infants as evaluated by auditory brainstem response. J. Pediatr. 2010, 156, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.B.; Orlando, M.; Wang, H. Latent iron deficiency in utero is associated with abnormal auditory neural myelination in ≥35 weeks gestational age infants. J. Pediatr. 2013, 163, 1267–1271. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, V.; Amin, S.B.; Agarwal, A.; Srivastava, L.M.; Soni, A.; Saluja, S. Latent iron deficiency at birth influences auditory neural maturation in late preterm and term infants. Am. J. Clin. Nutr. 2015, 102, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Yamashita, D.; Katsunuma, S.; Hasegawa, S.; Tanimoto, H.; Nibu, K. Increased inner ear susceptibility to noise injury in mice with streptozotocin-induced diabetes. Diabetes 2012, 61, 2980–2986. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Liu, L.; He, T.; Guo, X.; Yu, Z.; Yin, S.; Wang, J. Ribbon synapse plasticity in the cochleae of guinea pigs after noise-induced silent damage. PLoS ONE 2013, 8, e81566. [Google Scholar] [CrossRef] [PubMed]
- Selander, J.; Albin, M.; Rosenhall, U.; Rylander, L.; Lewné, M.; Gustavsson, P. Maternal occupational exposure to noise during pregnancy and hearing dysfunction in children: A nationwide prospective cohort study in Sweden. Environ. Health Perspect. 2016, 124, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Moser, T.; Brandt, A.; Lysakowski, A. Hair cell ribbon synapses. Cell Tissue Res. 2006, 326, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Glowatzki, E.; Fuchs, P.A. Transmitter release at the hair cell ribbon synapse. Nat. Neurosci. 2002, 5, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.L.; Forge, A.; Knipper, M.; Münkner, S.; Marcotti, W. Tonotopic variation in the calcium dependence of neurotransmitter release and vesicle pool replenishment at mammalian auditory ribbon synapses. J. Neurosci. 2008, 28, 7670–7678. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.C.; Frank, T.; Khimich, D.; Hoch, G.; Riedel, D.; Chapochnikov, N.M.; Yarin, Y.M.; Harke, B.; Hell, S.W.; Egner, A.; et al. Tuning of synapse number, structure and function in the cochlea. Nat. Neurosci. 2009, 12, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, F.; Königstorfer, A.; Südhof, T.C. RIBEYE, a component of synaptic ribbons: A protein’s journey through evolution provides insight into synaptic ribbon function. Neuron 2000, 28, 857–872. [Google Scholar] [CrossRef]
- Tom Dieck, S.; Altrock, W.D.; Kessels, M.M.; Qualmann, B.; Regus, H.; Brauner, D.; Fejtová, A.; Bracko, O.; Gundelfinger, E.D.; Brandstätter, J.H. Molecular dissection of the photoreceptor ribbon synapse: Physical interaction of Bassoon and RIBEYE is essential for the assembly of the ribbon complex. J. Cell Biol. 2005, 168, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, G.P.; Zeng, R.; Guo, J.Y.; Chen, C.F.; Gong, S.S. Temporospatial expression and cellular localization of VGLUT3 in the rat cochlea. Brain Res. 2013, 1537, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Ji, F.; Xu, Y.; Wang, X.; Hou, Z.; Yang, S. Myosin VIIa and otoferlin in cochlear inner hair cells have distinct response to ototoxic exposure. Acta Otolaryngol. 