Anticancer Effect of Ursodeoxycholic Acid in Human Oral Squamous Carcinoma HSC-3 Cells through the Caspases

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparations of Ursodeoxycholic Acid (UDCA)
2.2. Cell Preparation
2.3. 3-(4,5-Dimethyl-2-Thiazolyl)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
2.4. Nuclear Staining with 4’,6-Diamidino-2-Phenylindole (DAPI)
2.5. Flow Cytometry Analysis
2.6. RT-PCR Assay

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence |
|---|---|
| Caspase-3 | Forward: 5′-CAA ACT TTT TCA GAG GGG ATC G-3′ |
| Reverse: 5′-GCA TAC TGT TTC AGC ATG GCA-3′ | |
| Caspase-8 | Forward: 5′-CCC CAC CCT CAC TTT GCT-3′ |
| Reverse: 5′-GGA GGA CCA GGC TCA CTT A-3′ | |
| Caspase-9 | Forward: 5′-GGC CCT TCC TCG CTT CAT CTC-3′ |
| Reverse: 5′-GGT CCT TGG GCC TTC CTG GTA T-3′ | |
| Bax | Forward: 5′-AAG CTG AGC GAG TGT CTC CGG CG-3′ |
| Reverse: 5′-CAG ATG CCG GTT CAG GTA CTC AGT C-3′ | |
| Bcl-2 | Forward: 5′-CTC GTC GCT ACC GTC GTG ACT TGG-3′ |
| Reverse: 5′-CAG ATG CCG GTT CAG GTA CTC AGT C-3′ | |
| Bcl-xL | Forward: 5′-CCC AGA AAG GAT ACA GCT GG-3′ |
| Reverse: 5′-GCG ATC CGA CTC ACC AAT AC-3′ | |
| XIAP | Forward: 5′-CCG TGC GGT TGC TTT AGT TGT C-3′ |
| Reverse: 5′-ATG GCA GGG TTC CTC GGG TAT-3′ | |
| cIAP-1 | Forward: 5′-TGAGCATGCAGACACATGC-3′ |
| Reverse: 5′-TGACGGATGAACTCCTGTCC-3′ | |
| cIAP-2 | Forward: 5′-AATGGAAGATAGCACGAT-3′ |
| Reverse: 5′-AGAAAGGCTGGAGTAAGA-3′ | |
| Survivin | Forward: 5′-CTT TCT CAA GGC CCA CCG CAT CT-3′ |
| Reverse: 5′-GCA CTT TCT CCG CAG TTT CCT C-3′ | |
