Resveratrol and Cardiovascular Diseases

Abstract
:1. Introduction
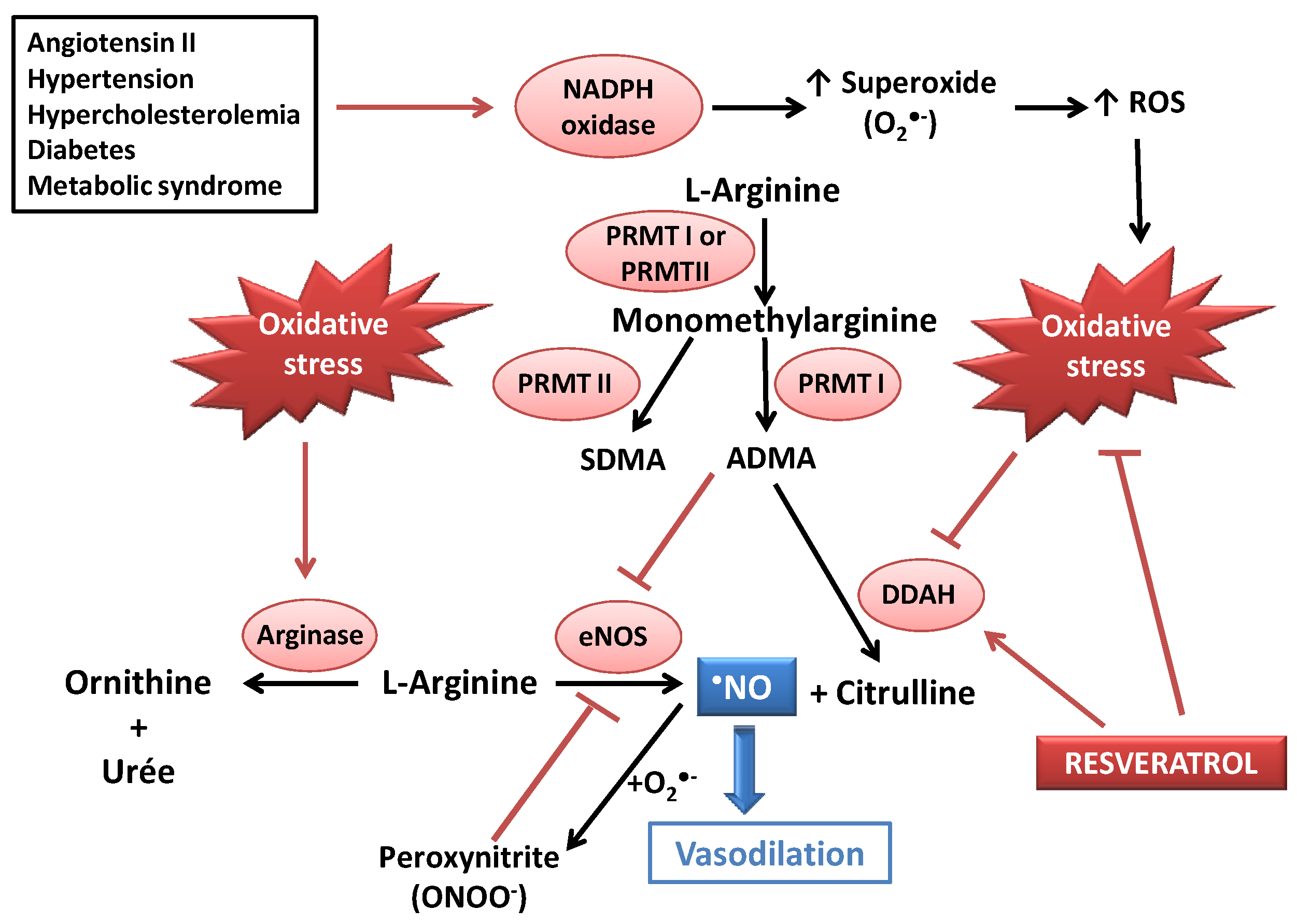
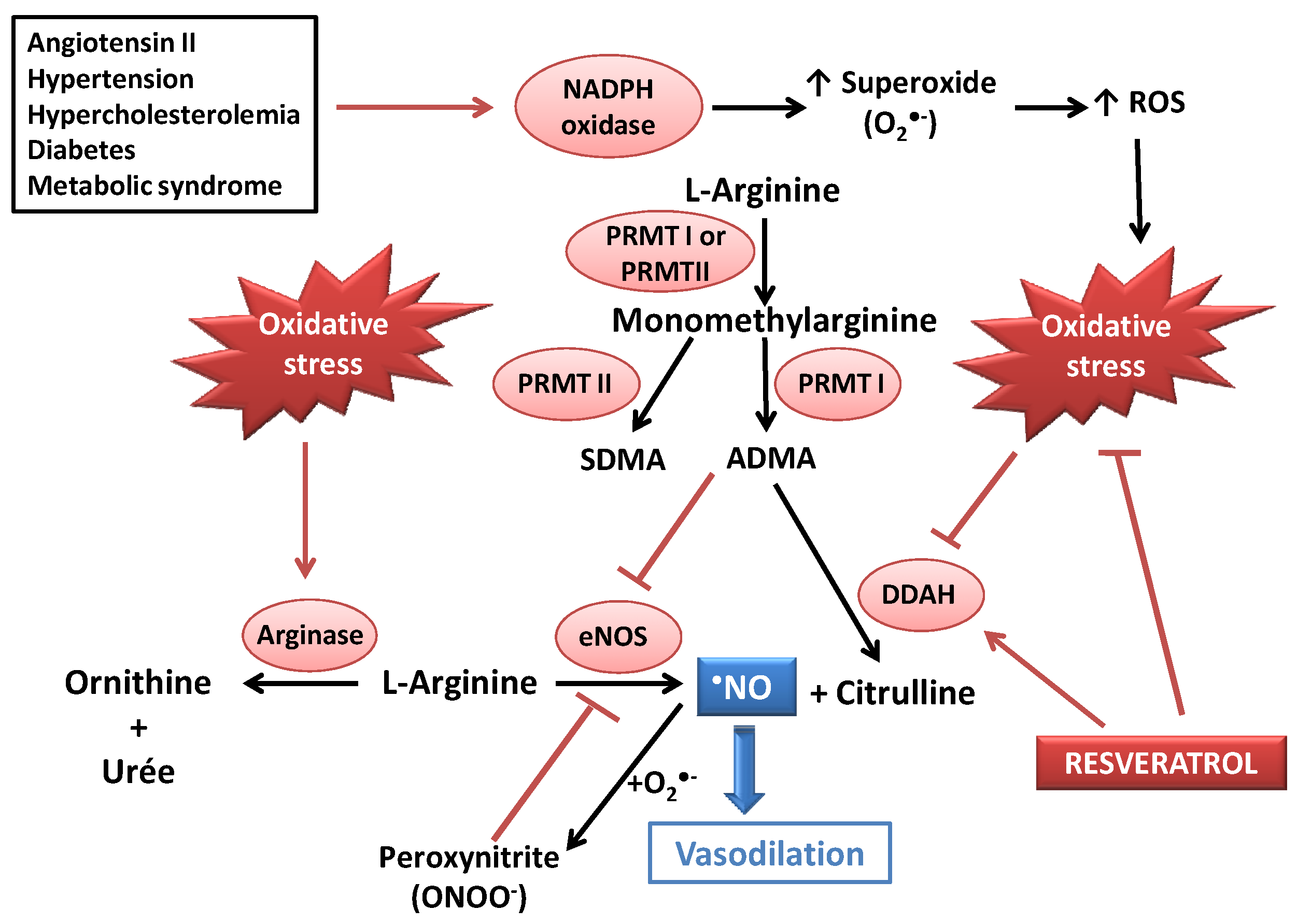
2. Resveratrol as an Antioxidant Able to Improve Nitric Oxide Availability in Vitro
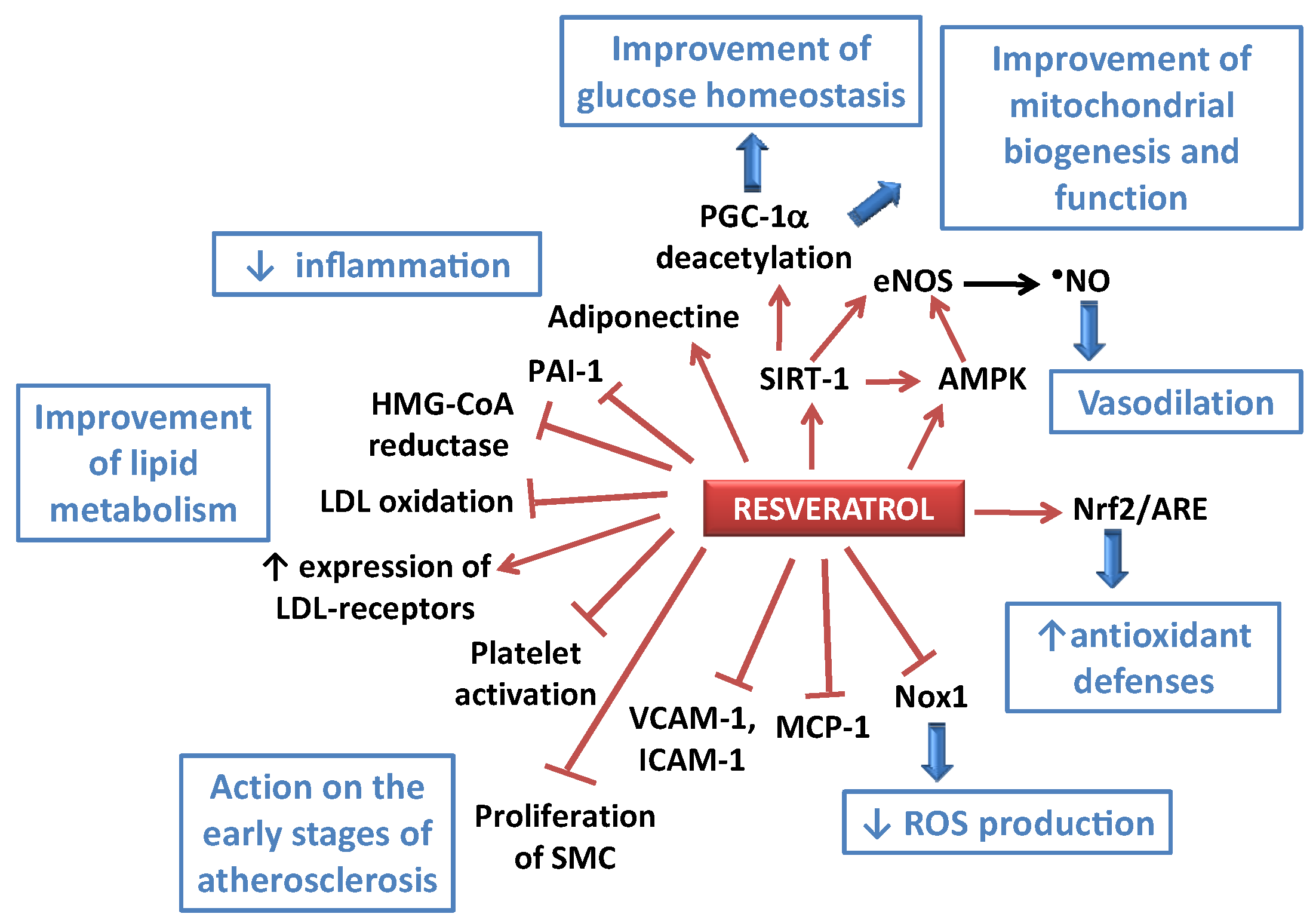
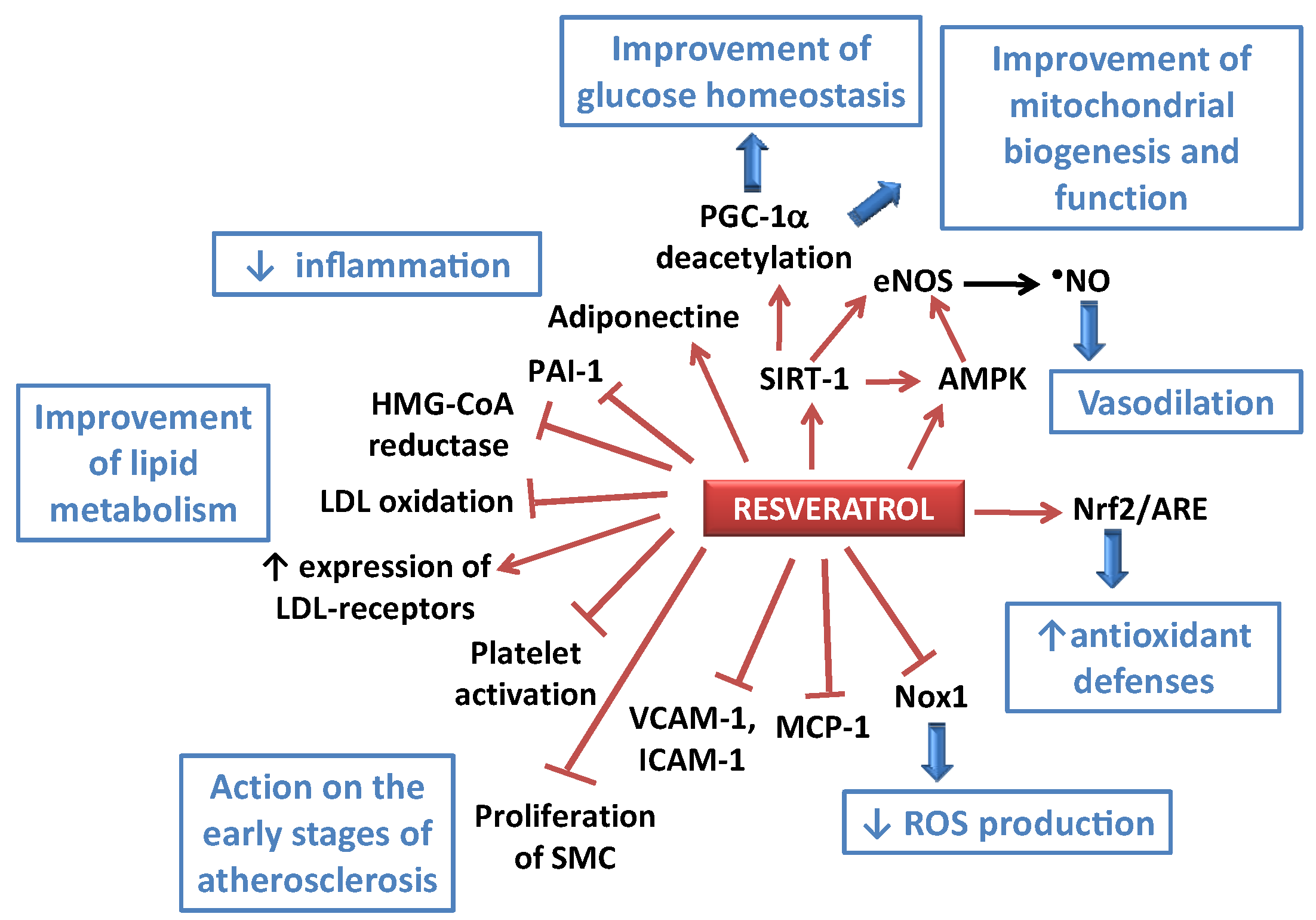
3. Health Benefits of Resveratrol against Cardiovascular Diseases: Examples of Preclinical and Clinical Studies
3.1. Anti-Atherosclerotic Effects of RES
3.2. Anti-Hypertensive Effects of RES
3.3. Protective Effect of RES in Stroke
3.4. Effects of RES on Myocardial Ischemia
3.5. Effects of RES on Heart Failure
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Laslett, L.J.; Alagona, P., Jr.; Clark, B.A., 3rd; Drozda, J.P., Jr.; Saldivar, F.; Wilson, S.R.; Poe, C.; Hart, M. The worldwide environment of cardiovascular disease: Prevalence, diagnosis, therapy, and policy issues: A report from the American College of Cardiology. J. Am. Coll. Cardiol. 2012, 60, S1–S49. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Smoliga, J.M.; Baur, J.A.; Hausenblas, H.A. Resveratrol and health—A comprehensive review of human clinical trials. Mol. Nutr. Food Res. 2011, 55, 1129–1141. [Google Scholar] [CrossRef] [PubMed]
- Delmas, D.; Jannin, B.; Latruffe, N. Resveratrol: Preventing properties against vascular alterations and ageing. Mol. Nutr. Food Res. 2005, 49, 377–395. [Google Scholar] [CrossRef] [PubMed]
- Zordoky, B.N.; Robertson, I.M.; Dyck, J.R. Preclinical and clinical evidence for the role of resveratrol in the treatment of cardiovascular diseases. Biochim. Biophys. Acta 2015, 1852, 1155–1177. [Google Scholar] [CrossRef] [PubMed]
- Cottart, C.H.; Nivet-Antoine, V.; Beaudeux, J.L. Review of recent data on the metabolism, biological effects, and toxicity of resveratrol in humans. Mol. Nutr. Food Res. 2014, 58, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Wallerath, T.; Deckert, G.; Ternes, T.; Anderson, H.; Li, H.; Witte, K.; Förstermann, U. Resveratrol, a polyphenolic phytoalexin present in red wine, enhances expression and activity of endothelial nitric oxide synthase. Circulation 2002, 106, 1652–1658. [Google Scholar] [CrossRef] [PubMed]
- Leikert, J.F.; Räthel, T.R.; Wohlfart, P.; Cheynier, V.; Vollmar, A.M.; Dirsch, V.M. Red wine polyphenols enhance endothelial nitric oxide synthase expression and subsequent nitric oxide release from endothelial cells. Circulation 2002, 106, 1614–1617. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.I.; Abdelkhalek, T.M.; Saleh, M.M.; Kamel, M.A.; Youssef, M.; Tawfik, S.H.; Dominguez, H. Insights into the molecular mechanisms of diabetes-induced endothelial dysfunction: Focus on oxidative stress and endothelial progenitor cells. Endocrine 2015, 50, 537–567. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.Y.; Ito, A.; Asagami, T.; Tsao, P.S.; Adimoolam, S.; Kimoto, M.; Tsuji, H.; Reaven, G.M.; Cooke, J.P. Impaired nitric oxide synthase pathway in diabetes mellitus: Role of asymmetric dimethylarginine and dimethylarginine dimethylaminohydrolase. Circulation 2002, 106, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Förstermann, U. Nitric oxide in the pathogenesis of vascular disease. J. Pathol. 2000, 190, 244–254. [Google Scholar] [CrossRef]
- Camont, L.; Collin, F.; Marchetti, C.; Jore, D.; Gardes-Albert, M.; Bonnefont-Rousselot, D. Liquid chromatographic/electrospray ionization mass spectrometric identification of the oxidation end-products of trans-resveratrol in aqueous solutions. Rapid Commun. Mass Spectrom. 2010, 24, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Camont, L.; Collin, F.; Couturier, M.; Thérond, P.; Jore, D.; Gardès-Albert, M.; Bonnefont-Rousselot, D. Radical-induced oxidation of trans-resveratrol. Biochimie 2012, 94, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Rhayem, Y.; Thérond, P.; Camont, L.; Couturier, M.; Beaudeux, J.L.; Legrand, A.; Jore, D.; Gardés-Albert, M.; Bonnefont-Rousselot, D. Chain-breaking activity of resveratrol and piceatannol in a linoleate micellar model. Chem. Phys. Lipids 2008, 155, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Scalera, F.; Kielstein, J.T.; Martens-Lobenhoffer, J.; Postel, S.C.; Täger, M.; Bode-Böger, S.M. Erythropoietin increases asymmetric dimethylarginine in endothelial cells: Role of dimethylarginine dimethylaminohydrolase. J. Am. Soc. Nephrol. 2005, 16, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Frombaum, M.; le Clanche, S.; Bonnefont-Rousselot, D.; Borderie, D. Antioxidant effects of resveratrol and other stilbene derivatives on oxidative stress and •NO bioavailability: Potential benefits to cardiovascular diseases. Biochimie 2012, 94, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Frombaum, M.; Thérond, P.; Djelidi, R.; Beaudeux, J.L.; Bonnefont-Rousselot, D.; Borderie, D. Piceatannol is more effective than resveratrol in restoring endothelial cell dimethylargininedimethylaminohydrolase expression and activity after high-glucose oxidative stress. Free Radic. Res. 2011, 45, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Bivalacqua, T.J.; Hellstrom, W.J.; Kadowitz, P.J.; Champion, H.C. Increased expression of arginase II in human diabetic corpus cavernosum: In diabetic-associated erectile dysfunction. Biochem. Biophys. Res. Commun. 2001, 283, 923–927. [Google Scholar] [CrossRef] [PubMed]
- White, A.R.; Ryoo, S.; Li, D.; Champion, H.C.; Steppan, J.; Wang, D.; Nyhan, D.; Shoukas, A.A.; Hare, J.M.; Berkowitz, D.E. Knockdown of arginase I restores NO signaling in the vasculature of old rats. Hypertension 2006, 47, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Fast, W. Inhibition of human dimethylarginine dimethylaminohydrolase-1 by S-nitroso-l-homocysteine and hydrogen peroxide. Analysis, quantification, and implications for hyperhomocysteinemia. J. Biol. Chem. 2007, 282, 34684–34692. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Peng, J.; Liu, S.Y.; Wang, C.J.; Xiang, D.X.; Xiong, X.M.; Hu, C.P.; Li, Y.J. Inhibitory effect of resveratrol derivative BTM-0512 on high glucose-induced cell senescence involves dimethylaminohydrolase/asymmetric dimethylarginine pathway. Clin. Exp. Pharmacol. Physiol. 2010, 37, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Dolinsky, V.W.; Dyck, J.R. Calorie restriction and resveratrol in cardiovascular health and disease. Biochim. Biophys. Acta 2011, 1812, 1477–1489. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xia, N.; Förstermann, U. Cardiovascular effects and molecular targets of resveratrol. Nitric Oxide 2012, 26, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Fichtlscherer, S.; Zeiher, A.M.; Dimmeler, S. Circulating microRNAs: Biomarkers or mediators of cardiovascular diseases? Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2383–2390. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Pacher, P.; Das, D.K. MicroRNA signatures of resveratrol in the ischemic heart. Ann. N. Y. Acad. Sci. 2011, 1215, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Larrosa, M.; González-Sarrías, A.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Resveratrol and clinical trials: The crossroad from in vitro studies to human evidence. Curr. Pharm. Des. 2013, 19, 6064–6093. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Witztum, J.L. Atherosclerosis. The road ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef]
