Vitamin D and Inflammatory Cytokines in Healthy and Preeclamptic Pregnancies



Abstract
:1. Introduction
2. Calcitriol in Non-Pregnant and Pregnant Women
3. Calcitriol in Preeclamptic Pregnancy
3.1. Association between Calcitriol and Hypocalciuria in Preeclampsia
3.2. Alteration of Vitamin D Metabolism in Preeclampsia
3.3. Fetoplacental Synthesis of Calcitriol in Pregnancy
3.4. Placental Vitamin D Metabolism in Normotensive and Preeclamptic Pregnancies

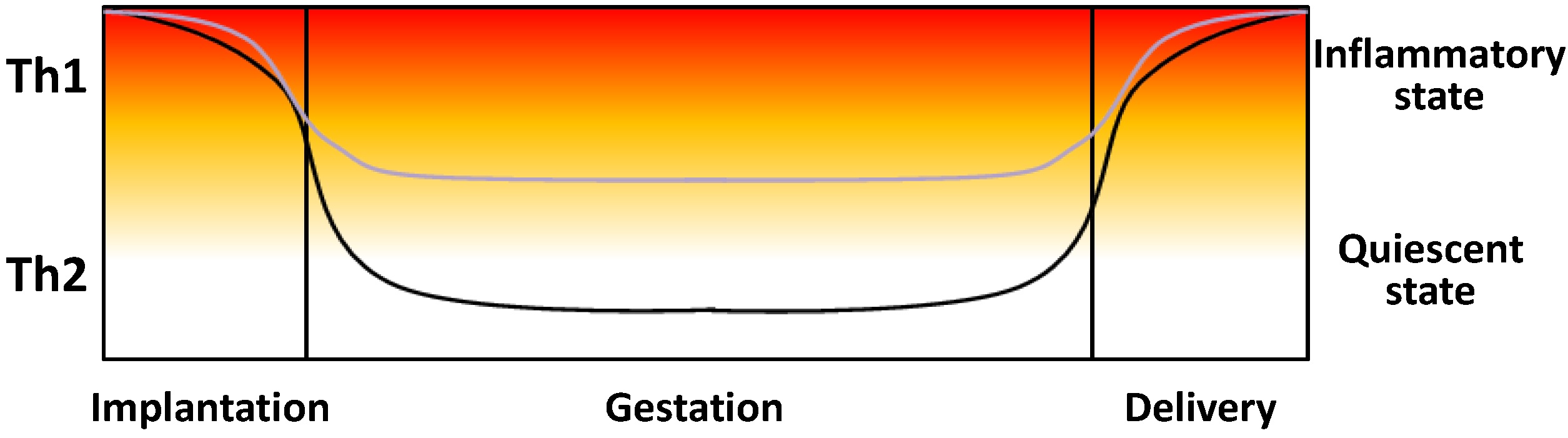
4. Circulating Cytokines in Healthy and Preeclamptic Pregnancies

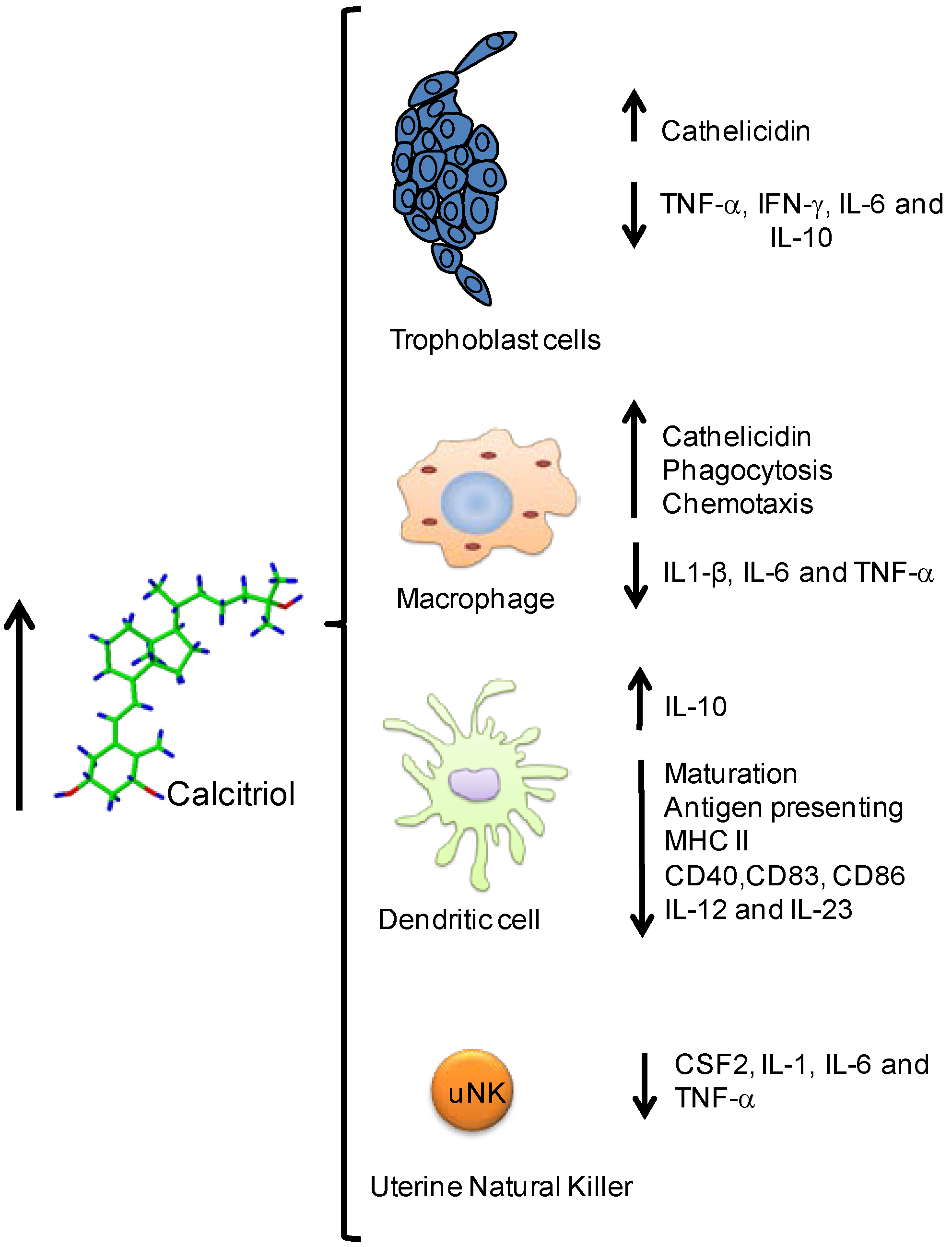{kind=link}
{kind=link}
{kind=link}
| Cytokines in Preeclamptic Pregnancies | References |
|---|---|
| Genetic polymorphisms of TNF-α and IL-1 result in increased levels of these pro-inflammatory cytokines and are associated with preeclampsia. | [106] |
| Serum and placental levels of pro-inflammatory IL-18 are increased in preeclampsia. | [107] |
| The ratios IL-2/IL-10 and TNF-α/IL-10 in maternal serum are higher in preeclampsia than in normal pregnancy. | [108] |
| Pro-inflammatory cytokines IL-6 and IL-8 are increased in maternal serum from women with preeclampsia as compared with normal pregnancy. | [109] |
| Serum levels of pro-inflammatory IL-15 and IL-16 are significantly higher in preeclampsia than in normal pregnancy. | [110] |
| Gene expression of anti-inflammatory IL-4 is low in preeclampsia. | [111] |
| In preeclampsia, the pro-inflammatory TNF-α and IL-6 and C-reactive protein are higher compared to normal pregnancy. | [112] |
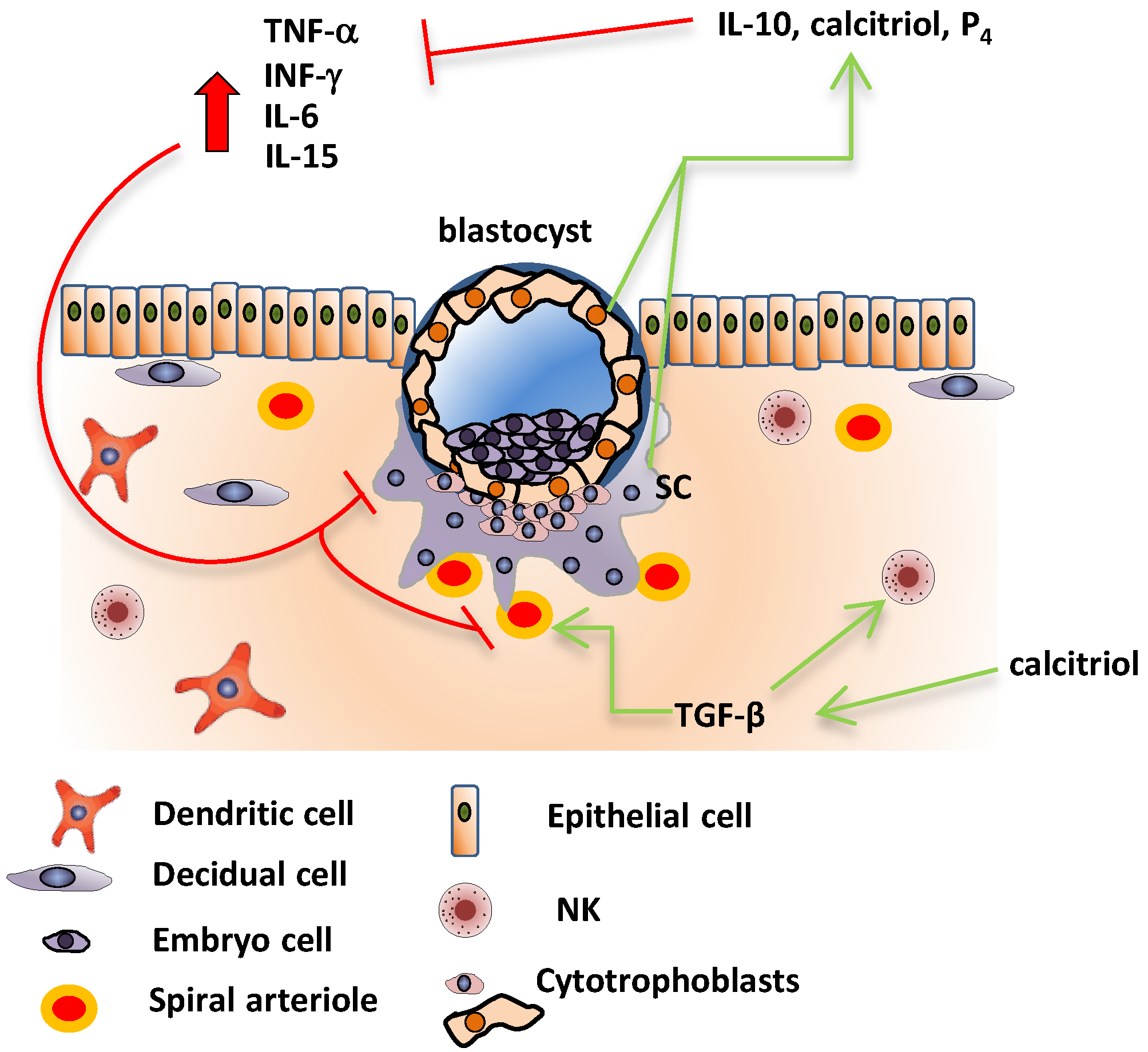
5. Fetoplacental Cytokines in Healthy and Preeclamptic Pregnancies
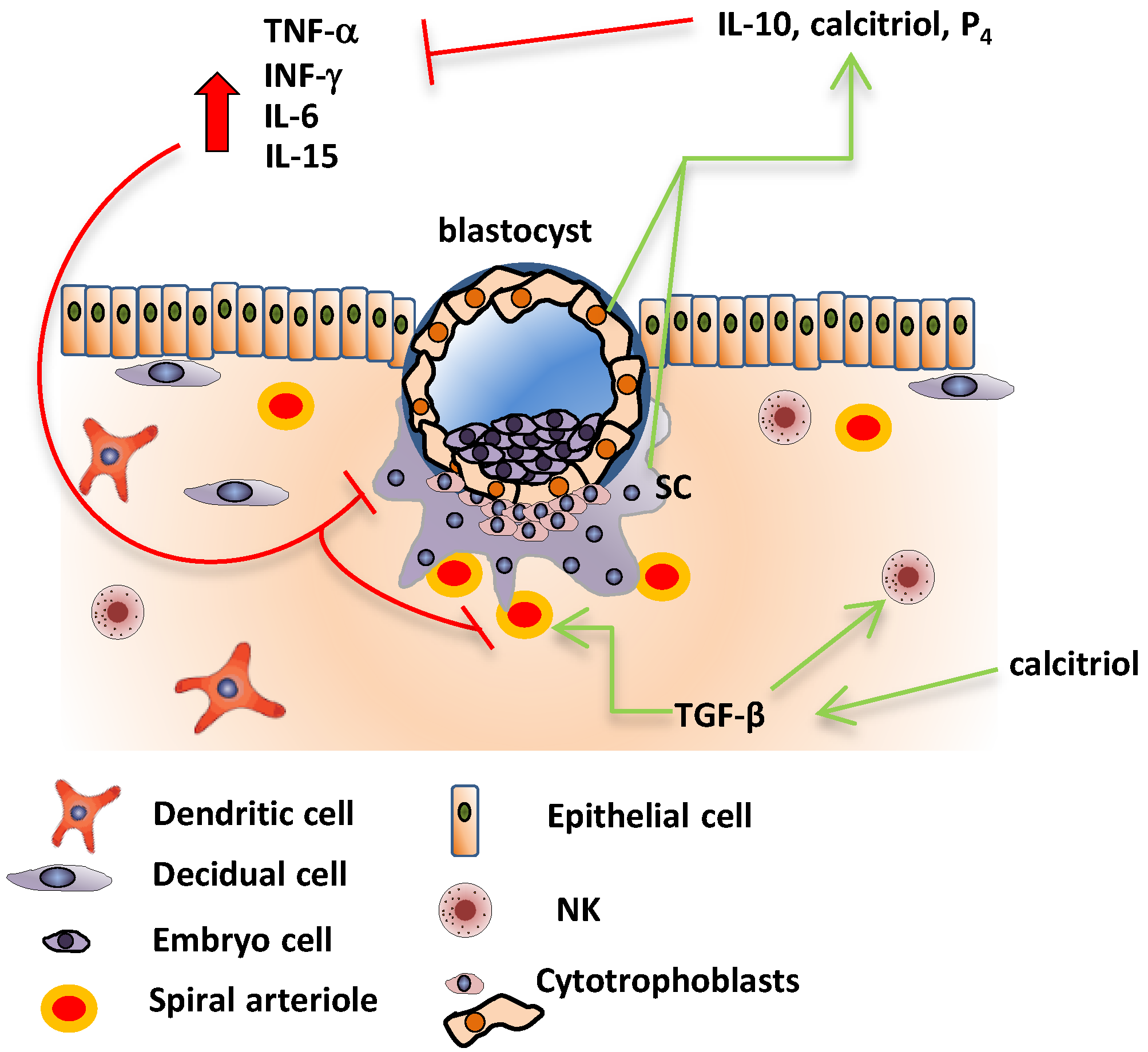
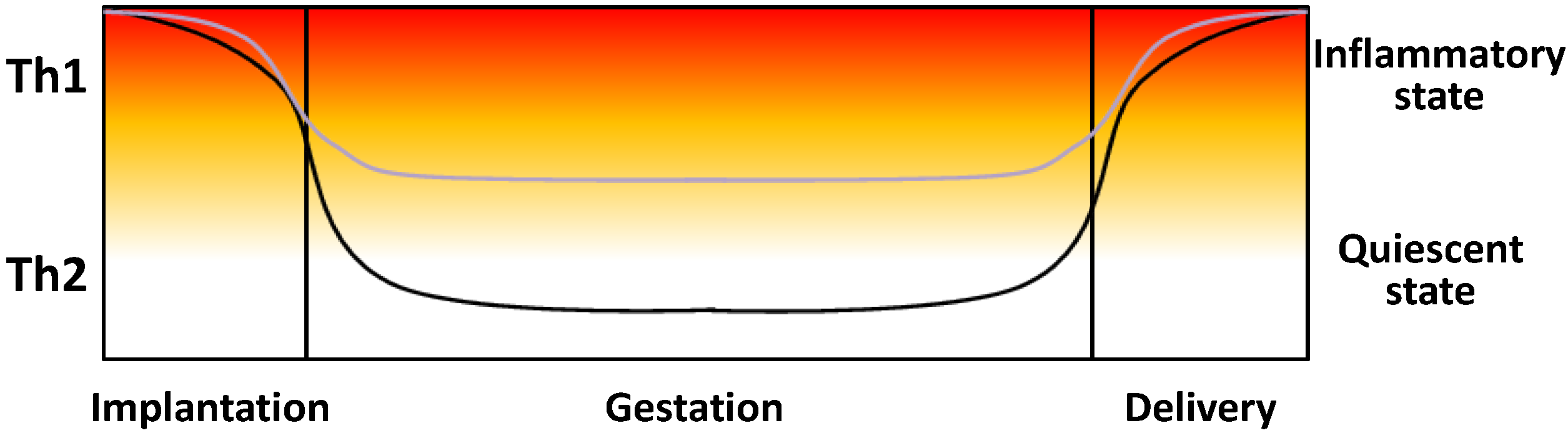
6. Effects of Calcitriol upon Inflammatory Cytokines in Human Placenta
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- National High Blood Pressure Education Program Working Group on High Blood Pressure in Pregnancy. Report of the National High Blood Pressure Education Program Working Group on high blood pressure in pregnancy. Am. J. Obstet. Gynecol. 2000, 183, S1–S22. [Google Scholar]
- August, P.; Marcaccio, B.; Gertner, J.M.; Druzin, M.L.; Resnick, L.M.; Laragh, J.H. Abnormal 1,25-dihydroxyvitamin D metabolism in preeclampsia. Am. J. Obstet. Gynecol. 1992, 166, 1295–1299. [Google Scholar] [CrossRef]
- Halhali, A.; Bourges, H.; Carrillo, A.; Garabedian, M. Lower circulating insulin-like growth factor I and 1,25-dihydroxyvitamin D levels in preeclampsia. Rev. Investig. Clin. 1995, 47, 259–266. [Google Scholar]
- Halhali, A.; Diaz, L.; Avila, E.; Ariza, A.C.; Garabedian, M.; Larrea, F. Decreased fractional urinary calcium excretion and serum 1,25-dihydroxyvitamin D and IGF-I levels in preeclampsia. J. Steroid Biochem. Mol. Biol. 2007, 103, 803–806. [Google Scholar] [CrossRef] [PubMed]
- Halhali, A.; Tovar, A.R.; Torres, N.; Bourges, H.; Garabedian, M.; Larrea, F. Preeclampsia is associated with low circulating levels of insulin-like growth factor I and 1,25-dihydroxy vitamin D in maternal and umbilical cord compartments. J. Clin. Endocrinol. Metab. 2000, 85, 1828–1833. [Google Scholar] [PubMed]
- Seely, E.W.; Wood, R.J.; Brown, E.M.; Graves, S.W. Lower serum ionized calcium and abnormal calciotropic hormone levels in preeclampsia. J. Clin. Endocrinol. Metab. 1992, 74, 1436–1440. [Google Scholar] [PubMed]
- LaMarca, B.; Cornelius, D.; Wallace, K. Elucidating immune mechanisms causing hypertension during pregnancy. Physiology 2013, 28, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Laresgoiti-Servitje, E. A leading role for the immune system in the pathophysiology of preeclampsia. J. Leukoc. Biol. 2013, 94, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Laresgoiti-Servitje, E.; Gomez-Lopez, N.; Olson, D.M. An immunological insight into the origins of pre-eclampsia. Hum. Reprod. Update 2010, 16, 510–524. [Google Scholar] [CrossRef] [PubMed]
- Perez-Sepulveda, A.; Torres, M.J.; Khoury, M.; Illanes, S.E. Innate immune system and preeclampsia. Front. Immunol. 2014, 5, 244. [Google Scholar] [CrossRef] [PubMed]
- DeLuca, H.F. Overview of general physiologic features and functions of vitamin D. Am. J. Clin. Nutr. 2004, 80, 1689S–1696S. [Google Scholar] [PubMed]
- Hewison, M. Vitamin D and immune function: Autocrine, paracrine or endocrine? Scand. J. Clin. Lab. Iinvestig. Suppl. 2012, 243, 92–102. [Google Scholar]
- Hewison, M. An update on vitamin D and human immunity. Clin. Endocrinol. 2012, 76, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Hewison, M. Vitamin D and immune function: An overview. Proc. Nutr. Soc. 2012, 71, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Thota, C.; Laknaur, A.; Farmer, T.; Ladson, G.; Al-Hendy, A.; Ismail, N. Vitamin D regulates contractile profile in human uterine myometrial cells via NF-κB pathway. Am. J. Obstet. Gynecol. 2014, 210, e341–e347. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.W. From vitamin D to hormone D: Fundamentals of the vitamin D endocrine system essential for good health. Am. J. Clin. Nutr. 2008, 88, 491S–499S. [Google Scholar] [PubMed]
