Mangiferin and Cancer: Mechanisms of Action



Abstract
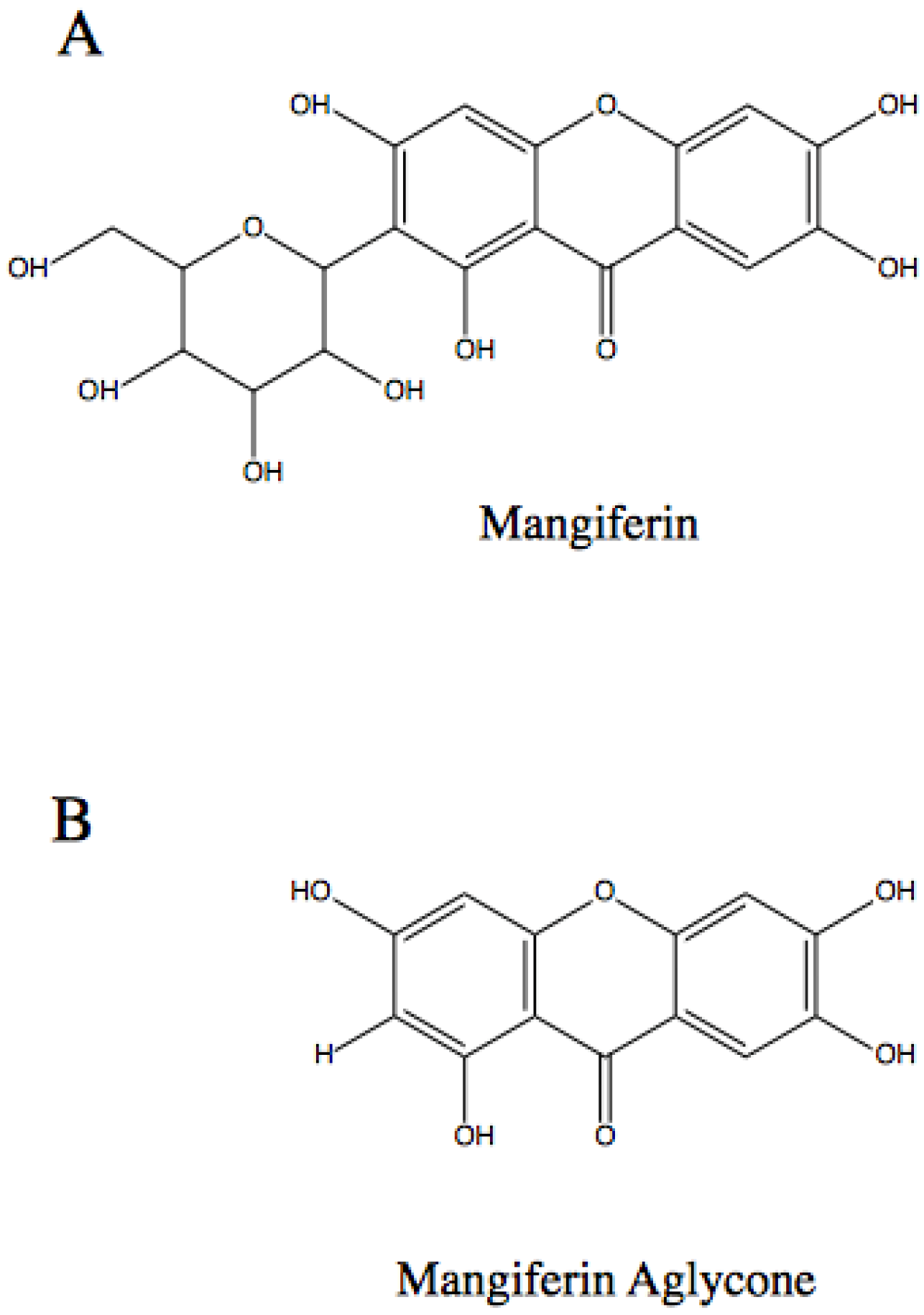
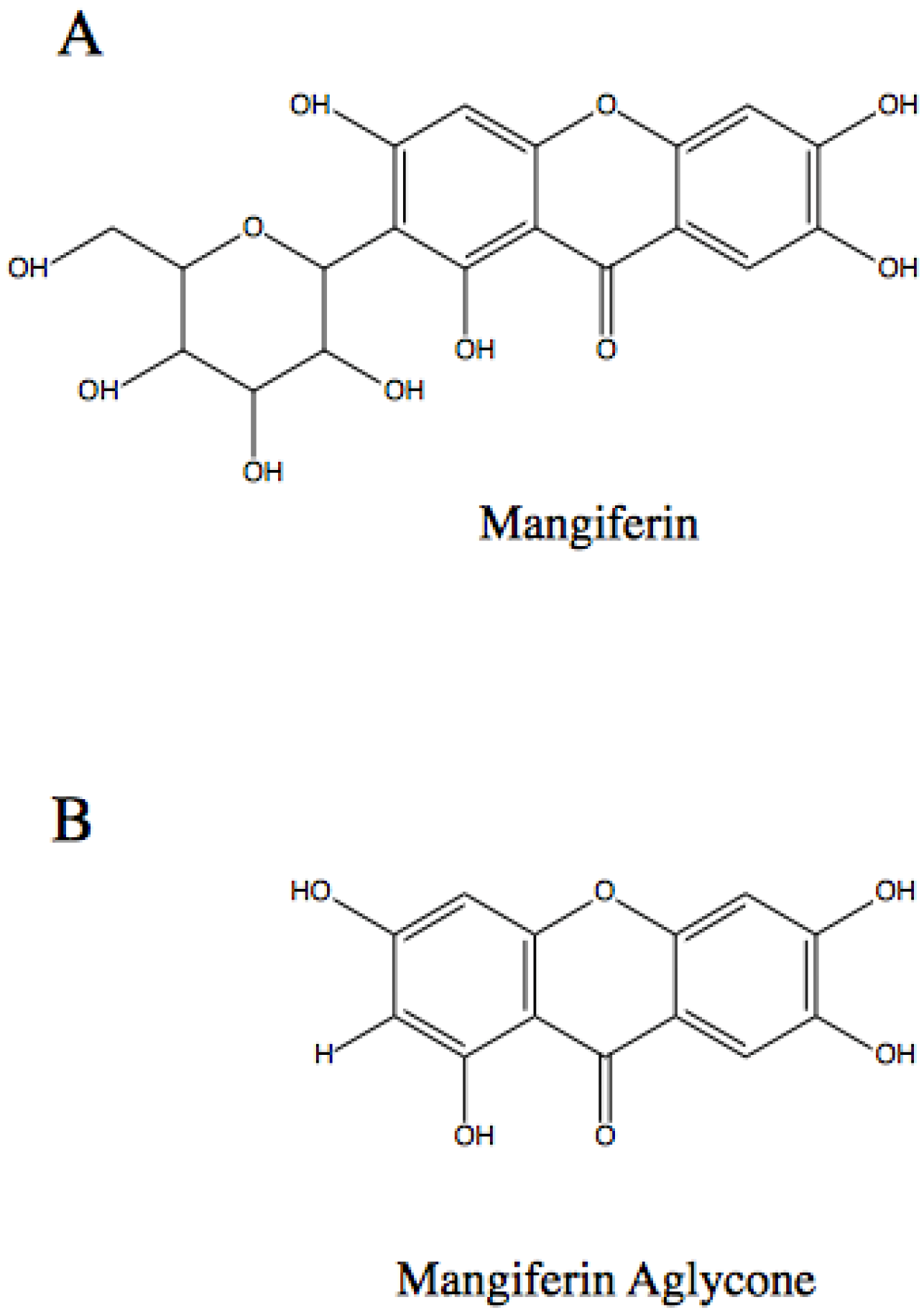
:1. Introduction
2. Molecular Mechanisms of the Anti-Cancer Action of Mangiferin
2.1. Inflammation
2.1.1. Nuclear Factor -Light-Chain-Enhancer of Activated B Cells Activity
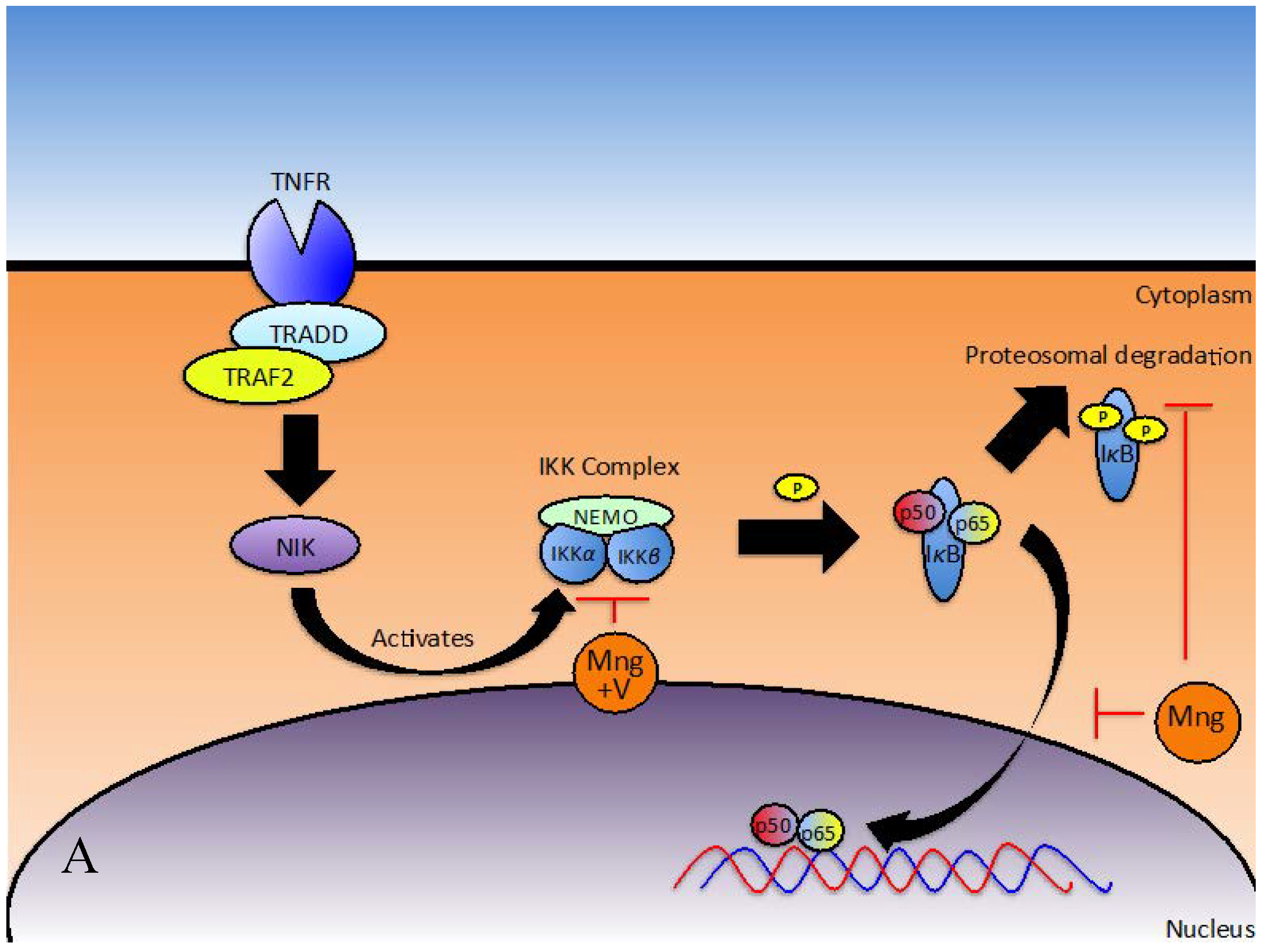
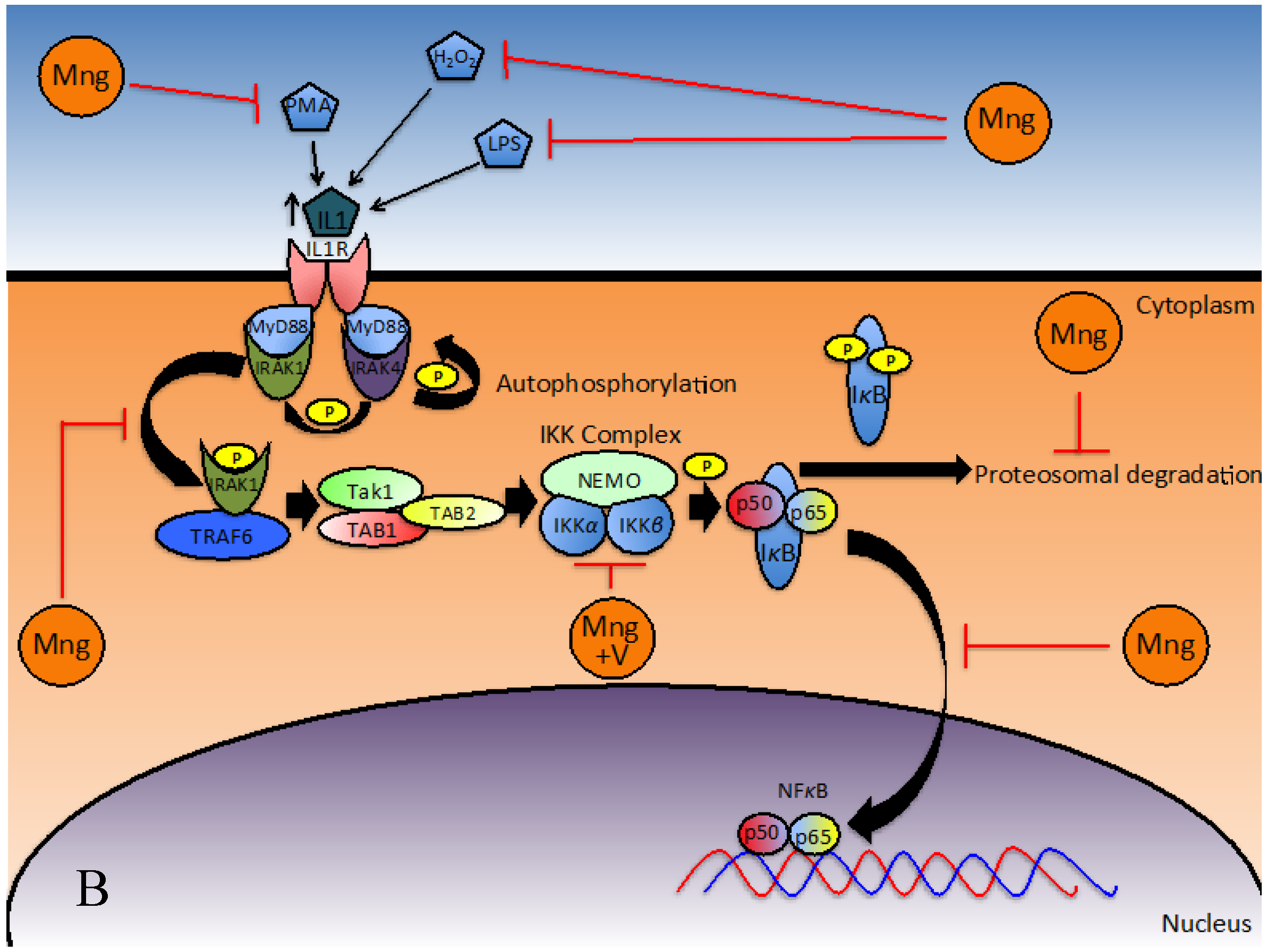
Initial Stimulus for NFκB Activation
Signal Transduction to Activate NFκB
Consequential Effects of NFκB Downregulation
2.1.2. Peroxisome Proliferator-Activated Receptor ү (PPARү)
2.1.3. Immune Response
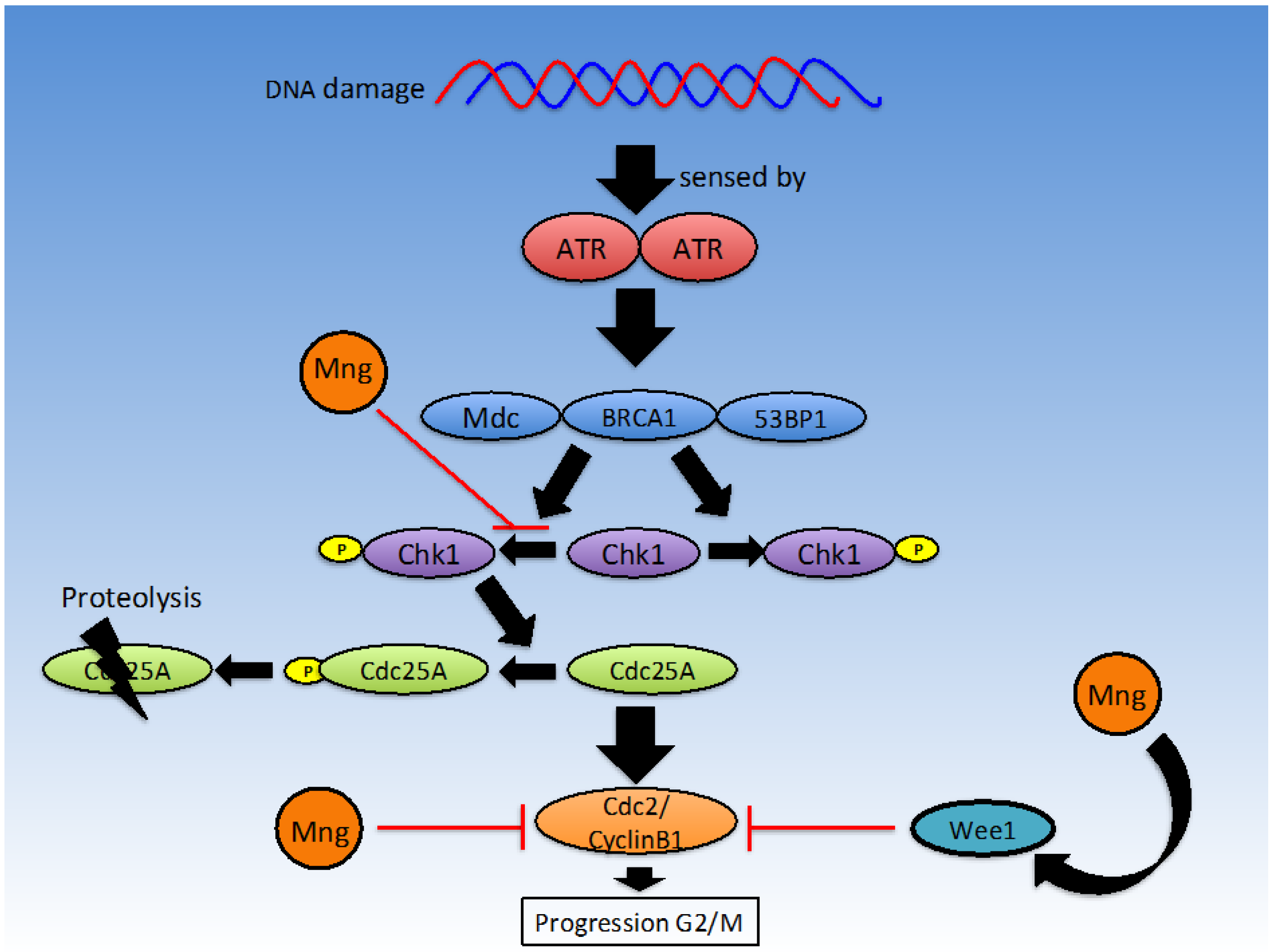
2.2. Cell Cycle
2.2.1. Mitogen Activated Protein Kinase Pathway
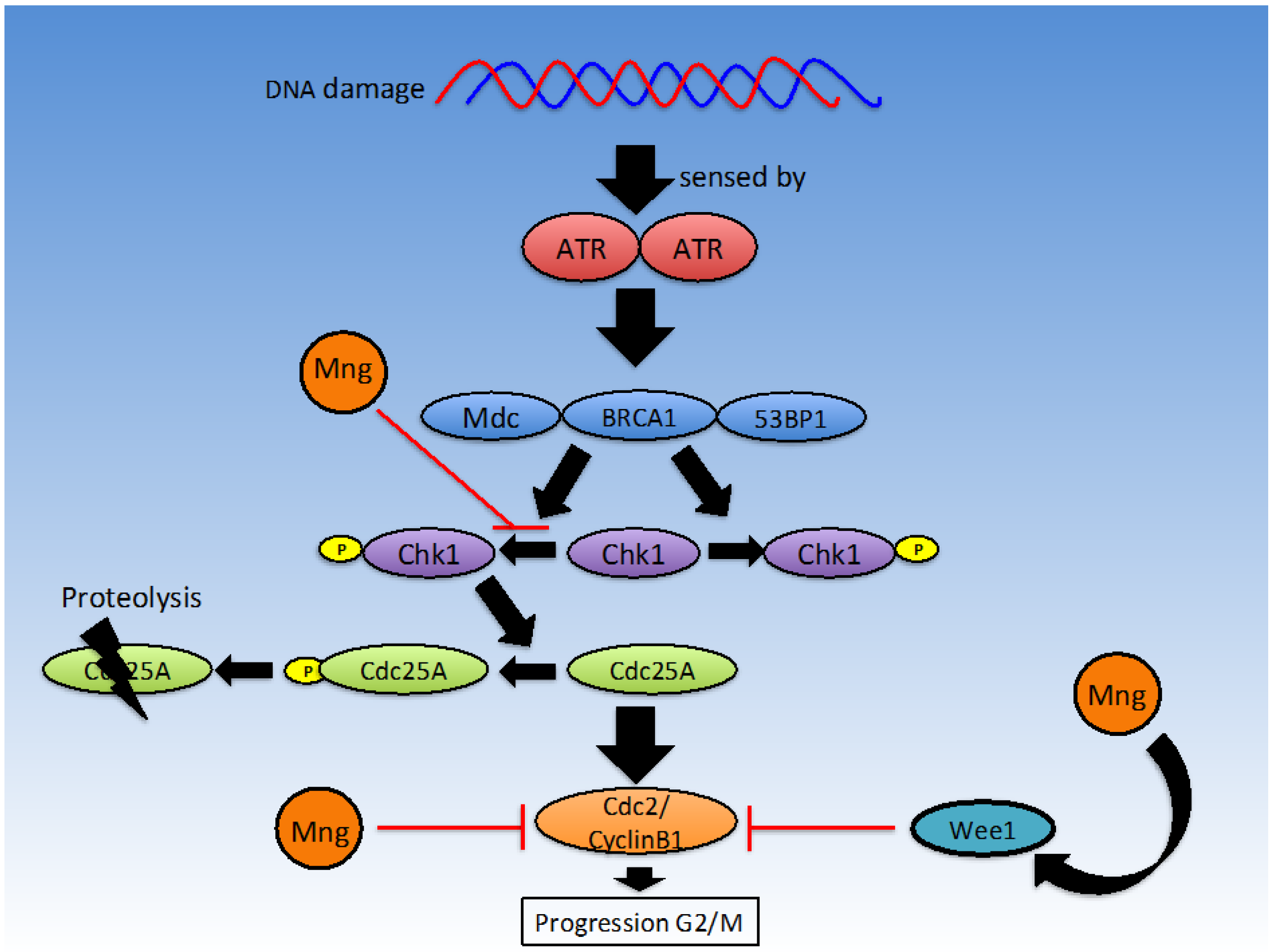
2.2.2. G2/M Checkpoint
2.3. Proliferation/Metastasis
2.3.1. Glycogen Synthase Kinase-3β/β-Catenin
2.3.2. Matrix Metalloproteinases
2.3.3. Epithelial to Mesenchymal Transition
2.3.4. Angiogenesis
2.3.5. Tumour Volume
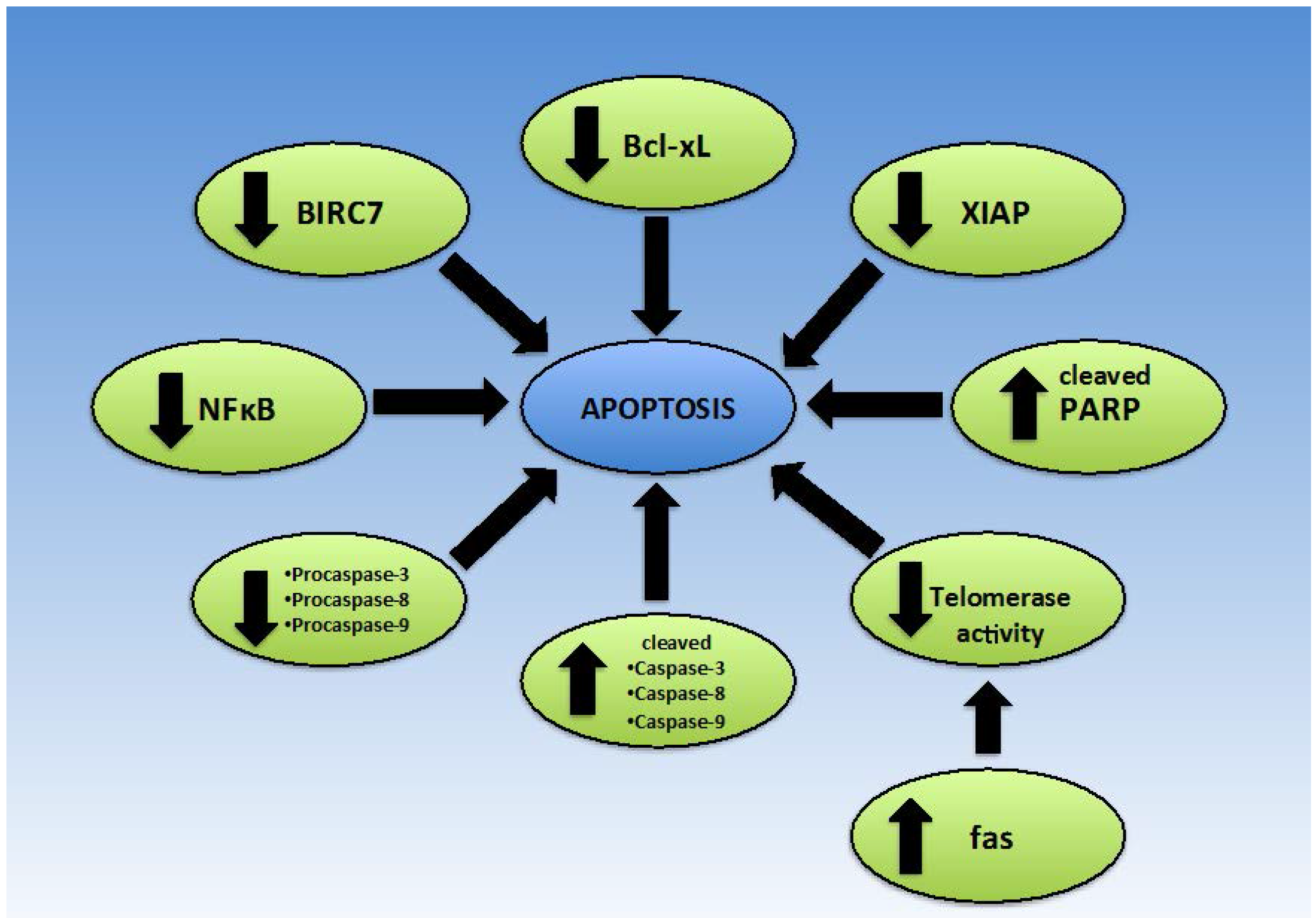
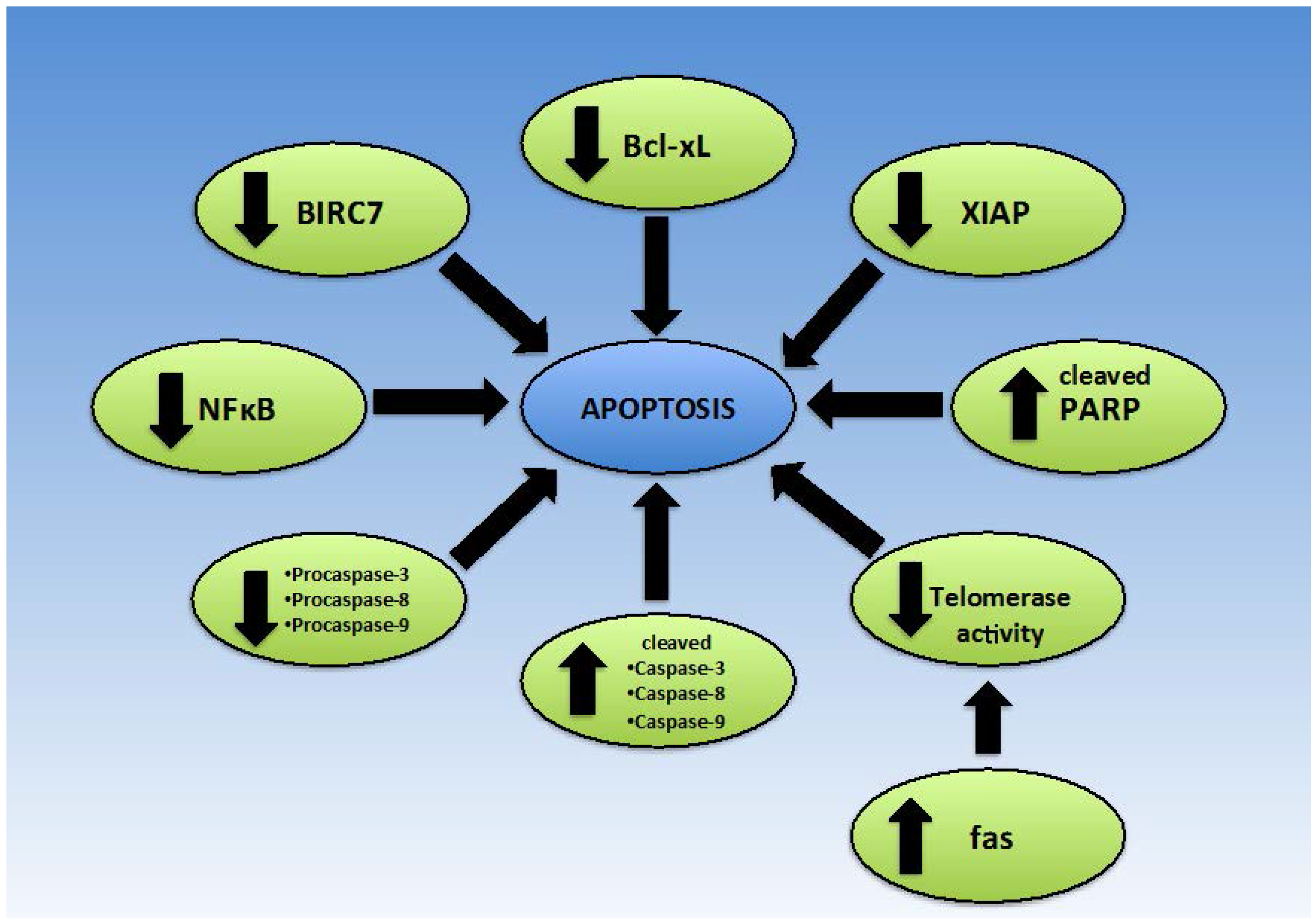
2.4. Apoptosis