2014, 134, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Song, Y.; Wang, R.; Gao, S.S.; Clifton, W.; Raphael, P.; Chao, S.I.; Pereira, F.A.; Groves, A.K.; Oghalai, J.S. Prestin regulation and function in residual outer hair cells after noise-induced hearing loss. PLoS ONE 2013, 8, e82602. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Hao, S.; Yang, B.; Zhao, Y.; Zhang, W.; Yang, J. Mild maternal iron deficiency anemia induces hearing impairment associated with reduction of ribbon synapse density and dysregulation of VGLUT3, myosin VIIa, and prestin expression in young guinea pigs. Neurotox. Res. 2016, 29, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [PubMed]
- Hu, X.; Teng, X.; Zheng, H.; Shan, Z.; Li, J.; Jin, T.; Xiong, C.; Zhang, H.; Fan, C.; Teng, W. Iron deficiency without anemia causes maternal hypothyroxinemia in pregnant rats. Nutr. Res. 2014, 34, 604–612. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, C.P.; Fiset, S.; Surette, M.E.; Turgeon O’Brien, H.; Rioux, F.M. Maternal iron deficiency alters essential fatty acid and eicosanoid metabolism and increases locomotion in adult guinea pig offspring. J. Nutr. 2009, 139, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Hao, S.; Yang, B.; Zhao, Y.; Zhang, R.; Zhang, W.; Yang, J.; Chen, J. Insulin resistance due to dietary iron overload disrupts inner hair cell ribbon synapse plasticity in male mice. Neurosci. Lett. 2015, 597, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, N.; Yan, K.; Liu, X.; Zhang, Y.; Hong, Z.; Wang, M.; Yin, Q.; Wu, F.; Lei, Y.; et al. Inner hair cell ribbon synapse plasticity might be molecular basis of temporary hearing threshold shifts in mice. Int. J. Clin. Exp. Pathol. 2015, 8, 8680–8691. [Google Scholar] [PubMed]
- Yu, F.; Hao, S.; Zhao, Y.; Ren, Y.; Yang, J.; Sun, X.; Chen, J. Mild maternal iron deficiency anemia induces DPOAE suppression and cochlear hair cell apoptosis by caspase activation in young guinea pigs. Environ. Toxicol. Pharmacol. 2014, 37, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Jiang, X.; Shi, C.; Shi, L.; Yang, B.; Shi, L.; Xu, Y.; Yang, W.; Yang, S. Cochlear inner hair cell ribbon synapse is the primary target of ototoxic aminoglycoside stimuli. Mol. Neurobiol. 2013, 48, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Werner, M.; van de Water, T.R.; Andersson, T.; Arnoldsson, G.; Berggren, D. Morphological and morphometric characteristics of vestibular hair cells and support cells in long term cultures of rat utricle explants. Hear. Res. 2012, 283, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.L.; Strathmann, F.G.; Gelein, R.; Walton, J.; Mayer-Pröschel, M. Iron deficiency disrupts axon maturation of the developing auditory nerve. J. Neurosci. 2012, 32, 5010–5015. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zheng, G.; Wu, Y.; Shen, X.; Jing, J.; Yu, T.; Song, H.; Chen, J.; Luo, W. Lead exposure results in hearing loss and disruption of the cochlear blood-labyrinth barrier and the protective role of iron supplement. Neurotoxicology 2013, 39, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, A.; Ciorba, A.; Aimoni, C.; Orioli, E.; Zeri, G.; Vigliano, M.; Gemmati, D. Sudden sensorineural hearing loss and polymorphisms in iron homeostasis genes: New insights from a case-control study. BioMed Res. Int. 2015, 2015, 834736. [Google Scholar] [CrossRef] [PubMed]