- Garabedian, M.; Holick, M.F.; Deluca, H.F.; Boyle, I.T. Control of 25-hydroxycholecalciferol metabolism by parathyroid glands. Proc. Natl. Acad. Sci. USA 1972, 69, 1673–1676. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Hewison, M.; Gardner, D.G.; Wagner, C.L.; Sergeev, I.N.; Rutten, E.; Pittas, A.G.; Boland, R.; Ferrucci, L.; Bikle, D.D. Vitamin D: Beyond bone. Ann. N. Y. Acad. Sci. 2013, 1287, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Tamblyn, J.A.; Hewison, M.; Wagner, C.L.; Bulmer, J.N.; Kilby, M.D. Immunological role of vitamin D at the maternal-fetal interface. J. Endocrinol. 2015, 224, R107–R121. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, C.S. Vitamin D in pregnancy and lactation: Maternal, fetal, and neonatal outcomes from human and animal studies. Am. J. Clin. Nutr. 2008, 88, 520S–528S. [Google Scholar] [PubMed]
- Salle, B.L.; Delvin, E.E.; Lapillonne, A.; Bishop, N.J.; Glorieux, F.H. Perinatal metabolism of vitamin D. Am. J. Clin. Nutr. 2000, 71, 1317S–1324S. [Google Scholar] [PubMed]
- Kovacs, C.S.; Kronenberg, H.M. Maternal-fetal calcium and bone metabolism during pregnancy, puerperium, and lactation. Endocr. Rev. 1997, 18, 832–872. [Google Scholar] [CrossRef] [PubMed]
- Ardawi, M.S.; Nasrat, H.A.; BA'Aqueel, H.S. Calcium-regulating hormones and parathyroid hormone-related peptide in normal human pregnancy and postpartum: A longitudinal study. Eur. J. Endocrinol. 1997, 137, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Halhali, A.; Villa, A.R.; Madrazo, E.; Soria, M.C.; Mercado, E.; Diaz, L.; Avila, E.; Garabedian, M.; Larrea, F. Longitudinal changes in maternal serum 1,25-dihydroxyvitamin D and insulin like growth factor I levels in pregnant women who developed preeclampsia: Comparison with normotensive pregnant women. J. Steroid Biochem. Mol. Biol. 2004, 89–90, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Hollis, B.W.; Johnson, D.; Hulsey, T.C.; Ebeling, M.; Wagner, C.L. Vitamin D supplementation during pregnancy: Double-blind, randomized clinical trial of safety and effectiveness. J. Bone Miner. Res. 2011, 26, 2341–2357. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Resurrection of vitamin D deficiency and rickets. J. Clin. Investig. 2006, 116, 2062–2072. [Google Scholar] [CrossRef] [PubMed]
- Brannon, P.M.; Picciano, M.F. Vitamin D in pregnancy and lactation in humans. Ann. Rev. Nutr. 2011, 31, 89–115. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M.; Halhali, A.; Guillozo, H.; Garabedian, M.; Balsan, S. Thyroid and parathyroid-independent increase in plasma 1,25-dihydroxyvitamin D during late pregnancy in the rat. J. Endocrinol. 1988, 116, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.S.; Norman, A.W.; Willis, D.M.; Goltzman, D.; Guyda, H.; Solomon, S.; Philips, D.R.; Bishop, J.E.; Mayer, E. Regulation of vitamin D metabolism in normal human pregnancy. J. Clin. Endocrinol. Metab. 1983, 56, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Breslau, N.A.; Zerwekh, J.E. Relationship of estrogen and pregnancy to calcium homeostasis in pseudohypoparathyroidism. J. Clin. Endocrinol. Metab. 1986, 62, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Menaa, C.; Vrtovsnik, F.; Friedlander, G.; Corvol, M.; Garabedian, M. Insulin-like growth factor I, a unique calcium-dependent stimulator of 1,25-dihydroxyvitamin D3 production. Studies in cultured mouse kidney cells. J. Biol. Chem. 1995, 270, 25461–25467. [Google Scholar] [CrossRef] [PubMed]
- Halhali, A.; Diaz, L.; Sanchez, I.; Garabedian, M.; Bourges, H.; Larrea, F. Effects of IGF-I on 1,25-dihydroxyvitamin D3 synthesis by human placenta in culture. Mol. Hum. Reprod. 1999, 5, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Rosol, T.J.; Capen, C.C.; Horst, R.L. Effects of infusion of human parathyroid hormone-related protein-(1–40) in nude mice: Histomorphometric and biochemical investigations. J. Bone Miner. Res. 1988, 3, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, T.; Drezner, M.K. Abnormal parathyroid hormone-related peptide stimulation of renal 25-hydroxyvitamin D-1-hydroxylase in hyp mice: Evidence for a generalized defect of enzyme activity in the proximal convoluted tubule. Endocrinology 1990, 127, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, M.J.; Tedesco, M.B.; Sereika, S.M.; Syed, M.A.; Garcia-Ocana, A.; Bisello, A.; Hollis, B.W.; Rosen, C.J.; Wysolmerski, J.J.; Dann, P.; et al. Continuous PTH and PTHrP infusion causes suppression of bone formation and discordant effects on 1,25(OH)2 vitamin D. J. Bone Miner. Res. 2005, 20, 1792–1803. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, M.J.; Tedesco, M.B.; Sereika, S.M.; Hollis, B.W.; Garcia-Ocana, A.; Stewart, A.F. Direct comparison of sustained infusion of human parathyroid hormone-related protein-(1-36) [hPTHrP-(1–36)] versus hPTH-(1–34) on serum calcium, plasma 1,25-dihydroxyvitamin D concentrations, and fractional calcium excretion in healthy human volunteers. J. Clin. Endocrinol. Metab. 2003, 88, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ramos, L.; Jones, D.C.; Cullen, M.T. Urinary calcium as an early marker for preeclampsia. Obstet. Ggynecol. 1991, 77, 685–688. [Google Scholar] [CrossRef]
