Hawthorn Fruit Extract Elevates Expression of Nrf2/HO-1 and Improves Lipid Profiles in Ovariectomized Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Hawthorn Fruit Extract
2.2. Animals and Diets
2.3. Sample Preparation
2.4. Determination of Blood Biochemical Variables
2.5. Liver Analysis
2.6. mRNA Expression Analysis
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
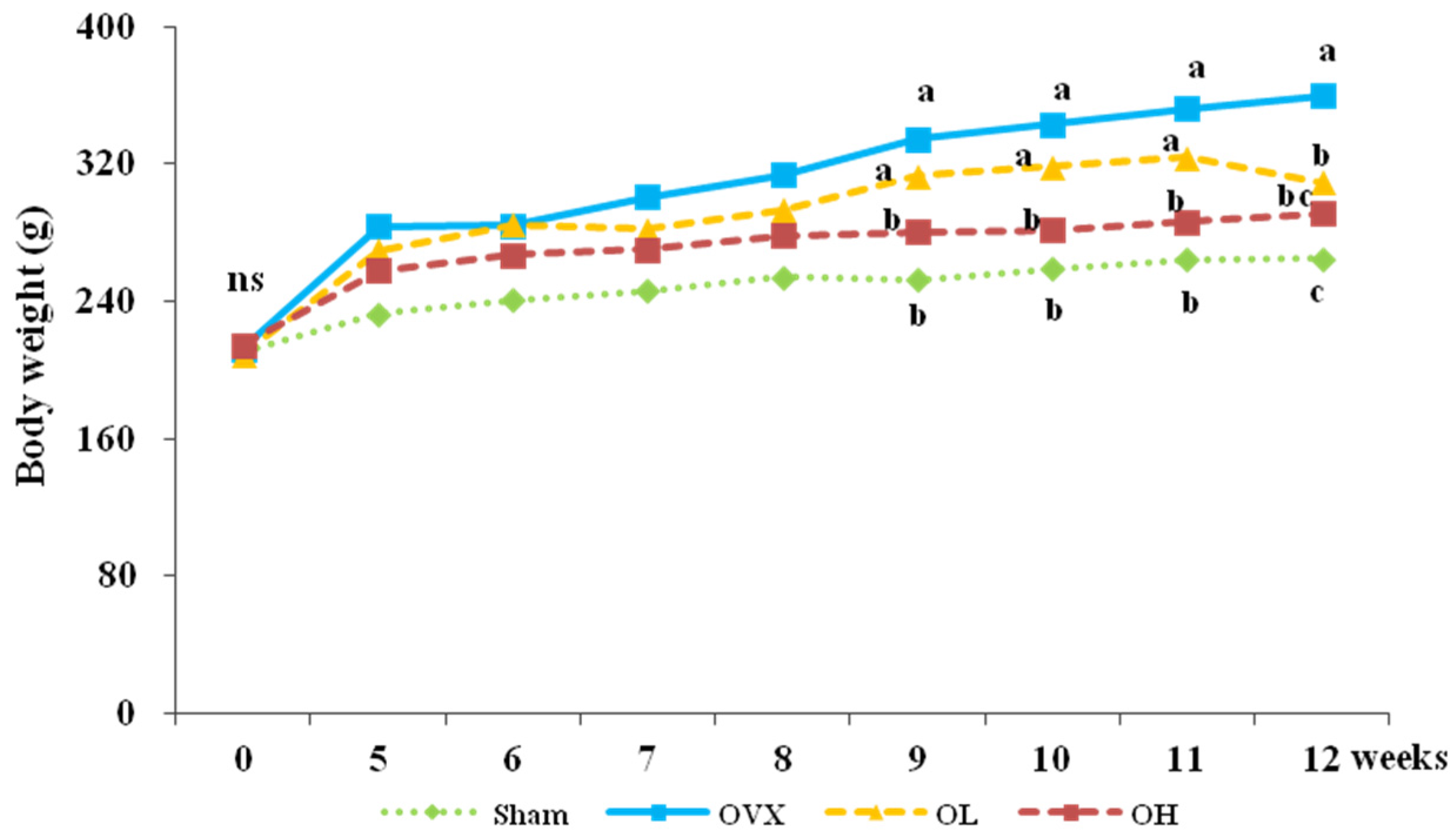
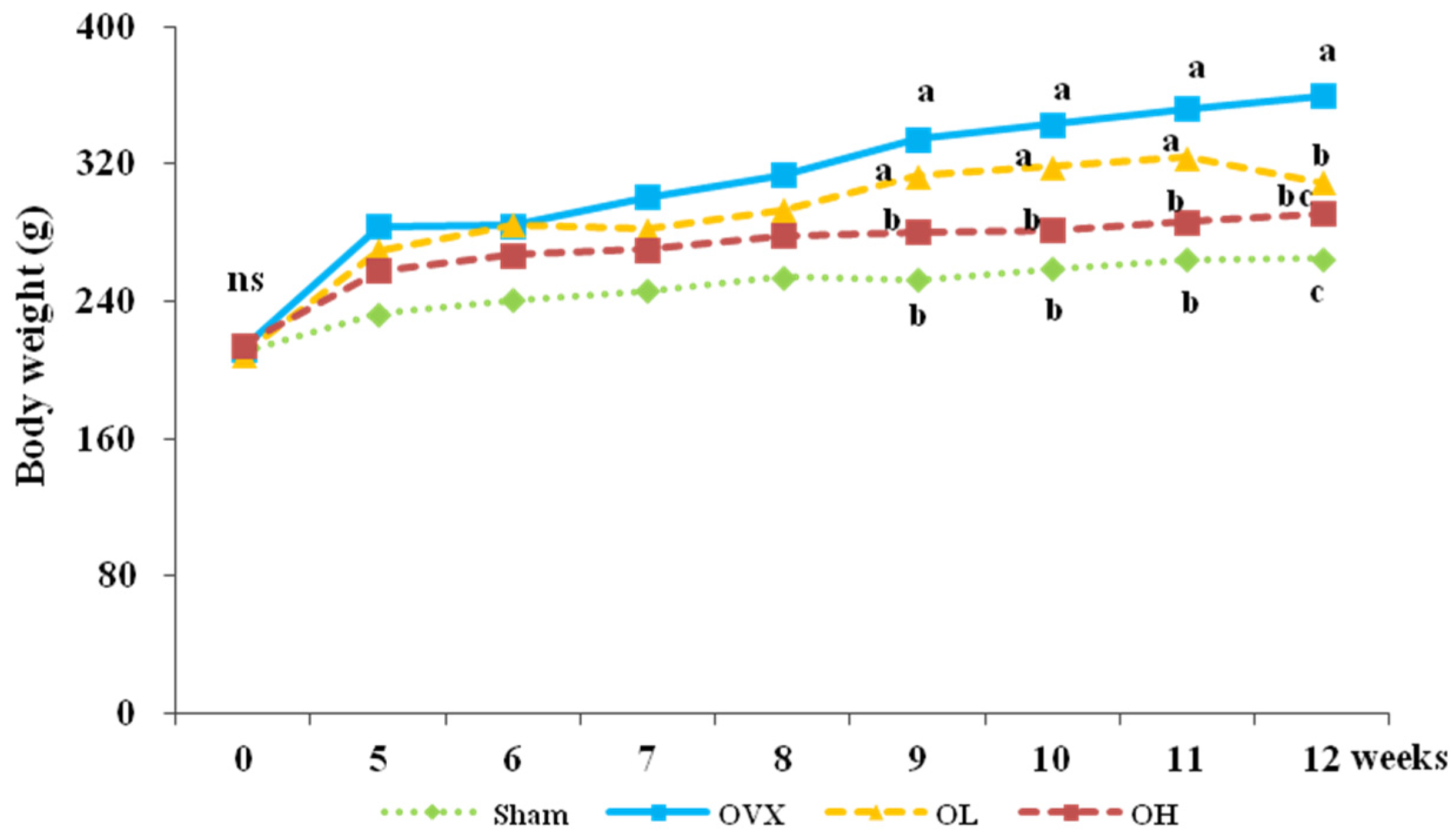
3.1. Food Intake, Food Efficiency Ratio, Body Weight, and Organ Weight
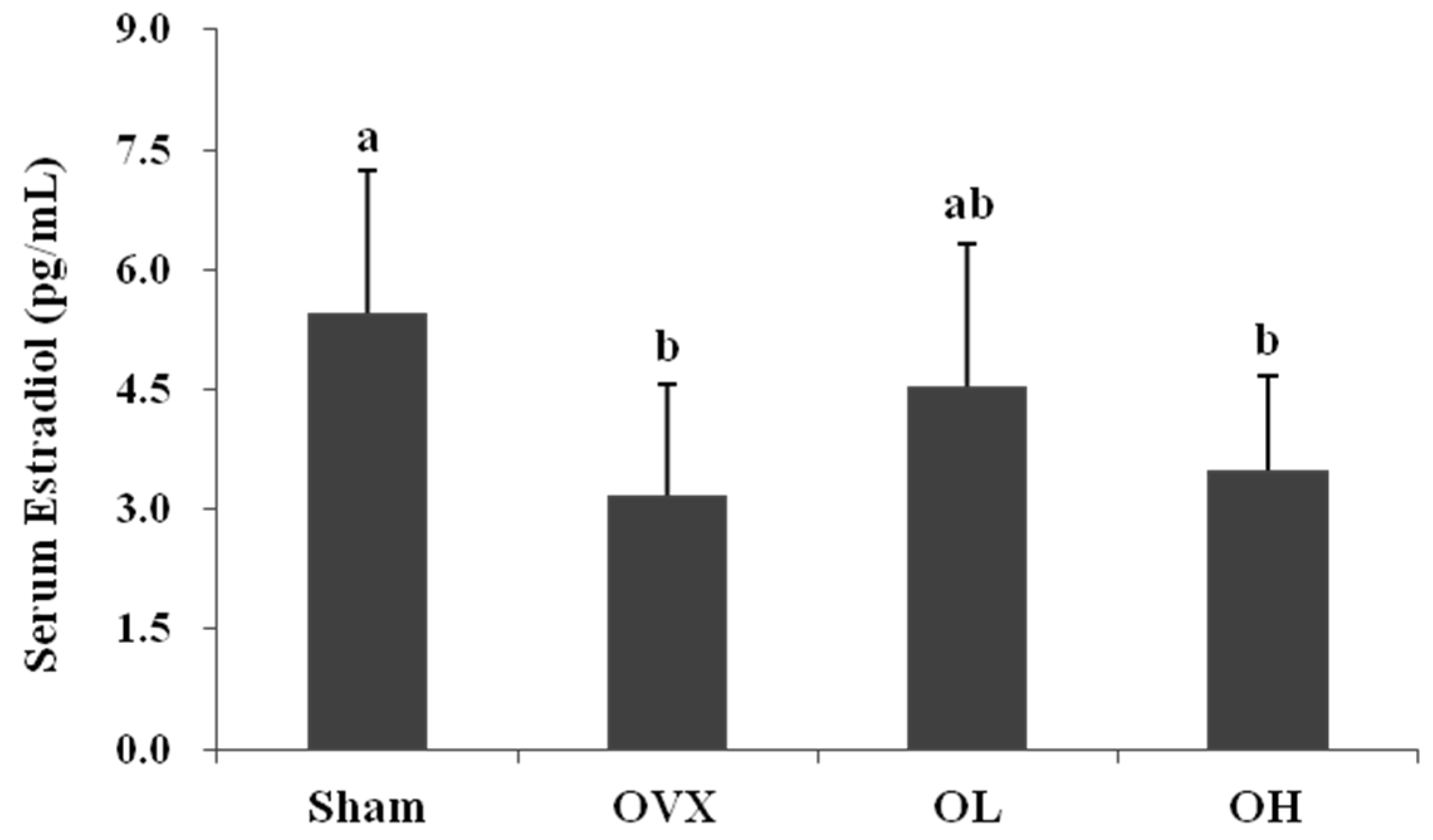
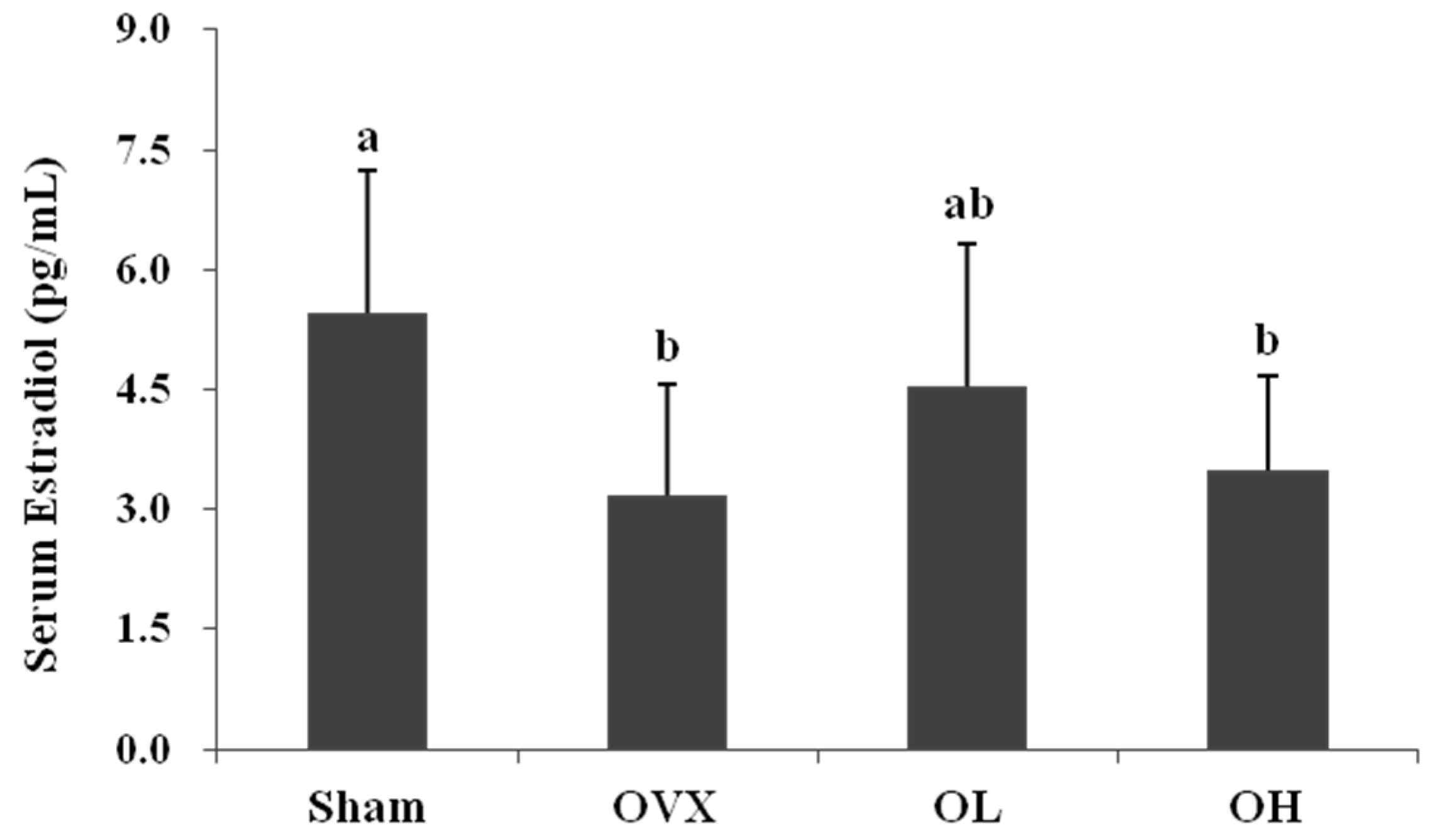
3.2. Serum Estradiol Level
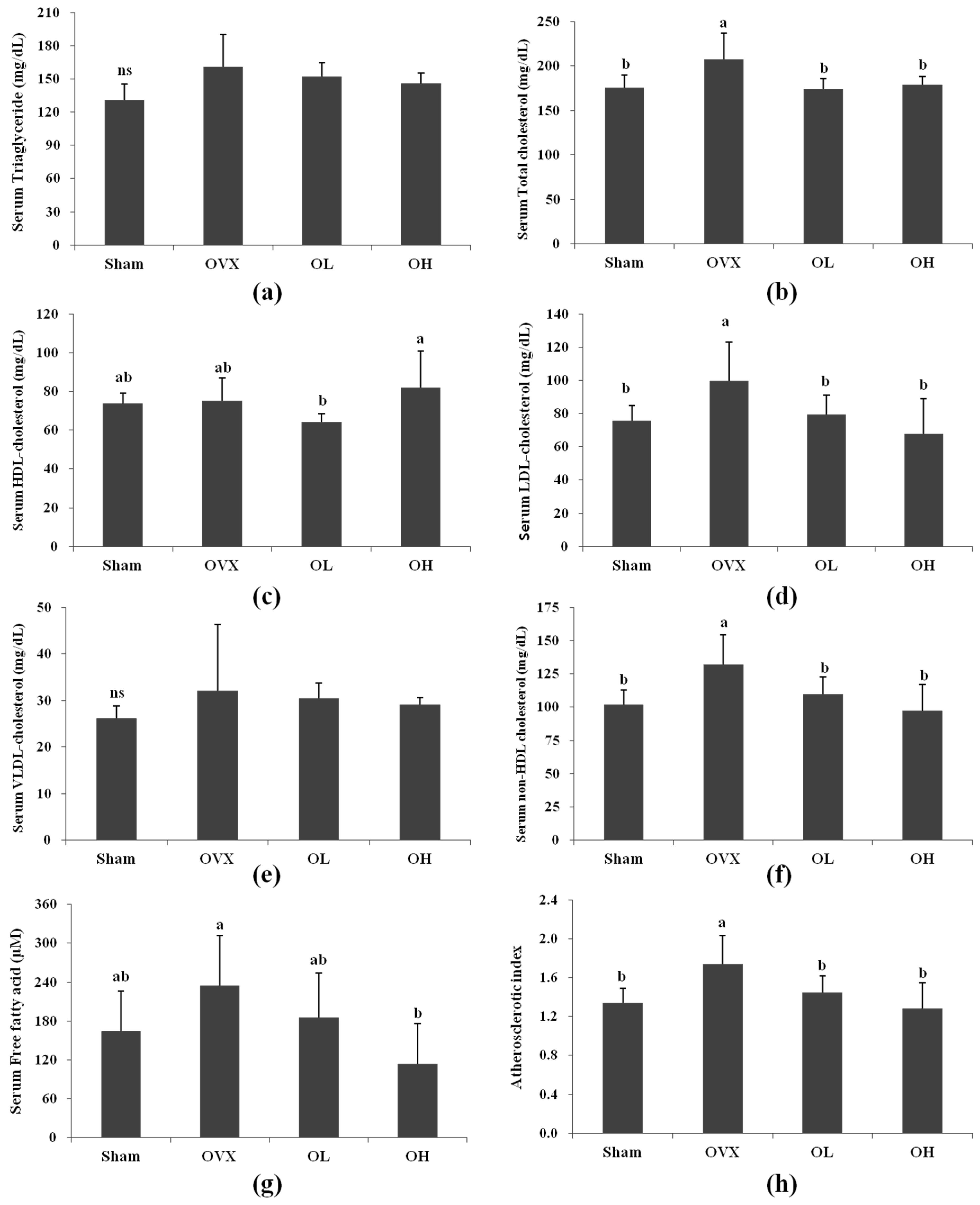
3.3. Serum Lipid Profiles
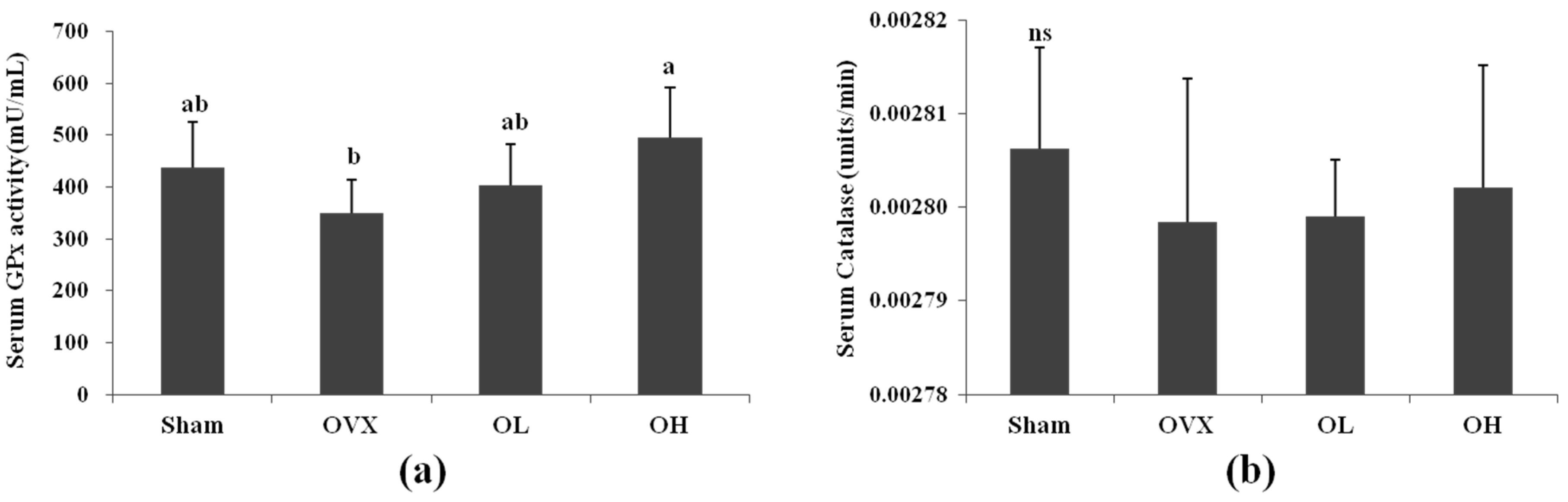
3.4. Serum Antioxidant Enzyme Activities
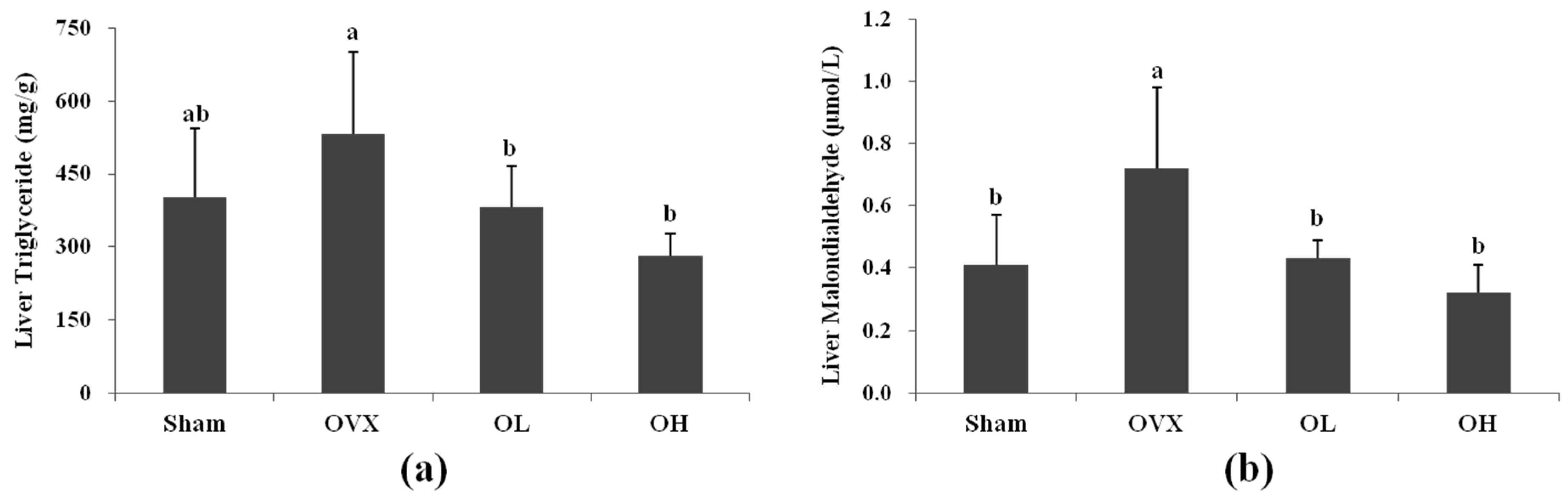
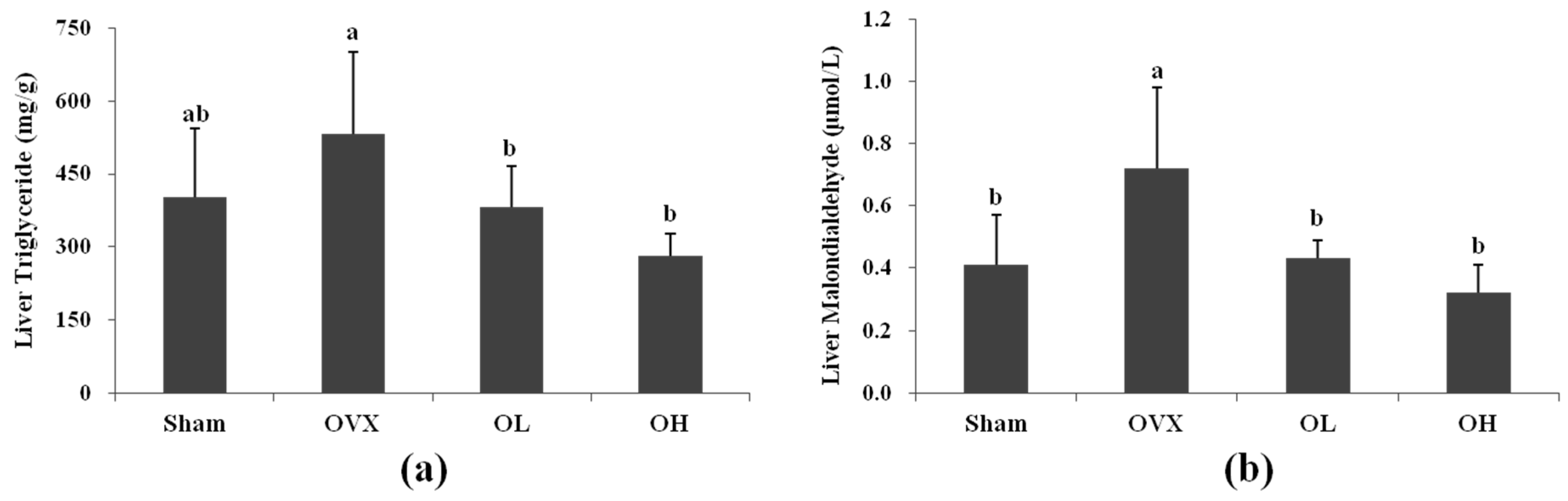
3.5. Liver TG and MDA Levels
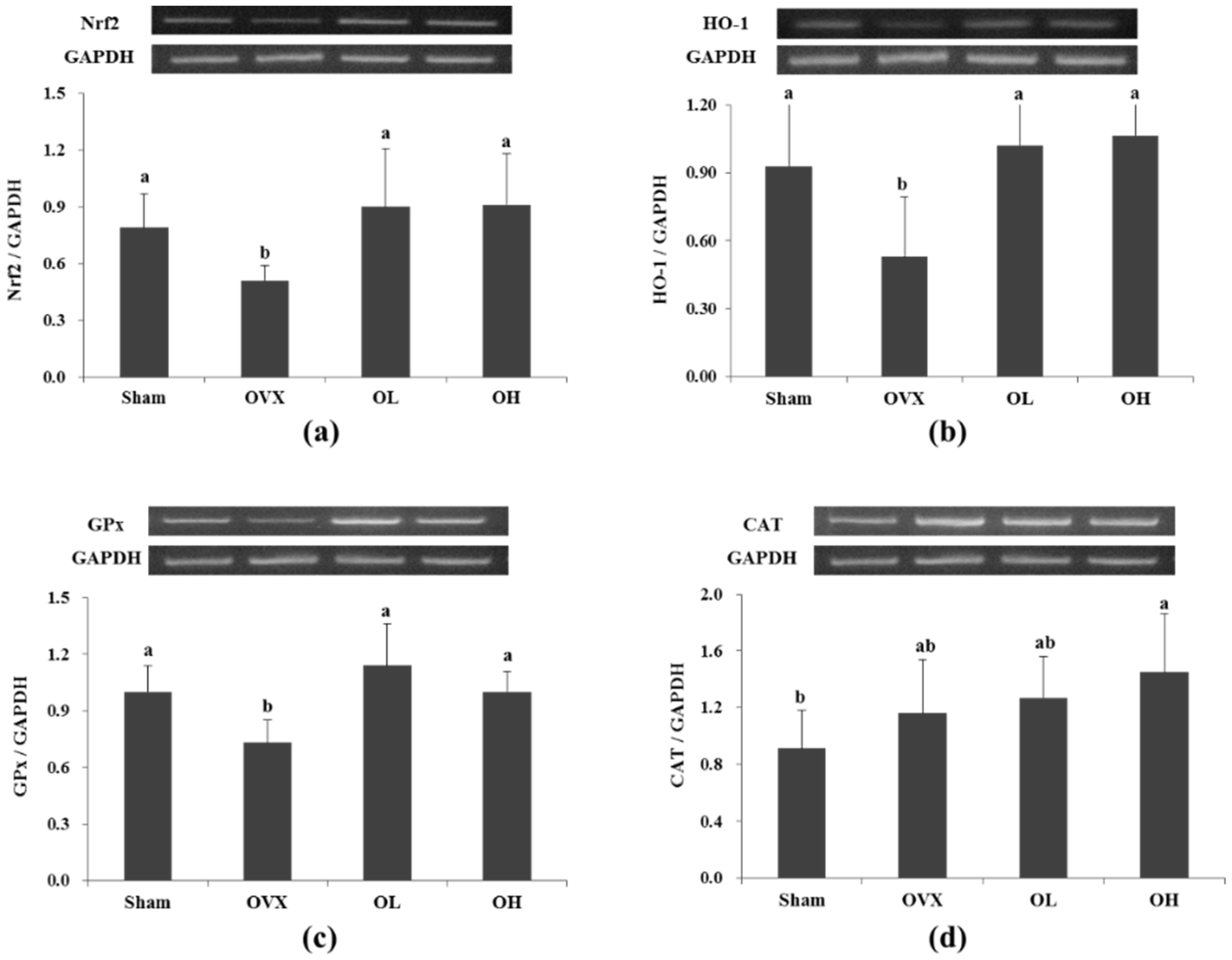
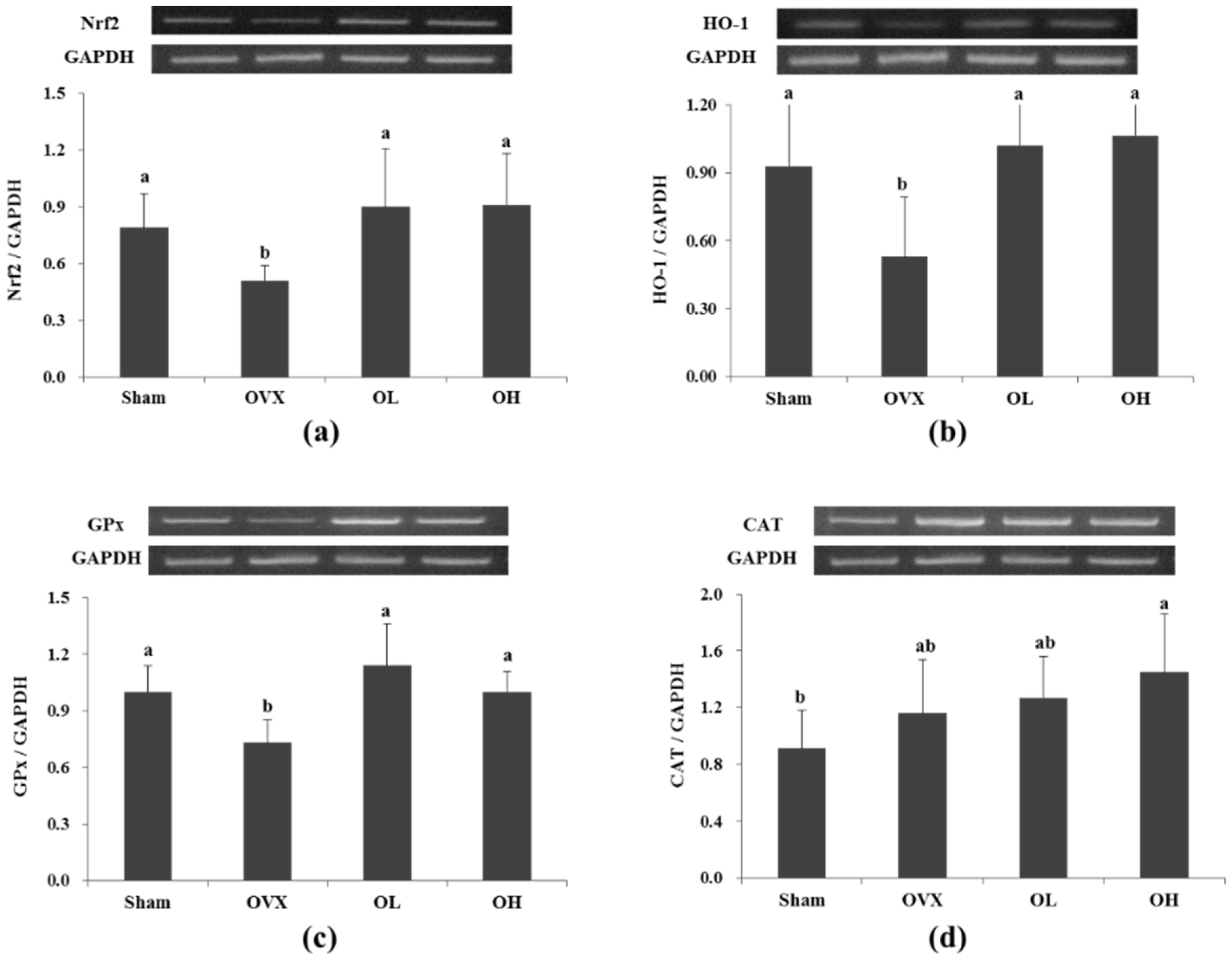
3.6. mRNA Expression of Nrf2, HO-1, GPx, and CAT in Liver
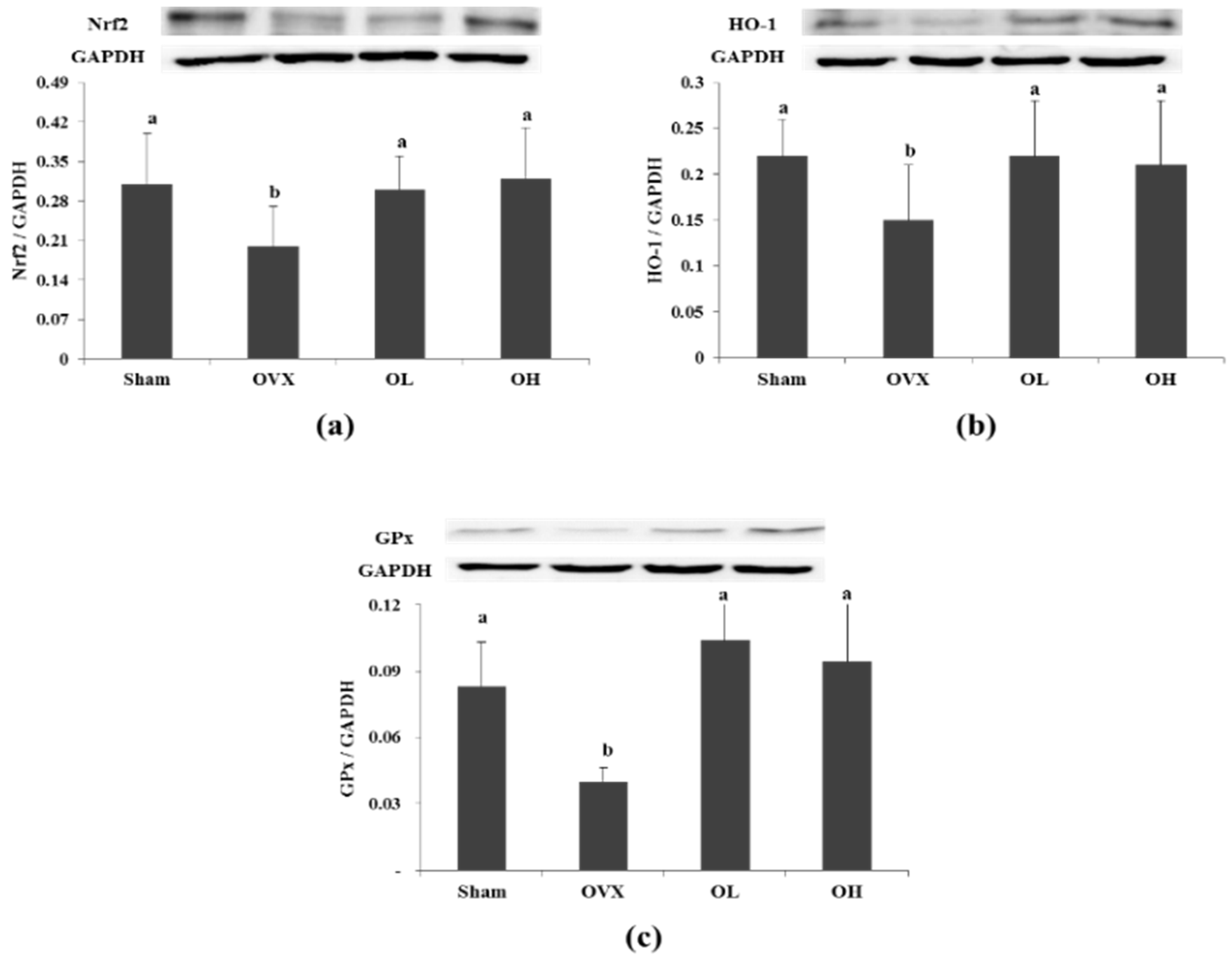
3.7. Protein Expression of Nrf2, HO-1, and GPx in Liver
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| LDL | low-density lipoprotein |
| VLDL | very low-density lipoprotein |
| HDL | high-density lipoprotein |
| ROS | reactive oxygen species |
| CAT | catalase |
| SOD | superoxide dismutase |
| GPx | glutathione peroxidase |
| Nrf2 | nuclear factor erythroid 2 related factor |
| HO-1 | heme oxygenase-1 |
| FFA | free fatty acids |
| GST | glutathione S-transferase |
| ARE | antioxidant response element |
| AI | Atherogenic Index |
| TG | triglyceride |
| MDA | malondialdehyde |
| TBA | thiobarbituric acid |
| TCA | trichloroacetic acid |
| RT-PCR | reverse transcription polymerase chain reaction |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| FER | food efficiency ratio |
| NAFLD | nonalcoholic fatty liver disease |
| ERKs | extracellular signal-regulated kinases |
| OVX | ovariectomized |
| PPAR α | peroxisome proliferator-activated receptor alpha |
| CYP7A1 | cholesterol 7alpha-hydroxylase |
References