- Göçmen, A.Y.; Burgucu, D.; Gümüşlü, S. Effect of resveratrol on platelet activation in hypercholesterolemic rats: CD40-CD40L system as a potential target. Appl. Physiol. Nutr. Metab. 2011, 36, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.J.; Ahn, J.Y.; Kim, S.; Choi, M.S.; Ha, T.Y. Resveratrol attenuates the expression of HMG-CoA reductase mRNA in hamsters. Biochem. Biophys. Res. Commun. 2008, 367, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Yashiro, T.; Nanmoku, M.; Shimizu, M.; Inoue, J.; Sato, R. Resveratrol increases the expression and activity of the low density lipoprotein receptor in hepatocytes by the proteolytic activation of the sterol regulatory element-binding proteins. Atherosclerosis 2012, 220, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Witztum, J.L.; Steinberg, D. Role of oxidized low density lipoprotein in atherogenesis. J. Clin. Investig. 1991, 88, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Berrougui, H.; Grenier, G.; Loued, S.; Drouin, G.; Khalil, A. A new insight into resveratrol as an atheroprotective compound: Inhibition of lipid peroxidation and enhancement of cholesterol efflux. Atherosclerosis 2009, 207, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Ramprasath, V.R.; Jones, P.J. Anti-atherogenic effects of resveratrol. Eur. J. Clin. Nutr. 2010, 64, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chen, L.H.; Varadharajan, T.; Tsai, M.J.; Chia, Y.C.; Yuan, T.C.; Sung, P.J.; Weng, C.F. Resveratrol inhibits glucose-induced migration of vascular smooth muscle cells mediated by focal adhesion kinase. Mol. Nutr. Food Res. 2014, 58, 1389–1401. [Google Scholar] [CrossRef] [PubMed]
- Haskó, G.; Pacher, P. Endothelial Nrf2 activation: A new target for resveratrol? Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H10–H12. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.H.; Alex, D.; Huang, H.Q.; Wang, N.; Yu, N.; Wang, Y.T.; Leung, G.P.; Lee, S.M. Inhibition of TNF-α-mediated endothelial cell-monocyte cell adhesion and adhesion molecules expression by the resveratrol derivative, trans-3,5,4′-trimethoxystilbene. Phytother. Res. 2011, 25, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Park, D.W.; Baek, K.; Kim, J.R.; Lee, J.J.; Ryu, S.H.; Chin, B.R.; Baek, S.H. Resveratrol inhibits foam cell formation via NADPH oxidase 1- mediated reactive oxygen species and monocyte chemotactic protein-1. Exp. Mol. Med. 2009, 41, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Latruffe, N.; Lançon, A.; Frazzi, R.; Aires, V.; Delmas, D.; Michaille, J.J.; Djouadi, F.; Bastin, J.; Cherkaoui-Malki, M. Exploring new ways of regulation by resveratrol involving miRNAs, with emphasis on inflammation. Ann. N. Y. Acad. Sci. 2015, 1348, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Hao, E.; Lang, F.; Chen, Y.; Zhang, H.; Cong, X.; Shen, X.; Su, G. Resveratrol alleviates endotoxin-induced myocardial toxicity via the Nrf2 transcription factor. PLoS ONE 2013, 8, e69452. [Google Scholar] [CrossRef] [PubMed]
- El-Mowafy, A.M.; Alkhalaf, M.; El-Kashef, H.A. Resveratrol reverses hydrogen peroxide-induced proliferative effects in human coronary smooth muscle cells: A novel signaling mechanism. Arch. Med. Res. 2008, 39, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A. Effects of resveratrol supplementation on plasma lipids: A systematic review and meta-analysis of randomized controlled trials. Nutr. Rev. 2013, 71, 822–835. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, J.K.; Thomas, S.; Nanjan, M.J. Resveratrol supplementation improves glycemic control in type 2 diabetes mellitus. Nutr. Res. 2012, 32, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Bo, S.; Ciccone, G.; Castiglione, A.; Gambino, R.; de Michieli, F.; Villois, P.; Durazzo, M.; Cavallo-Perin, P.; Cassader, M. Anti-inflammatory and antioxidant effects of resveratrol in healthy smokers a randomized, double-blind, placebo-controlled, cross-over trial. Curr. Med. Chem. 2013, 20, 1323–1231. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; García-Almagro, F.J.; Avilés-Plaza, F.; Parra, S.; Yáñez-Gascón, M.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; et al. Consumption of a grape extract supplement containing resveratrol decreases oxidized LDL and ApoB in patients undergoing primary prevention of cardiovascular disease: A triple-blind, 6-month follow-up, placebo-controlled, randomized trial. Mol. Nutr. Food Res. 2012, 56, 810–821. [Google Scholar] [CrossRef] [PubMed]
- Smulyan, H.; Mookherjee, S.; Safar, M.E. The two faces of hypertension: Role of aortic stiffness. J. Am. Soc. Hypertens. 2016, 10, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Rivera, L.; Morón, R.; Zarzuelo, A.; Galisteo, M. Long-term resveratrol administration reduces metabolic disturbances and lowers blood pressure in obese Zucker rats. Biochem. Pharmacol. 2009, 77, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Dolinsky, V.W.; Chakrabarti, S.; Pereira, T.J.; Oka, T.; Levasseur, J.; Beker, D.; Zordoky, B.N.; Morton, J.S.; Nagendran, J.; Lopaschuk, G.D.; et al. Resveratrol prevents hypertension and cardiac hypertrophy in hypertensive rats and mice. Biochim. Biophys. Acta 2013, 1832, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- Chan, V.; Fenning, A.; Iyer, A.; Hoey, A.; Brown, L. Resveratrol improves cardiovascular function in DOCA-salt hypertensive rats. Curr. Pharm. Biotechnol. 2011, 12, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Rimbaud, S.; Ruiz, M.; Piquereau, J.; Mateo, P.; Fortin, D.; Veksler, V.; Garnier, A.; Ventura-Clapier, R. Resveratrol improves survival, hemodynamics and energetics in a rat model of hypertension leading to heart failure. PLoS ONE 2011, 6, e26391. [Google Scholar] [CrossRef] [PubMed]
- Thandapilly, S.J.; Louis, X.L.; Behbahani, J.; Movahed, A.; Yu, L.; Fandrich, R.; Zhang, S.; Kardami, E.; Anderson, H.D.; Netticadan, T. Reduced hemodynamic load aids low-dose resveratrol in reversing cardiovascular defects in hypertensive rats. Hypertens. Res. 2013, 36, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Gordish, K.L.; Beierwaltes, W.H. Resveratrol induces acute endothelium-dependent renal vasodilation mediated through nitric oxide and reactive oxygen species scavenging. Am. J. Physiol. Ren. Physiol. 2014, 306, F542–F550. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, G.; Yao, H.; Sundar, I.K.; Caito, S.; Rahman, I. SIRT1 regulates oxidant- and cigarette smoke-induced eNOS acetylation in endothelial cells: Role of resveratrol. Biochem. Biophys. Res. Commun. 2010, 393, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Soylemez, S.; Sepici, A.; Akar, F. Resveratrol supplementation gender independently improves endothelial reactivity and suppresses superoxide production in healthy rats. Cardiovasc. Drugs Ther. 2009, 23, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Rush, J.W.; Quadrilatero, J.; Levy, A.S.; Ford, R.J. Chronic resveratrol enhances endothelium-dependent relaxation but does not alter eNOS levels in aorta of spontaneously hypertensive rats. Exp. Biol. Med. (Maywood) 2007, 232, 814–822. [Google Scholar] [PubMed]
- Cao, X.; Luo, T.; Luo, X.; Tang, Z. Resveratrol prevents AngII-induced hypertension via AMPK activation and RhoA/ROCK suppression in mice. Hypertens. Res. 2014, 37, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, W.; Zhang, P.; He, S.; Huang, D. Effect of resveratrol on blood pressure: A meta-analysis of randomized controlled trials. Clin. Nutr. 2015, 34, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Carrizzo, A.; Puca, A.; Damato, A.; Marino, M.; Franco, E.; Pompeo, F.; Traficante, A.; Civitillo, F.; Santini, L.; Trimarco, V.; et al. Resveratrol improves vascular function in patients with hypertension and dyslipidemia by modulating NO metabolism. Hypertension 2013, 62, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.; Tuor, U.I.; Thompson, R.; Institoris, A.; Kulynych, A.; Zhang, X.; Kinniburgh, D.W.; Bari, F.; Busija, D.W.; Barber, P.A. Protection against recurrent stroke with resveratrol: Endothelial protection. PLoS ONE 2012, 7, e47792. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Tsai, M.C.; Chih, C.L.; Hung, L.M.; Tsai, S.K. Resveratrol reduction of infarct size in Long-Evans rats subjected to focal cerebral ischemia. Life Sci. 2001, 69, 1057–1065. [Google Scholar] [CrossRef]