2.4.1. Mangiferin and Hesperidin in Cyclopia Sp. Extracts
2.4.2. Bax/Bcl-2
2.4.3. Intrinsic/Extrinsic Apoptotic Pathway
2.4.4. Telomerase
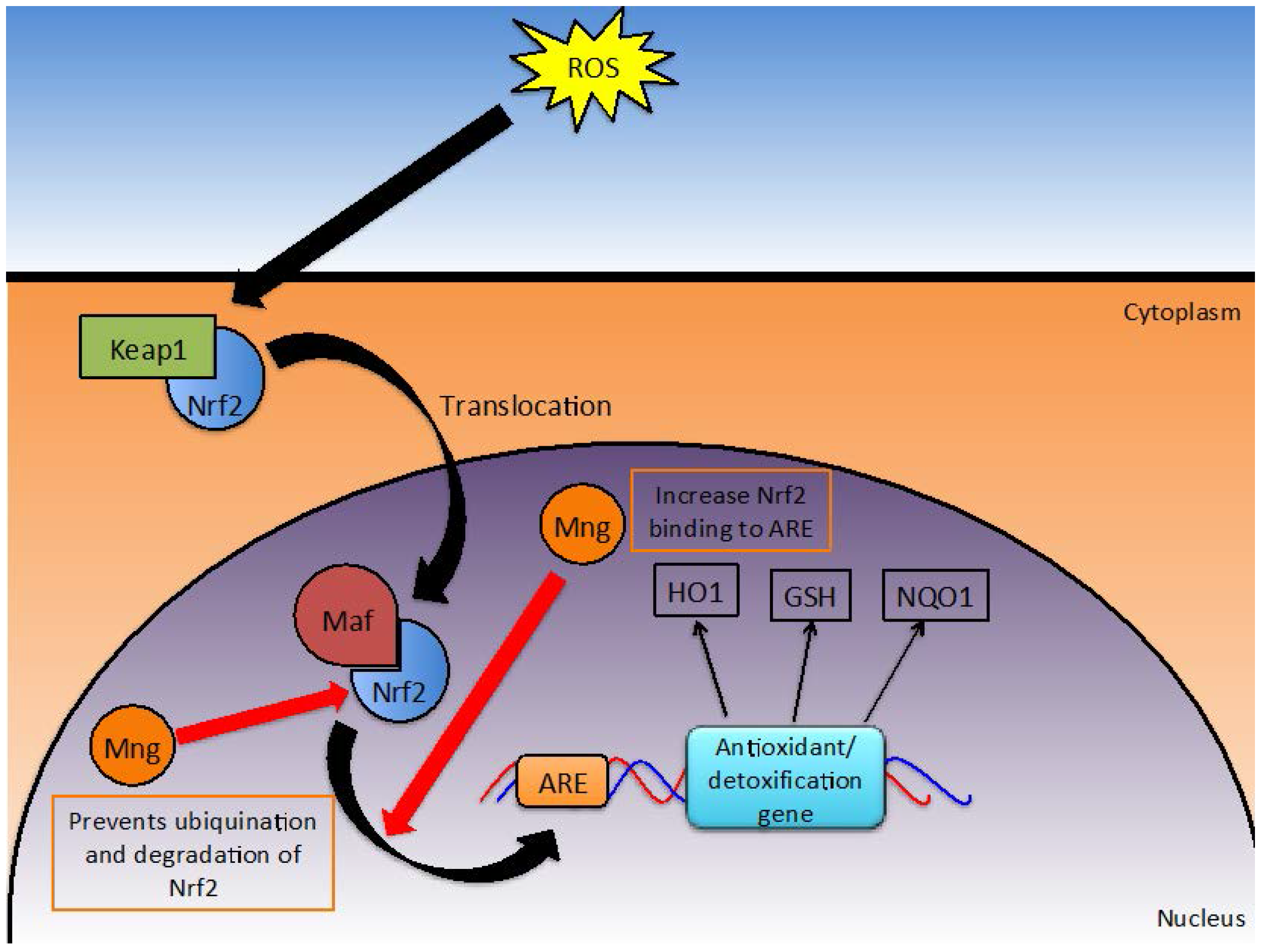
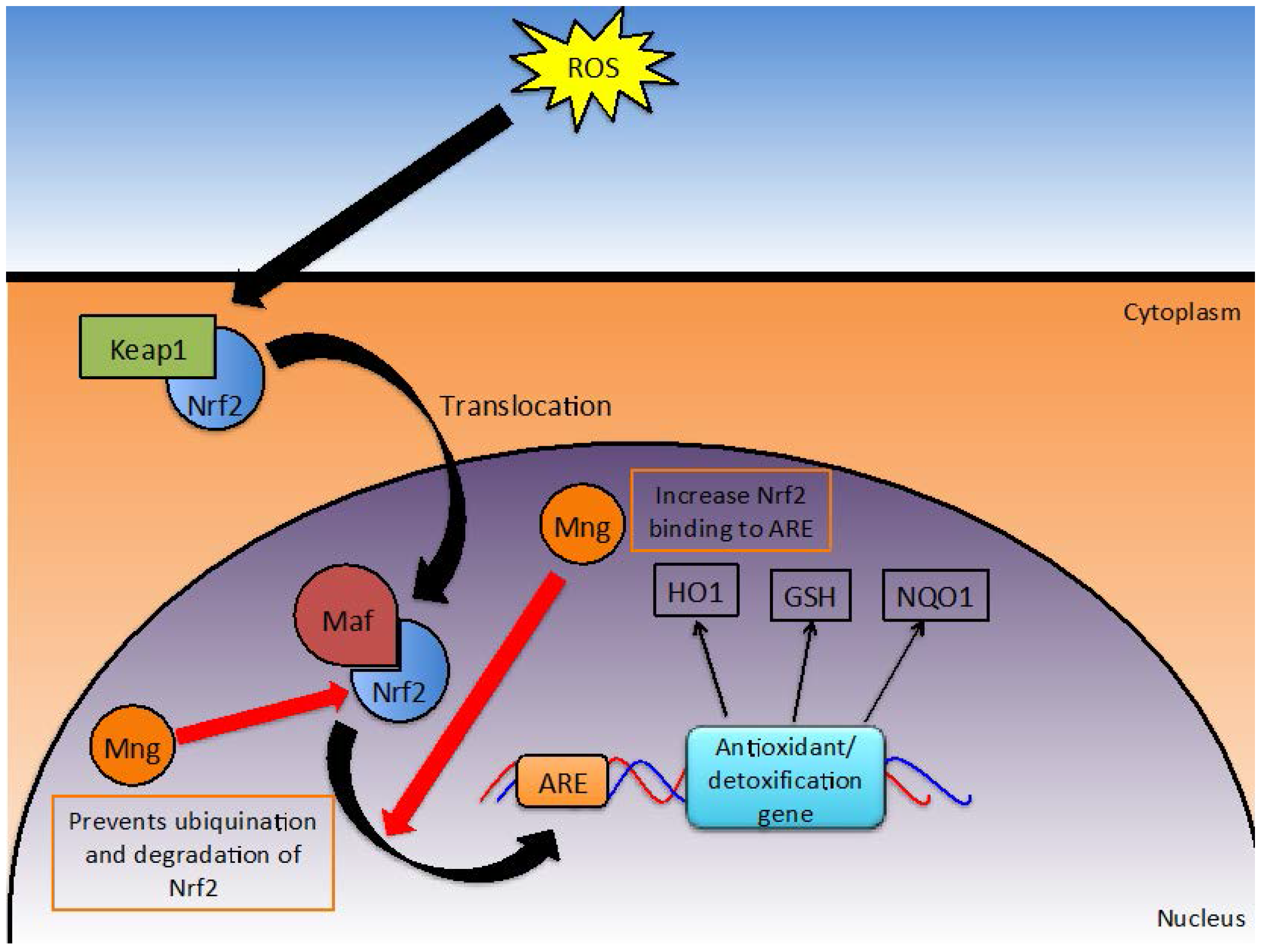
2.5. Oxidative Stress
2.5.1. Nrf2/ARE Detoxification Pathway
2.5.2. Elimination of Reactive Species
2.5.3. Catalase
2.6. DNA Damage
Radiation Damage
3. Synergistic Effects
3.1. Pro-Apoptotic Agents
3.1.1. Oxaliplatin
3.1.2. Etoposide
3.1.3. Doxorubicin
4. Bioavailability and Delivery of Mangiferin
5. Toxicity
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AML | acute myeloid leukaemia |
| ARE | antioxidant response element |
| ATM | Ataxia telangiectasia mutated protein |
| ATR | Ataxia Telangiectasia and Rad3-related protein |
| ATZ | aminotriazole |
| Bax | Bcl-2 associated X protein |
| bcl-2 | B Cell Lymphoma-2 |
| bcl-xL | B Cell Lymphoma-extra large |
| B(a)P | benzo(a)pyrene |
| Bid | BH3 interacting domain |
| BIRC7 | Baculoviral IAP Repeat Containing 7 |
| Chk1 | Checkpoint kinase 1 |
| CHk2 | Checkpoint Kinase 2 |