- Stachowiak, G.; Pertynski, T. Metabolic disorders in menopause. Prz Menopauzalny 2015, 14, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.M.; Vitale, C. Menopause and cardiovascular disease: The evidence. Climacteric 2007, 10 (Suppl. S1), 19–24. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.S.; Neri, S. Behaviour of some indicators of oxidative stress in postmenopausal and fertile women. Maturitas 2006, 53, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Seals, D.R.; Jablonski, K.L. Aging and vascular endothelial function in humans. Clin. Sci. 2011, 120, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Rodriguez, M.A.; Ruiz-Ramos, M. Oxidative stress as a risk factor for osteoporosis in elderly Mexicans as characterized by antioxidant enzymes. BMC Musculoskelet. Disord. 2007, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- Aleksunes, L.M.; Manautou, J.E. Emerging role of Nrf2 in protecting against hepatic and gastrointestinal disease. Toxicol. Pathol. 2007, 35, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Hong, S.W. Melatonin downregulates nuclear erythroid 2-related factor 2 and nuclear factor-kappaB during prevention of oxidative liver injury in a dimethylnitrosamine model. J. Pineal. Res. 2009, 47, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Jozkowicz, A.; Was, H. Heme oxygenase-1 in tumors: Is it a false friend? Antioxid. Redox Signal. 2007, 9, 2099–2117. [Google Scholar] [CrossRef] [PubMed]
- Posa, A.; Szabo, R. Endogenous estrogen-mediated heme oxygenase regulation in experimental menopause. Oxid. Med. Cell. Longev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Fattah, A.A.; Fahim, A.T. Resveratrol and curcumin ameliorate di-(2-ethylhexyl) phthalate induced testicular injury in rats. Gen. Comp. Endocrinol. 2016, 225, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Chow, J.M.; Shen, S.C. Quercetin, but not rutin and quercitrin, prevention of H2O2-induced apoptosis via anti-oxidant activity and heme oxygenase 1 gene expression in macrophages. Biochem. Pharmacol. 2005, 69, 1839–1851. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.C.; Cheng, T.H. Mechanism of heme oxygenase-1 gene induction by quercetin in rat aortic smooth muscle cells. Pharmacology 2004, 71, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Dahmer, S.; Scott, E. Health effects of hawthorn. Am. Fam. Physician 2010, 81, 465–468. [Google Scholar] [PubMed]