- Arrick, D.M.; Sun, H.; Patel, K.P.; Mayhan, W.G. Chronic resveratrol treatment restores vascular responsiveness of cerebral arterioles in type 1 diabetic rats. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H696–H703. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Agrawal, M.; Doré, S. Neuroprotective properties and mechanisms of resveratrol in in vitro and in vivo experimental cerebral stroke models. ACS Chem. Neurosci. 2013, 4, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Zhou, Y.; Wang, K.; Hou, Y.; Hou, R.; Ye, X. Resveratrol provides neuroprotection by inhibiting phosphodiesterases and regulating the cAMP/AMPK/SIRT1 pathway after stroke in rats. Brain Res. Bull. 2016, 121, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Wightman, E.L.; Reay, J.L.; Lietz, G.; Okello, E.J.; Wilde, A.; Haskell, C.F. Effects of resveratrol on cerebral blood flow variables and cognitive performance in humans: A double-blind, placebo-controlled, crossover investigation. Am. J. Clin. Nutr. 2010, 91, 1590–1597. [Google Scholar] [CrossRef] [PubMed]
- Wightman, E.L.; Reay, J.L.; Haskell, C.F.; Williamson, G.; Dew, T.P.; Kennedy, D.O. Effects of resveratrol alone or in combination with piperine on cerebral blood flow parameters and cognitive performance in human subjects: A randomised, double-blind, placebo-controlled, cross-over investigation. Br. J. Nutr. 2014, 112, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.M.; Howe, P.R.; Wong, R.H. Clinical evaluation of effects of chronic resveratrol supplementation on cerebrovascular function, cognition, mood, physical function and general well-being in postmenopausal women-rationale and study design. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Raj, P.; Louis, X.L.; Thandapilly, S.J.; Movahed, A.; Zieroth, S.; Netticadan, T. Potential of resveratrol in the treatment of heart failure. Life Sci. 2014, 95, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.M.; Su, M.J.; Chen, J.K. Resveratrol protects myocardial ischemia-reperfusion injury through both NO-dependent and NO-independent mechanisms. Free Radic. Biol. Med. 2004, 36, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Jia, G.L.; Wang, Y.M.; Ma, H. Cardioprotective effect of resvaratrol pretreatment on myocardial ischemia-reperfusion induced injury in rats. Vasc. Pharmacol. 2006, 45, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Shalwala, M.; Zhu, S.G.; Das, A.; Salloum, F.N.; Xi, L.; Kukreja, R.C. Sirtuin 1 (SIRT1) activation mediates sildenafil induced delayed cardioprotection against ischemia-reperfusion injury in mice. PLoS ONE 2014, 9, e86977. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Cordis, G.A.; Maulik, N.; Das, D.K. Pharmacological preconditioning with resveratrol: Role of CREB-dependent Bcl-2 signaling via adenosine A3 receptor activation. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H328–H335. [Google Scholar] [CrossRef] [PubMed]
- Kaga, S.; Zhan, L.; Matsumoto, M.; Maulik, N. Resveratrol enhances neovascularization in the infarcted rat myocardium through the induction of thioredoxin-1, heme oxygenase-1 and vascular endothelial growth factor. J. Mol. Cell. Cardiol. 2005, 39, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Kaga, S.; Zhan, L.; Bagchi, D.; Das, D.K.; Bertelli, A.; Maulik, N. Resveratrol ameliorates myocardial damage by inducing vascular endothelial growth factor-angiogenesis and tyrosine kinase receptor Flk-1. Cell Biochem. Biophys. 2006, 44, 43–49. [Google Scholar] [CrossRef]
- Chen, Y.R.; Yi, F.F.; Li, X.Y.; Wang, C.Y.; Chen, L.; Yang, X.C.; Su, P.X.; Cai, J. Resveratrol attenuates ventricular arrhythmias and improves the long-term survival in rats with myocardial infarction. Cardiovasc. Drugs Ther. 2008, 22, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Robich, M.P.; Osipov, R.M.; Chu, L.M.; Han, Y.; Feng, J.; Nezafat, R.; Clements, R.T.; Manning, W.J.; Sellke, F.W. Resveratrol modifies risk factors for coronary artery disease in swine with metabolic syndrome and myocardial ischemia. Eur. J. Pharmacol. 2011, 664, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Gurusamy, N.; Lekli, I.; Mukherjee, S.; Ray, D.; Ahsan, M.K.; Gherghiceanu, M.; Popescu, L.M.; Das, D.K. Cardioprotection by resveratrol: A novel mechanism via autophagy involving the mTORC2 pathway. Cardiovasc. Res. 2010, 86, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Wu, B.; Chen, C.; Chen, B.; Zhang, W.; Xu, D.; Bin, J.; Liao, Y. Resveratrol improves myocardial ischemia and ischemic heart failure in mice by antagonizing the detrimental effects of fractalkine. Crit. Care Med. 2012, 40, 3026–3033. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.S.; Wang, Z.B.; Ye, Z.; Lei, J.P.; Li, L.; Su, D.F.; Zheng, X. Resveratrol, an activator of SIRT1, upregulates AMPK and improves cardiac function in heart failure. Genet. Mol. Res. 2014, 13, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, H.; Takemura, G.; Goto, K.; Tsujimoto, A.; Ogino, A.; Takeyama, T.; Kawaguchi, T.; Watanabe, T.; Morishita, K.; Kawasaki, M.; et al. Resveratrol reverses remodeling in hearts with large, old myocardial infarctions through enhanced autophagy-activating AMP kinase pathway. Am. J. Pathol. 2013, 182, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Sabe, A.A.; Elmadhun, N.Y.; Dalal, R.S.; Robich, M.P.; Sellke, F.W. Resveratrol regulates autophagy signaling in chronically ischemic myocardium. J. Thorac. Cardiovasc. Surg. 2014, 147, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Wu, R.X.; Zhao, L.; Li, J.; Guo, H.T.; Fan, R.; Cui, Y.; Wang, Y.M.; Yue, S.Q.; Pei, J.M. Resveratrol attenuates ischemia/reperfusion injury in neonatal cardiomyocytes and its underlying mechanism. PLoS ONE 2012, 7, e51223. [Google Scholar] [CrossRef] [PubMed]
- Sabe, A.A.; Sadek, A.A.; Elmadhun, N.Y.; Dalal, R.S.; Robich, M.P.; Bianchi, C.; Sellke, F.W. Investigating the effects of resveratrol on chronically ischemic myocardium in a swine model of metabolic syndrome: A proteomics analysis. J. Med. Food 2015, 18, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Yang, R.; Yang, J.; Yang, J.; Ding, J.; Wu, H.; Zhang, J. Resveratrol pretreatment protects rat hearts from ischemia/reperfusion injury partly via a NALP3 inflammasome pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 8731–8741. [Google Scholar] [PubMed]
- Mukhopadhyay, P.; Mukherjee, S.; Ahsan, K.; Bagchi, A.; Pacher, P.; Das, D.K. Restoration of altered microRNA expression in the ischemic heart with resveratrol. PLoS ONE 2010, 5, e15705. [Google Scholar] [CrossRef] [PubMed]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signaling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Khanna, S.; Hussain, S.R.; Biswas, S.; Azad, A.; Rink, C.; Gnyawali, S.; Shilo, S.; Nuovo, G.J.; Sen, C.K. MicroRNA expression in response to murine myocardial infarction: miR-21 regulates fibroblast metalloprotease-2 via phosphatase and tensin homologue. Cardiovasc. Res. 2009, 82, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Cascio, S.; D’Andrea, A.; Ferla, R.; Surmacz, E.; Gulotta, E.; Amodeo, V.; Bazan, V.; Gebbia, N.; Russo, A. miR-20b modulates VEGF expression by targeting HIF-1 alpha and STAT3 in MCF-7 breast cancer cells. J. Cell. Physiol. 2010, 224, 242–249. [Google Scholar] [PubMed]
- Guttilla, I.K.; White, B.A. Coordinate regulation of FOXO1 by miR-27a, miR-96, and miR-182 in breast cancer cells. J. Biol. Chem. 2009, 284, 23204–23216. [Google Scholar] [CrossRef] [PubMed]