| CDK1 | Cyclin-Dependent Kinase 1 |
| COX | Cyclooxygenase-2 |
| CXCR4 | C-X-C Chemokine Receptor type-4 |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| EMT | Epithelial to Mesenchymal Transition |
| ERK | Extracellular signal-Regulated Kinase |
| FADD | Fas Associated Death Domain |
| GSH | glutathione S-transferase |
| HO-1 | heme oxygenase |
| H2O2 | hydrogen peroxide |
| IARC | International Agency for Research on Cancer |
| ICAM-1 | Intercellular Adhesion Molecule-1 |
| IκB | Inhibitor of κB |
| IKK-α | Inhibitor of NFB Kinase subunit-α |
| IKK-β | Inhibitor of NFB Kinase subunit-β |
| IL-1R | Interleukin-1 Receptors |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| IRAK1 | Interleukin-1 Receptor Activated Kinase 1 |
| IRAK4 | Interleukin-1 Receptor Activated Kinase 4 |
| KEAP-1 | Kelch-like ECH-associated protein-1 |
| LDC | Less Developed Countries |
| LPS | lipopolysaccharide |
| maf | musculoaponeurotic fibrosarcoma |
| MAPK | Mitogen Activated Protein Kinase |
| MDCs | More developed countries |
| MMP | matrix metalloproteinase |
