Anti-Inflammatory Effect of Angelica gigas via Heme Oxygenase (HO)-1 Expression

Abstract
:1. Introduction
2. Experimental Section
2.1. Sample Preparation and Reagents
2.2. Determination of Nitric Oxide (NO) Production
2.3. ORAC Assay
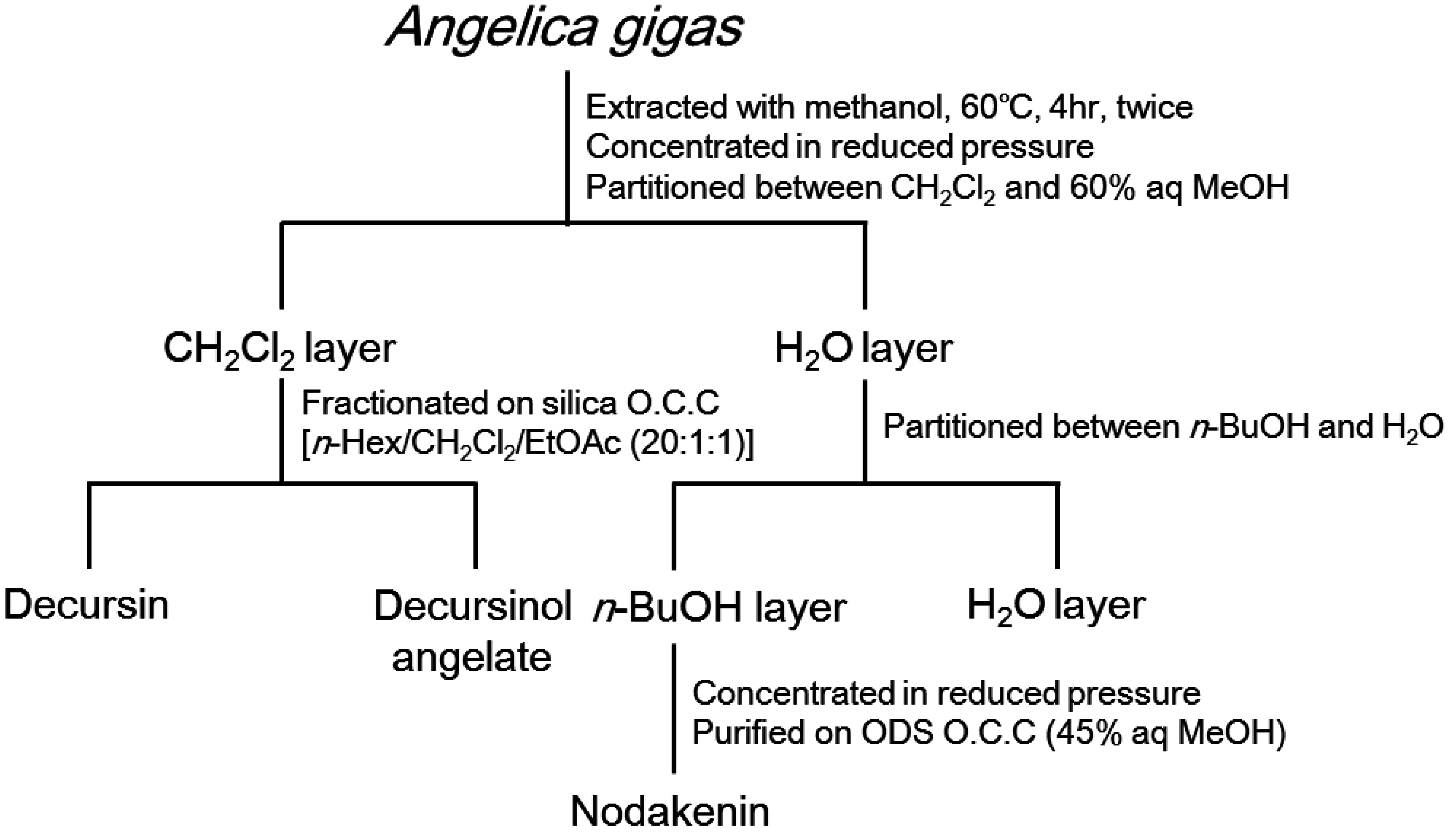
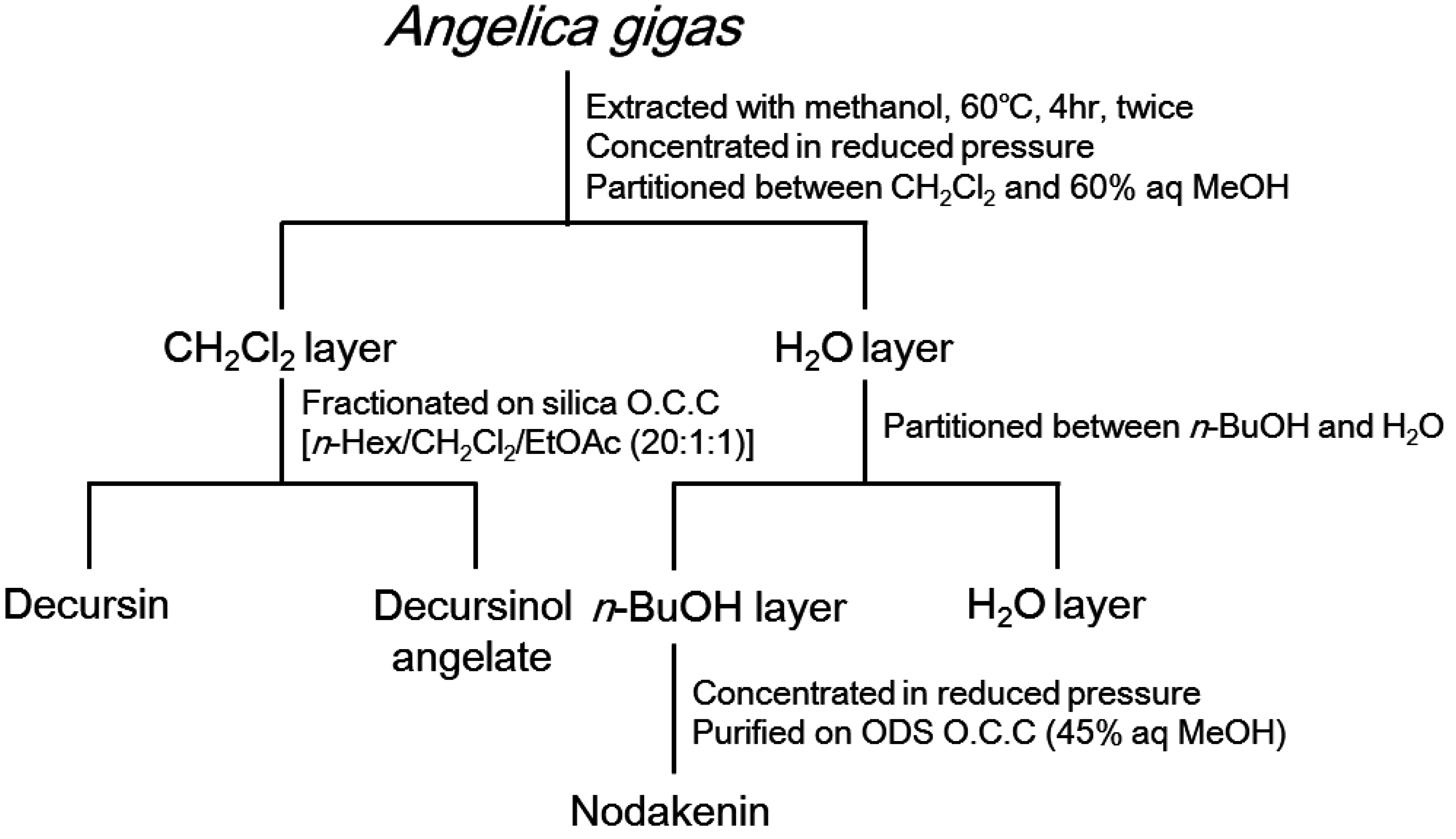
2.4. Extraction and Isolation
- Decursin (1)1H-NMR (500 MHz, CDCl3) δ 7.57 (1H, d, J = 9.5 Hz, H-4), 7.15 (1H, s, H-5), 6.76 (1H, s, H-8), 6.20 (1H, d, J = 9.5 Hz, H-3), 5.64 (1H, s, C-2’’), 5.07 (1H, t, J = 4.9 Hz, H-3’), 3.18 (1H, dd, J = 17.0, 4.6 Hz, H-4’a), 2.85 (1H, dd, J = 17.0, 4.9 Hz, H-4’b), 2.13 (3H, s, 3’’-CH3), 1.86 (3H, s, H-4’’), 1.37 (3H, s, gem.-CH3), 1.35 (3H, s, gem.-CH3); 13C-NMR (125 MHz, CDCl3) δ 165.73 (C-1’’), 161.28 (C-2), 158.43 (C-3’’), 156.46 (C-7), 154.14 (C-9), 143.23 (C-4), 128.71 (C-5), 115.99 (C-6), 115.52 (C-2’’), 113.17 (C-3), 112.79 (C-10), 104.61 (C-8), 76.75 (C-2’), 69.11 (C-3’), 27.88 (C-4’), 27.46 (C-4’’), 25.00 (gem.-CH3), 23.13 (gem.-CH3), 20.32 (3’’-CH3)
- Decursinol angelate (2)1H-NMR (500 MHz, CDCl3) δ 7.59 (1H, d, J = 9.5 Hz, H-4), 7.16 (1H, s, H-5), 6.76 (1H, s, H-8), 6.20 (1H, d, J = 9.5 Hz, H-3), 6.10 (1H, d, J = 7.3 Hz, H-3’’), 5.12 (1H, t, J = 4.8 Hz, H-3’), 3.22 (1H, dd, J = 17.8, 4.8 Hz H-4’a), 2.90 (1H, dd, J = 17.8, 4.9 Hz, H-4’b), 1.87 (3H, d, J = 7.1 Hz, H-4’’), 1.82 (3H, s, 2’’-CH3), 1.39 (3H, s, gem.-CH3), 1.37 (3H, s, gem.-CH3); 13C-NMR (125 MHz, CDCl3) δ 167.01 (C-1’’), 161.26 (C-2), 156.45 (C-7), 154.17 (C-9), 143.21 (C-4), 139.40 (C-3’’), 128.67 (C-5), 127.29 (C-2’’), 115.88 (C-6), 113.22 (C-3), 112.79 (C-10), 104.55 (C-8), 76.67 (C-2’), 70.02 (C-3’), 27.87 (C-4’), 25.07 (gem.-CH3), 23.19 (gem.-CH3), 20.50 (2’’-CH3), 15.73 (C-4’’)