- Saunders, L.R.; Sharma, A.D.; Tawney, J.; Nakagawa, M.; Okita, K.; Yamanaka, S.; Willenbring, H.; Verdin, E. miRNAs regulate SIRT1 expression during mouse embryonic stem cell differentiation and in adult mouse tissues. Aging (Albany NY) 2010, 2, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of microRNAs after myocardial infarction reveals a role of miR-29 in cardiac fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Zhao, L.; Feng, D.; Ma, W.; Liu, Y.; Wang, Y.; Liang, J.; Yang, F.; Bi, C.; Chen, H.; et al. Resveratrol attenuated low ambient temperature-induced myocardial hypertrophy via inhibiting cardiomyocyte apoptosis. Cell. Physiol. Biochem. 2015, 35, 2451–2462. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Grape resveratrol increases serum adiponectin and downregulates inflammatory genes in peripheral blood mononuclear cells: A triple-blind, placebo-controlled, one-year clinical trial in patients with stable coronary artery disease. Cardiovasc. Drugs Ther. 2013, 27, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Magyar, K.; Halmosi, R.; Palfi, A.; Feher, G.; Czopf, L.; Fulop, A.; Battyany, I.; Sumegi, B.; Toth, K.; Szabados, E. Cardioprotection by resveratrol: A human clinical trial in patients with stable coronary artery disease. Clin. Hemorheol. Microcirc. 2012, 50, 179–187. [Google Scholar] [PubMed]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Militaru, C.; Donoiu, I.; Craciun, A.; Scorei, I.D.; Bulearca, A.M.; Scorei, R.I. Oral resveratrol and calcium fructoborate supplementation in subjects with stable angina pectoris: Effects on lipid profiles, inflammation markers, and quality of life. Nutrition 2013, 29, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; DiPette, D.J.; Supowit, S.C. Protective effect of resveratrol against pressure overload-induced heart failure. Food Sci. Nutr. 2014, 2, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Tanno, M.; Kuno, A.; Yano, T.; Miura, T.; Hisahara, S.; Ishikawa, S.; Shimamoto, K.; Horio, Y. Induction of manganese superoxide dismutase by nuclear translocation and activation of SIRT1 promotes cell survival in chronic heart failure. J. Biol. Chem. 2010, 285, 8375–8382. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.Y.; Dolinsky, V.W.; Soltys, C.L.; Viollet, B.; Baksh, S.; Light, P.E.; Dyck, J.R. Resveratrol inhibits cardiac hypertrophy via AMP-activated protein kinase and Akt. J. Biol. Chem. 2008, 283, 24194–24201. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, M.; Matta, M.J.; Sunderesan, N.R.; Gupta, M.P.; Periasamy, M.; Gupta, M. Resveratrol, an activator of SIRT1, upregulates sarcoplasmic calcium ATPase and improves cardiac function in diabetic cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H833–H843. [Google Scholar] [CrossRef] [PubMed]
- Dolinsky, V.W.; Soltys, C.L.; Rogan, K.J.; Chan, A.Y.; Nagendran, J.; Wang, S.; Dyck, J.R. Resveratrol prevents pathological but not physiological cardiac hypertrophy. J. Mol. Med. (Berl.) 2015, 93, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Azorín-Ortuño, M.; Yáñez-Gascón, M.J.; Vallejo, F.; Pallarés, F.J.; Larrosa, M.; Lucas, R.; Morales, J.C.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Metabolites and tissue distribution of resveratrol in the pig. Mol. Nutr. Food Res. 2011, 55, 1154–1168. [Google Scholar] [CrossRef] [PubMed]
- Bresciani, L.; Calani, L.; Bocchi, L.; Delucchi, F.; Savi, M.; Ray, S.; Brighenti, F.; Stilli, D.; del Rio, D. Bioaccumulation of resveratrol metabolites in myocardial tissue is dose-time dependent and related to cardiac hemodynamics in diabetic rats. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Brantley, S.J.; Argikar, A.A.; Lin, Y.S.; Nagar, S.; Paine, M.F. Herb-drug interactions: Challenges and opportunities for improved predictions. Drug Metab. Dispos. 2014, 42, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Cottart, C.H.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J.L. Resveratrol bioavailability and toxicity in humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, D.M.; Yan, J.; Soleas, G.J. Absorption of three wine-related polyphenols in three different matrices by healthy subjects. Clin. Biochem. 2003, 36, 79–87. [Google Scholar] [CrossRef]
- Calamini, B.; Ratia, K.; Malkowski, M.G.; Cuendet, M.; Pezzuto, J.M.; Santarsiero, B.D.; Mesecar, A.D. Pleiotropic mechanisms facilitated by resveratrol and its metabolites. Biochem. J. 2010, 429, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Boocock, D.J.; Faust, G.E.; Patel, K.R.; Schinas, A.M.; Brown, V.A.; Ducharme, M.P.; Booth, T.D.; Crowell, J.A.; Perloff, M.; Gescher, A.J.; et al. Phase I dose escalation pharmacokinetic study in healthy volunteers of resveratrol, a potential cancer chemopreventive agent. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Ng, K.Y. Impact of glutaraldehyde on in vivo colon-specific release of resveratrol from biodegradable pectin-based formulation. J. Pharm. Sci. 2010, 99, 4903–4916. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Ji, C.; Xu, H.; Li, X.; Ding, H.; Ye, M.; Zhu, Z.; Ding, D.; Jiang, X.; Ding, X.; et al. Resveratrol-loaded polymeric micelles protect cells from Abeta-induced oxidative stress. Int. J. Pharm. 2009, 375, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Amri, A.; le Clanche, S.; Thérond, P.; Bonnefont-Rousselot, D.; Borderie, D.; Lai-Kuen, R.; Chaumeil, J.C.; Sfar, S.; Charrueau, C. Resveratrol self-emulsifying system increases the uptake by endothelial cells and improves protection against oxidative stress-mediated death. Eur. J. Pharm. Biopharm. 2014, 86, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Martins, S.; Segundo, M.A.; Reis, S. Nanoscale delivery of resveratrol towards enhancement of supplements and nutraceuticals. Nutrients 2016, 8. [Google Scholar] [CrossRef]
- Diaz-Gerevini, G.T.; Repossi, G.; Dain, A.; Tarres, M.C.; Das, U.N.; Eynard, A.R. Beneficial action of resveratrol: How and why? Nutrition 2016, 32, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Penalva, R.; Esparza, I.; Larraneta, E.; González-Navarro, C.J.; Gamazo, C.; Irache, J.M. Zein-based nanoparticles improve the oral bioavailability of resveratrol and its anti-inflammatory effects in a mouse model of endotoxic shock. J. Agric. Food Chem. 2015, 63, 5603–5611. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha Lindner, G.; Santos, D.B.; Colle, D.; Moreira, E.L.G.; Prediger, R.D.; Farina, M.; Khalil, N.M.; Mainardes, R.M. Improved neuroprotective effects of resveratrol-loadedpolysorbate 80-coated poly (lactide) nanoparticles in MPTP-induced parkinsonism. Nanomedicine (Lond.) 2015, 10, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Gepner, Y.; Golan, R.; Harman-Boehm, I.; Henkin, Y.; Schwarzfuchs, D.; Shelef, I.; Durst, R.; Kovsan, J.; Bolotin, A.; Leitersdorf, E.; et al. Effects of initiating moderate alcohol intake on cardiometabolic risk in adults with type 2 diabetes: A 2-year randomized, controlled trial. Ann. Intern. Med. 2015, 163, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, B.; Mukherjee, S.; Das, D.K. Hormetic response of resveratrol against cardioprotection. Exp. Clin. Cardiol. 2010, 15, e134–e138. [Google Scholar] [PubMed]
- Kucinska, M.; Piotrowska, H.; Luczak, M.W.; Mikula-Pietrasik, J.; Ksiazek, K.; Wozniak, M.; Wierzchowski, M.; Dudka, J.; Jäger, W.; Murias, M. Effects of hydroxylated resveratrol analogs on oxidative stress and cancer cells death in human acute T cell leukemia cell line: Prooxidativepotential of hydroxylated resveratrol analogs. Chem. Biol. Interact. 2014, 209, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.K.; Chen, J.; Lee, W.B. Differential inhibition and inactivation of human CYP1 enzymes by trans-resveratrol: Evidence for mechanism-based inactivation of CYP1A2. J. Pharmacol. Exp. Ther. 2001, 299, 874–882. [Google Scholar] [PubMed]