| MPP+ | 1-methyl-4-phenylpyridine |
| MTT | 3-(4,5-dimethyl-2-thiozolyl)-2,5-diphenyl-2H-tetrazolium bromide |
| Myd88 | Myeloid Differentiation Primary Response Gene 88 |
| NEMO | NFκB Essential Modulator |
| NFκB | Nuclear Factor -light-chain-enhancer of activated B cells |
| NIK | NCK Interacting Kinase |
| NQO1 | NAD(P)H: quinine reductases |
| Nrf2 | Nuclear factor erythroid 2-Related Factor 2 |
| PDG | peptidoglycan |
| PK | pharmacokinetics |
| PMA | phorbol-12-myristate-13-acetate |
| PPARү | Peroxisome Proliferator-Activated Receptor ү |
| ROS | Reactive oxygen species |
| SEAP | Secreted Embryonic Alkaline Phosphatase |
| SOD | superoxide |
| TAB1 | Transforming growth factor beta-activated kinase 1-binding protein 1 |
| TAB2 | Transforming growth factor beta-activated kinase 1-binding protein 2 |
| TAK1 | Transforming growth factor beta-activated kinase 1 |
| TLRs | Toll-like receptors |
| TNF | Tumour Necrosis Factor |
| TNFR | Tumour Necrosis Factor Receptor |
| TRADD | TNFR with Tumour Necrosis Factor Receptor type-1-Associated Death Domain protein |
| TRAF2 | Tumour Necrosis Factor Receptor-Associated Factor 2 |
| TRAF6 | Tumour necrosis factor Receptor-Associated Factor 6 |
| VEGF | Vascular Endothelial Growth Factor |
| XIAP | X linked Inhibitor of Apoptosis Protein |
References
- WHO. In Health in 2015 from Millenium Development Goals to Sustainable Development Goals 2015. Available online: http://www.who.int/gho/publications/mdgs-sdgs/MDGs-SDGs2015_chapter6.pdf (accessed on 20 April 2016).
- Torre, L.; Bray, F.; Siegel, R.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics 2012. Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- United Nations Department of Economic and Social Affairs. World Population Prospects: The 2012 Revision, Highlights and Advance Tables; United Nations: New York, NY, USA, 2013. [Google Scholar]
- World Cancer Research Fund International. Comparing More and Less Developed Countries. Available online: http://www.wcrf.org/int/cancer-facts-figures/comparing-more-less-developed-countries (accessed on 12 November 2015).
