A Maternal High-Energy Diet Promotes Intestinal Development and Intrauterine Growth of Offspring

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Blood Sampling and Analyses
2.3. Tissue Sample Collection
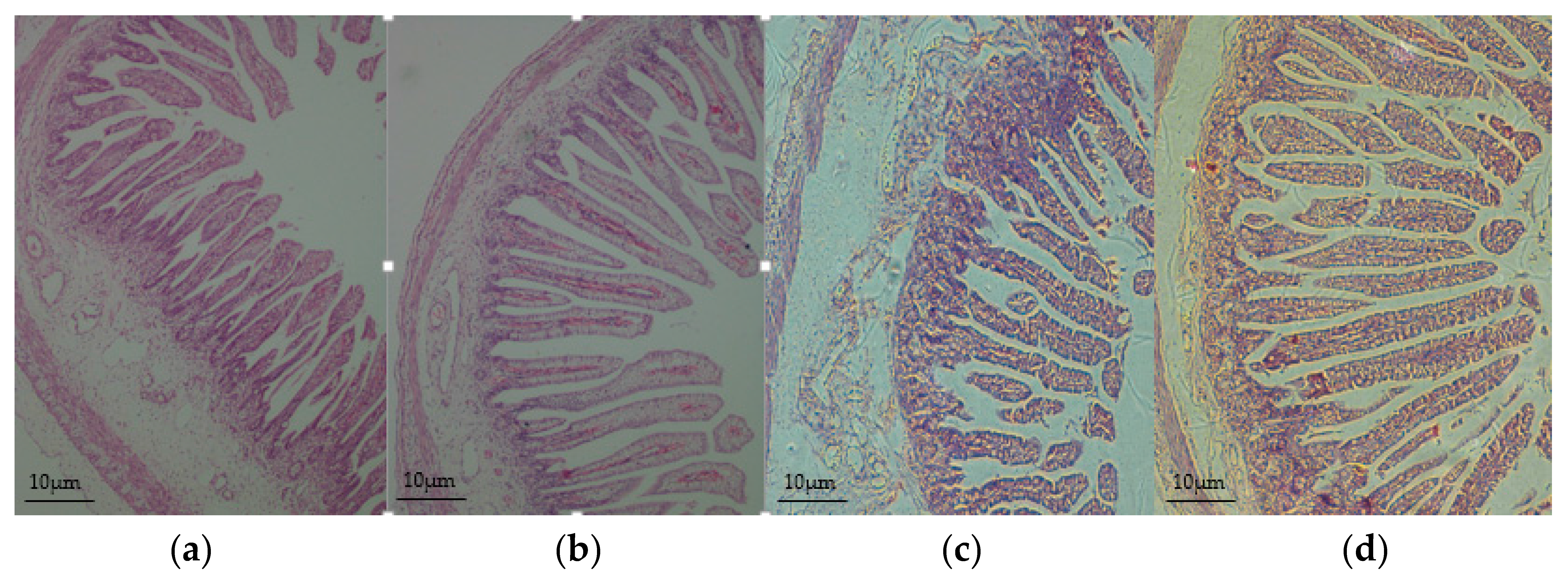
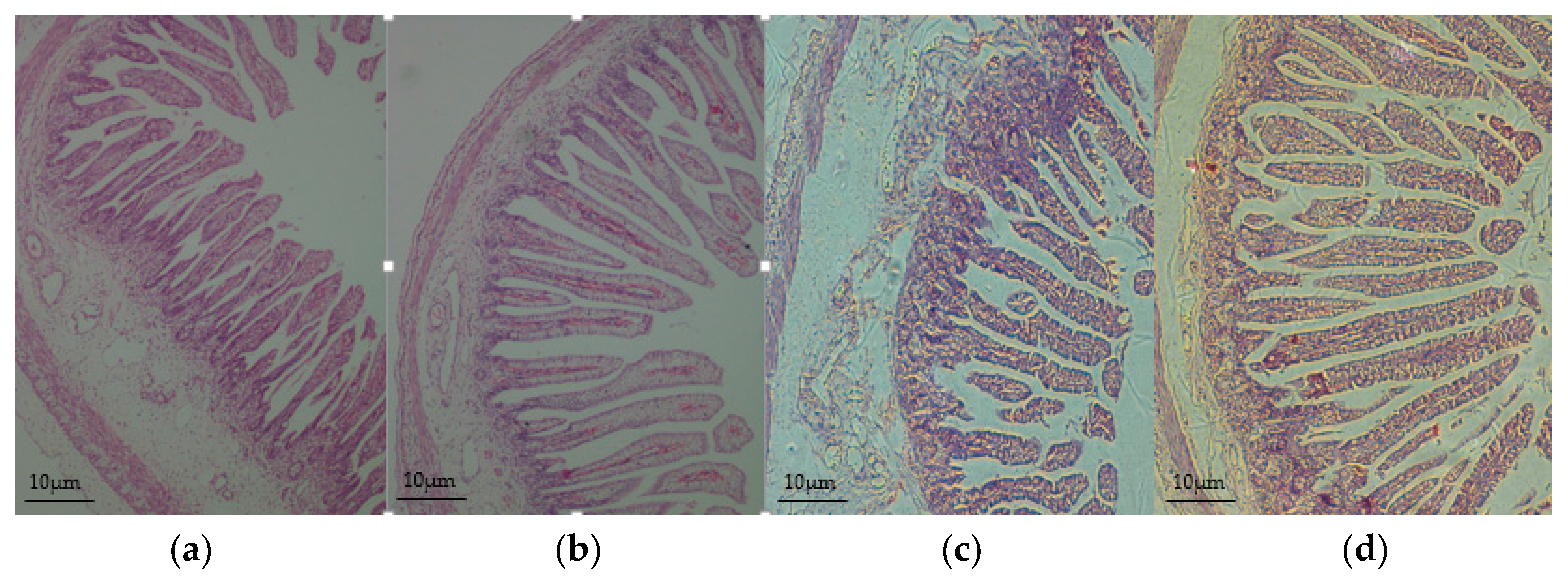
2.4. Small Intestinal Morphology
2.5. Enzyme Analyses
2.6. Total RNA Extraction and Real-Time RT-PCR
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. The BW and Small Intestine (SI) Index of Offspring Increased in the HED Group
3.2. Intestinal Morphology Improved in the Offspring of HED-Fed Mothers
3.3. The Activities of Digestive Enzyme Increased in the Jejunum and Ileum of the Offspring of HED-Fed Mothers
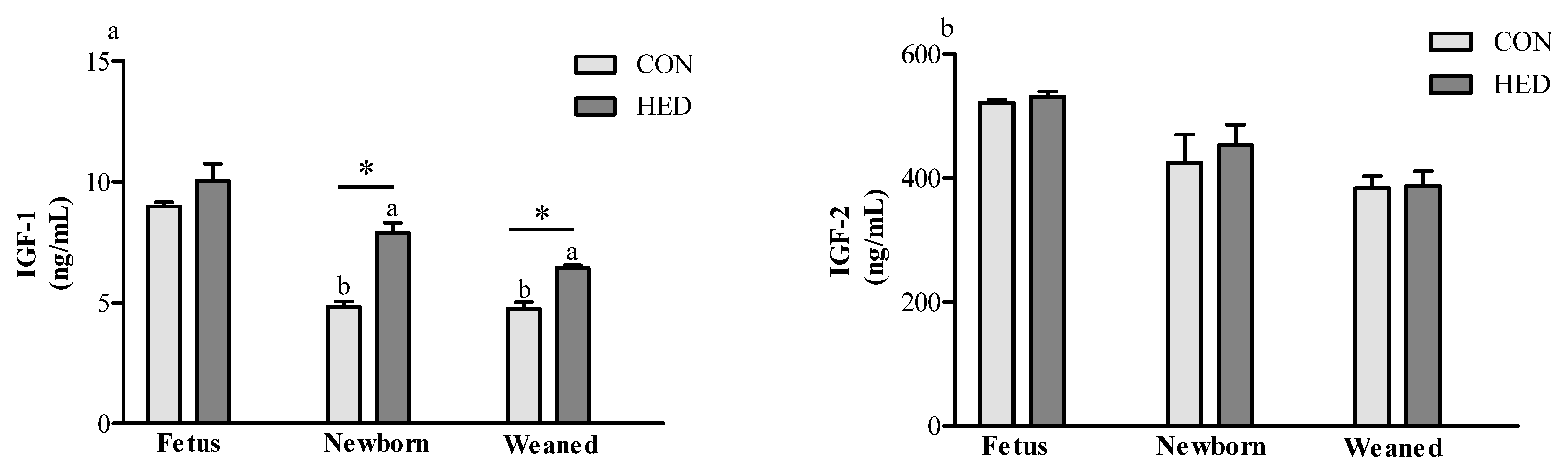
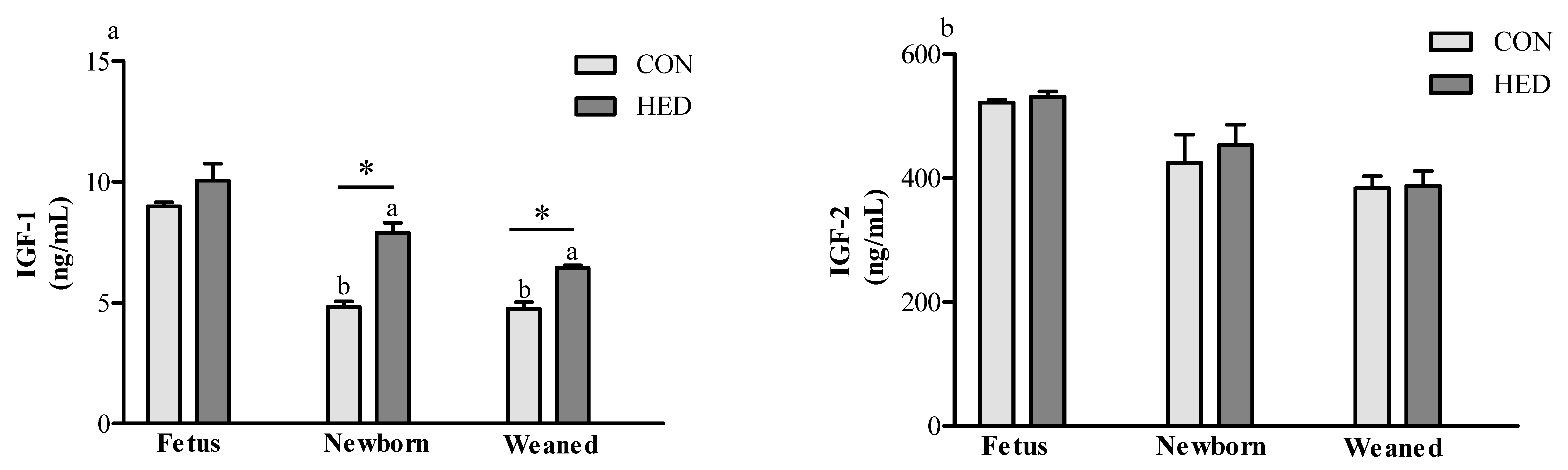
3.4. Serum Insulin-Like Growth Factor 1 (IGF-1) Concentration Increased in the Offspring of HED-Fed Mothers
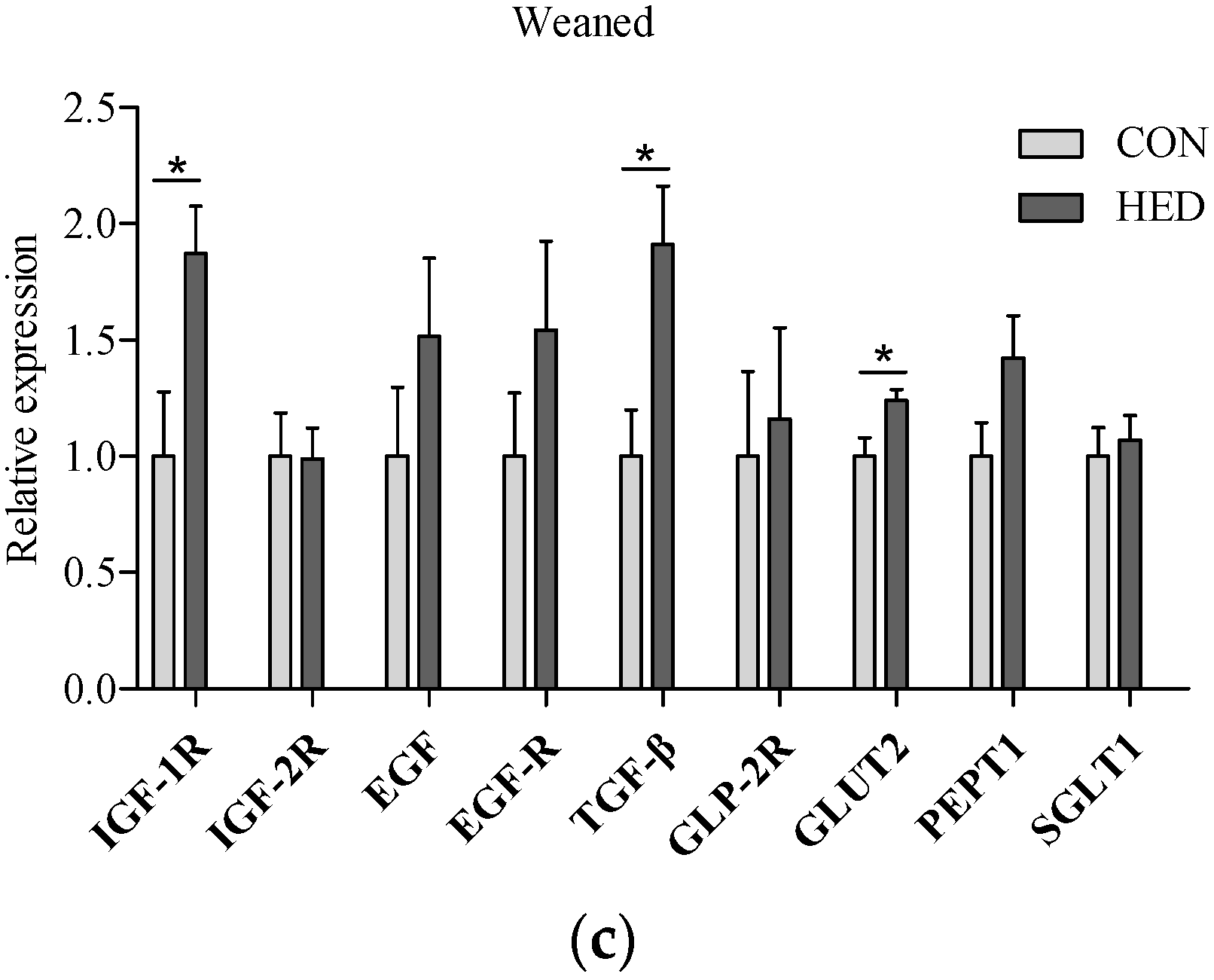
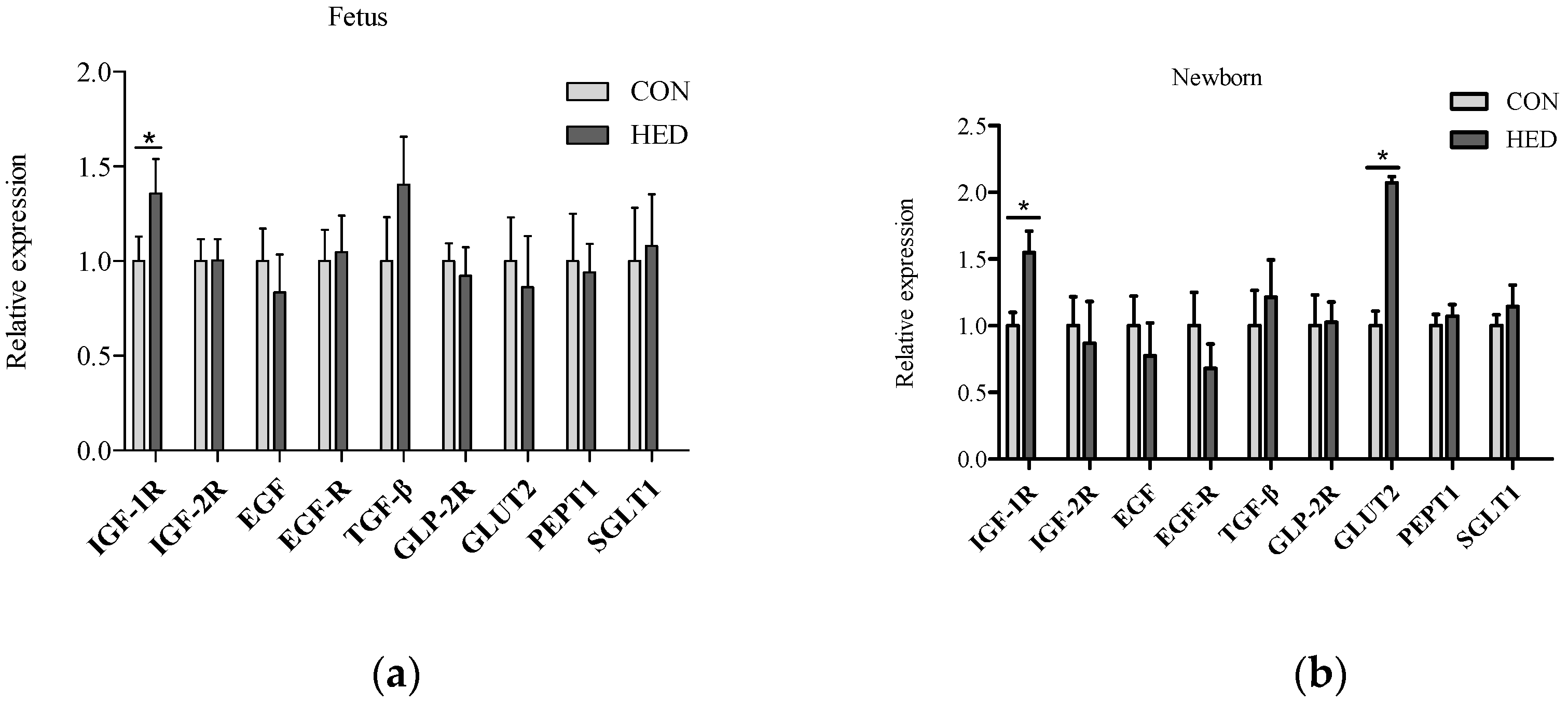
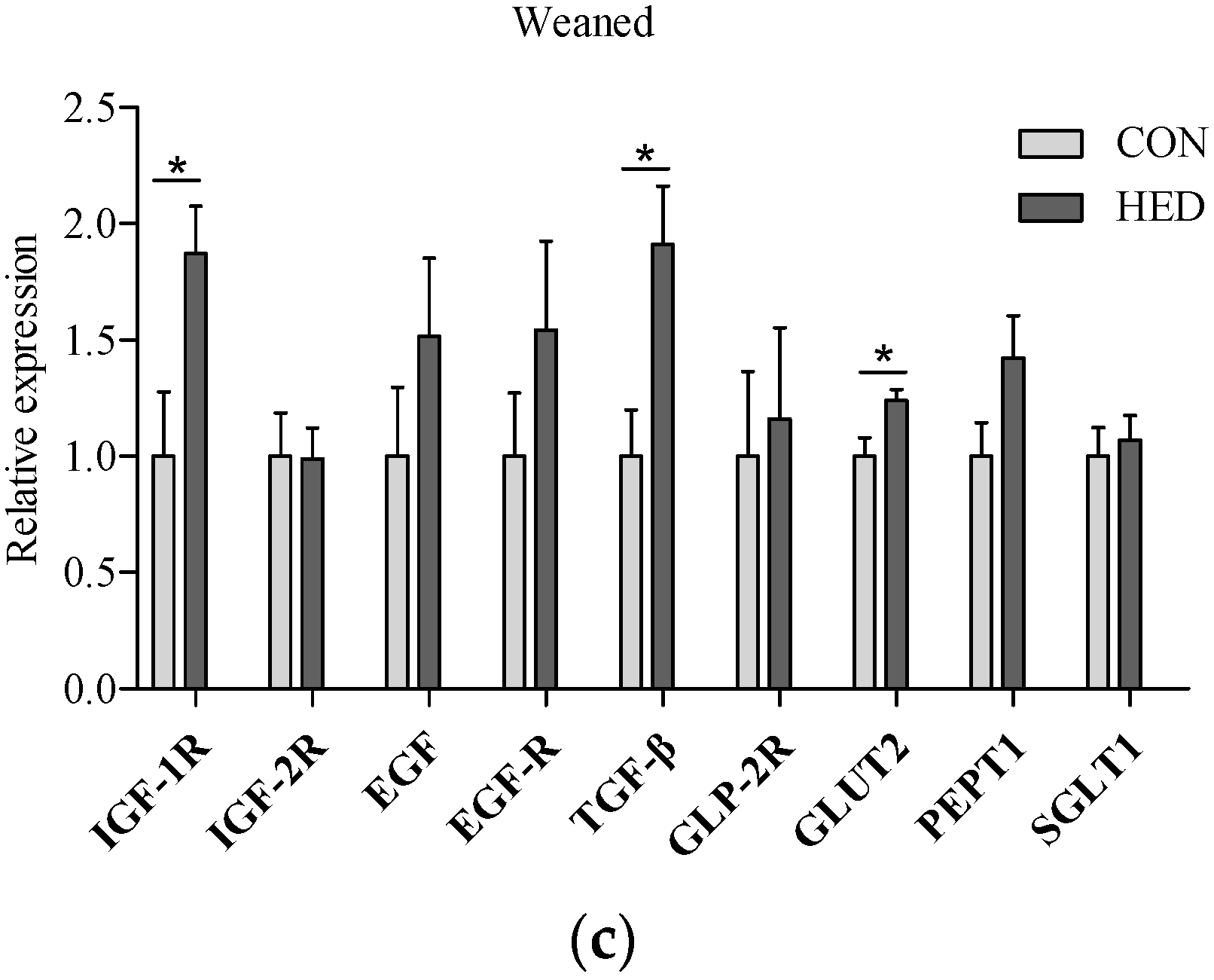
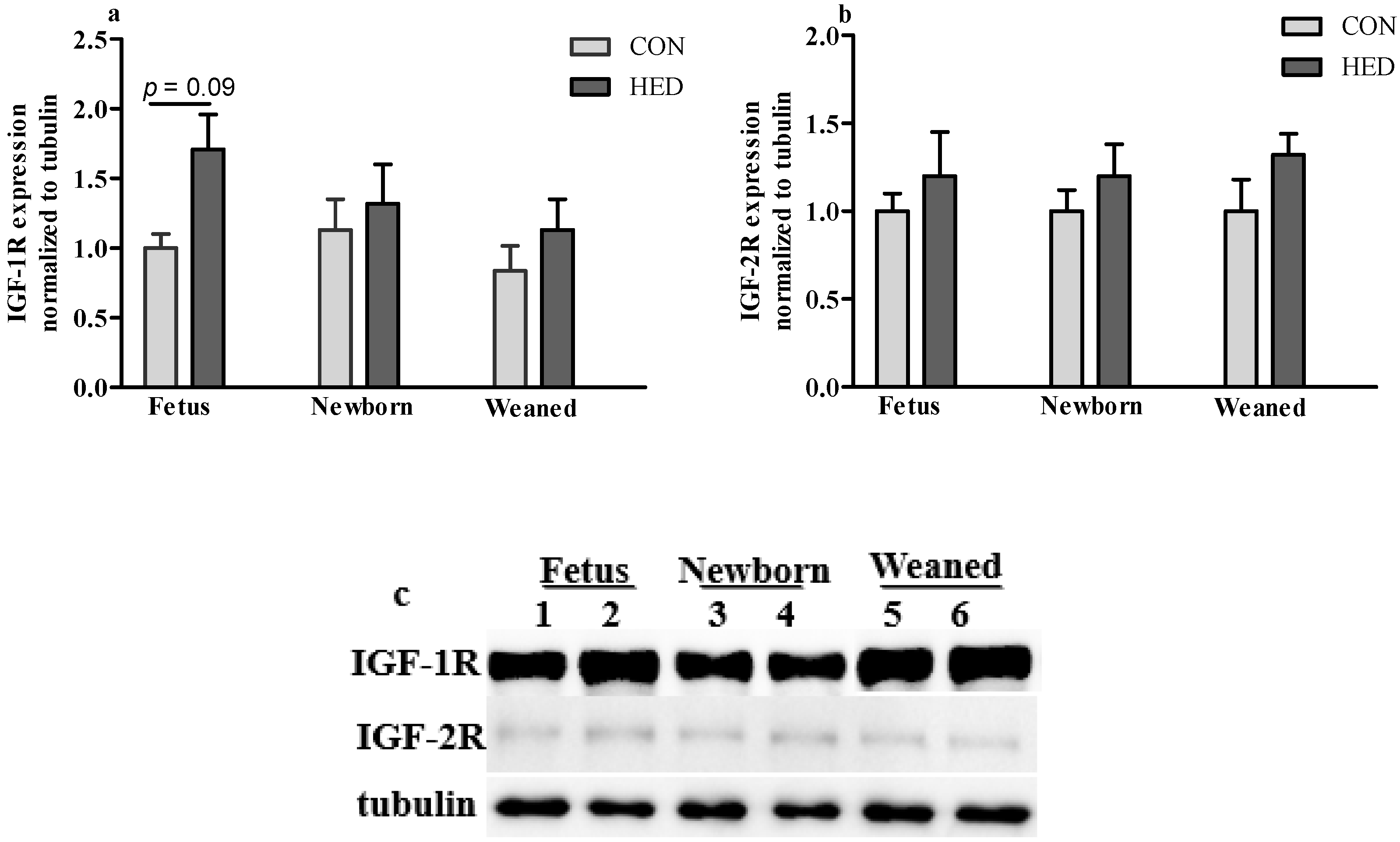
3.5. The Expression of Growth Factors Increased in Jejunum
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cromwell, G.L.; Hall, D.D.; Clawson, A.J.; Combs, G.E.; Knabe, D.A.; Maxwell, C.V.; Noland, P.R.; Orr, D.E.; Prince, T.J. Effects of additional feed during late gestation on reproductive performance of sows: A cooperative study. J. Anim. Sci. 1989, 67, 3–14. [Google Scholar] [PubMed]
- Kongsted, A.G. A review of the effect of energy intake on pregnancy rate and litter size–discussed in relation to group-housed non-lactating sows. Livest. Prod. Sci. 2005, 97, 13–26. [Google Scholar] [CrossRef]