- Chow, H.H.; Garland, L.L.; Hsu, C.H.; Vining, D.R.; Chew, W.M.; Miller, J.A.; Perloff, M.; Crowell, J.A.; Alberts, D.S. Resveratrol modulates drug- and carcinogen-metabolizing enzymes in a healthy volunteer study. Cancer Prev. Res. (Phila) 2010, 3, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Iwuchukwu, O.F.; Tallarida, R.J.; Nagar, S. Resveratrol in combination with other dietary polyphenols concomitantly enhances antiproliferation and UGT1A1 induction in Caco-2 cells. Life Sci. 2011, 88, 1047–1054. [Google Scholar] [CrossRef] [PubMed]
- Detampel, P.; Beck, M.; Krähenbühl, S.; Huwyler, J. Drug interaction potential of resveratrol. Drug Metab. Rev. 2012, 44, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Shin, Y.G.; Kosmeder, J.W.; Pezzuto, J.M.; van Breemen, R.B. Liquid chromatography/tandemmass spectrometric determination of inhibition of human cytochrome P450 isozymes by resveratrol and resveratrol-3-sulfate. Rapid Commun. Mass Spectrom. 2003, 17, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Ogu, C.C.; Maxa, J.L. Drug interactions due to cytochrome P450. Proceeding (Bayl. Univ. Med. Cent.) 2000, 13, 421–423. [Google Scholar]
- Visioli, F. The resveratrol fiasco. Pharmacol. Res. 2014, 90, 87. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Larrosa, M.; Yáñez-Gascón, M.J.; Dávalos, A.; Gil-Zamorano, J.; Gonzálvez, M.; García-Almagro, F.J.; Ros, J.A.R.; Tomás-Barberán, F.A.; Espín, J.C.; et al. One-year supplementation with a grape extract containing resveratrol modulates inflammatory-related microRNAs and cytokines expression in peripheral blood mononuclear cells of type 2 diabetes and hypertensive patients with coronary artery disease. Pharmacol. Res. 2013, 72, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Tang, P.C.-T.; Ng, Y.-F.; Ho, S.; Gyda, M.; Chan, S.-W. Resveratrol and cardiovascular health—Promising therapeutic or hopeless illusion? Pharmacol. Res. 2014, 90, 88–115. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.N.; Luksha, L.; Logman, H.; Poston, L.; Agewall, S.; Kublickiene, K. Acute responses to phytoestrogens in small arteries from men with coronary heart disease. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1969–H1975. [Google Scholar] [CrossRef] [PubMed]
- Rakici, O.; Kiziltepe, U.; Coskun, B.; Aslamaci, S.; Akar, F. Effects of resveratrol on vascular tone and endothelial function of human saphenous vein and internal mammary artery. Int. J. Cardiol. 2005, 105, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Amadio, P.; Baldassarre, D.; Tarantino, E.; Zacchi, E.; Gianellini, S.; Squellerio, I.; Amato, M.; Weksler, B.B.; Tremoli, E.; Barbieri, S.S. Production of prostaglandin E2 induced by cigarette smoke modulates tissue factor expression and activity in endothelial cells. FASEB J. 2015, 29, 4001–4010. [Google Scholar] [CrossRef] [PubMed]
- Thushara, R.M.; Hemshekhar, M.; Paul, M.; Sundaram, M.S.; Shankar, R.L.; Kemparaju, K.; Girish, K.S. Crocin prevents sesamol-induced oxidative stress and apoptosis in human platelets. J. Thromb. Thrombolysis 2014, 38, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Urpí-Sardà, M.; Lamuela-Raventós, R.M.; Estruch, R.; Vázquez-Agell, M.; Serrano-Martínez, M.; Jaeger, W.; Andres-Lacueva, C. Diagnostic performance of urinary resveratrol metabolites as a biomarker of moderate wine consumption. Clin. Chem. 2006, 52, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Faghihzadeh, F.; Adibi, P.; Hekmatdoost, A. The effects of resveratrol supplementation on cardiovascular risk factors in patients with non-alcoholic fatty liver disease: Arandomised, double-blind, placebo-controlled study. Br. J. Nutr. 2015, 114, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Van der Made, S.M.; Plat, J.; Mensink, R.P. Resveratrol does not influence metabolic risk markers related to cardiovascular health in overweight and slightly obese subjects: A randomized, placebo-controlled crossover trial. PLoS ONE 2015, 10, e0118393. [Google Scholar]
- Semba, R.D.; Ferrucci, L.; Bartali, B.; Urpí-Sarda, M.; Zamora-Ros, R.; Sun, K.; Cherubini, A.; Bandinelli, S.; Andres-Lacueva, C. Resveratrol levels and all-cause mortality in older community-dwelling adults. JAMA Intern. Med. 2014, 174, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Soare, A.; Weiss, E.P.; Holloszy, J.O.; Fontana, L. Multiple dietary supplements do not affect metabolic and cardio-vascular health. Aging (Albany NY) 2014, 6, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.P.; Huang, S.S.; Deng, J.Y.; Chang, C.C.; Day, Y.J.; Hung, L.M. Insulin and resveratrol act synergistically, preventing cardiac dysfunction in diabetes, but the advantage of resveratrol in diabetics with acute heart attack is antagonized by insulin. Free Radic. Biol. Med. 2010, 49, 1710–1721. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Morgan, B.; Potter, B.J.; Ma, L.; Dellsperger, K.C.; Ungvari, Z.; Zhang, C. Resveratrol improves left ventricular diastolic relaxation in type 2 diabetes by inhibiting oxidative/nitrative stress: In vivo demonstration with magnetic resonance imaging. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H985–H994. [Google Scholar] [CrossRef] [PubMed]
- Barger, J.L.; Kayo, T.; Vann, J.M.; Arias, E.B.; Wang, J.; Hacker, T.A.; Wang, Y.; Raederstorff, D.; Morrow, J.D.; Leeuwenburgh, C.; et al. A low dose of dietary resveratrol partially mimics caloric restriction and retards aging parameters in mice. PLoS ONE 2008, 3, e2264. [Google Scholar] [CrossRef]
- Dang, W. The controversial world of sirtuins. Drug Discov. Today Technol. 2014, 12, e9–e17. [Google Scholar] [CrossRef] [PubMed]
- Pacholec, M.; Bleasdale, J.E.; Chrunyk, B.; Cunningham, D.; Flynn, D.; Garofalo, R.S.; Griffith, D.; Griffor, M.; Loulakis, P.; Pabst, B.; et al. SRT1720, SRT2183, SRT1460, and resveratrol are not direct activators of SIRT1. J. Biol. Chem. 2010, 285, 8340–8351. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Marcos, P.J.; Auwerx, J. Regulation of PGC-1α, a nodal regulator of mitochondrial biogenesis. Am. J. Clin. Nutr. 2011, 93, 884S–890S. [Google Scholar] [CrossRef] [PubMed]
- Csiszar, A.; Labinskyy, N.; Pinto, J.T.; Ballabh, P.; Zhang, H.; Losonczy, G.; Pearson, K.; de Cabo, R.; Pacher, P.; Zhang, C.; et al. Resveratrol induces mitochondrial biogenesis in endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H13–H20. [Google Scholar] [CrossRef] [PubMed]
- Louis, X.L.; Thandapilly, S.J.; MohanKumar, S.K.; Yu, L.; Taylor, C.G.; Zahradka, P.; Netticadan, T. Treatment with low-dose resveratrol reverses cardiac impairment in obese prone but not in obese resistant rats. J. Nutr. Biochem. 2012, 23, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Siwik, D.A.; Luptak, I.; Hou, X.; Wang, L.; Higuchi, A.; Weisbrod, R.M.; Ouchi, N.; Tu, V.H.; Calamaras, T.D.; et al. The polyphenols resveratrol and S17834 prevent the structural and functional sequelae of diet-induced metabolic heart disease in mice. Circulation 2012, 125, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Su, H.C.; Hung, L.M.; Chen, J.K. Resveratrol, a red wine antioxidant, possesses an insulin-like effect in streptozotocin-induced diabetic rats. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1339–E1346. [Google Scholar] [CrossRef] [PubMed]
- Bagul, P.K.; Middela, H.; Matapally, S.; Padiya, R.; Bastia, T.; Madhusudana, K.; Reddy, B.R.; Chakravarty, S.; Banerjee, S.K. Attenuation of insulin resistance, metabolic syndrome and hepatic oxidative stress by resveratrol in fructose-fed rats. Pharmacol. Res. 2012, 66, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Turan, B.; Tuncay, E.; Vassort, G. Resveratrol and diabetic cardiac function: Focus on recent in vitro and in vivo studies. J. Bioenerg. Biomembr. 2012, 44, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhou, R.; Wang, B.; Mi, M.T. Effect of resveratrol on glucose control and insulin sensitivity: Ameta-analysis of 11 randomized controlled trials. Am. J. Clin. Nutr. 2014, 99, 1510–1519. [Google Scholar] [CrossRef] [PubMed]