- Yang, B.; Liu, P. Composition and health effects of phenolic compounds in hawthorn (Crataegus spp.) of different origins. J. Sci. Food Agric. 2012, 92, 1578–1590. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Peng, W. Crataegus pinnatifida: Chemical constituents, pharmacology, and potential applications. Molecules 2014, 19, 1685–1712. [Google Scholar] [CrossRef] [PubMed]
- Yonghyun, P.; Hyun-Joo, L. Effects of Korean and Chinese crataegi fructrus on the antioxidant activity and antiproliferation of cancer cells. Korean J. Community Living Sci. 2015, 26, 103–113. [Google Scholar]
- Cui, T.; Nakamura, K. Polyphenolic content and physiological activities of Chinese hawthorn extracts. Biosci. Biotechnol. Biochem. 2006, 70, 2948–2956. [Google Scholar] [CrossRef] [PubMed]
- Cui, T.; Li, J.Z. Quantification of the polyphenols and triterpene acids in Chinese hawthorn fruit by high-performance liquid chromatography. J. Agric. Food Chem. 2006, 54, 4574–4581. [Google Scholar] [CrossRef] [PubMed]
- Kao, E.S.; Wang, C.J. Anti-inflammatory potential of flavonoid contents from dried fruit of crataegus pinnatifida in vitro and in vivo. J. Agric. Food Chem. 2005, 53, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Z. Hawthorn fruit increases the antioxidant capacity and reduces lipid peroxidation in senescence-accelerated mice. Eur. Food Res. Technol. 2011, 232, 743–751. [Google Scholar] [CrossRef]
- Chang, Q.; Zuo, Z. Hawthorn. J. Clin. Pharmacol. 2002, 42, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Ling, S.; Nheu, L. Effects of four medicinal herbs on human vascular endothelial cells in culture. Int. J. Cardiol. 2008, 128, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Topal, G.; Koc, E. Effects of crataegus microphylla on vascular dysfunction in streptozotocin-induced diabetic rats. Phytother. Res. 2013, 27, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Pittler, M.H.; Schmidt, K. Hawthorn extract for treating chronic heart failure: Meta-analysis of randomized trials. Am. J. Med. 2003, 114, 665–674. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Sagor, A.T.; Chowdhury, M.R.H. Supplementation of fresh ucche (Momordica charantia L. var. muricata Willd) prevented oxidative stress, fibrosis and hepatic damage in CCl4 treated rats. BMC Complement. Altern. Med. 2015, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [PubMed]
- Folch, J.; Lees, M. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Bahorun, T.; Gressier, B. Oxygen species scavenging activity of phenolic extracts from hawthorn fresh plant organs and pharmaceutical preparations. Arzneimittelforschung 1996, 46, 1086–1089. [Google Scholar] [PubMed]
- Bahorun, T.; Trotin, F. Antioxidant activities of Crataegus monogyna extracts. Planta Med. 1994, 60, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Kallio, H. Quantitative analysis of phenolic compounds in Chinese hawthorn (Crataegus spp.) fruits by high performance liquid chromatography-electrospray ionisation mass spectrometry. Food Chem. 2011, 127, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Hwang, S.I.; Lee, M.H.; Jang, Y.S. Fruit characteristics and variation of phenolic compounds in the fruit of hawthorn (Crataegus pinnatifida Bunge) selected from Korea and Chinese cultivars. Korean J. Plant Resour. 2010, 23, 223–227. [Google Scholar]
- Muthusami, S.; Ramachandran, I. Ovariectomy induces oxidative stress and impairs bone antioxidant system in adult rats. Clin. Chim. Acta 2005, 360, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.D.; Kuo, W.W. Cardiac Fas-dependent and mitochondria-dependent apoptosis in ovariectomized rats. Maturitas 2008, 61, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Goss, P.E.; Qi, S. The effects of atamestane and toremifene alone and in combination compared with letrozole on bone, serum lipids and the uterus in an ovariectomized rat model. Breast Cancer Res. Treat. 2007, 103, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Arjmandi, B.H.; Khan, D.A. The ovarian hormone deficiency-induced hypercholesterolemia is reversed by soy protein and the synthetic isoflavone, ipriflavone. Nutr. Res. 1997, 17, 885–894. [Google Scholar] [CrossRef]