- Herpin, P.; Damon, M.; le Dividich, J. Development of thermoregulation and neonatal survival in pigs. Livest. Prod. Sci. 2002, 78, 25–45. [Google Scholar] [CrossRef]
- Wolf, J.; Žáková, E.; Groeneveld, E. Within-litter variation of birth weight in hyperprolific Czech Large White sows and its relation to litter size traits, stillborn piglets and losses until weaning. Livest. Sci. 2008, 115, 195–205. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Sangild, P.T. Gut responses to enteral nutrition in preterm infants and animals. Exp. Biol. Med. 2006, 231, 1695–1711. [Google Scholar]
- Innis, S.M.; Dai, C.; Wu, X.; Buchan, A.M.; Jacobson, K. Perinatal lipid nutrition alters early intestinal development and programs the response to experimental colitis in young adult rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1376–G1385. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Che, L.; Wang, J.; Yang, M.; Su, G.; Fang, Z.; Lin, Y.; Xu, S.; Wu, D. Effects of maternal over- and undernutrition on intestinal morphology, enzyme activity, and gene expression of nutrient transporters in newborn and weaned pigs. Nutrition 2014, 30, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- He, Z.X.; Wu, D.Q.; Sun, Z.H.; Tan, Z.L.; Qiao, J.Y.; Ran, T.; Tang, S.X.; Zhou, C.S.; Han, X.F.; Wang, M.; et al. Protein or energy restriction during late gestation alters fetal growth and visceral organ mass: An evidence of intrauterine programming in goats. Anim. Reprod. Sci. 2013, 137, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bateson, P.; Barker, D.; Clutton-Brock, T.; Deb, D.; D’Udine, B.; Foley, R.A.; Gluckman, P.; Godfrey, K.; Kirkwood, T.; Lahr, M.M.; et al. Developmental plasticity and human health. Nature 2004, 430, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in utero and early-life conditions on adult health and disease. N. Engl. J. Med. 2008, 3, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Noblet, J.; Close, W.H.; Heavens, R.P.; Brown, D. Studies on the energy metabolism of the pregnant sow. Br. J. Nutr. 1985, 53, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Hu, L.; Xuan, Y.; Ding, X.; Luo, Y.; Bai, S.; He, S.; Zhang, K.; Che, L. Effects of high nutrient intake on the growth performance, intestinal morphology and immune function of neonatal intra-uterine growth-retarded pigs. Br. J. Nutr. 2013, 110, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Tounian, P. Programming towards childhood obesity. Ann. Nutr. Metab. 2011, 58, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Fix, J.S.; Cassady, J.P.; Herring, W.O.; Holl, J.W.; Culbertson, M.S.; See, M.T. Effect of piglet birth weight on body weight, growth, backfat, and longissimus muscle area of commercial market swine. Livest. Sci. 2010, 127, 51–59. [Google Scholar] [CrossRef]
- Drozdowski, L.A.; Clandinin, T.; Thomson, A.B. Ontogeny, growth and development of the small intestine: Understanding pediatric gastroenterology. World J. Gastroenterol. 2010, 16, 787–799. [Google Scholar] [PubMed]