- Boon, V.; Carr, J.; Klebe, S. The role of viruses in carcinogenesis. AMSJ 2013, 4, 11–15. [Google Scholar]
- Parkin, D.M.; Boyd, L.; Walker, L.C. The fraction of cancer attributable to lifestyle and environmental factors in the UK in 2010. Br. J. Cancer. 2011, 105, S77–S81. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Rengarajan, T.; Nishigaki, I.; Ekambaram, G.; Sakthisekaran, D. Potent chemopreventive effect of mangiferin on lung carcinogenesis in experimental Swiss albino mice. J. Cancer Res. Ther. 2014, 10, 1033–1039. [Google Scholar] [PubMed]
- Lv, J.; Wang, Z.; Zhang, L.; Wang, H.L.; Liu, Y.; Li, C.; Deng, J.; Yi, W.; Bao, J.K. Mangiferin induces apoptosis and cell cycle arrest in MCF-7 cells both in vitro andin vivo. J. Anim. Vet. Adv. 2013, 12, 352–359. [Google Scholar]
- Rajendran, P.; Jayakumar, T.; Nishigaki, I.; Ekambaram, G.; Nishigaki, Y.; Vetriselvi, J.; Sakthisekaran, D. Immunomodulatory effect of mangiferin in experimental animals with Benzo(a)pyrene-induced lung carcinogenesis. Int. J. Biomed. Sci. 2013, 9, 68–74. [Google Scholar] [PubMed]
- Hu, X.Y.; Deng, J.G.; Wang, L.; Yuan, Y.F. Synthesis and anti-tumor activity evaluation of gallic acid-mangiferin hybrid molecule. Med. Chem. 2013, 9, 1058–1062. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Sreenivasan, Y.; Ramesh, G.T.; Manna, S.K. β-d-glucoside suppresses tumor necrosis factor-induced activation of nuclear transcription factor κB but potentiates apoptosis. J. Biol. Chem. 2004, 279, 33768–33781. [Google Scholar] [CrossRef] [PubMed]
- Rodeiro, I.; Delgado, R.; Garrido, G. Effects of a Mangifera indica L. stem bark extract and mangiferin on radiation-induced DNA damage in human lymphocytes and lymphoblastoid cells. Cell Prolif. 2014, 47, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.-P.; Zhao, J.; Li, S.-S.; Yang, L.-J.; Zeng, L.-L.; Chen, Y.; Fang, J. Mangiferin activates Nrf2-antioxidant response element signaling without reducing the sensitivity to etoposide of human myeloid leukemia cells in vitro. APS 2014, 35, 257–266. [Google Scholar] [PubMed]
- Li, H.; Huang, J.; Yang, B.; Xiang, T.; Yin, X.; Peng, W.; Cheng, W.; Wan, J.; Luo, F.; Li, H. Mangiferin exerts antitumor activity in breast cancer cells by regulating matrix metalloproteinases, epithelial to mesenchymal transition, and β-catenin signaling pathway. Toxicol. Appl. Pharmacol. 2013, 272, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Nageshwar Rao, B.; Satish Rao, B.S. Mangiferin attenuates methylmercury induced cytotoxicity against IMR-32, human neuroblastoma cells by the inhibition of oxidative stress and free radical scavenging potential. Chem. Biol. Interact. 2011, 193, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Liu, L.; Zhong, Z.; Xiao, C.; Zhang, J. Mangiferin regulates proliferation and apoptosis in glioma cells by induction of microRNA-15b and inhibition of MMP-9 expression. Oncol. Rep. 2015, 33, 2815–2820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhao, J.; Li, S.; Zeng, L.; Chen, Y.; Fang, J. Mangiferin activates the Nrf2-ARE pathway and reduces etoposide-induced DNA damage in human umbilical cord mononuclear blood cells. Pharm. Biol. 2015, 53, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, B.; Li, S.; Zeng, L.; Chen, Y.; Fang, J. Mangiferin increases Nrf2 protein stability by inhibiting its ubiquitination and degradation in human HL60 myeloid leukemia cells. Int. J. Mol. Med. 2014, 33, 1348–1354. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.S.; Jung, K.; Kim, D.H.; Kim, H.S. Selective inhibition of MMP-9 gene expression by mangiferin in PMA-stimulated human astroglioma cells: Involvement of PI3K/Akt and MAPK signaling pathways. Pharmacol. Res. 2012, 66, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Ghosh, J.; Roy, A.; Sil, P.C. Mangiferin exerts hepatoprotective activity against D-galactosamine induced acute toxicity and oxidative/nitrosative stress via Nrf2-NFκB pathways. Toxicol. Appl. Pharmacol. 2012, 260, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Matkowski, A.; Kuś, P.; Góralska, E.; Woźniak, D. Mangiferin—A bioactive xanthonoid, not only from mango and not just antioxidant. Mini Rev. Med. Chem. 2013, 13, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, M.; Nataraj, J.; Essa, M.M.; Memon, M.A.; Manivasagam, T. Mangiferin attenuates MPTP induced dopaminergic neurodegeneration and improves motor impairment, redox balance and Bcl-2/Bax expression in experimental Parkinson’s disease mice. Chem. Biol. Interact. 2013, 206, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, L.; Rodeiro, I.; Donato, M.T.; Herrera, J.A.; Delgado, R.; Castell, J.V.; Gómez-Lechón, M.J. Multiparametric evaluation of the cytoprotective effect of the Mangifera indica L. stem bark extract and mangiferin in HepG2 cells. J. Pharm. Pharmacol. 2013, 65, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.; Wu, H.; Li, H.; Jia, Q.; Song, G. Comparison of microwave-assisted and conventional extraction of mangiferin from mango (Mangifera indica L.) leaves. J. Sep. Sci. 2013, 36, 3457–3462. [Google Scholar] [PubMed]
- Chellan, N.; Joubert, E.; Strijdom, H.; Roux, C.; Louw, J.; Muller, C.J.F. Aqueous extract of unfermented honeybush (Cyclopia maculata) attenuates STZ-induced diabetes and β-cell cytotoxicity. Planta Med. 2014, 80, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Chavan, J.J.; Ghadage, D.M.; Kshirsagar, P.R.; Kudale, S.S. Optimization of extraction techniques and RP-HPLC analysis of antidiabetic and anticancer drug mangiferin from roots of saptarangi (Salacia chinensis L.). J. Liq. Chromatogr. Relat. Technol. 2015, 38, 963–969. [Google Scholar] [CrossRef]
- Chavan, J.J.; Ghadage, D.M.; Bhoite, A.S.; Umdale, S.D. Micropropagation, molecular profiling and RP-HPLC determination of mangiferin across various regeneration stages of Saptarangi (Salacia chinensis L.). Ind. Crops Prod. 2015, 76, 1123–1132. [Google Scholar] [CrossRef]
- Lei, J.; Zhou, C.; Hu, H.; Hu, L.; Zhao, M.; Yang, Y.; Chuai, Y.; Ni, J.; Cai, J. Mangiferin aglycone attenuates radiation-induced damage on human intestinal epithelial cells. J. Cell. Biochem. 2012, 113, 2633–2642. [Google Scholar] [CrossRef] [PubMed]
- Duang, X.Y.; Wang, Q.; Zhou, X.D.; Huang, D.M. Mangiferin: A possible strategy for periodontal disease to therapy. Med. Hypotheses 2011, 76, 486–488. [Google Scholar] [CrossRef] [PubMed]
- Kaivalya, M.; Rao, B.N.; Satish Rao, B.S. Mangiferin: A xanthone attenuates mercury chloride induced cytotoxicity and genotoxicity in HepG2 cells. J. Biochem. Mol. Toxicol. 2011, 25, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Barreto, J.C.; Trevisan, M.T.; Hull, W.E.; Erben, G.; de Brito, E.S.; Pfundstein, B.; Wurtele, G.; Spiegelhalder, B.; Owen, R.W. Characterization and quantitation of polyphenolic compounds in bark, kernel, leaves, and peel of mango (Mangifera indica L.). J. Agric. Food Chem. 2008, 56, 5599–5610. [Google Scholar] [CrossRef] [PubMed]
- Hewavitharana, A.K.; Tan, Z.W.; Shimada, R.; Shaw, P.N.; Flanagan, B.M. Between fruit variability of the bioactive compounds, β-carotene and mangiferin, in mango (Mangifera indica). Nutr. Diet. 2013, 70, 158–163. [Google Scholar] [CrossRef]
- Louisa, M.; Soediro, T.M.; Suyatna, F.D. In vitro modulation of P-glycoprotein, MRP-1 and BCRP expression by mangiferin in doxorubicin-treated MCF-7 cells. Asian Pac. J. Cancer Prev. 2014, 15, 1639–1642. [Google Scholar] [CrossRef] [PubMed]