- Nodakenin (3)1H-NMR (500 MHz, DMSO-d6) δ 7.92 (1H, d, J = 9.7 Hz, H-4), 7.45 (1H, s, H-5), 6.77 (1H, s, H-8), 6.19 (1H, d, J = 9.7 Hz, H-3), 4.96 (1H, m, H-3’), 4.42 (1H, d, J = 7.8 Hz, H-1’’), 2.80~3.75 (9H, m, H-2’,3’,2’’,3’’,4’’,5’’,6’’), 1.30 (3H, s, gem.-CH3), 1.12 (3H, s, gem.-CH3); 13C-NMR (125 MHz, DMSO-d6) δ 163.51 (C-7), 161.00 (C-2), 154.17 (C-9), 145.17 (C-4), 126.09 (C-6), 124.41 (C-5), 112.70 (C-3), 111.72 (C-10), 97.65 (C-8), 97.25 (C-1’’), 90.23 (C-2’), 77.53 (C-5’’), 77.31 (C-3’’), 77.07 (C-4’), 73.94 (C-2’’), 70.68 (C-4’’), 61.64 (C-6’’), 29.60 (C-3’), 23.62 (gem.-CH3), 21.13 (gem.-CH3)
2.5. HO-1 Expression Assay
3. Results
3.1. NO Inhibition Effects

{kind=link}
{kind=link}
| Treat-Ment Conc. (μg/mL) | Positive Control (DEX) | AGE | n-Hexane | CH2Cl2 | EtOAc | n-BuOH | H2O | Decursin | Decursinol Angelate | Nodakenin |
|---|---|---|---|---|---|---|---|---|---|---|
| 25 | 18.2 ± 0.36 | 24.0 ± 0.75 | 23.3 ± 0.28 | 18.5 ± 0.49 | 21.0 ± 0.39 | 21.3 ± 0.36 | 21.6 ± 0.50 | 19.4 ± 0.24 | 21.7 ± 0.21 | 22.1 ± 0.35 |
| 50 | 13.5 ± 0.22 | 20.7 ± 0.64 | 22.4 ± 0.46 | 14.8 ± 0.54 | 20.2 ± 0.14 | 20.1 ± 0.45 | 20.6 ± 0.53 | 17.1 ± 0.16 | 20.1 ± 0.30 | 23.1 ± 0.50 |
| 100 | 11.1 ± 0.21 | 12.9 ± 0.20 | 20.3 ± 0.53 | 12.5 ± 0.87 | 19.2 ± 0.20 | 18.3 ± 0.32 | 19.0 ± 0.38 | 15.6 ± 0.71 | 16.4 ± 0.33 | 21.3 ± 0.26 |
| 200 | 8.15 ± 0.17 | 9.5 ± 0 29 | 19.4 ± 0.49 | 8.95 ± 1.68 | 19.5 ± 1.27 | 14.0 ± 0.38 | 18.5 ± 0.39 | 10.9 ± 0.13 | 14.6 ± 0.85 | 21.6 ± 0.12 |
3.2. Anti-Oxidative Effect of AGE and Isolated Coumarins
| Treatment Conc. (μg/mL) | Positive Control (Trolox, Vitamin E) | AGE | Decursin | Decursinol Angelate | Nodakenin |
|---|---|---|---|---|---|
| 100 | 1.00 ± 0.020 | 0.91 ± 0.043 | 0.50 ± 0.008 | 0.40 ± 0.009 | 0.33 ± 0.015 |
| 50 | 1.00 ± 0.017 | 0.59 ± 0.026 | 0.27 ± 0.008 | 0.28 ± 0.009 | 0.22 ± 0.037 |
| 25 | 1.00 ± 0.027 | 0.46 ± 0.015 | 0.16 ± 0.010 | 0.18 ± 0.006 | 0.16 ± 0.009 |
| 12.5 | 1.00 ± 0.009 | 0.32 ± 0.023 | 0.10 ± 0.004 | 0.11 ± 0.006 | 0.08 ± 0.007 |