- Henning, S.J. Ontogeny of enzymes in the small intestine. Annu. Rev. Physiol. 1985, 47, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, L.; Zhou, G.; Liao, Z.; Ahmad, H.; Liu, W.; Wang, T. Dietary l-arginine supplementation improves the intestinal development through increasing mucosal Akt and mammalian target of rapamycin signals in intra-uterine growth retarded piglets. Br. J. Nutr. 2012, 108, 1371–1381. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Wang, T.; Zhang, X.; Li, W. Heat shock protein 70 is upregulated in the intestine of intrauterine growth retardation piglets. Cell Stress Chaperons 2010, 15, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.M.; Neville, T.L.; Reed, J.J.; Taylor, J.B.; Reynolds, L.P.; Redmer, D.A.; Hammer, C.J.; Vonnahme, K.A.; Caton, J.S. Maternal nutritional plane and selenium supply during gestation impact visceral organ mass and intestinal growth and vascularity of neonatal lamb offspring. J. Anim. Sci. 2013, 91, 2628–2639. [Google Scholar] [CrossRef] [PubMed]
- Trahair, J.F.; Harding, R. Ultrastructural anomalies in the fetal small intestine indicate that fetal swallowing is important for normal development: An experimental study. Virchows Arch. 1992, 420, 305–312. [Google Scholar] [CrossRef]
- Spencer, J.A.; Chang, T.C.; Jones, J.; Robson, S.C.; Preece, M.A. Third trimester fetal growth and umbilical venous blood concentrations of IGF-1, IGFBP-1, and growth hormone at term. Arch. Dis. Child. Fetal Neonatal Ed. 1995, 73, F87–F90. [Google Scholar] [CrossRef] [PubMed]
- Kimble, R.M.; Breier, B.H.; Gluckman, P.D.; Harding, J.E. Enteral IGF-I enhances fetal growth and gastrointestinal development in oesophageal ligated fetal sheep. J. Endocrinol. 