- Taufield, P.A.; Ales, K.L.; Resnick, L.M.; Druzin, M.L.; Gertner, J.M.; Laragh, J.H. Hypocalciuria in preeclampsia. N. Engl. J. Med. 1987, 316, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Halhali, A.; Diaz, L.; Barrera, D.; Avila, E.; Larrea, F. Placental calcitriol synthesis and IGF-I levels in normal and preeclamptic pregnancies. J. Steroid Biochem. Mol. Biol. 2014, 144, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, E.B.; Johannesen, P.; Kristensen, S.; Rasmussen, A.B.; Emmertsen, K.; Moller, J.; Lauritsen, J.G.; Wohlert, M. Calcium, parathyroid hormone and calcitonin in normal pregnancy and preeclampsia. Gynecol. Oobstet. Investig. 1984, 18, 156–164. [Google Scholar] [CrossRef]
- Tolaymat, A.; Sanchez-Ramos, L.; Yergey, A.L.; Vieira, N.E.; Abrams, S.A.; Edelstein, P. Pathophysiology of hypocalciuria in preeclampsia: Measurement of intestinal calcium absorption. Obstet. Gynecol. 1994, 83, 239–243. [Google Scholar] [CrossRef]
- Diaz, E.; Halhali, A.; Luna, C.; Diaz, L.; Avila, E.; Larrea, F. Newborn birth weight correlates with placental zinc, umbilical insulin-like growth factor I, and leptin levels in preeclampsia. Arch. Med. Res. 2002, 33, 40–47. [Google Scholar] [CrossRef]
- Kocyigit, Y.; Atamer, Y.; Atamer, A.; Tuzcu, A.; Akkus, Z. Changes in serum levels of leptin, cytokines and lipoprotein in pre-eclamptic and normotensive pregnant women. Gynecol. Endocrinol. 2004, 19, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Williams, M.A.; Vadachkoria, S.; Muy-Rivera, M.; Frederick, I.O.; Luthy, D.A. Maternal plasma concentrations of insulinlike growth factor-1 and insulinlike growth factor-binding protein-1 in early pregnancy and subsequent risk of preeclampsia. Clin. Biochem. 2004, 37, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Olmos, A.; Diaz, L.; Avila, E.; Barrera, D.; Lopez-Marure, R.; Biruete, B.; Larrea, F.; Halhali, A. Associations between insulin-like growth factor I, vascular endothelial growth factor and its soluble receptor 1 in umbilical serum and endothelial cells obtained from normotensive and preeclamptic pregnancies. Growth Factors 2013, 31, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.Y.; Xue, M.; Xia, A.B. Study on changes of IGF-I and leptin levels in serum and placental tissue of prceclampsia patients and their associativity. Chin. J. Cel. Mol. Immunol. 2011, 27, 192–194. [Google Scholar]
- Shang, L.X.; Wang, J.; Zhang, L.J.; Gao, H.; Qu, D.Y.; Wang, J.H. Relationship between changes of insulin like growth factor-1 and insulin like growth factor binding protein-1 in maternal serum and placenta and pathogenesis of hypertensive disorder complicating pregnancy. Zhonghua Fu Chan Ke Za Zhi 2005, 40, 516–520. [Google Scholar] [PubMed]
- Sifakis, S.; Akolekar, R.; Kappou, D.; Mantas, N.; Nicolaides, K.H. Maternal serum insulin-like growth factor-I at 11–13 weeks in preeclampsia. Prenat. Diagn. 2010, 30, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Valerio, E.G.; Muller, A.L.; Martins-Costa, S.H.; Lopes Ramos, J.G.; Rodini, G. Are insulin resistance index, IGF-1 and metabolic syndrome components correlates with severe preeclampsia? Hypertens. Pregnancy 2011, 30, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Dubova, E.A.; Pavlov, K.A.; Lyapin, V.M.; Kulikova, G.V.; Shchyogolev, A.I.; Sukhikh, G.T. Expression of insulin-like growth factors in the placenta in preeclampsia. Bull. Exp. Biol. Med. 2014, 157, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Halhali, A.; Wimalawansa, S.J.; Berentsen, V.; Avila, E.; Thota, C.S.; Larrea, F. Calcitonin gene- and parathyroid hormone-related peptides in preeclampsia: Effects of magnesium sulfate. Obstet. Gynecol. 2001, 97, 893–897. [Google Scholar] [CrossRef]
- Yadav, S.; Yadav, Y.S.; Goel, M.M.; Singh, U.; Natu, S.M.; Negi, M.P. Calcitonin gene- and parathyroid hormone-related peptides in normotensive and preeclamptic pregnancies: A nested case-control study. Arch. Gynecol. Obstet. 2014, 290, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Abedi, P.; Mohaghegh, Z.; Afshary, P.; Latifi, M. The relationship of serum vitamin D with pre-eclampsia in the iranian women. Mater. Child Nutr. 2014, 10, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Achkar, M.; Dodds, L.; Giguere, Y.; Forest, J.C.; Armson, B.A.; Woolcott, C.; Agellon, S.; Spencer, A.; Weiler, H.A. Vitamin D status in early pregnancy and risk of preeclampsia. Am. J. Obstet. Gynecol. 2015, 212, e511–e517. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.; Basu, J.; Das, S.K.; Misra, A.K. Effects of age, stage of disease, and educational level on cognitive dysfunction in non-demented idiopathic parkinsonism: A preliminary report. Ind. Psychiatry J. 2012, 21, 32–38. [Google Scholar] [PubMed]
- Adhikari, S.; Karmahapatra, S.K.; Karve, T.M.; Bandyopadhyay, S.; Woodrick, J.; Manthena, P.V.; Glasgow, E.; Byers, S.; Saha, T.; Uren, A. Characterization of magnesium requirement of human 5′-tyrosyl DNA phosphodiesterase mediated reaction. BMC Res. Notes 2012, 5, 134. [Google Scholar] [CrossRef] [PubMed]