- Dou, W.; Zhang, J.; Ren, G.; Ding, L.; Sun, A.; Deng, C.; Wu, X.; Wei, X.; Mani, S.; Wang, Z. Mangiferin attenuates the symptoms of dextran sulfate sodium-induced colitis in mice via NF-κB and MAPK signaling inactivation. Int. Immunopharmacol. 2014, 23, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.L.; Wang, A.Y.; Huang, Y.Q.; Luo, Y.; Ling, M. Mangiferin induces apoptosis by regulating Bcl-2 and bax expression in the CNE2 nasopharyngeal carcinoma cell line. Asian Pac. J. Cancer Prev. 2014, 15, 7065–7068. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.; Mosaddik, A.; Gyawali, R.; Ahn, K.S.; Cho, S.K. Induction of apoptosis by ethanolic extract of mango peel and comparative analysis of the chemical constitutes of mango peel and flesh. Food Chem. 2012, 133, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Maganda, J.; Blancas-Benítez, F.J.; Zamora-Gasga, V.M.; García-Magaña, M.d.L.; Bello-Pérez, L.A.; Tovar, J.; Sáyago-Ayerdi, S.G. Nutritional properties and phenolic content of a bakery product substituted with a mango (Mangifera indica)‘Ataulfo’processing by-product. Food Res. Int. 2015, 73, 117–123. [Google Scholar] [CrossRef]
- Yatnatti, S.; Vijayalakshmi, D.; Chandru, R. Processing and Nutritive Value of Mango Seed Kernel Flour. Curr. Res. Nutr. Food Sci. J. 2014, 2, 170–175. [Google Scholar] [CrossRef]
- Ajila, C.M.; Aalami, M.; Leelavathi, K.; Prasada Rao, U.J.S. Mango peel powder: A potential source of antioxidant and dietary fiber in macaroni preparations. Innov. Food Sci. Emerg. Technol. 2010, 11, 219–224. [Google Scholar] [CrossRef]
- Bandyopadhyay, K.; Chakraborty, C.; Bhattacharyya, S. Fortification of Mango Peel and Kernel Powder in Cookies Formulation. J. Acad. Ind. Res. 2014, 2, 661. [Google Scholar]
- García-Rivera, D.; Delgado, R.; Bougarne, N.; Haegeman, G.; Vanden Berghe, W. Gallic acid indanone and mangiferin xanthone are strong determinants of immunosuppressive anti-tumour effects of Mangifera indica L. bark in MDA-MB231 breast cancer cells. Cancer Lett. 2011, 305, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Fang, J.; Chen, Y. Antioxidant effect of mangiferin and its potential to be a cancer chemoprevention agent. Lett. Drug Des. Discov. 2013, 10, 239–244. [Google Scholar] [CrossRef]
- Telang, M.; Dhulap, S.; Mandhare, A.; Hirwani, R. Therapeutic and cosmetic applications of mangiferin: A patent review. Expert Opin. Ther. Pat. 2013, 23, 1561–1580. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.Q.; Yang, D.; Ma, Q.Y.; Yi, X.H.; Zhou, J.; Zhao, Y.X. A new benzophenone from Dobinea delavayi. Chem. Nat. Compd. 2013, 49, 46–48. [Google Scholar] [CrossRef]
- Xu, G.A. Drug Composition for Treating 2 Type Diabetes and its Chronicity Neopathy. Google Patents EP 2070540 A1, 17 June 2009. [Google Scholar]
- Zhang, W.; Li, P.; Gong, Y.; Gao, X. Mangiferin Aglycone Crystal form i and Preparation Method Thereof. Google Patents EP 2716637 A1, 9 April 2014. [Google Scholar]
- Ahmad, A.; Padhye, S.; Sarkar, F.H. Role of novel nutraceuticals garcinol, plumbagin and mangiferin in the prevention and therapy of human malignancies: Mechanisms of anticancer activity. In Nutraceuticals and Cancer; Springer: New York, NY, USA, 2012; Volume 1, pp. 179–199. [Google Scholar]
- Du Plessis-Stoman, D.; du Preez, J.G.H.; Van de Venter, M. Combination treatment with oxaliplatin and Mangiferin Causes Increased Apoptosis and downregulation of NFκB in cancer cell lines. AJOL 2011, 8, 177–184. [Google Scholar]
- Hudecová, A.; Kusznierewicz, B.; Rundén-Pran, E.; Magdolenová, Z.; Hašplová, K.; Rinna, A.; Fjellsbo, L.M.; Kruszewski, M.; Lankoff, A.; Sandberg, W.J.; et al. Silver nanoparticles induce premutagenic DNA oxidation that can be prevented by phytochemicals from Gentiana asclepiadea. Mutagenesis 2012, 27, 759–769. [Google Scholar] [CrossRef] [PubMed]
- De Souza, J.R.R.; Feitosa, J.P.A.; Ricardo, N.M.P.S.; Trevisan, M.T.S.; de Paula, H.C.B.; Ulrich, C.M.; Owen, R.W. Spray-drying encapsulation of mangiferin using natural polymers. Food Hydrocoll. 2013, 33, 10–18. [Google Scholar] [CrossRef]
- Kawpoomhae, K.; Sukma, M.; Ngawhirunpat, T.; Opanasopit, P.; Sripattanaporn, A. Antioxidant and neuroprotective effects of standardized extracts of Mangifera indica leaf. Thai J. Pharm. Sci. 2010, 34, 32–43. [Google Scholar]
- Peng, Z.G.; Yao, Y.B.; Yang, J.; Tang, Y.L.; Huang, X. Mangiferin induces cell cycle arrest at G2/M phase through ATR-Chk1 pathway in HL-60 leukemia cells. Genet. Mol. Res. 2015, 14, 4989–5002. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben-Neriah, Y. NF-κB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Vendramini-Costa, D.B.; Carvalho, J.E. Molecular link mechanisms between inflammation and cancer. Curr. Pharm. Des. 2012, 18, 3831–3852. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Hu, R.; Gou, Q.L. Inflammation and cancer. Pharm. Biotechnol. 2011, 18, 372–376. [Google Scholar]
- Bhoj, V.G.; Chen, Z.J. Ubiquitylation in innate and adaptive immunity. Nature 2009, 458, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Rhyasen, G.W.; Starczynowski, D.T. IRAK signalling in cancer. Br. J. Cancer 2015, 112, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Tully, J.E.; Nolin, J.D.; Guala, A.S.; Hoffman, S.M.; Roberson, E.C.; Lahue, K.G.; van der Velden, J.; Anathy, V.; Blackwell, T.S.; Janssen-Heininger, Y.M.W. Cooperation between classical and alternative NF-κB pathways regulates proinflammatory Responses in Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2012, 47, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.J.; Jang, S.E.; Hyam, S.R.; Han, M.J.; Kim, D.H. Mangiferin ameliorates colitis by inhibiting IRAK1 phosphorylation in NF-kappaB and MAPK pathways. Eur. J. Pharmacol. 2014, 740, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, B.K.; Zaidi, A.H.; Gupta, P.; Mokhamatam, R.B.; Raviprakash, N.; Mahali, S.K.; Manna, S.K. A natural xanthone increases catalase activity but decreases NF-kappa B and lipid peroxidation in U-937 and HepG2 cell lines. Eur. J. Pharmacol. 2015, 764, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Wee, Z.N.; Yatim, S.M.J.M.; Kohlbauer, V.K.; Feng, M.; Goh, J.Y.; Bao, Y.; Lee, P.L.; Zhang, S.; Wang, P.P.; Lim, E. IRAK1 is a therapeutic target that drives breast cancer metastasis and resistance to paclitaxel. Nat. Commun. 2015, 6, 8746. [Google Scholar] [CrossRef] [PubMed]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-kappaB and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The complexity of NF-kappaB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Belfiore, A.; Genua, M.; Malaguarnera, R. PPAR-agonists and their effects on IGF-I receptor signaling: Implications for cancer. PPAR Res. 2009, 2009, 18. [Google Scholar] [CrossRef] [PubMed]
- Vandoros, G.P.; Konstantinopoulos, P.A.; Sotiropoulou-Bonikou, G.; Kominea, A.; Papachristou, G.I.; Karamouzis, M.V.; Gkermpesi, M.; Varakis, I.; Papavassiliou, A.G. PPAR-gamma is expressed and NF-kB pathway is activated and correlates positively with COX-2 expression in stromal myofibroblasts surrounding colon adenocarcinomas. J. Cancer Res. Clin. Oncol. 2006, 132, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Hugo, H.J.; Saunders, C.; Ramsay, R.G.; Thompson, E.W. New Insights on COX-2 in Chronic Inflammation Driving Breast Cancer Growth and Metastasis. J. Mammary Gland Biol. 2015, 20, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud-Awny, M.; Attia, A.S.; Abd-Ellah, M.F.; El-Abhar, H.S. Mangiferin mitigates gastric ulcer in ischemia/reperfused rats: Involvement of PPAR-γ, NF-κB and Nrf2/HO-1 signaling pathways. PLoS ONE 2015, 10, e0132497. [Google Scholar] [CrossRef] [PubMed]
- Santarpia, L.; Lippman, S.M.; El-Naggar, A.K. Targeting the MAPK-RAS-RAF signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Malakhova, M.; Mottamal, M.; Reddy, K.; Kurinov, I.; Carper, A.; Langfald, A.; Oi, N.; Kim, M.O.; Zhu, F.; et al. Norathyriol suppresses skin cancers induced by solar ultraviolet radiation by targeting ERK kinases. Cancer Res. 2012, 72, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Shimada, M.; Nakanishi, M. DNA damage checkpoints and cancer. J. Mol. Histol. 2006, 37, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Mitra, J.; Enders, G.H. Cyclin A/Cdk2 complexes regulate activation of Cdk1 and Cdc25 phosphatases in human cells. Oncogene 2004, 23, 3361–3367. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Nong, C.; Guo, L.; Meng, G.; Zha, X.L. The proliferation inhibition effect and apoptosis induction of Mangiferin on BEL-7404 human hepatocellular carcinoma cell. Chin. J. Dig. 2002, 6, 341–343. [Google Scholar]