1999, 162, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Toms, D.; Yin, Y.; Cheung, Q.; Gong, J.; De Lange, K.; Li, J. Epidermal growth factor-expressing Lactococcus lactis enhances intestinal development of early-weaned pigs. J. Nutr. 2010, 140, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Osswald, C.; Baumgarten, K.; Stümpel, F.; Gorboulev, V.; Akimjanova, M.; Knobeloch, K.P.; Horak, I.; Kluge, R.; Joost, H.G.; Koepsell, H. Mice without the regulator gene Rsc1A1 exhibit increased Na+-d-glucose cotransport in small intestine and develop obesity. Mol. Cell. Biol. 2005, 25, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Gabler, N.K.; Spencer, J.D.; Webel, D.M.; Spurlock, M.E. In utero and postnatal exposure to long chain (n-3) PUFA enhances intestinal glucose absorption and energy stores in weanling pigs. J. Nutr. 2007, 137, 2351–2358. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′-3′) | GenBank Accession | |
|---|---|---|---|
| 28S | Sense | TACCCATATCCGCAGCAGGTC | 4W24_5 |
| Antisense | CCCTTAGAGCCAATCCTTATCCC | ||
| IGF-1R | Sense | GGAGGAAGTGACAGGGACTAAAGG | NM_214172.1 |
| Antisense | GGTGCCAGGTGATGATGATGC | ||
| IGF-2R | Sense | CGCTTTCATCATCCGCTTCG | JQ250827.1 |
| Antisense | CAGCGCGGTTTCAAAGTCAA | ||
| EGF | Sense | TCCTGTCAGCTAACCCATTACG | NM_214020.1 |
| Antisense | TGAGTTACCGAGTGATTCTCCC | ||
| EGFR | Sense | GGGATAGGGATTGGCGAGTT | NM-2140075 |
| Antisense | GCCACGTATGATTTCCAGGTTC | ||
| TGF-β | Sense | AAGCGGCAACAAAATCTATG | AF101033.1 |
| Antisense | CCCGAGAGAGCAATACAGGT | ||
| GLP-2-R | Sense | TGGCAGGACGACTCCCAGTG | NM_001246266.1 |
| Antisense | CAGGATGAAAGAGGCAAACAGG | ||
| SGLT1 | Sense | CCACTTTCCCTATAAAACCTCAC | NM_001164021.1 |