- Aghajafari, F.; Nagulesapillai, T.; Ronksley, P.E.; Tough, S.C.; O’Beirne, M.; Rabi, D.M. Association between maternal serum 25-hydroxyvitamin D level and pregnancy and neonatal outcomes: Systematic review and meta-analysis of observational studies. BMJ 2013, 346, f1169. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.M.; Haeri, S.; Camargo, C.A., Jr.; Espinola, J.A.; Stuebe, A.M. A nested case-control study of midgestation vitamin D deficiency and risk of severe preeclampsia. J. Clin. Endocrinol. Metab. 2010, 95, 5105–5109. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Catov, J.M.; Simhan, H.N.; Holick, M.F.; Powers, R.W.; Roberts, J.M. Maternal vitamin D deficiency increases the risk of preeclampsia. J. Clin. Endocrinol. Metab. 2007, 92, 3517–3522. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Simhan, H.N.; Catov, J.M.; Roberts, J.M.; Platt, R.W.; Diesel, J.C.; Klebanoff, M.A. Maternal vitamin D status and the risk of mild and severe preeclampsia. Epidemiology 2014, 25, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Mohaghegh, Z.; Abedi, P.; Dilgouni, T.; Namvar, F.; Ruzafza, S. The relation of preeclampsia and serum level of 25-hydroxyvitamin D in mothers and their neonates: A case control study in iran. Horm. Metab. Res. 2015, 47, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.J.; Alanis, M.C.; Wagner, C.L.; Hollis, B.W.; Johnson, D.D. Plasma 25-hydroxyvitamin D levels in early-onset severe preeclampsia. Am. J. Obstet. Gynecol. 2010, 203, e361–e366. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.J.; Wagner, C.L.; Hollis, B.W.; Baatz, J.E.; Johnson, D.D. Maternal vitamin D and fetal growth in early-onset severe preeclampsia. Am. J. Obstet. Gynecol. 2011, 204, e551–e556. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.J.; Wagner, C.L.; Hollis, B.W.; Baatz, J.E.; Johnson, D.D. Association of maternal vitamin D and placenta growth factor with the diagnosis of early onset severe preeclampsia. Am. J. Perinatol. 2013, 30, 167–172. [Google Scholar] [PubMed]
- Singla, R.; Gurung, P.; Aggarwal, N.; Dutta, U.; Kochhar, R. Relationship between preeclampsia and vitamin D deficiency: A case control study. Arch. Gynecol. Obstet. 2015, 29, 1247–1251. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.I.; Koch, C.A.; Tamanna, S.; Rouf, S.; Shamsuddin, L. Vitamin D deficiency and the risk of preeclampsia and eclampsia in bangladesh. Horm. Metab. Res. 2013, 45, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.Q.; Audibert, F.; Hidiroglou, N.; Sarafin, K.; Julien, P.; Wu, Y.; Luo, Z.C.; Fraser, W.D. Longitudinal vitamin D status in pregnancy and the risk of pre-eclampsia. BJOG 2012, 119, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Woodham, P.C.; Brittain, J.E.; Baker, A.M.; Long, D.L.; Haeri, S.; Camargo, C.A., Jr.; Boggess, K.A.; Stuebe, A.M. Midgestation maternal serum 25-hydroxyvitamin D level and soluble fms-like tyrosine kinase 1/placental growth factor ratio as predictors of severe preeclampsia. Hypertension 2011, 58, 1120–1125. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lee, M.; Jeyabalan, A.; Roberts, J.M. The relationship of hypovitaminosis D and IL-6 in preeclampsia. Am. J. Obstet. Gynecol. 2014, 210, e141–e147. [Google Scholar] [CrossRef] [PubMed]
- Burris, H.H.; Rifas-Shiman, S.L.; Huh, S.Y.; Kleinman, K.; Litonjua, A.A.; Oken, E.; Rich-Edwards, J.W.; Camargo, C.A., Jr.; Gillman, M.W. Vitamin D status and hypertensive disorders in pregnancy. Ann. Epidemiol. 2014, 24, e399–e403. [Google Scholar] [CrossRef] [PubMed]
- Dalmar, A.; Raff, H.; Chauhan, S.P.; Singh, M.; Siddiqui, D.S. Serum 25-hydroxyvitamin D, calcium, and calcium-regulating hormones in preeclamptics and controls during first day postpartum. Endocrine 2015, 48, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Skowronska-Jozwiak, E.; Lebiedzinska, K.; Smyczynska, J.; Lewandowski, K.C.; Glowacka, E.; Lewinski, A. Effects of maternal vitamin D status on pregnancy outcomes, health of pregnant women and their offspring. Neuro Endocrinol. Lett. 2014, 35, 367–372. [Google Scholar] [PubMed]
- Zhou, J.; Su, L.; Liu, M.; Liu, Y.; Cao, X.; Wang, Z.; Xiao, H. Associations between 25-hydroxyvitamin D levels and pregnancy outcomes: A prospective observational study in southern china. Eur. J. Clin. Nutr. 2014, 68, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Wacker, M.; Holick, M.F. Sunlight and vitamin D: A global perspective for health. Dermatoendocrinology 2013, 5, 51–108. [Google Scholar] [CrossRef] [PubMed]
- Lechtermann, C.; Hauffa, B.P.; Herrmann, R.; Schundeln, M.M.; Gellhaus, A.; Schmidt, M.; Grasemann, C. Maternal vitamin D status in preeclampsia: Seasonal changes are not influenced by placental gene expression of vitamin d metabolizing enzymes. PLoS ONE 2014, 9, e105558. [Google Scholar] [CrossRef] [PubMed]
- Olmos-Ortiz, A.; Avila, E.; Durand-Carbajal, M.; Diaz, L. Regulation of calcitriol biosynthesis and activity: Focus on gestational vitamin D deficiency and adverse pregnancy outcomes. Nutrients 2015, 7, 443–480. [Google Scholar] [CrossRef] [PubMed]
- Tabesh, M.; Salehi-Abargouei, A.; Tabesh, M.; Esmaillzadeh, A. Maternal vitamin D status and risk of pre-eclampsia: A systematic review and meta-analysis. J. Clin. Endocrinol. Metab. 2013, 98, 3165–3173. [Google Scholar] [CrossRef] [PubMed]