| 6.25 | 1.00 ± 0.022 | 0.24 ± 0.022 | 0.05 ± 0.001 | 0.07 ± 0.002 | 0.04 ± 0.007 |
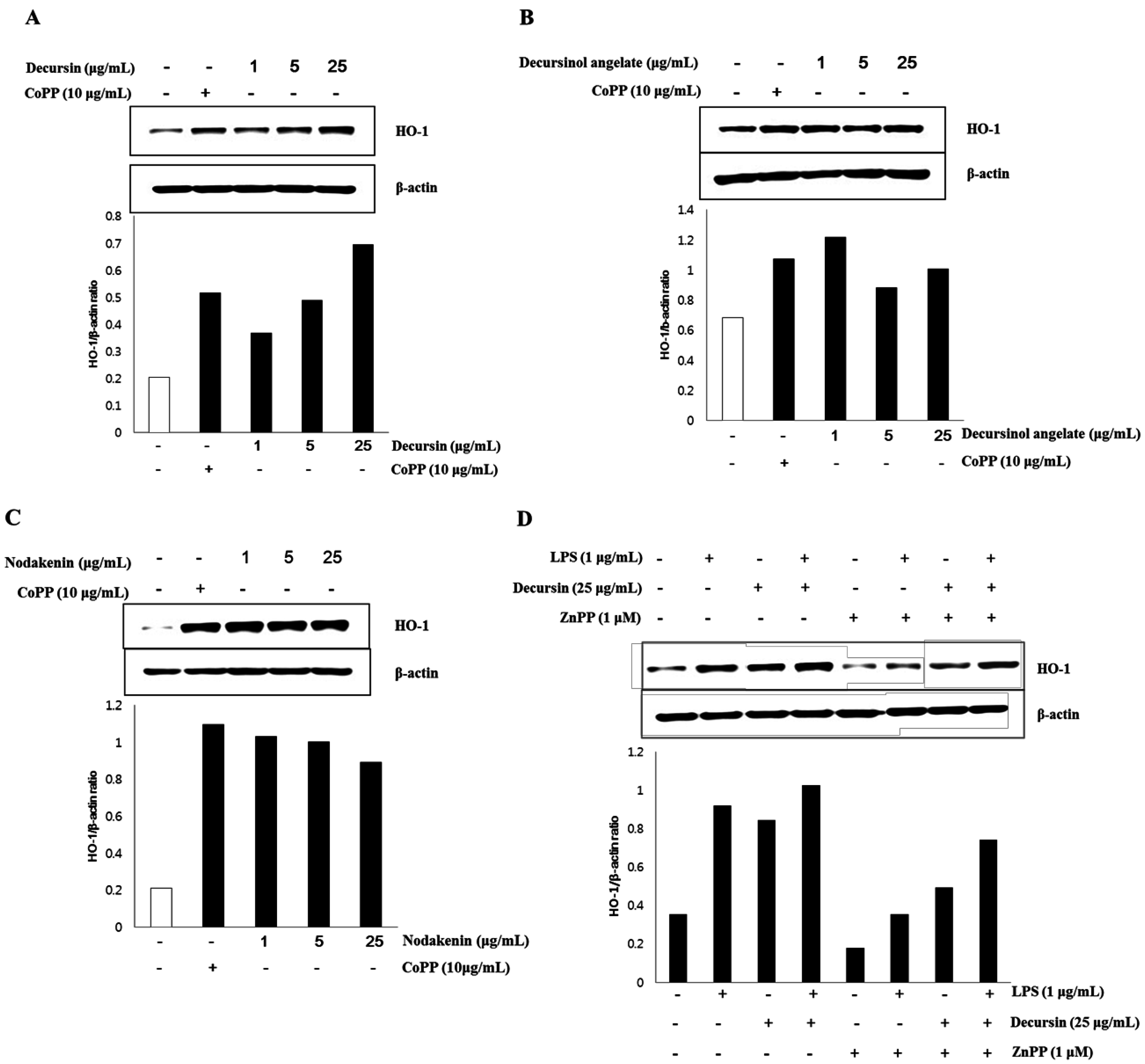
3.3. HO-1 Expression of Isolated Coumarins
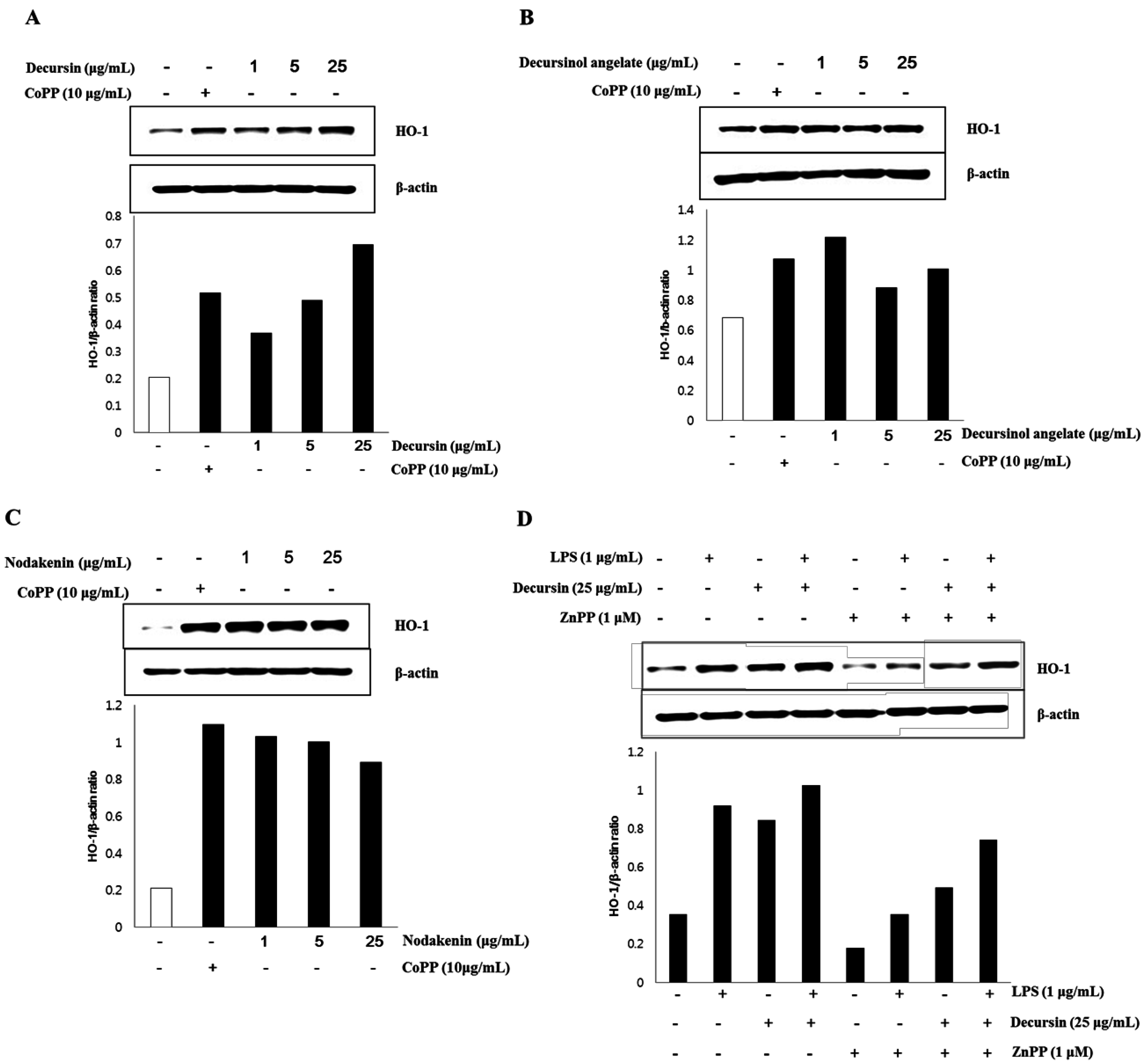
4. Discussions
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Son, C.Y.; Baek, I.H.; Song, G.Y.; Kang, J.S.; Kwon, K.I. Pharmacological effect of decursin and decursinol angelate from Angelica gigas Nakai. Yakhak Hoeji 2009, 53, 303–313. [Google Scholar]
- Son, S.H.; Park, K.K.; Park, S.K.; Kim, Y.C.; Kim, Y.S.; Lee, S.K.; Chung, W.Y. Decursin and decursinol from Angelica gigas inhibit the lung metastasis of murine colon carcinoma. Phytother. Res. 2011, 25, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, S.S.; Shin, K.H. Coumarins and a pyrimidine from Angelica gigas roots. Nat. Prod. Sci. 2002, 8, 58–61. [Google Scholar]
- Chi, H.J.; Kim, H.S. Studies on essential oils of plants of Angelica genus in Korea (I) essential oils of Angelicae gigantis radix. Korean J. Pharmacogn. 1988, 19, 239. [Google Scholar]
- Choi, Y.E.; Ahn, H.; Ryu, J.H. Polyacetylenes from Angelica gigas and their inhibitory activity on nitric oxide synthesis in activated macrophages. Biol. Pharm. Bull. 2000, 23, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.S.; Sim, W.S.; Lee, I.K.; Seu, Y.B.; Kim, I.H. Decursinol angelate: A cytotoxic and protein kinase C activating agent from the root of Angelica gigas. Planta Med. 1997, 63, 360–361. [Google Scholar] [CrossRef] [PubMed]
- Hata, K.; Sano, K. The constitution of decursin, a new coumarin isolated from the root of fr. et sav. (umbelliferae). Tetrahedron Lett. 1966, 7, 1461–1465. [Google Scholar] [CrossRef]
- Ng, T.B.; Ling, J.M.; Wang, Z.T.; Cai, J.N.; Xu, G.J. Examination of coumarins, flavonoid and polysaccharopeptide for antibacterial activity. Gen. Pharmacol. 1996, 27, 1237–1240. [Google Scholar] [CrossRef]