- Stevenson, J.C.; Crook, D. Influence of age and menopause on serum lipids and lipoproteins in healthy women. Atherosclerosis 1993, 98, 83–90. [Google Scholar] [CrossRef]
- Ryou, S.H.; Kang, M.S. Effects of green tea or sasa quelpaertensis bamboo leaves on plasma and liver lipids, erythrocyte na efflux, and platelet aggregation in ovariectomized rats. Nutr. Res. Pract. 2012, 6, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Hart-Unger, S.; Korach, K.S. Estrogens and obesity: Is it all in our heads? Cell Metab. 2011, 14, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Song, J. Effect of genistein on insulin resistance, renal lipid metabolism, and antioxidative activities in ovariectomized rats. Nutrition 2009, 25, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, X. Long-term resveratrol treatment prevents ovariectomy-induced osteopenia in rats without hyperplastic effects on the uterus. Br. J. Nutr. 2014, 111, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhu, J. Differential effects of polyphenols-enriched extracts from hawthorn fruit peels and fleshes on cell cycle and apoptosis in human MCF-7 breast carcinoma cells. Food Chem. 2013, 141, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Kuo, D.H.; Yeh, C.H. Effect of shanzha, a Chinese herbal product, on obesity and dyslipidemia in hamsters receiving high-fat diet. J. Ethnopharmacol. 2009, 124, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Creatsas, G.; Christodoulakos, G. Cardiovascular disease: Screening and management of the a-symptomatic high-risk post-menopausal woman. Maturitas 2005, 52 (Suppl. S1), 32–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ho, W.K. Hawthorn fruit is hypolipidemic in rabbits fed a high cholesterol diet. J. Nutr. 2002, 132, 5–10. [Google Scholar] [PubMed]
- Kwok, C.-Y.; Li, C. Cholesterol lowering and vascular protective effects of ethanolic extract of dried fruit of Crataegus pinnatifida, hawthorn (shan zha), in diet-induced hypercholesterolaemic rat model. J. Funct. Foods 2013, 5, 1326–1335. [Google Scholar] [CrossRef]