- Haohao, Z.; Guijun, Q.; Juan, Z.; Wen, K.; Lulu, C. Resveratrol improves high-fat diet induced insulin resistance by rebalancing subsarcolemmal mitochondrial oxidation and antioxidantion. J. Physiol. Biochem. 2015, 71, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, M.S.; Perry, C.G.; Arkell, A.M.; Chabowski, A.; Simpson, J.A.; Wright, D.C.; Holloway, G.P. Impairments in mitochondrial palmitoyl-CoA respiratory kinetics that precede development of diabetic cardiomyopathy are prevented by resveratrol in ZDF rats. J. Physiol. 2014, 592, 2519–2533. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, T.; Caso, R.; McMahon, D.; Nymark, M. A novel, multi-ingredient supplement to manage elevated blood lipids in patients with no evidence of cardiovascular disease: A pilot study. Altern. Ther. Health Med. 2014, 20, 18–23. [Google Scholar] [PubMed]
- Micans, P. Aortic wave velocity: A noninvasive method to measure the stiffness of arteries and the clinical results of supplements that appear to improve arterial stiffness. Curr. Aging Sci. 2013, 6, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Popat, R.; Plesner, T.; Davies, F.; Cook, G.; Cook, M.; Elliott, P.; Jacobson, E.; Gumbleton, T.; Oakervee, H.; Cavenagh, J. A phase 2 study of SRT501 (resveratrol) with bortezomib for patients with relapsed and or refractory multiple myeloma. Br. J. Haematol. 2013, 160, 714–717. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, B.; Campen, M.J.; Channell, M.M.; Wherry, S.J.; Varamini, B.; Davis, J.G.; Baur, J.A.; Smoliga, J.M. Resveratrol for primary prevention of atherosclerosis: Clinical trial evidence for improved gene expression in vascular endothelium. Int. J. Cardiol. 2013, 166, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. One-year consumption of a grape nutraceutical containing resveratrol improves the inflammatory and fibrinolytic status of patients in primary prevention of cardiovascular disease. Am. J. Cardiol. 2012, 110, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Urpi-Sarda, M.; Lamuela-Raventós, R.M.; Martínez-González, M.Á.; Salas-Salvadó, J.; Arós, F.; Fitó, M.; Lapetra, J.; Estruch, R.; Andres-Lacueva, C. PREDIMED study investigators. High urinary levels of resveratrol metabolites are associated with a reduction in the prevalence of cardiovascular risk factors in high-risk patients. Pharmacol. Res. 2012, 65, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Fujitaka, K.; Otani, H.; Jo, F.; Jo, H.; Nomura, E.; Iwasaki, M.; Nishikawa, M.; Iwasaka, T.; Das, D.K. Modified resveratrol Longevinex improves endothelial function in adults with metabolic syndrome receiving standard treatment. Nutr. Res. 2011, 31, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.H.; Howe, P.R.; Buckley, J.D.; Coates, A.M.; Kunz, I.; Berry, N.M. Acute resveratrol supplementation improves flow-mediated dilatation in overweight/obese individuals with mildly elevated blood pressure. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, A.; Paur, I.; Bøhn, S.K.; Sakhi, A.K.; Borge, G.I.; Serafini, M.; Erlund, I.; Laake, P.; Tonstad, S.; Blomhoff, R. Bilberry juice modulates plasma concentration of NF-kappaB related inflammatory markers in subjects at increased risk of CVD. Eur. J. Nutr. 2010, 49, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Gresele, P.; Pignatelli, P.; Guglielmini, G.; Carnevale, R.; Mezzasoma, A.M.; Ghiselli, A.; Momi, S.; Violi, F. Resveratrol, at concentrations attainable with moderate wine consumption, stimulates human platelet nitric oxide production. J. Nutr. 2008, 138, 1602–1608. [Google Scholar] [PubMed]
- Lekakis, J.; Rallidis, L.S.; Andreadou, I.; Vamvakou, G.; Kazantzoglou, G.; Magiatis, P.; Skaltsounis, A.L.; Kremastinos, D.T. Polyphenolic compounds from red grapes acutely improve endothelial function in patients with coronary heart disease. Eur. J. Cardiovasc. Prev. Rehabil. 2005, 12, 596–600. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Authors | Date | Place | Aim of the Study and Population Studied | Dose of RES | Summary of Main Findings | Side Effects |
|---|---|---|---|---|---|---|
| Faghihzadeh et al. [132] | 2015 | Iran | Evaluation of the effects of RES supplementation on CV risk factors in 55 patients with non alcoholic fatty liver disease (NAFLD), in a randomised double-blinded placebo-controlled clinical trial | Supplementation with a 500-mg RES capsule or a placebo capsule for 12 weeks | Supplementation with RES did not have any beneficial effect on anthropometric measurements, insulin resistance markers, lipid profile and blood pressure; however, it reduced alanine aminotransferase and hepatic steatosis in patients with NAFLD. | None reported |
| Amadio et al. [129] | 2015 | Italy | Study of the effect of prostaglandin E2 (PGE2) induced by cigarette smoke on tissue factor (TF) expression and activity in endothelial cells. No CVD. | 10 µM RES only used in vitro as SIRT-1 activator | (1) In vivo: positive correlation between PGE2 levels and TF antigen and activity in human smokers; similar findings in mice; (2) in vitro: exogenous or endogenous PGE2 up-regulated TF expression and activity via the EP1/SIRT1 pathway in endothelial cells. Cell treatment with exogenous PGI2 prevented the inhibition of SIRT1 and the induction of TF mediated by PGE2. | Not applicable (in vitro use of RES as SIRT-1 activator) |
| Van der Made et al. [133] | 2015 | The Netherlands | Randomized, placebo-controlled crossover study conducted in 45 overweight and slightly obese men (n = 25) and women (n = 20), with a mean age of 61 ± 7 years. Study of the effect of RES on apoA-I concentrations, and on other markers of lipid and lipoprotein metabolism, glucose metabolism, and markers of inflammation and endothelial function. | Subjects received in random order RES (150 mg per day) or placebo capsules for 4 weeks, separated by a 4-week wash-out period. | RES did not change metabolic risk markers related to CV health in overweight and slightly obese men and women. Effects on glucose metabolism not significant. | None reported |
| Hobbs et al. [151] | 2014 | USA | Effects of a multi-ingredient supplement featuring red yeast rice in patients with no CVD history or symptoms other than elevated blood lipids. | Study conducted in 19 hypercholesterolemic patients, to evaluate the effects of the supplement alone (in patients with trigly cerides <140 mg/dL) or associated with ω3-polyunsaturated fatty acids and vitamin E (in patients with triglycerides >140 mg/dL, receiving 1384 mg of ω-3 daily). The supplement contained a blend of red yeast rice, biofla-vonoids, polycosanol, 525 mg natural ω-3 fatty acids, RES, coenzyme Q10, folic acid, niacin, B6, B12, and black pepper. | The supplement decreased total cholesterol and LDL-cholesterol significantly, and addition of an ω-3 supplement also non significantly decreased triglycerides in the subgroup with triglycerides >140 mg/dL. Very small size of the group and RES associated with other components. | None reported |
| Semba et al. [134] | 2014 | USA | Prospective cohort study, the Invecchiare in Chianti (InCHIANTI) Study (“Aging in the Chianti Region”), conducted (1998–2009) in a population-based sample of 783 community-dwelling men and women ≥ 65 years, to determine whether RES levels achieved with diet were associated with inflammation, cancer, cardiovascular disease, and mortality in humans | 783 subjects had 24-h urine samples available for measurements of RES (no data on the dietary amount of RES) | Total urinary RES metabolite concentration was not associated with inflammatory markers, CVD or cancer, or predictive of all-cause mortality, therefore RES did not substantially influenced health status and mortality risk of this population. | None reported |