| Antisense | CTCCATCAAACTTCCATCCTCAG | ||
| GLUT2 | Sense | CCTGCTTGGTCTATCTGCTGTG | NM_001097417.1 |
| Antisense | TTGATGCTTCTTCCCTTTCTTT | ||
| PEPT1 | Sense | GATGAAATGTGAGCGTATGGG | AY180903.1 |
| Antisense | AAAGAGGGAGGATCTGGAAAA | ||
| Parameter | Fetus | Newborn Piglets | Weaned Piglets | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | HED | p-Value | CON | HED | p-Value | CON | HED | p-Value | |
| BW (Kg) | 0.65 ± 0.03 | 0.78 ± 0.01 | 0.016 | 1.33 ± 0.21 | 1.58 ± 0.13 | 0.031 | 5.53 ± 0.31 | 6.91 ± 0.29 | 0.009 |
| SI (cm) | 96.23 ± 3.65 | 110.45 ± 4.60 | 0.053 | 277.17 ± 50.63 | 288.67 ± 14.51 | 0.604 | 653.38 ± 38.77 | 738.67 ± 35.30 | 0.135 |
| SI (g) | 9.93 ± 0.80 | 15.84 ± 1.44 | <0.001 | 34.87 ± 6.72 | 47.65 ± 7.80 | 0.013 | 200.30 ± 6.23 | 233.43 ± 12.56 | 0.040 |
| SI (cm·kg−1 BW) | 151.48 ± 4.85 | 144.89 ± 6.13 | 0.446 | 210.83 ± 18.70 | 183.74 ± 5.56 | 0.195 | 119.06 ± 6.23 | 107.57 ± 5.65 | 0.202 |
| SI (g·kg−1 BW) | 15.63 ± 0.50 | 20.80 ± 1.00 | 0.004 | 26.10 ± 1.13 | 30.20 ± 1.68 | 0.071 | 36.91 ± 2.69 | 33.94 ± 1.80 | 0.380 |
| SI weight/length (mg·cm−1) | 103.19 ± 4.25 | 137.41 ± 3.47 | 0.002 | 128.36 ± 12.40 | 164.73 ± 9.50 | 0.042 | 311.47 ± 19.01 | 315.66 ± 4.37 | 0.834 |
| Parameter | D 0 | D 90 | D 114 | ||||||
| CON | HED | p-Value | CON | HED | p-value | CON | HED | p-Value | |
| Sows BW (Kg) | 135.60 ± 0.80 | 135.48 ± 1.07 | 0.927 | 189.30 ± 1.92 | 202.65 ± 3.55 | 0.005 | 210.85 ± 6.19 | 221.93 ± 13.01 | 0.049 |
| Parameter | Fetus | Newborn Piglets | Weaned Piglets | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | HED | p-Value | CON | HED | p-Value | CON | HED | p-Value | |