- Eigeliene, N.; Härkönen, P. Effects of estradiol and medroxyprogesterone acetate on expression of the cell cycle proteins cyclin D1, p21 and p27 in cultured human breast tissues. Cell Cycle 2008, 7, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Kankofer, M.; Radzki, R.P. Anti-oxidative/oxidative status of rat liver after ovariectomy. J. Vet. Med. Ser. A 2007, 54, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Oztekin, E.; Tiftik, A.M. Lipid peroxidation in liver tissue of ovariectomized and pinealectomized rats: Effect of estradiol and progesterone supplementation. Cell Biochem. Funct. 2007, 25, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Ha, B.J.; Lee, S.H. The role of salicornia herbacea in ovariectomy-induced oxidative stress. Biol. Pharm. Bull. 2006, 29, 1305–1309. [Google Scholar] [CrossRef] [PubMed]
- Kwok, C.-Y.; Wong, C.N.-Y. Consumption of dried fruit of Crataegus pinnatifida (hawthorn) suppresses high-cholesterol diet-induced hypercholesterolemia in rats. J. Funct. Foods 2010, 2, 179–186. [Google Scholar] [CrossRef]
- Lemini, C.; Franco, Y. Estrogenic effects of 17 β -aminoestrogens assessed in uteri of rats and mice. Eur. J. Pharmacol. 2005, 510, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Topcuoglu, A.; Uzun, H. Effects of estrogens on oxidative protein damage in plasma and tissues in ovariectomised rats. Clin. Invest. Med. 2009, 32, 133–143. [Google Scholar]
- Prasannarong, M.; Vichaiwong, K. Calorie restriction prevents the development of insulin resistance and impaired insulin signaling in skeletal muscle of ovariectomized rats. Biochim. Biophys. Acta 2012, 1822, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.J.; Carvalho, R.A. Fatty acid oxidation and cardiovascular risk during menopause: A mitochondrial connection? J. Lipids 2012, 2012, 365798. [Google Scholar] [CrossRef] [PubMed]
- Campos, L.B.; Gilglioni, E.H. Cimicifuga racemosa impairs fatty acid β-oxidation and induces oxidative stress in livers of ovariectomized rats with renovascular hypertension. Free Radic. Biol. Med. 2012, 53, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Collin, P.; Chapados, N. Time course of changes in in vitro lipolysis of intra-abdominal fat depots in relation to high-fat diet-induced hepatic steatosis in rats. Br. J. Nutr. 2006, 96, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, Y.H. Estrogen deprivation and excess energy supply accelerate 7,12-dimethylbenz(a)anthracene-induced mammary tumor growth in C3H/HeN mice. Nutr. Res. Pract. 