- Jewett, D.L.; Romano, M.N.; Williston, J.S. Human auditory evoked potentials: Possible brain stem components detected on the scalp. Science 1970, 167, 1517–1518. [Google Scholar] [CrossRef] [PubMed]
- Khimich, D.; Nouvian, R.; Pujol, R.; Tom Dieck, S.; Egner, A.; Gundelfinger, E.D.; Moser, T. Hair cell synaptic ribbons are essential for synchronous auditory signaling. Nature 2005, 434, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Saluja, S.; Modi, M. Fetal-neonatal iron deficiency is associated with poorer auditory recognition memory at 2 months of age. Evid. Based Med. 2016, 21, 112. [Google Scholar] [CrossRef] [PubMed]
- Brumwell, C.L.; Hossain, W.A.; Morest, D.K.; Bernd, P. Role for basic fibroblast growth factor (FGF-2) in tyrosine kinase (TrkB) expression in the early development and innervation of the auditory receptor: In vitro and in situ studies. Exp. Neurol. 2000, 162, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, S.G.; Liberman, M.C. Synaptopathy in the noise-exposed and aging cochlea: Primary neural degeneration in acquired sensorineural hearing loss. Hear. Res. 2015, 330, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Ohlemiller, K.K. Age-related loss of spiral ganglion neurons. Hear. Res. 2010, 264, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Kalinec, G.M.; Fernandez-Zapico, M.E.; Urrutia, R.; Esteban-Cruciani, N.; Chen, S.; Kalinec, F. Pivotal role of Harakiri in the induction and prevention of gentamicin-induced hearing loss. Proc. Natl. Acad. Sci. USA 2005, 102, 16019–16024. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg Diet) | Control | ID |
|---|---|---|
| Cornstarch | 529.5 | 529.92 |
| Casein (>85% protein) | 200.0 | 200.0 |
| Sucrose | 100.0 | 100.0 |
| Soybean oil | 70.0 | 70.0 |
| Fiber | 50.0 | 50.0 |
| Mineral mix (Ferric citrate) | 35.0 (0.735) | 34.58 (0.315) |
| Vitamin mix | 10.0 | 10.0 |
| l-Cysteine | 3.0 | 3.0 |
| Choline bitartrate | 2.5 | 2.5 |
| Group | Control | ID |
|---|---|---|
| Number of dams | 10 | 10 |
| Number of successful parturition | 10 | 10 |
| Duration of gestation (day) a | 21.0 ± 0.0 | 21.2 ± 0.5 |
| Maternal weight gain (g) a GD 0–21 | 98.3 ± 18.3 | 96.9 ± 22.5 |
| PND 4 sex ratio (male:female) | 50:45 | 56:44 |
| PND 4 body weights (g) a | 10.11 ± 2.22 | 8.61 ± 1.49 |
| PND 7 body weights (g) a | 13.62 ± 2.12 | 12.21 ± 3.29 |
| PND 14 body weights (g) a | 25.93 ± 3.34 | 23.36 ± 5.31 |
| PND 21 body weights (g) a | 45.79 ± 1.43 | 43.11 ± 5.03 |
| PND 28 body weights (g) a | 82.23 ± 10.16 | 79.77 ± 10.09 |
| PND 35 body weights (g) a | 122.92 ± 12.79 | 112.85 ± 19.56 |
| GD 13 | ||
| SI (µmol/L) a | 95.07 ± 18.81 | 90.50 ± 13.22 |
| Hct | 0.45 ± 0.13 | 0.44 ± 0.11 |