- Rehman, S.U.; Chohan, Z.H.; Gulnaz, F.; Supuran, C.T. In-vitro antibacterial, antifungal and cytotoxic activities of some coumarins and their metal complexes. J. Enzym. Inhib. Med. Chem. 2005, 20, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.R.; Lee, J.H.; Kim, J.Y.; Park, K.W.; Jeong, I.Y.; Shim, K.H.; Lee, M.K.; Seo, K.I. Decursin from Angelica gigas Nakai induces apoptosis in RC-58T/h/SA#4 primary human prostate cancer cells via a mitochondria-related caspase pathway. Food Chem. Toxicol. 2011, 49, 2517–2523. [Google Scholar] [PubMed]
- Yim, N.H.; Lee, J.H.; Cho, W.K.; Yang, M.C.; Kwak, D.H.; Ma, J.Y. Decursin and decursinol angelate from Angelica gigas Nakai induce apoptosis via induction of TRAIL expression on cervical cancer cells. Eur. J. Integr. Med. 2011, 3, 299–307. [Google Scholar] [CrossRef]
- Lee, S.; Lee, Y.S.; Jung, S.H.; Shin, K.H.; Kim, B.K.; Kang, S.S. Anti-tumor activities of decursinol angelate and decursin from Angelica gigas. Arch. Pharm. Res. 2003, 26, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Du, J.K.; Zou, L.Y.; Wu, T.; Lee, Y.W.; Kim, Y.H. Decursin isolated from Angelica gigas Nakai rescues PC12 Cells from amyloid beta-protein-induced neurotoxicity through Nrf2-mediated upregulation of heme oxygenase-1: Potential roles of MAPK. Evid. Based Complement. Altern. Med. 2013, 2013, 467245. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Kim, D.H.; Moon, Y.S.; Jung, J.S.; Ahn, E.M.; Baek, N.I.; Song, D.K. Protection against β-amyloid peptide-induced memory impairment with long-term administration of extract of Angelica gigas or decursinol in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2004, 28, 25–30. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Lee, S.; Jin, J.L.; Yun-Choi, H.S. Platelet anti-aggregatory effects of coumarins from the roots of Angelica genuflexa and A. gigas. Arch. Pharm. Res. 2003, 26, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Liew, F.Y. Regulation of nitric oxide synthesis in infectious and autoimmune diseases. Immunol. Lett. 1994, 43, 95–98. [Google Scholar] [CrossRef]