| Jejunum | |||||||||
| Villus height (μm) | 25.62 ± 1.12 | 29.45 ± 1.43 | 0.033 | 109.65 ± 4.77 | 126.06 ± 3.24 | 0.004 | 68.58 ± 7.69 | 72.66 ± 3.36 | 0.182 |
| Crypt depth (μm) | 3.82 ± 0.26 | 4.45 ± 0.38 | 0.791 | 22.37 ± 0.51 | 23.39 ± 0.55 | 0.212 | 33.73 ± 0.90 | 38.87 ± 1.09 | <0.001 |
| VCR | 5.90 ± 0.36 | 6.98 ± 0.66 | 0.174 | 2.08 ± 0.28 | 5.17 ± 0.25 | 0.001 | 2.15 ± 0.07 | 2.18 ± 0.09 | 0.787 |
| Ileum | |||||||||
| Villus height (μm) | 24.12 ± 1.20 | 28.44 ± 1.23 | 0.043 | 112.05 ± 4.10 | 113.22 ± 4.82 | 0.854 | 54.02 ± 1.54 | 60.28 ± 2.31 | 0.031 |
| Crypt depth (μm) | 3.92 ± 0.26 | 4.05 ± 0.38 | 0.791 | 33.48 ± 1.82 | 34.11 ± 1.84 | 0.606 | 27.40 ± 0.86 | 30.17 ± 1.50 | 0.098 |
| VCR | 6.90 ± 0.36 | 7.98 ± 0.66 | 0.174 | 3.75 ± 0.16 | 4.12 ± 0.25 | 0.189 | 2.03 ± 0.14 | 2.45 ± 0.09 | 0.014 |
| Duodenum | |||||||||
| Villus height (μm) | 15.62 ± 1.12 | 19.45 ± 1.43 | 0.333 | 76.55 ± 1.91 | 84.31 ± 4.48 | 0.066 | 105.21 ± 2.70 | 103.87 ± 3.36 | 0.757 |
| Crypt depth (μm) | 4.42 ± 0.26 | 4.45 ± 0.38 | 0.791 | 38.42 ± 0.84 | 38.37 ± 1.06 | 0.970 | 54.56 ± 1.33 | 50.54 ± 1.54 | 0.054 |
| VCR | 2.90 ± 0.36 | 2.98 ± 0.66 | 0.874 | 2.08 ± 0.07 | 2.26 ± 0.12 | 0.148 | 2.10 ± 0.08 | 2.22 ± 0.10 | 0.367 |
| U/mg Protein | Fetus | Newborn Piglets | Weaned Piglets | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | HED | p-Value | CON | HED | p-Value | CON | HED | p-Value | |
| Jejunum | |||||||||
| Lactase | 105.91 ± 10.21 | 68.71 ± 2.36 | 0.012 | 96.89 ± 2.52 | 93.57 ± 44.38 | 0.943 | 50.61 ± 6.34 | 85.08 ± 11.86 | 0.043 |