| Fas | Forward: 5′-GAA ATG AAA TCC AAA GCT-3′ |
| Reverse: 5′-TAA TTT AGA GGC AAA GTG GC-3′ | |
| FasL | Forward: 5′-GGA TTG GGC CTG GGG ATG TTT CA-3′ |
| Reverse: 5′-TTG TGG CTC AGG GGC AGG TTG TTG-3′ | |
| TRAIL | Forward: 5′-GGA ACC CAA GGT GGG TAG AT-3′ |
| Reverse: 5′-TCT CAC CAC ACT GCA ACC TC-3′ | |
| DR4 | Forward: 5′-AAG TCC CTG CAC CAC GAC-3′ |
| Reverse: 5′-CCA CAA CCT GAG CCG ATG-3′ | |
| DR5 | Forward: 5′-TGA GAT AAA GGT GGC TAA A -3′ |
| Reverse: 5′-AAA GGT AAA CCA GGG AAG -3′ | |
| NF-κB | Forward: 5′-CAC TTA TGG ACA ACT ATG AGG TCT CTG G-3′ |
| Reverse: 5′-CTG TCT TGT GGA CAA CGC AGT GGA ATT TTA GG-3′ | |
| IκB-α | Forward: 5′-GCT GAA GAA GGA GCG GCT ACT-3′ |
| Reverse: 5′-TCG TAC TCC TCG TCT TTC ATG GA-3′ | |
| GAPDH | Forward: 5′-CGG AGT CAA CGG ATT TGG TC-3′ |