- Yang, A.; Wang, S.; Zong, H.; Zheng, G.; Chen, H.; Tao, J.; Tao, Y. Anti-inflammatory effects of ethanol extract from Melilotus suaveolens Ledeb: Involvement of pro- and anti-inflammatory cytokines and mediators. J. Med. Plants Res. 2012, 6, 516–525. [Google Scholar] [CrossRef]
- Clancy, R.M.; Leszczynska-Piziak, J.; Abramson, S.B. Nitric oxide, an endothelial cell relaxation factor, inhibits neutrophil superoxide anion production via a direct action on the NADPH oxidase. J. Clin. Investig. 1992, 90, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Torres, A.; Jones-Carson, J.; Mastroeni, P.; Ischiropoulos, H.; Fang, F.C. Antimicrobial actions of the NADPH phagocyte oxidase and inducible nitric oxide synthase in experimental salmonellosis. I. effects on microbial killing by activated peritoneal macrophages in vitro. J. Exp. Med. 2000, 192, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Jung, J.Y.; Jung, Y.J.; Choi, J.H.; Jeong, W.S.; Song, Y.S.; Kang, J.S.; Bi, K.; Kim, M.J. Anti-inflammatory activities of coumarins isolated from Angelica gigas Nakai on LPS-stimulated RAW 264.7 cell. J. Food Sci. Nutr. 2009, 14, 179–187. [Google Scholar] [CrossRef]
- Loscalzo, J.; Welch, G. Nitric oxide and its role in the cardiovascular system. Prog. Cardiovasc. Dis. 1995, 38, 87–104. [Google Scholar] [CrossRef]
- Marletta, M.A.; Yoon, P.S.; Iyengar, R.; Leaf, C.D.; Wishnok, J.S. Macrophage oxidation of l-arginine to nitrite and nitrate: Nitric oxide is an intermediate. Biochemistry 1988, 27, 8706–8711. [Google Scholar] [CrossRef] [PubMed]
- Motterlini, R.; Foresti, R.; Intaglietta, M.; Winslow, R.M. NO-mediated activation of heme oxygenase: Endogenous cytoprotection against oxidative stress to endothelium. Am. J. Physiol. 1996, 270, H107–H114. [Google Scholar] [PubMed]