- Hofmeyr, G.J.; Belizan, J.M.; von Dadelszen, P. Low-dose calcium supplementation for preventing pre-eclampsia: A systematic review and commentary. BJOG 2014, 121, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Delvin, E.E.; Arabian, A.; Glorieux, F.H.; Mamer, O.A. In vitro metabolism of 25-hydroxycholecalciferol by isolated cells from human decidua. J. Clin. Endocrinol. Metab. 1985, 60, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Weisman, Y.; Vargas, A.; Duckett, G.; Reiter, E.; Root, A.W. Synthesis of 1,25-dihydroxyvitamin D in the nephrectomized pregnant rat. Endocrinology 1978, 103, 1992–1996. [Google Scholar] [CrossRef] [PubMed]
- Gray, T.K.; Lester, G.E.; Lorenc, R.S. Evidence for extra-renal 1 alpha-hydroxylation of 25-hydroxyvitamin D3 in pregnancy. Science 1979, 204, 1311–1313. [Google Scholar] [CrossRef] [PubMed]
- Weisman, Y.; Harell, A.; Edelstein, S.; David, M.; Spirer, Z.; Golander, A. 1 alpha, 25-dihydroxyvitamin D3 and 24,25-dihydroxyvitamin D3 in vitro synthesis by human decidua and placenta. Nature 1979, 281, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Tanamura, A.; Nomura, S.; Kurauchi, O.; Furui, T.; Mizutani, S.; Tomoda, Y. Purification and characterization of 1,25(OH)2 D3 receptor from human placenta. J. Obstet. Gynecol. 1995, 21, 631–639. [Google Scholar]
- Zerwekh, J.E.; Breslau, N.A. Human placental production of 1 alpha,25-dihydroxyvitamin D3: Biochemical characterization and production in normal subjects and patients with pseudohypoparathyroidism. J. Clin. Endocrinol. Metab. 1986, 62, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Hollis, B.W.; Iskersky, V.N.; Chang, M.K. In vitro metabolism of 25-hydroxyvitamin D3 by human trophoblastic homogenates, mitochondria, and microsomes: Lack of evidence for the presence of 25-hydroxyvitamin D3-1 alpha- and 24R-hydroxylases. Endocrinology 1989, 125, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.; Sanchez, I.; Avila, E.; Halhali, A.; Vilchis, F.; Larrea, F. Identification of a 25-hydroxyvitamin D3 1alpha-hydroxylase gene transcription product in cultures of human syncytiotrophoblast cells. J. Clin. Endocrinol. Metab. 2000, 85, 2543–2549. [Google Scholar] [PubMed]
- Adams, J.S.; Hewison, M. Extrarenal expression of the 25-hydroxyvitamin D-1-hydroxylase. Arch. Biochem. Biophys. 2012, 523, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S.; Rafison, B.; Witzel, S.; Reyes, R.E.; Shieh, A.; Chun, R.; Zavala, K.; Hewison, M.; Liu, P.T. Regulation of the extrarenal CYP27B1-hydroxylase. J. Steroid Biochem. Mol. Biol. 2014, 144, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.; Arranz, C.; Avila, E.; Halhali, A.; Vilchis, F.; Larrea, F. Expression and activity of 25-hydroxyvitamin D-1 alpha-hydroxylase are restricted in cultures of human syncytiotrophoblast cells from preeclamptic pregnancies. J. Clin. Endocrinol. Metab. 2002, 87, 3876–3882. [Google Scholar] [PubMed]
- Fischer, D.; Schroer, A.; Ludders, D.; Cordes, T.; Bucker, B.; Reichrath, J.; Friedrich, M. Metabolism of vitamin D3 in the placental tissue of normal and preeclampsia complicated pregnancies and premature births. Clin. Exp. Obstet. Ggynecol. 2007, 34, 80–84. [Google Scholar]
- Ma, R.; Gu, Y.; Zhao, S.; Sun, J.; Groome, L.J.; Wang, Y. Expressions of vitamin d metabolic components VDBP, CYP2R1, CYP27B1, CYP24A1, and VDR in placentas from normal and preeclamptic pregnancies. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E928–E935. [Google Scholar] [CrossRef] [PubMed]
- Noyola-Martinez, N.; Diaz, L.; Zaga-Clavellina, V.; Avila, E.; Halhali, A.; Larrea, F.; Barrera, D. Regulation of CYP27B1 and CYP24A1 gene expression by recombinant pro-inflammatory cytokines in cultured human trophoblasts. J. Steroid Biochem. Mol. Biol. 2014, 144, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.; Noyola-Martinez, N.; Barrera, D.; Hernandez, G.; Avila, E.; Halhali, A.; Larrea, F. Calcitriol inhibits TNF-α-induced inflammatory cytokines in human trophoblasts. J. Reprod. Immunol. 2009, 81, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, B.; Sibson, M.; Ng, H.K.; Manuelpillai, U.; Rakyan, V.; Down, T.; Beck, S.; Fournier, T.; Evain-Brion, D.; Dimitriadis, E.; et al. Placenta-specific methylation of the vitamin D 24-hydroxylase gene: Implications for feedback autoregulation of active vitamin D levels at the fetomaternal interface. J. Biol. Chem. 2009, 284, 14838–14848. [Google Scholar] [CrossRef] [PubMed]