2015, 9, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Moreno, F.N.; Campos-Shimada, L.B. Vitex agnus-castus L. (Verbenaceae) improves the liver lipid metabolism and redox state of ovariectomized rats. Evid. Based Complement. Alternat. Med. 2015, 2015, 212378. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kang, M.I. Oxidative and electrophilic stresses activate Nrf2 through inhibition of ubiquitination activity of Keap1. Mol. Cell. Biol. 2006, 26, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.K.; Itoh, K. Enhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: Role of antioxidant response element-like sequences in the Nrf2 promoter. Mol. Cell. Biol. 2002, 22, 2883–2892. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S.H.; Baranano, D.E. Heme oxygenase: A font of multiple messengers. Neuropsychopharmacology 2001, 25, 294–298. [Google Scholar] [CrossRef]
- Ogborne, R.M.; Rushworth, S.A. Haem oxygenase-1: A target for dietary antioxidants. Biochem. Soc. Trans. 2004, 32, 1003–1005. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-Q.; Gao, B.-W. Critical role of FoxO1 in granulosa cell apoptosis caused by oxidative stress and protective effects of grape seed procyanidin B2. Oxid. Med. Cell. Longev. 2016, 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yue, Y. Procyanidin B2 attenuates neurological deficits and blood-brain barrier disruption in a rat model of cerebral ischemia. Mol. Nutr. Food Res. 2015, 59, 1930–1941. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Ramiro, I.; Ramos, S. Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur. J. Nutr. 2012, 51, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Tiano, J.P.; Mauvais-Jarvis, F. Importance of oestrogen receptors to preserve functional β-cell mass in diabetes. Nat. Rev. Endocrinol. 2012, 8, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Miquel, J.; Ramirez-Bosca, A. Menopause: A review on the role of oxygen stress and favorable effects of dietary antioxidants. Arch. Gerontol. Geriatr. 2006, 42, 289–306. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Molecule Sequence (5′-3′) | Annealing | Cynyle |