| Reverse: 5′-AGC CTT CTC CAT GGT CGT GA-3′ |
2.7. Western Blot Assay
2.8. Statistical Analysis
3. Results
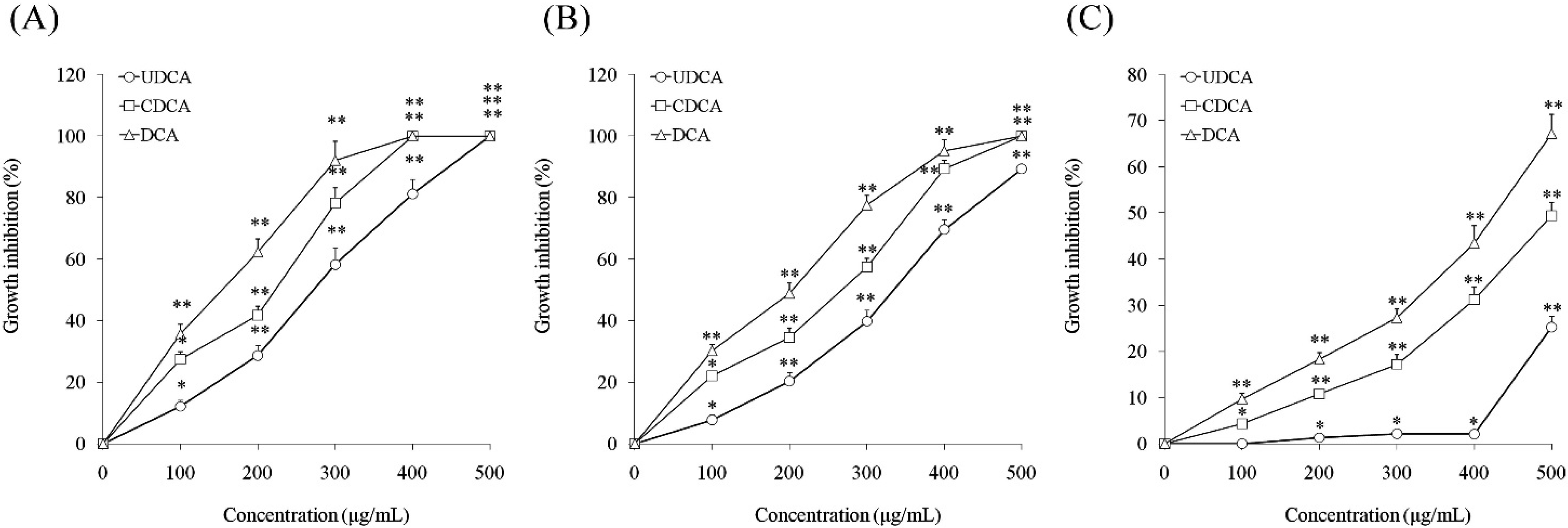
3.1. Inhibitory Effects of Ursodeoxycholic Acid (UDCA), Chenodeoxycholic Acid (CDCA) and Deoxycholic Acid (DCA) on Cell Growth
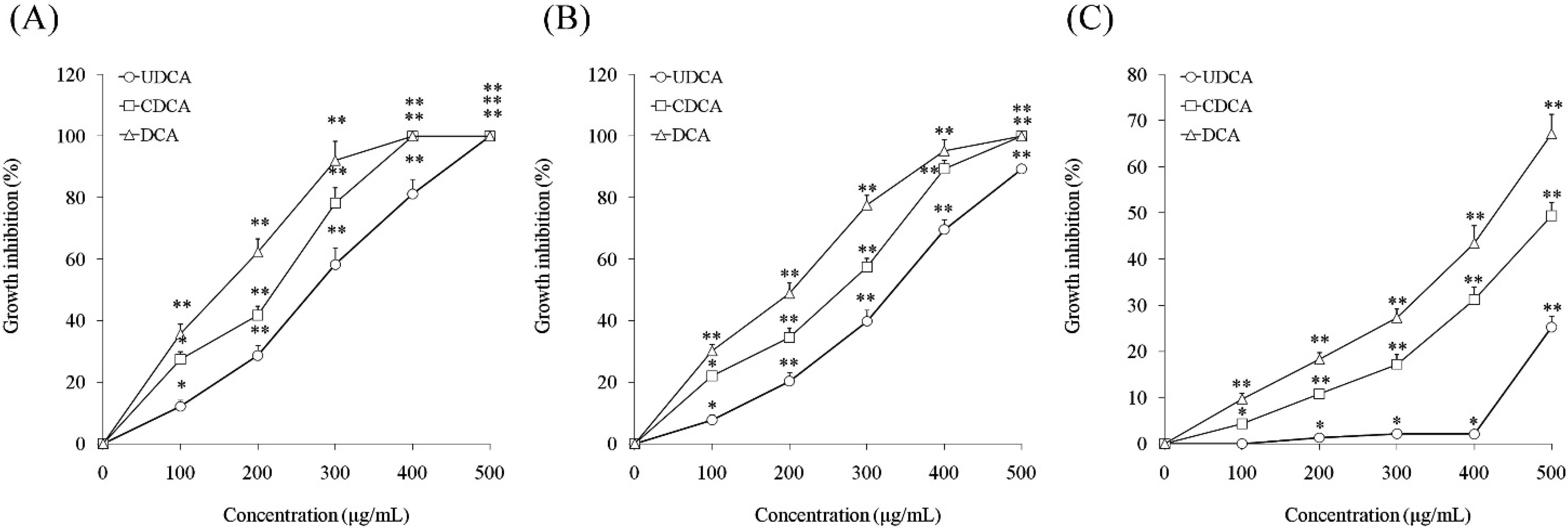
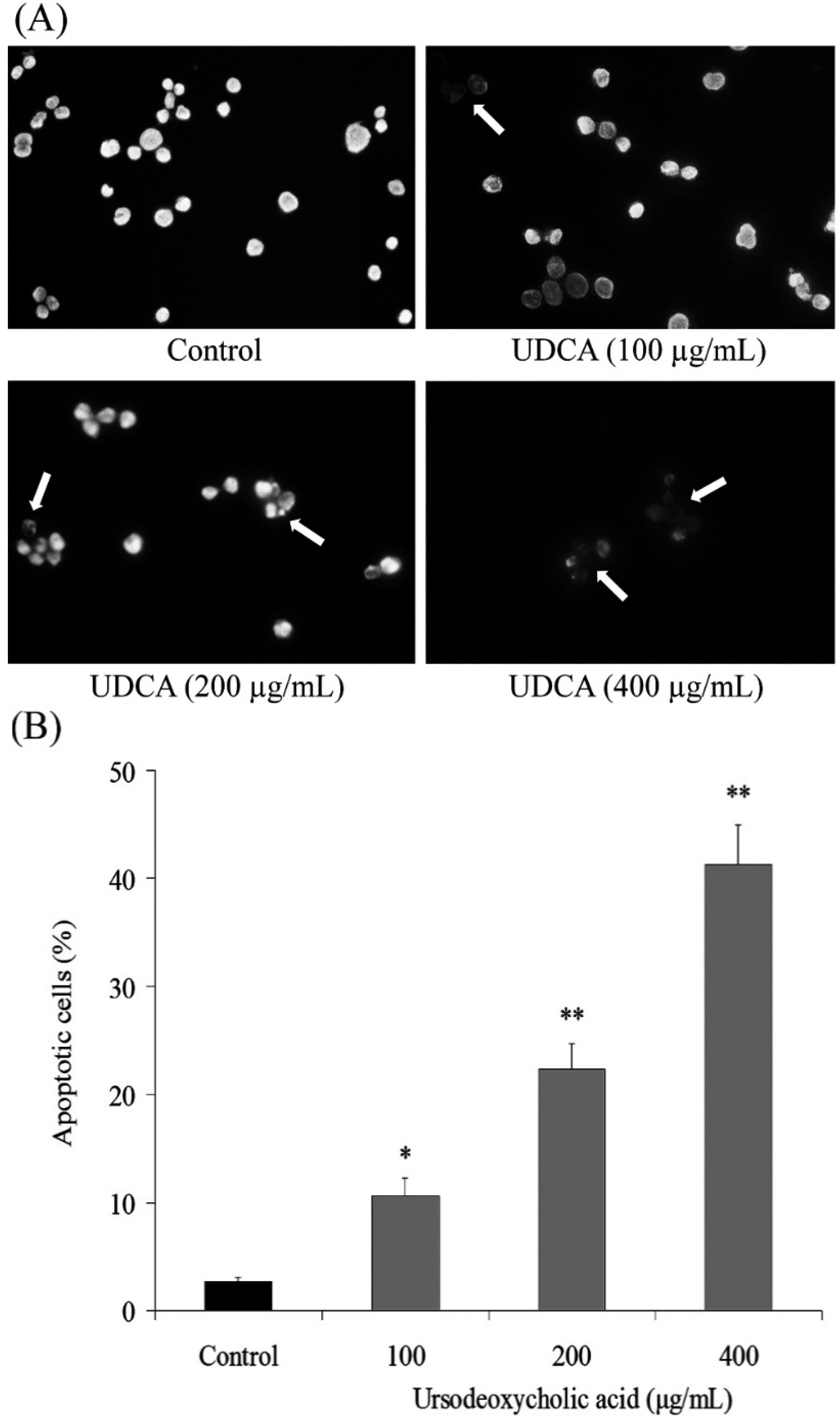
3.2. Induction of Apoptosis by UDCA

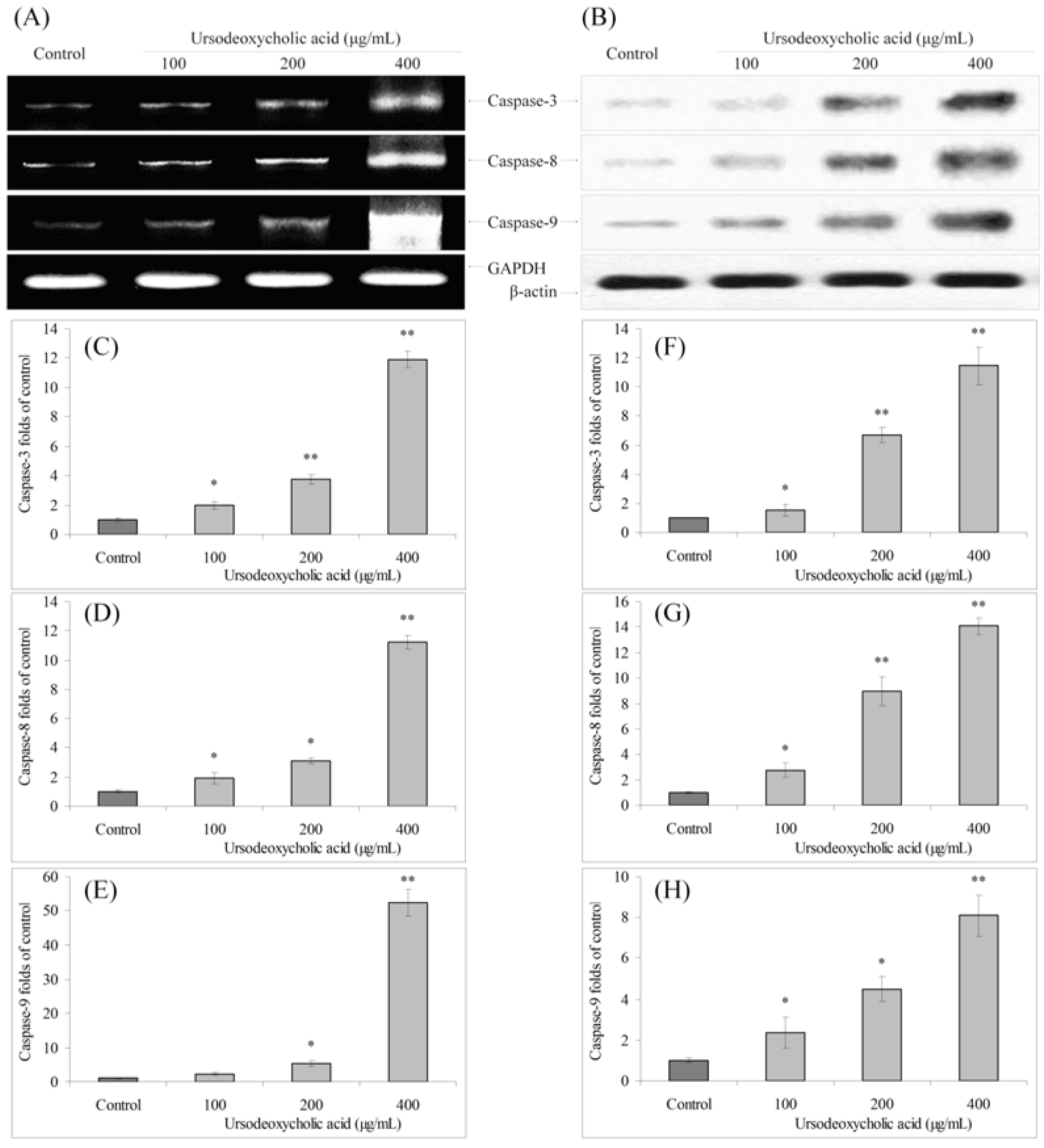
3.3. Gene Expression of Caspase Family
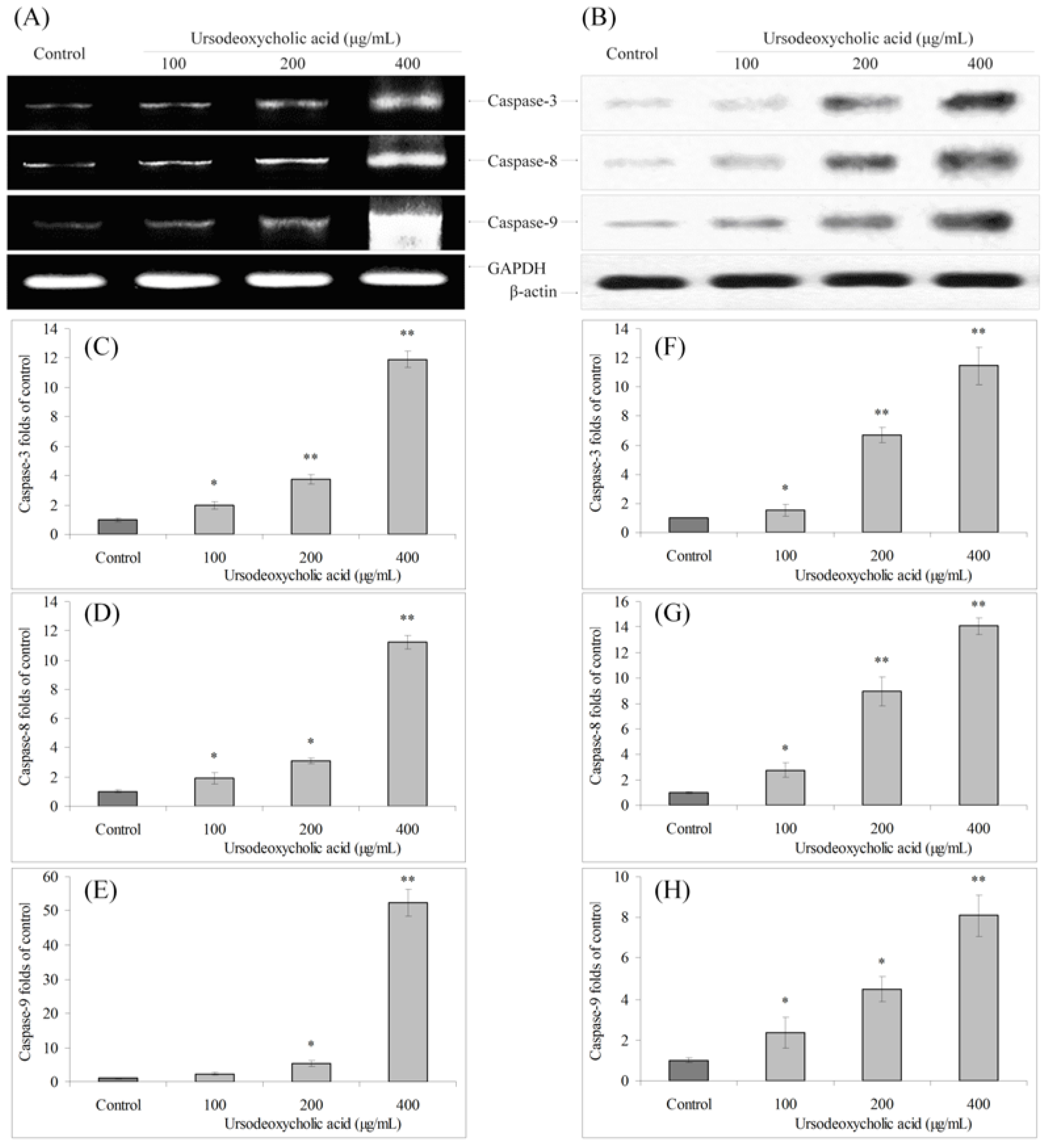
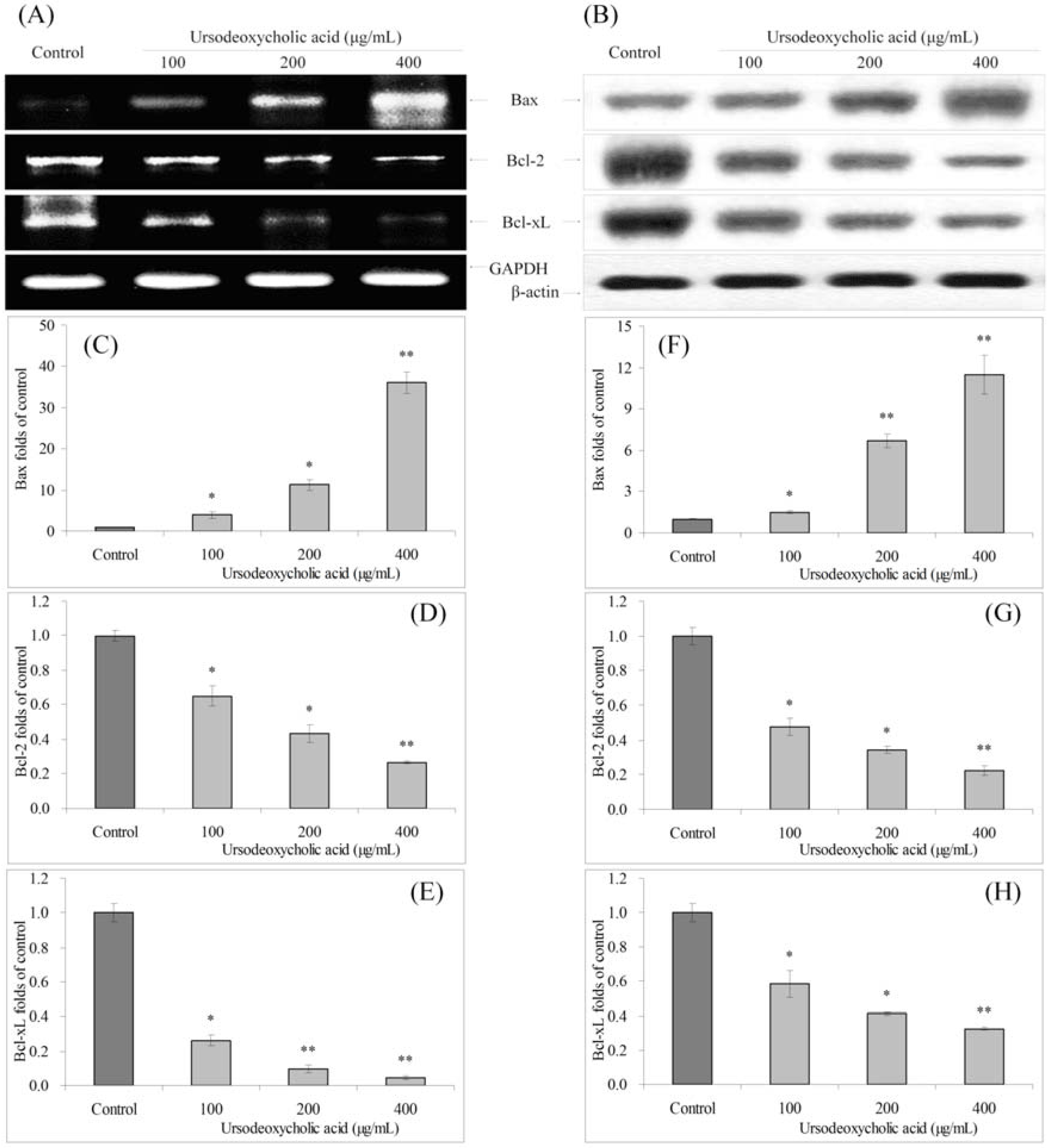
3.4. Gene Expression of Bcl-2 Family
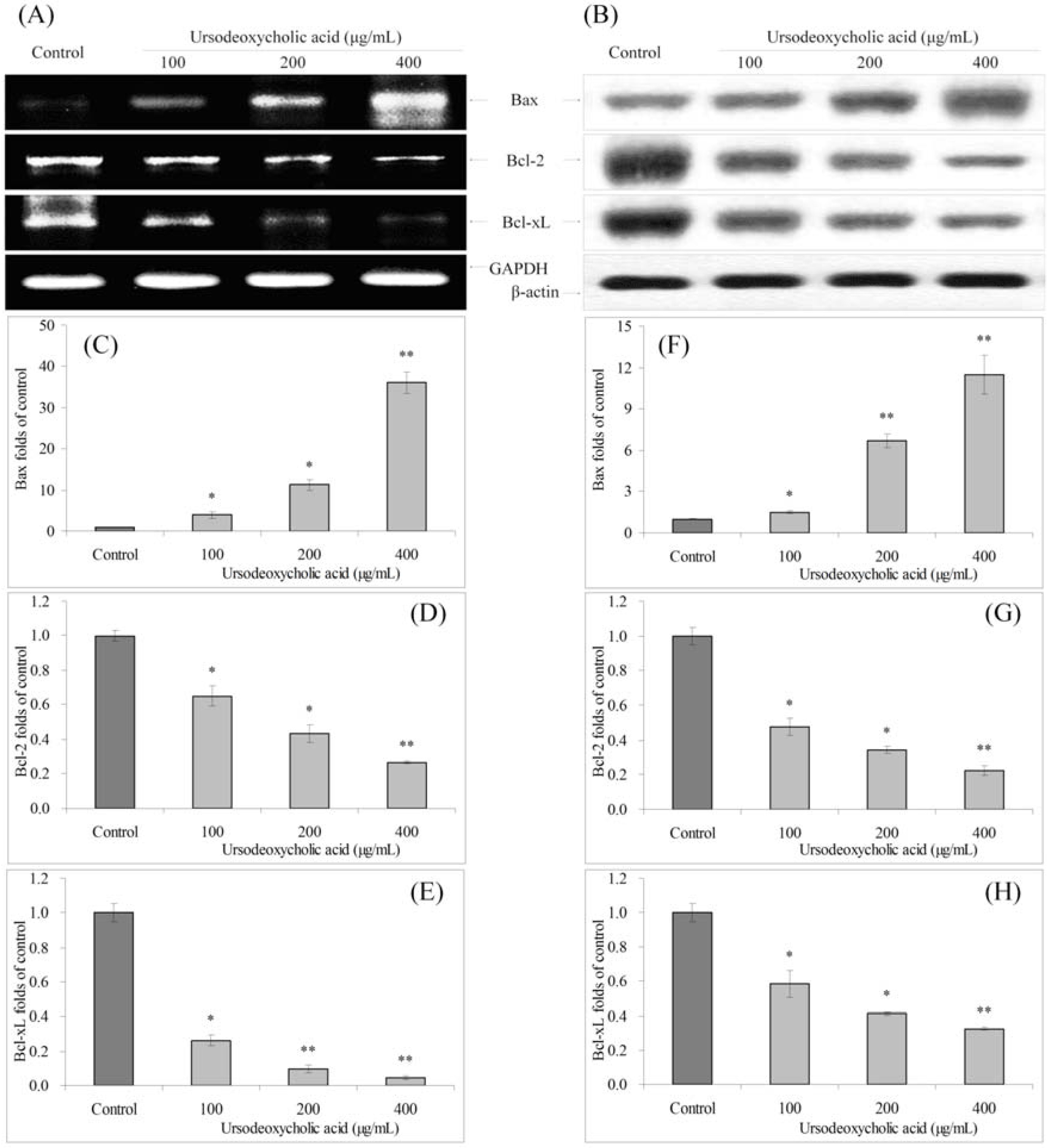
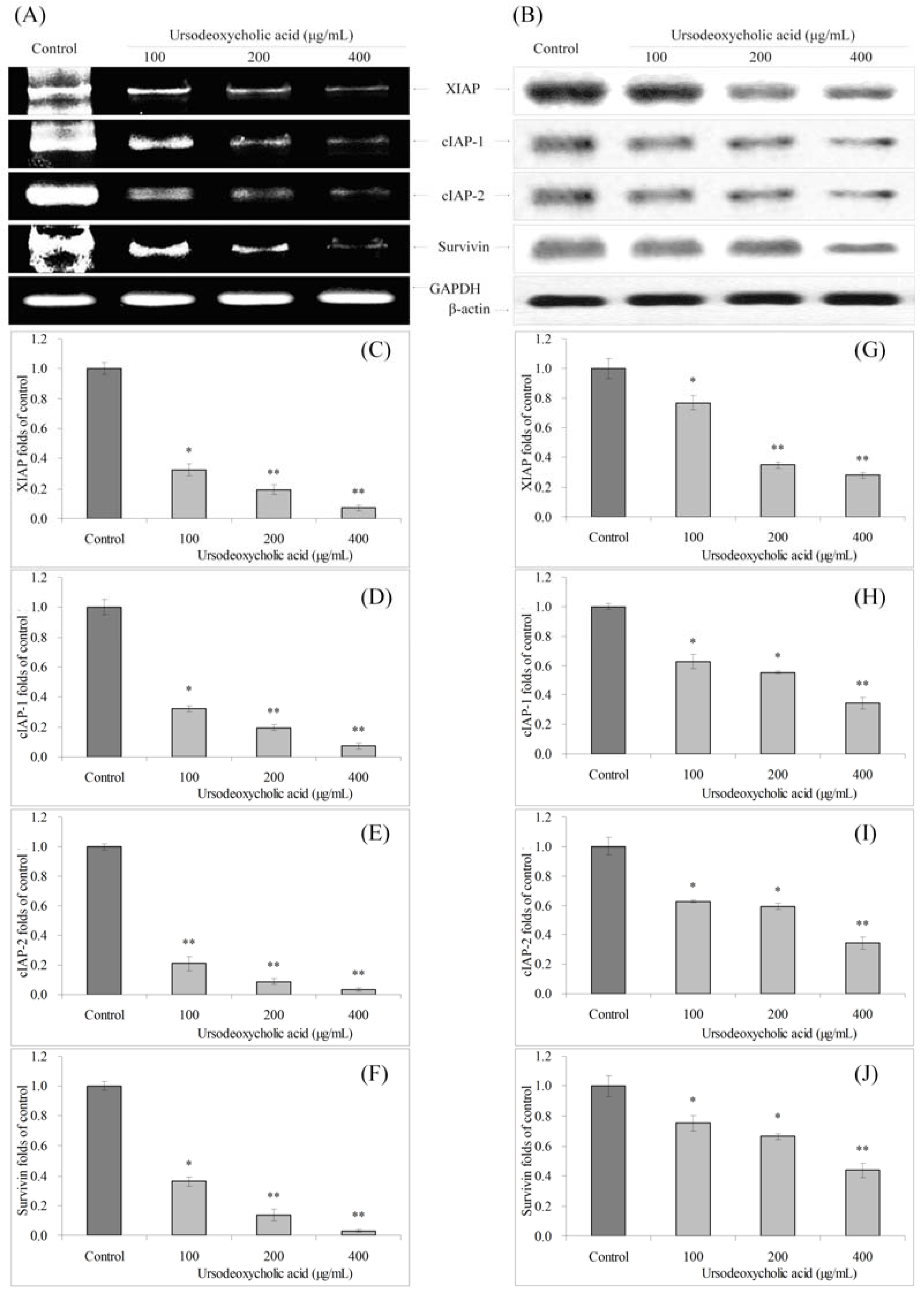
3.5. Gene Expression of XIAP, cIAP-1, cIAP-2 and Survivin
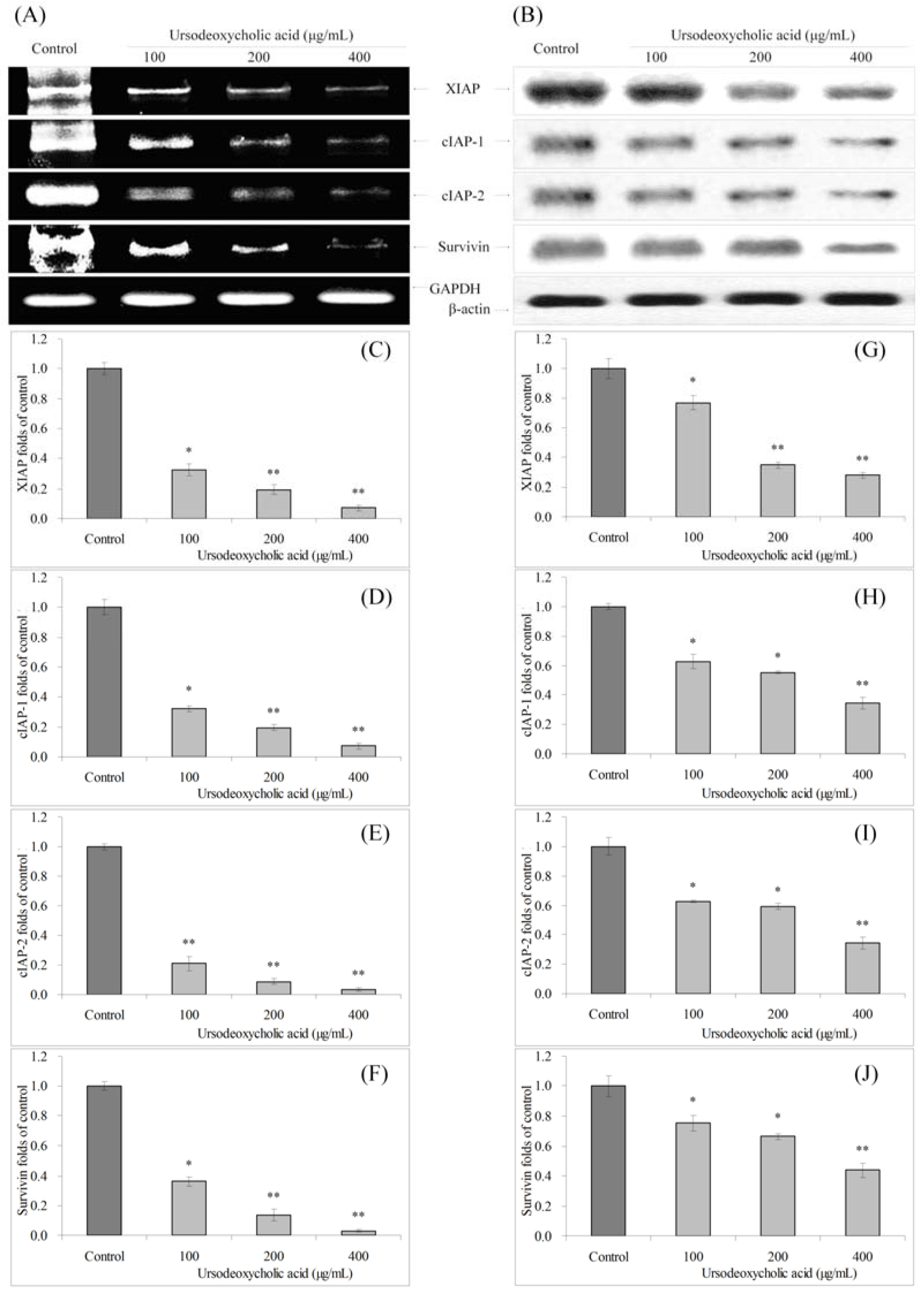