| Thushara et al. [130] | 2014 | India | No study on RES effects: study on ameliorative efficacy of crocin on sesamol-induced platelet apoptosis | |||
| Soare et al. [135] | 2014 | USA | 6-month randomized, single-blind controlled trial, in 56 non-obese men and women, aged 38 to 55 years, assigned to a dietary supplement (SUP) group or control (CON) group (24 weeks of daily supplementation), with a 6-month follow-up | SUP group : 10 dietary supplements each day (100 mg RES, a complex of 800 mg each of green, black, and white tea extract, 250 mg of pomegranate extract, 650 mg of quercetin, 500 mg of acetyl-l-carnitine, 600 mg of lipoic acid, 900 mg of curcumin, 1 g of sesamin, 1.7 g of cinnamon bark extract, and 1 g fish oil). | No effect on arterial stiffness, endothelial function or blood pressure in nonobese men and women. No effect on key metabolic variables involved in aging and in CVD, including plasma markers of inflammation, oxidative stress and glycation, plasma lipids, growth factors, or body composition. Lack of beneficial metabolic effects perhaps related to the low phytochemical bioavailability or inadequate supplement potency of the phytochemicals. | Adverse events limited to mild gastro intestinal discomfort associated with taking the large number of oral supplements in 19% of the participants. |
| Micans [152] | 2013 | UK | Measurement of aortic wave velocity as a noninvasive method to evaluate the stiffness of arteries. Small clinical trial (20 patients) to appreciate the effects of supplements that appear to improve arterial stiffness | 4 groups of patients: placebo, arginine, trans-RES (45 mg), l-carnosine, aminoguanidine: aortic wave pressure test before the supplement (baseline), and 1 h, 1 week and 1 month after supplement | Results for RES were given after 45 mg trans-RES given dialy for 5 weeks, showing an improvement of 15% in the aortic wave velocity test. No effect observed in the placebo roup. | None reported |
| Tomé-Carneiro et al. [125] | 2013 | Spain | Study of the molecular changes in peripheral blood mononuclear cells (PBMCs) associated to the one-year daily intake of a RES-enriched grape extract (GE-RES) in hypertensive male patients with type 2 diabetes mellitus, constituting a subset of 35 patients from a larger intervention study previously described [83] | Patients randomly allocated as follows: placebo (n = 9), GE (n = 13) and GE-RES (containing 8 mg RES) (n = 13) | Supplementation with GE or GE-RES did not affect body weight, blood pressure, glucose, HbA1c or lipids; no significant change in serum inflammatory markers, only a significant reduction of alkaline phosphatase and IL-6 levels. The expression of the pro-inflammatory cytokines CCL3, IL-1β and TNF-α was significantly reduced and that of the transcriptional repressor LRRFIP-1 increased in PBMCs from patients taking the GE-RES extract. A group of miRNAs involved in the regulation of the inflammatory response were found to be highly correlated and altered in the group consuming the GE-RES for 12 months. | None reported |
| Bo et al. [44] | 2013 | Italy | Randomized, double-blind, cross-over trial performed in 50 healthy adult smokers | 25 subjects randomly allocated to “RES-first” (30-days: 500 mg RES/day, 30-days wash-out, 30-days placebo) and 25 to “placebo-first” (30-days placebo, 30-days wash-out, 30-days 500 mg RES/day) | RES significantly reduced C-reactive protein and triglyceride concentrations, and increased total antioxidant status values. No change in uric acid, glucose, insulin, cholesterol, liver enzyme concentrations, weight, waist circumference, and blood pressure. | None reported |
| Tomé-Carneiro et al. [92] | 2013 | Spain | Triple-blind, randomized, placebo-controlled, one-year follow-up, 3-arm pilot clinical trial of dose-depending effects of a RES-containing grape supplement on 75 stable patients with coronary artery disease treated according to guidelines for secondary prevention of CVD | 3 arms: 350 mg/day of placebo, RES-containing grape extract (grape phenolics plus 8 mg RES) or conventional grape extract lacking RES during 6 months, and a double dose for the following 6 months | After 1 year, in contrast to the placebo and conventional grape extract groups, the RES-containing grape extract group showed an increase of the anti-inflammatory serum adiponectin and a decrease of the thrombogenic PAI-1; inhibition of atherothrombotic signals in peripheral blood mononuclear cells. | None reported |
| Popat et al. [153] | 2013 | UK | Not applicable: phase 2 study of SRT501 (RES) with bortezomib for patients with relapsed and or refractory multiple myeloma | |||
| Militaru et al. [95] | 2013 | Romania | Randomized, double-blinded, active-controlled, parallel clinical trial with 60 days oral supplementation with calcium fructoborate, RES, and their combination, in 166 subjects with stable angina pectoris | 3 groups (with usual medical care and treatment): group 1 received a single daily capsule with RES 20 mg/day (10 mg trans-RES); group 2 received a single daily capsule of RES 20 mg/day (10 mg trans-RES) combined with calcium fructoborate (CF) 112 mg/day (3 mg boron); group 3 received a single daily capsule of CF 112 mg/day (3 mg boron). Non-randomized control group: only usual medical care and treatment. | Significant decrease of hs-CRP in all groups at the 30-day and 60-day visits, greater for group 3, followed by group 2. The NT-proBNP was significantly lowered in groups 1 and 3, but combination RES + CF (group 2) was the most effective. Lipid markers showed slight changes from baseline in all groups. Improvement in the quality of life: best for group 2. | None reported |
| Agarwal et al. [154] | 2013 | USA | Double-blind, randomized, placebo-controlledtudy of the effects of RES on endothelial response and plasma biomarkers in 44 healthy individuals using a novel unbiased assay to assess the overall inflammatory capacity of plasma on expression of genes associated with inflammation and atherosclerosis | 1 month treatment RES supplement : 400 mg trans-RES (98% pure, from Polygonum Cuspidatum), 400 mg of mixed Bordeaux whole grape extract (90% polyphenols), and 100 mg quercetin | Exposing cultured human coronary artery endothelial cells to plasma drawn post-RES resulted in significantly lower mRNA expression of VCAM, ICAM, and IL-8 than plasma drawn from the same subjects at baseline (no effect of placebo). Significant reduction in plasma interferon-γ with RES supplement (not with placebo), and significant reduction in fasting insulin concentration with RES supplement | 2 subjects with RES and 1 with placebo reported mild gastrointestinal side effects |
| Tomé-Carneiro et al. [45] | 2012 | Spain | Study of the effect of a grape supplement in oxidized LDL (LDLox), apolipoprotein-B (ApoB), and serum lipids on 75 statin-treated patients in primary CVD prevention | Grape exctract containing 8 mg RES. 3 parallel arms: one capsule (350 mg) daily for 6 months containing RES-enriched grape extract (GE-RES, Stilvid®), grape extract (GE, similar polyphenolic content but no RES), or placebo (maltodextrin) | LDL-cholesterol, ApoB, LDLox and LDLox/ApoB decreased in the Stilvid® group, whereas the ratio non-HDL-cholesterol/ApoB increased, which might exert additional cardioprotection | None reported |
| Tomé-Carneiro et al. [155] | 2012 | Spain | 75 patients undergoing primary prevention of CVD participated in this triple-blinded, randomized, parallel, dose-response, placebo-controlled, 1-year follow-up trial: effects of a dietary RES-rich grape supplement on the inflammatory and fibrinolytic status | 3 groups: placebo (maltodextrin), or RES-rich grape supplement (8 mg RES), or conventional grape supplement lacking RES, for the first 6 months, and a double dose for the next 6 months | 1-year consumption of a RES-rich grape supplement improved the inflammatory and fibrinolytic status in patients who were on statins for primary prevention of CVD and at high CVD risk (i.e., with diabetes or hypercholesterolemia plus ≥1 other CV risk factor) | None reported |