- Barrera, D.; Avila, E.; Hernandez, G.; Mendez, I.; Gonzalez, L.; Halhali, A.; Larrea, F.; Morales, A.; Diaz, L. Calcitriol affects hCG gene transcription in cultured human syncytiotrophoblasts. Reprod. Biol. Endocrinol. 2008, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Barrera, D.; Avila, E.; Hernandez, G.; Halhali, A.; Biruete, B.; Larrea, F.; Diaz, L. Estradiol and progesterone synthesis in human placenta is stimulated by calcitriol. J. Steroid Biochem. Mol. Biol. 2007, 103, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Halhali, A.; Figueras, A.G.; Diaz, L.; Avila, E.; Barrera, D.; Hernandez, G.; Larrea, F. Effects of calcitriol on calbindins gene expression and lipid peroxidation in human placenta. J. Steroid Biochem. Mol. Biol. 2010, 121, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.Y.; Susarla, R.; Canovas, D.; Vasilopoulou, E.; Ohizua, O.; McCabe, C.J.; Hewison, M.; Kilby, M.D. Vitamin D promotes human extravillous trophoblast invasion in vitro. Placenta 2015, 36, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Gu, B.; Gu, Y.; Groome, L.J.; Sun, J.; Wang, Y. Activation of vitamin D receptor promotes VEGF and CuZn-SOD expression in endothelial cells. J. Steroid Biochem. Mol. Biol. 2014, 140, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Brodowski, L.; Burlakov, J.; Myerski, A.C.; von Kaisenberg, C.S.; Grundmann, M.; Hubel, C.A.; von Versen-Hoynck, F. Vitamin D prevents endothelial progenitor cell dysfunction induced by sera from women with preeclampsia or conditioned media from hypoxic placenta. PLoS ONE 2014, 9, e98527. [Google Scholar] [CrossRef] [PubMed]
- Barrera, D.; Noyola-Martinez, N.; Avila, E.; Halhali, A.; Larrea, F.; Diaz, L. Calcitriol inhibits interleukin-10 expression in cultured human trophoblasts under normal and inflammatory conditions. Cytokine 2012, 57, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Noyola-Martinez, N.; Diaz, L.; Avila, E.; Halhali, A.; Larrea, F.; Barrera, D. Calcitriol downregulates TNF-α and IL-6 expression in cultured placental cells from preeclamptic women. Cytokine 2013, 61, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Latest advances in understanding preeclampsia. Science 2005, 308, 1592–1594. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Immunology of pre-eclampsia. Am. J. Reprod. Immunol. 2010, 63, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Moffett, A.; Loke, Y.W. The immunological paradox of pregnancy: A reappraisal. Placenta 2004, 25, 1–8. [Google Scholar] [CrossRef]
- Haggerty, C.L.; Ferrell, R.E.; Hubel, C.A.; Markovic, N.; Harger, G.; Ness, R.B. Association between allelic variants in cytokine genes and preeclampsia. Am. J. Obstet. Gynecol. 2005, 193, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Huang, H.; Dong, M.; Yao, Q.; Wang, H. Serum and placental interleukin-18 are elevated in preeclampsia. J. Reprod. Immunol. 2005, 65, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Wang, Z.; He, J. Serum T helper 1- and 2-type cytokines in preeclampsia. Int. J. Gynaecol. Obst. 2005, 89, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, Y.; Ruber, M.; Matthiesen, L.; Berg, G.; Nieminen, K.; Sharma, S.; Ernerudh, J.; Ekerfelt, C. Cytokine mapping of sera from women with preeclampsia and normal pregnancies. J. Reprod. Immunol. 2006, 70, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, H.; Wang, Z.; Huang, H.; Dong, M. Elevated serum levels of interleukin-15 and interleukin-16 in preeclampsia. J. Reprod. Immunol. 2007, 73, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Rajakumar, A.; Chu, T.; Handley, D.E.; Bunce, K.D.; Burke, B.; Hubel, C.A.; Jeyabalan, A.; Peters, D.G. Maternal gene expression profiling during pregnancy and preeclampsia in human peripheral blood mononuclear cells. Placenta 2011, 32, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Mihu, D.; Razvan, C.; Malutan, A.; Mihaela, C. Evaluation of maternal systemic inflammatory response in preeclampsia. Taiwan J. Obstet. Gynecol. 2015, 54, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Denney, J.M.; Nelson, E.L.; Wadhwa, P.D.; Waters, T.P.; Mathew, L.; Chung, E.K.; Goldenberg, R.L.; Culhane, J.F. Longitudinal modulation of immune system cytokine profile during pregnancy. Cytokine 2011, 53, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Szarka, A.; Rigo, J., Jr.; Lazar, L.; Beko, G.; Molvarec, A. Circulating cytokines, chemokines and adhesion molecules in normal pregnancy and preeclampsia determined by multiplex suspension array. BMC Immunol. 2010, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Makhseed, M.; Raghupathy, R.; Azizieh, F.; Al-Azemi, M.M.; Hassan, N.A.; Bandar, A. Mitogen-induced cytokine responses of maternal peripheral blood lymphocytes indicate a differential Th-type bias in normal pregnancy and pregnancy failure. Am. J. Reprod. Immunol. 1999, 42, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Sakai, M.; Sasaki, Y.; Tanebe, K.; Tsuda, H.; Michimata, T. Quantitative analysis of peripheral blood Th0, Th1, Th2 and the Th1:Th2 cell ratio during normal human pregnancy and preeclampsia. Clin. Exp. Immunol. 1999, 117, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Holtan, S.G.; Chen, Y.; Kaimal, R.; Creedon, D.J.; Enninga, E.A.; Nevala, W.K.; Markovic, S.N. Growth modeling of the maternal cytokine milieu throughout normal pregnancy: Macrophage-derived chemokine decreases as inflammation/counterregulation increases. J. Immunol. Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Christian, L.M.; Porter, K. Longitudinal changes in serum proinflammatory markers across pregnancy and postpartum: Effects of maternal body mass index. Cytokine 2014, 70, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Austgulen, R.; Lien, E.; Liabakk, N.B.; Jacobsen, G.; Arntzen, K.J. Increased levels of cytokines and cytokine activity modifiers in normal pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 1994, 57, 149–155. [Google Scholar] [CrossRef]