| SF (mg/L) a | 5.19 ± 1.53 | 3.51 ± 1.07 * |
| GD 21 | ||
| SI (µmol/L) a | 97.38 ± 23.05 | 90.92 ± 20.34 |
| Hct | 0.46 ± 0.16 | 0.44 ± 0.19 |
| SF (mg/L) a | 6.04 ± 1.61 | 3.98 ± 1.36 * |
| PND 21 | ||
| Dam | ||
| Hb (g/L) a | 126.40 ± 17.62 | 115.08 ± 20.06 |
| SI (µmol/L) a | 110.19 ± 29.03 | 96.90 ± 26.11 |
| Hct | 0.40 ± 0.17 | 0.39 ± 0.15 |
| SF (mg/L) a | 6.29 ± 1.43 | 4.16 ± 1.19 * |
| PND 21 | ||
| Pups | ||
| Hb (g/L) a | 117.46 ± 13.51 | 108.72 ± 13.79 |
| SF (mg/L) a | 5.28 ± 1.33 | 3.61 ± 1.02 * |
| PND 36 | ||
| Pups | ||
| Hb (g/L) a | 127.23 ± 20.06 | 114.07 ± 16.45 |
| SF (mg/L) a | 5.53 ± 1.06 | 3.07 ± 1.10 * |
| Group | Control | ID |
|---|---|---|
| GD 3 | 21.49 ± 3.96 | 22.12 ± 3.88 |
| GD 6 | 26.74 ± 5.98 | 25.90 ± 4.45 |
| GD 7 | 27.85 ± 7.04 | 25.52 ± 3.80 |
| GD 9 | 30.74 ± 7.45 | 25.06 ± 4.11 |
| GD 11 | 27.41 ± 5.65 | 25.42 ± 5.54 |
| GD 14 | 29.30 ± 2.78 | 26.75 ± 3.91 |
| GD 17 | 29.02 ± 3.97 | 29.76 ± 4.90 |
| GD 20 | 22.59 ± 7.68 | 25.94 ± 6.95 |
| PND 1 | 24.57 ± 7.29 | 23.48 ± 5.56 |
| PND 4 | 34.95 ± 4.05 | 37.89 ± 7.89 |
| PND 7 | 44.96 ± 6.00 | 46.21 ± 6.38 |
| PND 14 | 63.75 ± 6.48 | 61.09 ± 7.68 |
| Analysis Method | Control (32 Ears) | ID (32 Ears) |
|---|---|---|
| Hearing test ABR recording | ||
| PND 21 | 8 (right ear) | 8 (right ear) |
| PND 22 | 8 (right ear) | 8 (right ear) |
| PND 26 | 8 (right ear) | 8 (right ear) |
| PND 29 | 8 (right ear) | 8 (right ear) |
| PND 36 | 8 (right ear) | 8 (right ear) |
| Morphological test | ||
| SEM (PND 21) | 4 (left ear) | 4 (left ear) |
| Immunofluorescence | ||
| DAPI-positive nuclei (PND 21) | 4 (left ear) | 4 (left ear) |
| RIBEYE/CtBP2 (PND 21) | 4 (left ear) | 4 (left ear) |
| (PND 36) | 4 (left ear) | 4 (left ear) |
| HE staining (PND 21) | 4 (right ear) | 4 (right ear) |
| (PND 36) | 4 (right ear) | 4 (right ear) |
| Western blotting | 3 (left ear) | 3 (left ear) |
| SPL (dB) | Control | ID | ||
|---|---|---|---|---|
| A of Wave Ip a | Latency of Ip a | A of Wave Ip a | Latency of Ip a | |
| 70 | 5.92 ± 1.67 | 1.49 ± 0.13 | 3.11 ± 0.96 * | 1.56 ± 0.17 |
| 80 | 6.53 ± 1.70 | 1.35 ± 0.15 | 3.52 ± 1.31 * | 1.44 ± 0.16 |
| 90 | 6.90 ± 1.76 | 1.29 ± 0.17 | 4.37 ± 1.08 * | 1.35 ± 0.10 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, F.; Hao, S.; Yang, B.; Zhao, Y.; Yang, J. RETRACTED: Low Iron Diet Increases Susceptibility to Noise-Induced Hearing Loss in Young Rats. Nutrients 2016, 8, 456. https://doi.org/10.3390/nu8080456
Yu F, Hao S, Yang B, Zhao Y, Yang J. RETRACTED: Low Iron Diet Increases Susceptibility to Noise-Induced Hearing Loss in Young Rats. Nutrients. 2016; 8(8):456. https://doi.org/10.3390/nu8080456
Chicago/Turabian StyleYu, Fei, Shuai Hao, Bo Yang, Yue Zhao, and Jun Yang. 2016. "RETRACTED: Low Iron Diet Increases Susceptibility to Noise-Induced Hearing Loss in Young Rats" Nutrients 8, no. 8: 456. https://doi.org/10.3390/nu8080456
APA StyleYu, F., Hao, S., Yang, B., Zhao, Y., & Yang, J. (2016). RETRACTED: Low Iron Diet Increases Susceptibility to Noise-Induced Hearing Loss in Young Rats. Nutrients, 8(8), 456. https://doi.org/10.3390/nu8080456