- Park, J.Y.; Cho, H.Y.; Kim, J.K.; Noh, K.H.; Yang, J.R.; Ahn, J.M.; Lee, M.O.; Song, Y.S. Chlorella dichloromethane extract ameliorates NO production and iNOS expression through the down-regulation of NFB activity mediated by suppressed oxidative stress in RAW 264.7 macrophages. Clin. Chim. Acta 2005, 351, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef] [PubMed]
- Busse, R.; Mulsch, A. Induction of nitric oxide synthase by cytokines in vascular smooth muscle cells. FEBS Lett. 1990, 275, 87–90. [Google Scholar] [CrossRef]
- Choi, A.M.; Alam, J. Heme oxygenase-1: Function, regulation, and implication of a novel stress-inducible protein in oxidant-induced lung injury. Am. J. Respir. Cell Mol. Biol. 1996, 15, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Krzysiek-Maczka, G.; Targosz, A.; Ptak-Belowska, A.; Korbut, E.; Szczyrk, U.; Strzalka, M.; Brzozowski, T. Molecular alterations in fibroblasts exposed to helicobacter pylori: A missing link in bacterial inflammation progressing into gastric carcinogenesis? J. Physiol. Pharmacol. 2013, 64, 77–87. [Google Scholar] [PubMed]
- Kikuchi, G.; Yoshida, T.; Noguchi, M. Heme oxygenase and heme degradation. Biochem. Biophys. Res. Commun. 2005, 338, 558–567. [Google Scholar] [CrossRef] [PubMed]
- McCoubrey, W.K., Jr.; Huang, T.J.; Maines, M.D. Isolation and characterization of a cDNA from the rat brain that encodes hemoprotein heme oxygenase-3. Eur. J. Biochem. 1997, 247, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Durante, W.; Kroll, M.H.; Christodoulides, N.; Peyton, K.J.; Schafer, A.I. Nitric oxide induces heme oxygenase-1 gene expression and carbon monoxide production in vascular smooth muscle cells. Circ. Res. 1997, 80, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Maines, M.D. Heme