- Belo, L.; Santos-Silva, A.; Rocha, S.; Caslake, M.; Cooney, J.; Pereira-Leite, L.; Quintanilha, A.; Rebelo, I. Fluctuations in C-reactive protein concentration and neutrophil activation during normal human pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2005, 123, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Kraus, T.A.; Sperling, R.S.; Engel, S.M.; Lo, Y.; Kellerman, L.; Singh, T.; Loubeau, M.; Ge, Y.; Garrido, J.L.; Rodriguez-Garcia, M.; et al. Peripheral blood cytokine profiling during pregnancy and post-partum periods. Am. J. Reprod. Immunol. 2010, 64, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Makhseed, M.; Raghupathy, R.; Azizieh, F.; Farhat, R.; Hassan, N.; Bandar, A. Circulating cytokines and CD30 in normal human pregnancy and recurrent spontaneous abortions. Hum. Reprod. 2000, 15, 2011–2017. [Google Scholar] [CrossRef] [PubMed]
- Opsjln, S.L.; Wathen, N.C.; Tingulstad, S.; Wiedswang, G.; Sundan, A.; Waage, A.; Austgulen, R. Tumor necrosis factor, interleukin-1, and interleukin-6 in normal human pregnancy. Am. J. Obstet. Gynecol. 1993, 169, 397–404. [Google Scholar] [CrossRef]
- Sharma, A.; Satyam, A.; Sharma, J.B. Leptin, IL-10 and inflammatory markers (TNF-α, IL-6 and IL-8) in pre-eclamptic, normotensive pregnant and healthy non-pregnant women. Am. J. Reprod. Immunol. 2007, 58, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Vassiliadis, S.; Ranella, A.; Papadimitriou, L.; Makrygiannakis, A.; Athanassakis, I. Serum levels of pro- and anti-inflammatory cytokines in non-pregnant women, during pregnancy, labour and abortion. Mediat. Inflamm. 1998, 7, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Conrad, K.P.; Miles, T.M.; Benyo, D.F. Circulating levels of immunoreactive cytokines in women with preeclampsia. Am. J. Reprod. Immunol. 1998, 40, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Kalantar, F.; Rajaei, S.; Heidari, A.B.; Mansouri, R.; Rashidi, N.; Izad, M.H.; Mirahmadian, M. Serum levels of tumor necrosis factor-α, interleukin-15 and interleukin-10 in patients with pre-eclampsia in comparison with normotensive pregnant women. Iran. J. Nurs. Midwifery Res. 2013, 18, 463–466. [Google Scholar] [PubMed]
- Lau, S.Y.; Guild, S.J.; Barrett, C.J.; Chen, Q.; McCowan, L.; Jordan, V.; Chamley, L.W. Tumor necrosis factor-alpha, interleukin-6, and interleukin-10 levels are altered in preeclampsia: A systematic review and meta-analysis. Am. J. Reprod. Immunol. 2013, 70, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Yao, M.Z.; Liu, J.B.; Xiong, L.K. A meta-analysis of tumor necrosis factor-α, interleukin-6, and interleukin-10 in preeclampsia. Cytokine 2011, 56, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Casart, Y.C.; Tarrazzi, K.; Camejo, M.I. Serum levels of interleukin-6, interleukin-1β and human chorionic gonadotropin in pre-eclamptic and normal pregnancy. Gynecol. Endocrinol. 2007, 23, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Laskowska, M.; Leszczynska-Gorzelak, B.; Laskowska, K.; Oleszczuk, J. Evaluation of maternal and umbilical serum TNF alpha levels in preeclamptic pregnancies in the intrauterine normal and growth-restricted fetus. J. Matern. Fetal Neonatal Med. 2006, 19, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Vince, G.S.; Starkey, P.M.; Austgulen, R.; Kwiatkowski, D.; Redman, C.W. Interleukin-6, tumour necrosis factor and soluble tumour necrosis factor receptors in women with pre-eclampsia. Brit. J. Obstet. Gynaecol. 1995, 102, 20–25. [Google Scholar] [CrossRef]
- PrabhuDas, M.; Bonney, E.; Caron, K.; Dey, S.; Erlebacher, A.; Fazleabas, A.; Fisher, S.; Golos, T.; Matzuk, M.; McCune, J.M.; et al. Immune mechanisms at the maternal-fetal interface: Perspectives and challenges. Nat. Immunol. 2015, 16, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Fest, S.; Aldo, P.B.; Abrahams, V.M.; Visintin, I.; Alvero, A.; Chen, R.; Chavez, S.L.; Romero, R.; Mor, G. Trophoblast-macrophage interactions: A regulatory network for the protection of pregnancy. Am. J. Reprod. Immunol. 2007, 57, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Goldman-Wohl, D.; Hamani, Y.; Avraham, I.; Greenfield, C.; Natanson-Yaron, S.; Prus, D.; Cohen-Daniel, L.; Arnon, T.I.; Manaster, I.; et al. Decidual NK cells regulate key developmental processes at the human fetal-maternal interface. Nat. Med. 2006, 12, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Crocker, I.P.; Wareing, M.; Ferris, G.R.; Jones, C.J.; Cartwright, J.E.; Baker, P.N.; Aplin, J.D. The effect of vascular origin, oxygen, and tumour necrosis factor alpha on trophoblast invasion of maternal arteries in vitro. J. Pathol. 2005, 206, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Lash, G.E.; Otun, H.A.; Innes, B.A.; Bulmer, J.N.; Searle, R.F.; Robson, S.C. Inhibition of trophoblast cell invasion by TGFB1, 2, and 3 is associated with a decrease in active proteases. Biol. Reprod. 2005, 73, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Otun, H.A.; Lash, G.E.; Innes, B.A.; Bulmer, J.N.; Naruse, K.; Hannon, T.; Searle, R.F.; Robson, S.C. Effect of tumour necrosis factor-α in combination with interferon-γ on first trimester extravillous trophoblast invasion. J. Reprod. Immunol. 2011, 88, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Paradowska, E.; Blach-Olszewska, Z.; Gejdel, E. Constitutive and induced cytokine production by human placenta and amniotic membrane at term. Placenta 1997, 18, 441–446. [Google Scholar] [CrossRef]