| Maltase | 35.73 ± 3.71 | 32.14 ± 2.15 | 0.435 | 23.59 ± 1.49 | 30.47 ± 3.83 | 0.145 | 92.93 ± 46.47 | 105.16 ± 52.56 | 0.115 |
| Sucrase | - | - | - | 1.42 ± 0.17 | 4.37 ± 0.18 | <0.001 | 66.88 ± 5.74 | 151.12 ± 18.33 | 0.005 |
| Ileum | |||||||||
| Lactase | 33.40 ± 4.73 | 28.49 ± 4.56 | 0.483 | 10.83 ± 0.99 | 20.48 ± 0.91 | <0.001 | 0.12 ± 0.02 | 0.18 ± 0.03 | 0.199 |
| Maltase | 16.84 ± 1.78 | 16.85 ± 0.81 | 0.996 | 13.82 ± 1.43 | 16.46 ± 2.29 | 0.366 | 50.32 ± 10.78 | 79.87 ± 11.02 | 0.104 |
| Sucrase | - | - | - | 0.22 ± 0.04 | 1.35 ± 0.21 | 0.002 | 8.67 ± 0.76 | 17.81 ± 1.19 | 0.001 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, P.; Che, L.; Yang, Z.; Feng, B.; Che, L.; Xu, S.; Lin, Y.; Fang, Z.; Li, J.; Wu, D. A Maternal High-Energy Diet Promotes Intestinal Development and Intrauterine Growth of Offspring. Nutrients 2016, 8, 258. https://doi.org/10.3390/nu8050258
Liu P, Che L, Yang Z, Feng B, Che L, Xu S, Lin Y, Fang Z, Li J, Wu D. A Maternal High-Energy Diet Promotes Intestinal Development and Intrauterine Growth of Offspring. Nutrients. 2016; 8(5):258. https://doi.org/10.3390/nu8050258
Chicago/Turabian StyleLiu, Peilin, Long Che, Zhenguo Yang, Bin Feng, Lianqiang Che, Shengyu Xu, Yan Lin, Zhengfeng Fang, Jian Li, and De Wu. 2016. "A Maternal High-Energy Diet Promotes Intestinal Development and Intrauterine Growth of Offspring" Nutrients 8, no. 5: 258. https://doi.org/10.3390/nu8050258
APA StyleLiu, P., Che, L., Yang, Z., Feng, B., Che, L., Xu, S., Lin, Y., Fang, Z., Li, J., & Wu, D. (2016). A Maternal High-Energy Diet Promotes Intestinal Development and Intrauterine Growth of Offspring. Nutrients, 8(5), 258. https://doi.org/10.3390/nu8050258