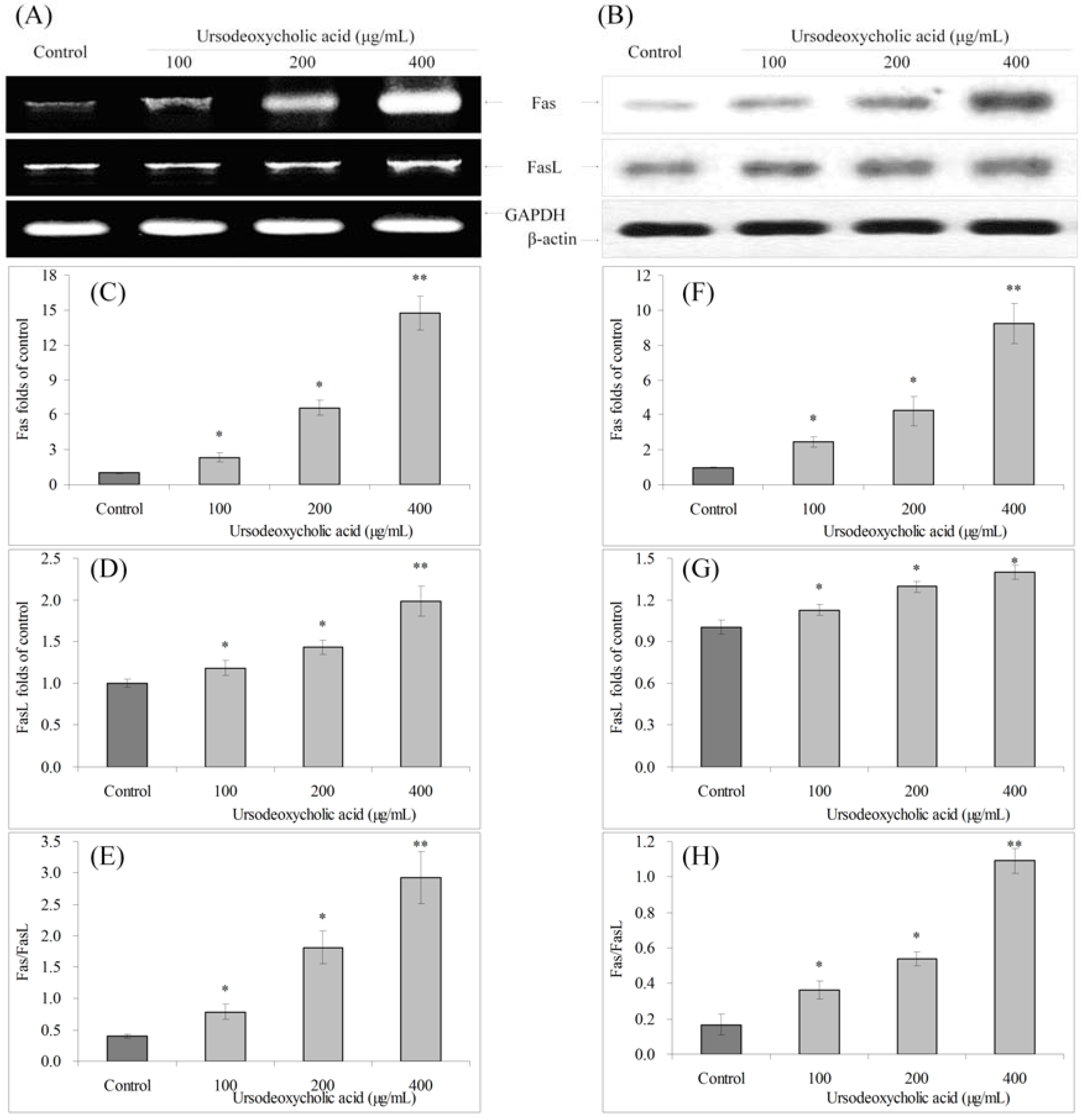
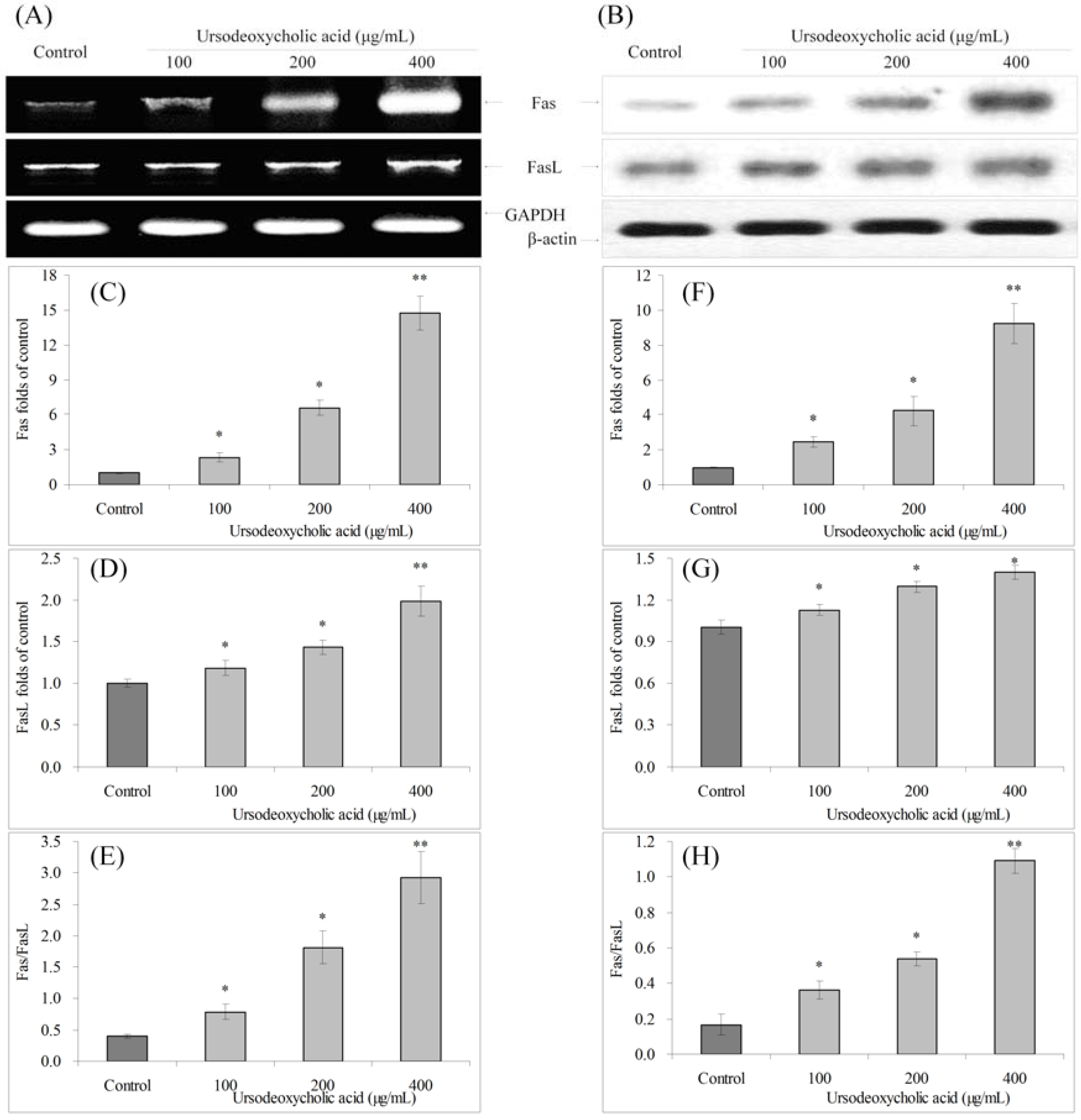
3.6. Gene Expression of Fas and FasL
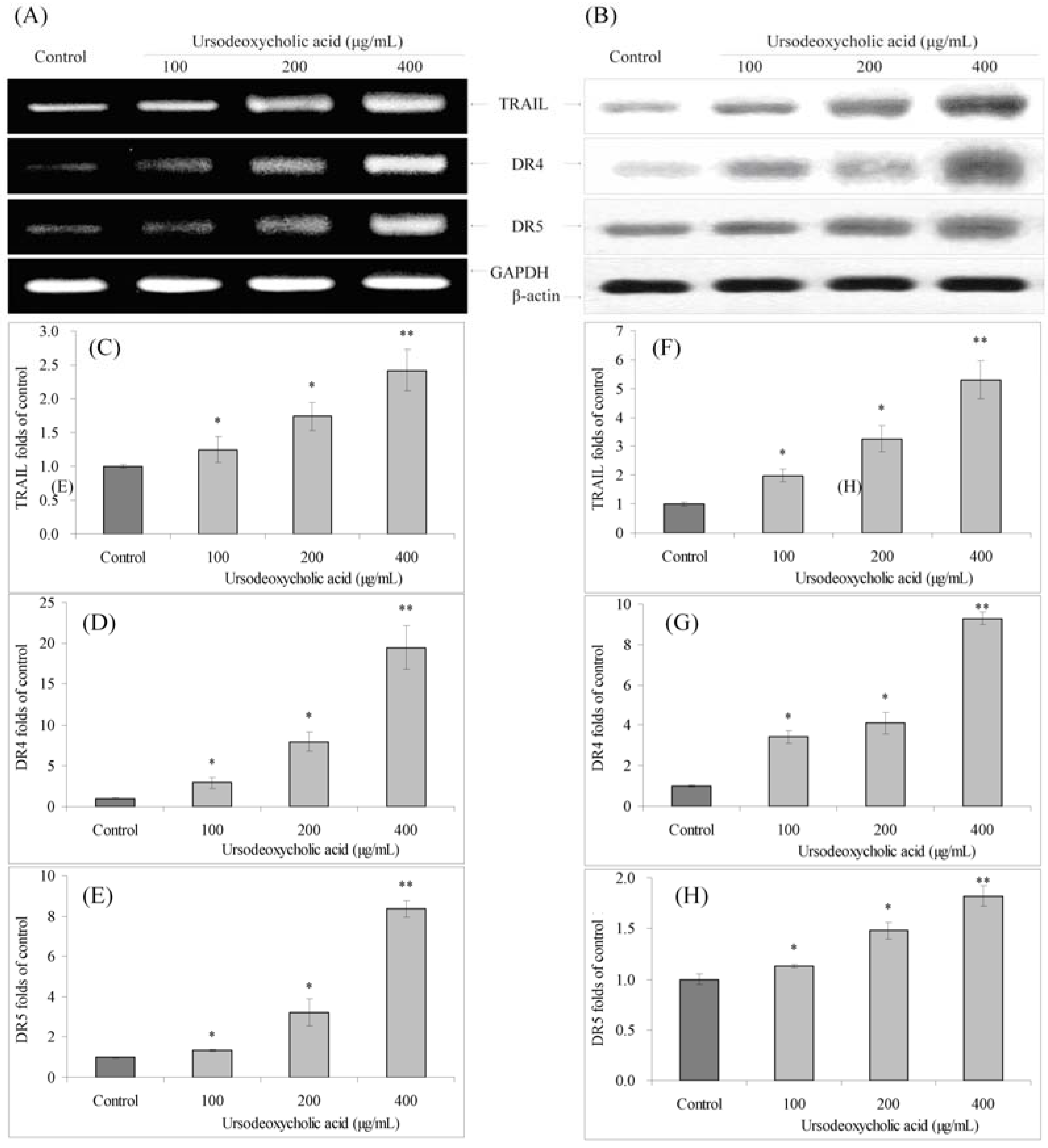
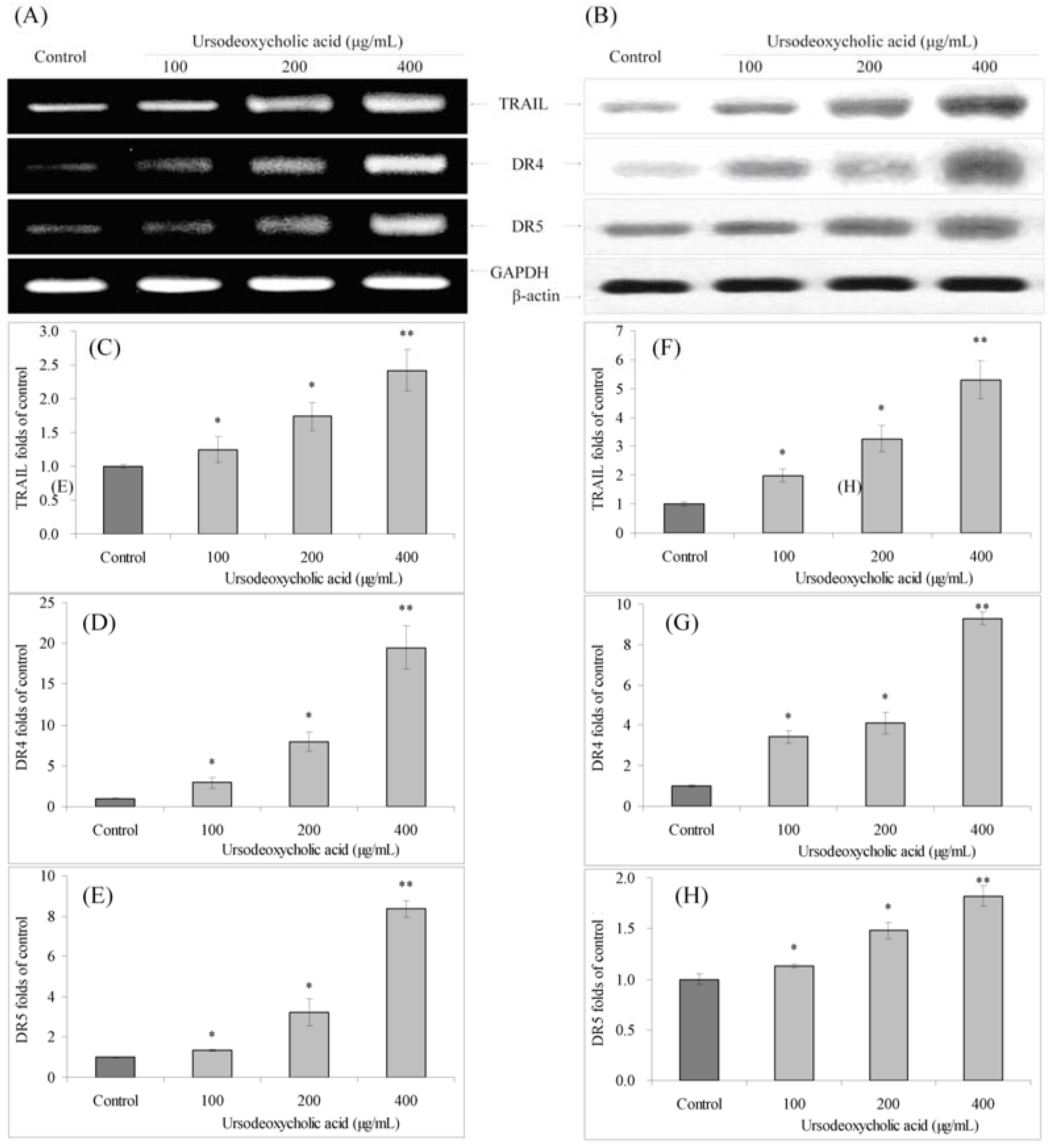
3.7. Gene Expression of TRAIL, DR4 and DR5
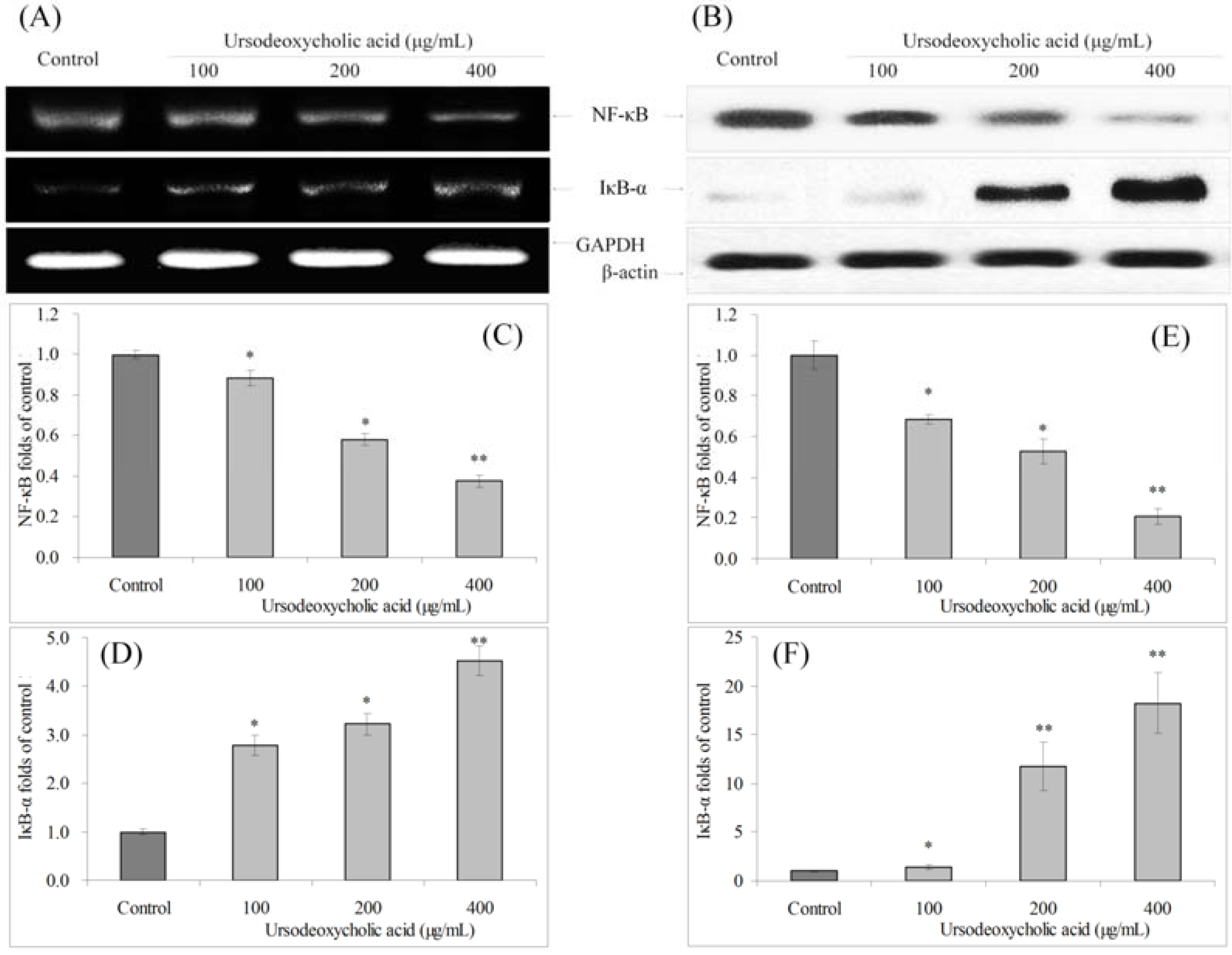
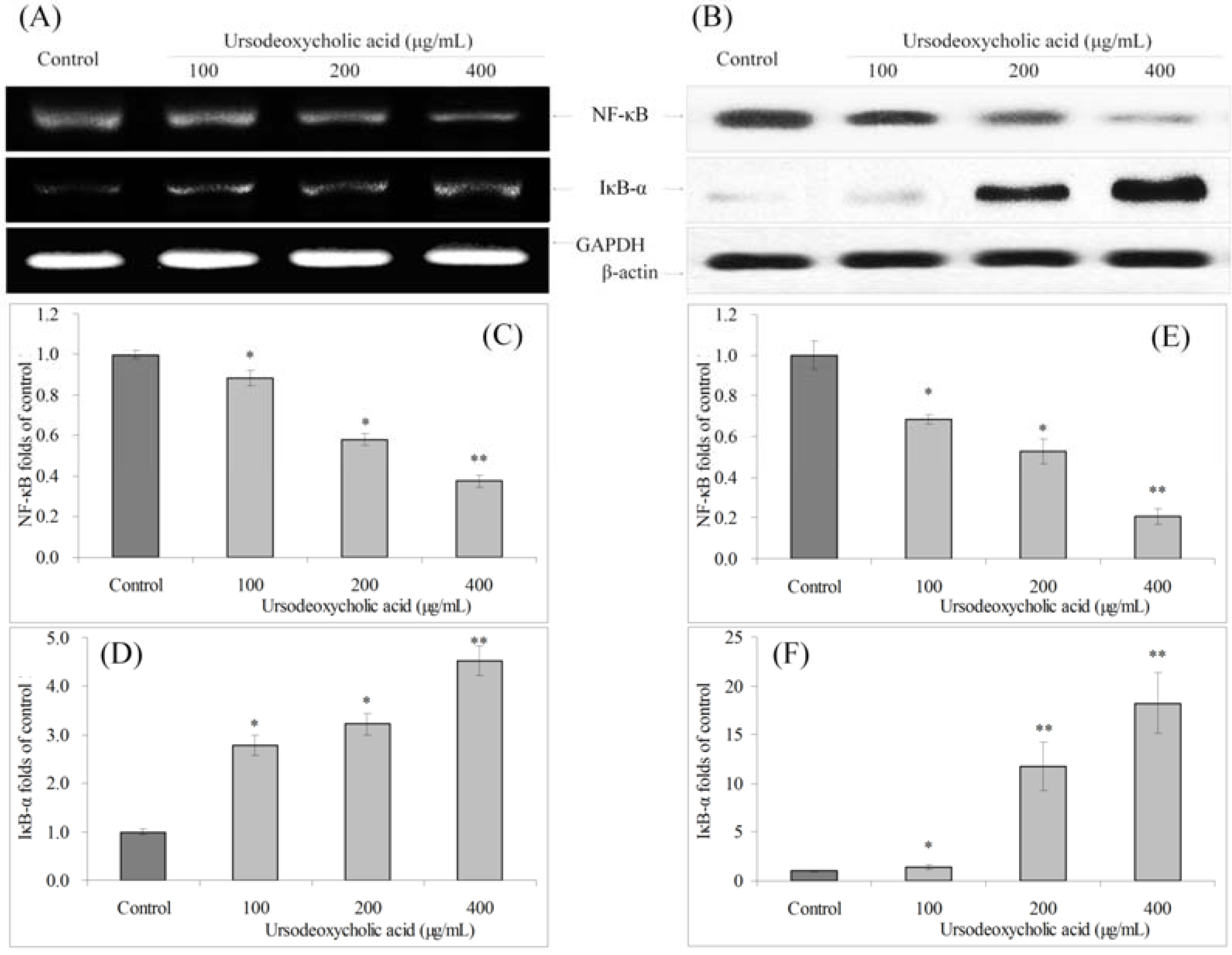
3.8. Gene Expression of NF-κB and IκB-α
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chi, C.G. Synthetic and pharmacological research progress of primary chemical components in bear bile. Strait. Pharm. J. 2008, 20, 5–8. [Google Scholar]
- Lee, S.H.; Cheon, G.J.; Kim, H.S.; Kim, Y.D.; Kim, S.G.; Kim, Y.S.; Jeong, S.W.; Jang, J.Y.; Kim, B.S. Comparison on the efficacy and safety of biphenyl dimethyl dicarboxylate and ursodeoxycholic