| Zamora-Ros et al. [156] | 2012 | Spain | Study of the associations between total urinary RES metabolites (TRMs) as biomarkers of wine and RES consumption and CV risk factors in a large cross-sectional study including high CV risk individuals (1000 participants—479 men and 521 women—of the PREDIMED Study) | TRMs were used as a biomarker of wine consumption (when the model was not adjusted for alcohol intake) or as biomarkers of RES intake (when the model was additionally adjusted for alcohol intake) | Both RES and wine intake, evaluated as TRMs, were associated with beneficial changes in blood lipid profiles, fasting blood glucose (only RES) and heart rate, suggesting that RES intake via wine consumption might help to decrease CV risk factors | None reported |
| Magyar et al. [93] | 2012 | Hungary | Double-blind, placebo controlled trial conducted in 40 post-infarction Caucasian patients to investigate if RES had a clinically measurable cardioprotective effect | 10 mg RES daily (capsule) for 3 months | RES improved left ventricle diastolic function, endothelial function, lowered LDL-cholesterol level and protected against unfavourable hemorheological changes measured in patients with coronary artery disease | None reported |
| Fujitaka et al. [157] | 2011 | Japan | Study of Longevinex, a modified RES, on endothelial function in 34 patients with metabolic syndrome | 2 groups: group A, with Longevinex administered for 3 months and then discontinued for 3 months, and group B, with Longevinex administered between 3 and 6 months. 1 capsule of Longevinex daily, containing 100 mg trans-RES | Longevinex specifically improved endothelial function (flow-mediated dilatation) in subjects with metabolic syndrome | None reported |
| Wong et al. [158] | 2011 | Australia | Is consumption of RES able to improve flow-mediated dilatation (FMD) of the brachial artery is a biomarker of endothelial function, in 19 overweight/obese (BMI 25–35) men or post-menopausal women, aged 30–70 years, with untreated borderline hypertension (systolic BP: 130–160 mmHg or diastolic BP: 85–100 mmHg) | 3 doses of RES in RES capsules (resVida™ 30, 90 and 270 mg) or placebo capsules, in randomised, double-blind, placebo-controlled, crossover human intervention trial comprising 4 visits at weekly intervals (note that the lowest RES dose (30 mg) used in this study cannot be obtained from normal dietary habits) | First study to evaluate the acute effects of RES consumption on human circulatory function: significant dose effect of RES on plasma RES concentration (p < 0.001) and on FMD (p < 0.01), which increased from 4.1 ± 0.8% (placebo) to 7.7 ± 1.5% after 270 mg RES | None reported |
| Karlsen et al. [159] | 2010 | Norway | Effect of bilberry juice (RES content: 1–12 mg/100 g fw) on serum and plasma biomarkers of inflammation and antioxidant status in men aged between 30 and 70 years and women between 45 and 70 years and at least 12 months postmenopausal, at elevated risk of CVD | One group consumed 330 mL bilberry juice/day (diluted in 1 L water) and one group consumed 1 L water/day | Supplementation with bilberry juice resulted in significant decreases in plasma concentrations of C-reactive protein (CRP), interleukin (IL)-6, IL-15, and monokine induced by interferon-g (MIG) | None reported |
| Gresele et al. [160] | 2008 | Italy | Study of the effects of RES, at concentrations attainable after moderate wine intake, on platelet ●NO production in 20 healthy volunteers. Moreover, RES at the concentrations detected in plasma after wine intake, was incubated in vitro with washed platelets and several variables related to ●NO production and to signal transduction were measured | Study before and after 15 days of controlled white or red wine intake (300 mL/day): total polyphenolic concentration = 1.8 g/L for red wine and 0.25 g/L for white wine. Maximum RES concentration for in vitro studies: 0.5 μmol/L | After wine intake, increase of plasma RES and release of ●NO by stimulated platelets. In vitro, RES enhanced production of ●NO by stimulated platelets, activity of platelet ●NO synthase (NOS), phosphorylation of protein kinase B, an activator of the endothelial NOS (eNOS), and phosphorylation of vasodilator-activated protein. | None reported |
| Zamora-Ros et al. [131] | 2006 | Spain | Analysis of urinary total RES metabolites (TRMs) as a biomarker of moderate wine (sparkling, white, or red wine) consumption, in 2 open, prospective, randomized, crossover, single-blinded clinical trials | Daily amount of total RES (for 4 weeks): 0.357, 0.398, and 2.56 mg for sparkling, white, and red wine, respectively | RES metabolites in urine may be useful biomarkers of wine intake in epidemiologic and intervention studies (the aim of the study was not to report beneficial effect of RES in patients with CV risk) | None reported |
| Cruz et al. [127] | 2006 | Sweden | Investigation of acute vasodilator responses to phytoestrogens and selective estrogen receptor-alpha (ERalpha) agonist in isolated small arteries from 15 men (38–71 years) with established coronary heart disease (CHD) and with a history of MI (1–12 months before study) vs. healthy male control subjects | 10-30 µM RES | Phytoestrogens (especially RES), at concentrations achievable by ingestion of phytoestrogen-rich food products, induced dilatation ex vivo of small peripheral arteries from normal men and from those with established CHD. The contribution of ●NO to dilatory responses by these compounds is pertinent to arteries from control males, whereas other ●NO-independent dilatory mechanism(s) are involved in arteries from CHD | Not applicable (ex vivo study) |
| Lekakis et al. [161] | 2005 | Greece | 30 male patients with coronary heart disease, randomly assigned either to a red grape polyphenol extract dissolved in 20 mL of water (n = 15) or 20 mL of water (placebo) (n = 15), to examine whether acute intake of the extract has a positive effect on brachial artery flow-mediated dilatation | Use of 600 mg extract containing 0.9 mg trans-RES and other polyphenolic compounds | Intake of the extract caused an increase in flow-mediated dilatation, which was significantly higher than the baseline values. No change was observed after intake of placebo. | None reported (the long-term effect of the extract on endothe-lial function has not been studied) |
| Rakici et al. [128] | 2005 | Turkey | First study of the relaxant effect of RES on human blood vessels of internal mammary artery and saphenous vein grafts from 38 randomized male patients undergoing coronary artery revascularization | 10–70 μM RES | 70 µM RES caused relaxation in saphenous vein and internal mammary artery (mainly endothelium-dependent and ●NO-mediated relaxations in internal mammary artery, partially in saphenous vein rings) | Not applica-ble (ex vivo study) |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonnefont-Rousselot, D. Resveratrol and Cardiovascular Diseases. Nutrients 2016, 8, 250. https://doi.org/10.3390/nu8050250
Bonnefont-Rousselot D. Resveratrol and Cardiovascular Diseases. Nutrients. 2016; 8(5):250. https://doi.org/10.3390/nu8050250
Chicago/Turabian StyleBonnefont-Rousselot, Dominique. 2016. "Resveratrol and Cardiovascular Diseases" Nutrients 8, no. 5: 250. https://doi.org/10.3390/nu8050250
APA StyleBonnefont-Rousselot, D. (2016). Resveratrol and Cardiovascular Diseases. Nutrients, 8(5), 250. https://doi.org/10.3390/nu8050250