|---|---|---|---|
| Nrf2 | |||
| Forward | CCATTTACGGAGACCCAC | 55 °C | 30 |
| Reverse | TGAGCGGCAACTTTATTC | ||
| HO-1 | |||
| Forward | TGCTCGCATGAACACTCTG | 60 °C | 45 |
| Reverse | TCCTCTGTCAGCAGTGCCT | ||
| GPx | |||
| Forward | CTCTCCGCGGTGGCACAGT | 58 °C | 30 |
| Reverse | CCACCACCGGGTCGGACATAC | ||
| CAT | |||
| Forward | GCGAATGGAGAGGCAGTGTAC | 55.5 °C | 30 |
| Reverse | GAGTGACGTTGTCTTCATTAGCACTG | ||
| GAPDH | |||
| Forward | CCTCTCTCTTGCTCTCAGTAT | 56 °C | 33 |
| Reverse | GTATCCGTTGTGGATCTGACA |
| Sham | OVX | OL | OH | |
|---|---|---|---|---|
| Food intake (g/day) | 14.44 ± 3.73 ns | 13.68 ± 0.32 | 13.78 ± 0.90 | 12.67 ± 0.93 |
| FER | 0.08 ± 0.02 c | 0.23 ± 0.03 a | 0.16 ± 0.08 b | 0.13 ± 0.06 b,c |
| Body weight (g) | ||||
| Initial | 210.49 ± 5.79 ns | 212.19 ± 7.12 | 209.12 ± 14.84 | 214.46 ± 6.98 |
| Final | 264.87 ± 9.95 c | 359.63 ± 19.57 a | 309.37 ± 41.19 b | 291.48 ± 36.65 b,c |
| Organ weight (g) | ||||
| Liver | 7.21 ± 0.60 b | 8.80 ± 0.95 a | 7.69 ± 0.86 b | 7.16 ± 1.06 b |
| Uterus | 0.54 ± 0.09 a | 0.13 ± 0.05 b | 0.33 ± 0.23 a,b | 0.41 ± 0.33 a |
| Uterine fat | 3.40 ± 1.16 b | 7.37 ± 2.26 a | 5.25 ± 1.93 b | 5.04 ± 1.53 b |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoo, J.-H.; Liu, Y.; Kim, H.-S. Hawthorn Fruit Extract Elevates Expression of Nrf2/HO-1 and Improves Lipid Profiles in Ovariectomized Rats. Nutrients 2016, 8, 283. https://doi.org/10.3390/nu8050283
Yoo J-H, Liu Y, Kim H-S. Hawthorn Fruit Extract Elevates Expression of Nrf2/HO-1 and Improves Lipid Profiles in Ovariectomized Rats. Nutrients. 2016; 8(5):283. https://doi.org/10.3390/nu8050283
Chicago/Turabian StyleYoo, Jeong-Hyun, Yanan Liu, and Hyun-Sook Kim. 2016. "Hawthorn Fruit Extract Elevates Expression of Nrf2/HO-1 and Improves Lipid Profiles in Ovariectomized Rats" Nutrients 8, no. 5: 283. https://doi.org/10.3390/nu8050283
APA StyleYoo, J.-H., Liu, Y., & Kim, H.-S. (2016). Hawthorn Fruit Extract Elevates Expression of Nrf2/HO-1 and Improves Lipid Profiles in Ovariectomized Rats. Nutrients, 8(5), 283. https://doi.org/10.3390/nu8050283