- Wilkinson, A.S.; Taing, M.W.; Pierson, J.T.; Lin, C.N.; Dietzgen, R.G.; Shaw, P.N.; Gidley, M.J.; Monteith, G.R.; Roberts-Thomson, S.J. Estrogen modulation properties of mangiferin and quercetin and the mangiferin metabolite norathyriol. Food Funct. 2015, 6, 1847–1854. [Google Scholar] [CrossRef] [PubMed]
- Dilshara, M.G.; Kang, C.H.; Choi, Y.H.; Kim, G.Y. Mangiferin inhibits tumor necrosis factor-α-induced matrix metalloproteinase-9 expression and cellular invasion by suppressing nuclear factor-κB activity. BMB Rep. 2015, 48, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Yang, J.; Moses, M.A. Matrix metalloproteinases as novel biomarker s and potential therapeutic targets in human cancer. J. Clin. Oncol. 2009, 27, 5287–5297. [Google Scholar] [CrossRef] [PubMed]
- Bartoszewski, R.; Hering, A.; Marsza, M.; Hajduk, J.S.; Bartoszewska, S.; Kapoor, N.; Kochan, K.; Ochocka, R. Mangiferin has an additive effect on the apoptotic properties of hesperidin in Cyclopia sp. tea extracts. PLoS ONE 2014, 9, e92128. [Google Scholar] [CrossRef] [PubMed]
- Salakou, S.; Kardamakis, D.; Tsamandas, A.C.; Zolota, V.; Apostolakis, E.; Tzelepi, V.; Papathanasopoulos, P.; Bonikos, D.S.; Papapetropoulos, T.; Petsas, T. Increased Bax/Bcl-2 ratio up-regulates caspase-3 and increases apoptosis in the thymus of patients with myasthenia gravis. In Vivo 2007, 21, 123–132. [Google Scholar] [PubMed]
- Kavitha, M.; Manivasagam, T.; Essa, M.M.; Tamilselvam, K.; Selvakumar, G.P.; Karthikeyan, S.; Thenmozhi, J.A.; Subash, S. Mangiferin antagonizes rotenone: Induced apoptosis through attenuating mitochondrial dysfunction and oxidative stress in SK-N-SH neuroblastoma cells. Neurochem. Res. 2014, 39, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Peng, Z.; Yang, J.; Song, S. The effect of mangiferin on telomerase activity and apoptosis in Leukemic K562 cells. Zhong Yao Cai 2007, 30, 306–309. [Google Scholar] [PubMed]
- Shay, J.W.; Wright, W.E. Role of telomeres and telomerase in cancer. Semin. Cancer Biol. 2012, 21, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Foresti, R. Anti-Inflammatory and Antioxidant Activities of Nrf2/HO-1 Activators: In Vitro Studies in Microglia and Retinal Cells; Univesity of Catania: Paris, France, 2014. [Google Scholar]
- Menkovic, N.; Juranic, Z.; Stanojkovic, T.; Raonic-Stevanovic, T.; Šavikin, K.; Zdunić, G.; Borojevic, N. Radioprotective activity of Gentiana lutea extract and mangiferin. Phytother. Res. 2010, 24, 1693–1696. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Chemocare.com. Oxaliplatin. Available online: http://www.chemocare.com/chemotherapy/drug-info/oxaliplatin.aspx (accessed on 2 September 2016).
- Schonn, I.; Hennesen, J.; Dartsch, D.C. Cellular responses to etoposide: Cell death despite cell cycle arrest and repair of DNA damage. Apoptosis 2010, 15, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frígola, A. Analytical methods for determining bioavailability and bioaccessibility of bioactive compounds from fruits and vegetables: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 155–171. [Google Scholar] [CrossRef]
- McClements, D.J.; Li, F.; Xiao, H. The Nutraceutical Bioavailability Classification Scheme: Classifying Nutraceuticals According to Factors Limiting their Oral Bioavailability. Ann. Rev. Food Sci. Technol. 2015, 6, 299–327. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Chen, C.; Zhang, C.; Zhang, Y.; Tang, X. Determination of mangiferin in rat plasma by liquid-liquid extraction with UPLC-MS/MS. J. Pharm. Biomed. Anal. 2010, 51, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Wang, F.; Li, Y.; Li, Y.; Wang, M.; Sun, D.; Sun, C. Pharmacokinetic study of mangiferin in human plasma after oral administration. Food Chem. 2012, 132, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, F.; Zeng, X.; Yang, L.; Deng, Y.; Wu, Z.; Feng, Y.; Li, X. Application of a liquid chromatography/tandem mass spectrometry method to pharmacokinetic study of mangiferin in rats. J. Chromatogr. B 2010, 878, 3345–3350. [Google Scholar] [CrossRef] [PubMed]
- Kammalla, A.K.; Ramasamy, M.K.; Inampudi, J.; Dubey, G.P.; Agrawal, A.; Kaliappan, I. Comparative Pharmacokinetic Study of Mangiferin After Oral Administration of Pure Mangiferin and US Patented Polyherbal Formulation to Rats. AAPS PharmSciTech 2015, 16, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Chen, H.; Sun, L.; Tong, L.; Zhang, T. Improving permeability and oral absorption of mangiferin by phospholipid complexation. Fitoterapia 2014, 93, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Al-Abd, A.M.; Aljehani, Z.K.; Gazzaz, R.W.; Fakhri, S.H.; Jabbad, A.H.; Alahdal, A.M.; Torchilin, V.P. Pharmacokinetic strategies to improve drug penetration and entrapment within solid tumors. J. Control Release 2015, 219, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Caro, C.; Pozo, D. Polysaccharide Colloids as Smart Vehicles in Cancer Therapy. Curr. Pharm Des. 2015, 21, 4822–4836. [Google Scholar] [CrossRef] [PubMed]
- Prado, Y.; Merino, N.; Acosta, J.; Herrera, J.A.; Luque, Y.; Hernández, I.; Prado, E.; Garrido, G.; Delgado, R.; Rodeiro, I. Acute and 28-day subchronic toxicity studies of mangiferin, a glucosyl xanthone isolated from Mangifera indica L. stem bark. J. Pharm. Pharm. Res. 2015, 3, 13–23. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemotherapeutic Agent | Cell Line | Reference | Evidence |
|---|---|---|---|
| Oxaliplatin | HeLa, HT29, HL60 | [48] | Reduction in oxaliplatin IC50 values; counteracts resistance to oxaliplatin. |
| Etoposide | HL60, U937 | [11,13] | Reduces oxidative stress. Protects normal cells without reducing sensitivity of HL60 to etoposide [13]. Activity of the drug is enhanced by mangiferin [11]. |
| Doxorubicin | MCF7, U937 | [13,33] | At high concentrations mangiferin can inhibit P-glycoprotein expression and chemosensitise for doxorubicin therapy [33]. Activity of the drug is enhanced by mangiferin [11]. |
| Paclitaxel | Triple negative breast cancer | [60,62] | IRAK1 overexpression confers a growth advantage [62]. Mangiferin may inhibit IRAK1 activation [60,62]. |
| Cisplatin | U937 | [11] | Inhibits ROS production [8]. Activity of the drug is enhanced by mangiferin; Impedes NFκB activation; Enhanced cell death in the presence of TNF [11]. |
| Vincristine | U937 | [11] | Inhibits ROS production [8]. Activity of the drug is enhanced by mangiferin; Impedes NFκB activation; Enhanced cell death in the presence of TNF [11]. |
| Adriamycin | U937 | [11] | Inhibits ROS production [8]. Activity of the drug is enhanced by mangiferin; Impedes NFκB activation; Enhanced cell death in the presence of TNF [11]. |
| AraC | U937 | [11] | Inhibits ROS production [8]. Activity of the drug is enhanced by mangiferin; Impedes NFκB activation; Enhanced cell death in the presence of TNF [11]. |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gold-Smith, F.; Fernandez, A.; Bishop, K. Mangiferin and Cancer: Mechanisms of Action. Nutrients 2016, 8, 396. https://doi.org/10.3390/nu8070396
Gold-Smith F, Fernandez A, Bishop K. Mangiferin and Cancer: Mechanisms of Action. Nutrients. 2016; 8(7):396. https://doi.org/10.3390/nu8070396
Chicago/Turabian StyleGold-Smith, Fuchsia, Alyssa Fernandez, and Karen Bishop. 2016. "Mangiferin and Cancer: Mechanisms of Action" Nutrients 8, no. 7: 396. https://doi.org/10.3390/nu8070396
APA StyleGold-Smith, F., Fernandez, A., & Bishop, K. (2016). Mangiferin and Cancer: Mechanisms of Action. Nutrients, 8(7), 396. https://doi.org/10.3390/nu8070396