oxygenase: Function, multiplicity, regulatory mechanisms, and clinical applications. FASEB J. 1988, 2, 2557–2568. [Google Scholar] [PubMed]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.K.; Lianos, E.A. Nitric oxide induces heme oxygenase-1 gene expression in mesangial cells. Kidney Int. 1999, 55, 1734–1739. [Google Scholar] [CrossRef] [PubMed]
- De Montellano, P.R.O. The mechanism of heme oxygenase. Curr. Opin. Chem. Biol. 2000, 4, 221–227. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeong, J.H.; Jeon, S.T.; Kim, H.; Ock, J.; Suk, K.; Kim, S.I.; Song, K.S.; Lee, W.H. Decursin inhibits induction of inflammatory mediators by blocking nuclear factor-kB activation in macrophages. Mol. Pharmacol. 2006, 69, 1783–1790. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, P.; Ferlazzo, V.; Milano, S.; La Rosa, M.; di Bella, G.; Caruso, R. Anti-inflammatory effects of chemically modified tetracyclines by the inhibition of nitric oxide and interleukin-12 synthesis in J774 cell line. Int. Immunopharmacol. 2001, 1, 1765–1776. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Teng, L.; Vu, D.; He, J.Q.; Guo, Y.; Li, Q.; Tang, X.L.; Rokosh, G.; Bhatnagar, A.; Bolli, R. The heme oxygenase 1 inducer (CoPP) protects human cardiac stem cells against apoptosis through activation of the extracellular signal-regulated kinase (ERK)/NRF2 signaling pathway and cytokine release. J. Biol. Chem. 2012, 287, 33720–33732. [Google Scholar] [CrossRef] [PubMed]
- Rim, H.K.; Cho, W.; Sung, S.H.; Lee, K.T. Nodakenin suppresses lipopolysaccharide-induced inflammatory responses in macrophage cells by inhibiting tumor necrosis factor receptor-associated factor 6 and nuclear factor-κB pathways and protects mice from lethal endotoxin shock. J. Pharmacol. Exp. Ther. 2012, 342, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Jang, F.; Jiang, R.; Zhu, X.; Zhang, X.; Zhan, Z. Genipin inhibits TNF-α-induced vascular smooth muscle cell proliferation and migration via induction of HO-1. PLoS ONE 2013, 8, e74826. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.R. Oxidative stress in the brain and arterial hypertension. Hypertens. Res. 2009, 32, 1047–1048. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Kang, Y.J.; Lee, K.Y.; Choi, H.C. Isoproterenol inhibits angiotensin II-stimulated proliferation and reactive oxygen species production in vascular smooth muscle cells through heme oxygenase-1. Biol. Pharm. Bull. 2009, 32, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- Ndisang, J.F. Role of heme oxygenase in inflammation, insulin-signalling, diabetes and obesity. Mediat. Inflamm. 2010, 359732. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Henning, S.M.; Heber, D. Limitations of MTT and MTS-based assays for measurement of anti-proliferative activity of green tea polyphenols. PLoS ONE 2010, 5, e10202. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, J.H.; Kwon, J.E.; Cho, Y.; Kim, I.; Kang, S.C. Anti-Inflammatory Effect of Angelica gigas via Heme Oxygenase (HO)-1 Expression. Nutrients 2015, 7, 4862-4874. https://doi.org/10.3390/nu7064862
Cho JH, Kwon JE, Cho Y, Kim I, Kang SC. Anti-Inflammatory Effect of Angelica gigas via Heme Oxygenase (HO)-1 Expression. Nutrients. 2015; 7(6):4862-4874. https://doi.org/10.3390/nu7064862
Chicago/Turabian StyleCho, Joon Hyeong, Jung Eun Kwon, Youngmi Cho, Inhye Kim, and Se Chan Kang. 2015. "Anti-Inflammatory Effect of Angelica gigas via Heme Oxygenase (HO)-1 Expression" Nutrients 7, no. 6: 4862-4874. https://doi.org/10.3390/nu7064862
APA StyleCho, J. H., Kwon, J. E., Cho, Y., Kim, I., & Kang, S. C. (2015). Anti-Inflammatory Effect of Angelica gigas via Heme Oxygenase (HO)-1 Expression. Nutrients, 7(6), 4862-4874. https://doi.org/10.3390/nu7064862