- Kupferminc, M.J.; Peaceman, A.M.; Wigton, T.R.; Rehnberg, K.A.; Socol, M.L. Tumor necrosis factor-alpha is elevated in plasma and amniotic fluid of patients with severe preeclampsia. Am. J. Obstet. Gynecol. 1994, 170, 1752–1757. [Google Scholar] [CrossRef]
- Wang, Y.; Walsh, S.W. TNF alpha concentrations and mrna expression are increased in preeclamptic placentas. J. Reprod. Immunol. 1996, 32, 157–169. [Google Scholar] [CrossRef]
- Southcombe, J.H.; Redman, C.W.; Sargent, I.L.; Granne, I. IL-1 family cytokines and their regulatory proteins in normal pregnancy and pre-eclampsia. Clin. Exp. Iimmunol. 2015. [Google Scholar] [CrossRef]
- Taki, A.; Abe, M.; Komaki, M.; Oku, K.; Iseki, S.; Mizutani, S.; Morita, I. Expression of angiogenesis-related factors and inflammatory cytokines in placenta and umbilical vessels in pregnancies with preeclampsia and chorioamnionitis/funisitis. Congenit. Anom. 2012, 52, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Wilczynski, J.R.; Tchorzewski, H.; Banasik, M.; Glowacka, E.; Wieczorek, A.; Lewkowicz, P.; Malinowski, A.; Szpakowski, M.; Wilczynski, J. Lymphocyte subset distribution and cytokine secretion in third trimester decidua in normal pregnancy and preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 109, 8–15. [Google Scholar] [CrossRef]
- Sargent, I.L.; Borzychowski, A.M.; Redman, C.W. NK cells and human pregnancy—An inflammatory view. Trends Immunol. 2006, 27, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Sacks, G.P.; Redman, C.W.; Sargent, I.L. Monocytes are primed to produce the Th1 type cytokine IL-12 in normal human pregnancy: An intracellular flow cytometric analysis of peripheral blood mononuclear cells. Clin. Exp. Immunol. 2003, 131, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Shi, Z.; Wang, P.; You, W.; Liang, G. Comparative proteome profile of human placenta from normal and preeclamptic pregnancies. PLoS ONE 2013, 8, e78025. [Google Scholar] [CrossRef] [PubMed]
- Duff, S.E.; Li, C.; Garland, J.M.; Kumar, S. CD105 is important for angiogenesis: Evidence and potential applications. FASEB J. 2003, 17, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Sorensen, L.K.; Brooke, B.S.; Urness, L.D.; Davis, E.C.; Taylor, D.G.; Boak, B.B.; Wendel, D.P. Defective angiogenesis in mice lacking endoglin. Science 1999, 284, 1534–1537. [Google Scholar] [CrossRef] [PubMed]
- Reister, F.; Frank, H.G.; Kingdom, J.C.; Heyl, W.; Kaufmann, P.; Rath, W.; Huppertz, B. Macrophage-induced apoptosis limits endovascular trophoblast invasion in the uterine wall of preeclamptic women. Lab. Investig. 2001, 81, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Stanczuk, G.A.; McCoy, M.J.; Hutchinson, I.V.; Sibanda, E.N. The genetic predisposition to produce high levels of TGF-β1 impacts on the severity of eclampsia/pre-eclampsia. Acta Obstet. Gynecol. Scand. 2007, 86, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Co, E.C.; Gormley, M.; Kapidzic, M.; Rosen, D.B.; Scott, M.A.; Stolp, H.A.; McMaster, M.; Lanier, L.L.; Barcena, A.; Fisher, S.J. Maternal decidual macrophages inhibit NK cell killing of invasive cytotrophoblasts during human pregnancy. Biol. Reprod. 2013, 88, 155. [Google Scholar] [CrossRef] [PubMed]
- Moffett-King, A. Natural killer cells and pregnancy. Nat. Rev. Immunol. 2002, 2, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Muralimanoharan, S.; Maloyan, A.; Myatt, L. Evidence of sexual dimorphism in the placental function with severe preeclampsia. Placenta 2013, 34, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.M.; Chamley, L.; Mitchell, M.D.; Keelan, J.A. Cytokines of the placenta and extra-placental membranes: Biosynthesis, secretion and roles in establishment of pregnancy in women. Placenta 2002, 23, 239–256. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrera, D.; Díaz, L.; Noyola-Martínez, N.; Halhali, A. Vitamin D and Inflammatory Cytokines in Healthy and Preeclamptic Pregnancies. Nutrients 2015, 7, 6465-6490. https://doi.org/10.3390/nu7085293
Barrera D, Díaz L, Noyola-Martínez N, Halhali A. Vitamin D and Inflammatory Cytokines in Healthy and Preeclamptic Pregnancies. Nutrients. 2015; 7(8):6465-6490. https://doi.org/10.3390/nu7085293
Chicago/Turabian StyleBarrera, David, Lorenza Díaz, Nancy Noyola-Martínez, and Ali Halhali. 2015. "Vitamin D and Inflammatory Cytokines in Healthy and Preeclamptic Pregnancies" Nutrients 7, no. 8: 6465-6490. https://doi.org/10.3390/nu7085293
APA StyleBarrera, D., Díaz, L., Noyola-Martínez, N., & Halhali, A. (2015). Vitamin D and Inflammatory Cytokines in Healthy and Preeclamptic Pregnancies. Nutrients, 7(8), 6465-6490. https://doi.org/10.3390/nu7085293