acid in patients with abnormal alanine aminotransferase: Multicenter, double-blinded, randomized, active-controlled clinical trial. Korean J. Gastroenterol. 2014, 64, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, A.K.; Engels, E.A.; Anderson, W.F.; Gillison, M.L. Incidence trends for human papillomavirus-related and -unrelated oral squamous cell carcinomas in the United States. J. Clin. Oncol. 2008, 26, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Michaud, D.S.; Izard, J.; Rubin, Z.; Johansson, I.; Weiderpass, E.; Tjønneland, A.; Olsen, A.; Overvad, K.; Boutron-Ruault, M.C.; Clavel-Chapelon, F.; et al. Lifestyle, dietary factors, and antibody levels to oral bacteria in cancer-free participants of a European cohort study. Cancer Causes Control 2013, 24, 1901–1909. [Google Scholar] [CrossRef] [PubMed]
- Tanis, T.; Cincin, Z.B.; Gokcen-Rohlig, B.; Bireller, E.S.; Ulusan, M.; Tanyel, C.R.; Cakmakoglu, B. The role of components of the extracellular matrix and inflammation on oral squamous cell carcinoma metastasis. Arch. Oral Biol. 2014, 59, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Li, G.J.; Sun, P.; Wang, R.; Qian, Y.; Zhao, X. In vitro and in vivo anti-cancer activities of Kuding tea (Ilex kudingcha C.J. Tseng) against oral cancer. Exp. Ther. Med. 2014, 7, 709–715. [Google Scholar] [PubMed]
- Zhao, X.; Wang, Q.; Li, G.J.; Chen, F.; Qian, Y.; Wang, R. In vitro antioxidant, anti-mutagenic, anti-cancer and anti-angiogenic effects of Chinese Bowl tea. J. Func. Food. 2014, 7, 590–598. [Google Scholar] [CrossRef]
- Zhao, X.; Deng, X.X.; Park, K.Y.; Qiu, L.H.; Pang, L. Purple bamboo salt has anticancer activity in TCA8113 cells in vitro and preventive effects on buccal mucosa cancer in mice in vivo. Exp. Ther. Med. 2013, 5, 549–554. [Google Scholar] [PubMed]
- Zhao, X.; Wang, Q.; Qian, Y.; Pang, L. Cassia tora L. (Jue-ming-zi) has anticancer activity in TCA8113 cells in vitro and exerts anti-metastatic effects in vivo. Oncol. Lett. 2013, 5, 1036–1042. [Google Scholar] [PubMed]
- Zhao, X.; Ju, J.H.; Kim, H.M.; Park, K.Y. Antimutagenic activity and in vitro anticancer effects of bamboo salt on HepG2 human hepatoma cells. J. Environ. Pathol. Toxicol. Oncol. 2013, 32, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Pang, L.; Li, J.; Song, J.L.; Qiu, L.H. Apoptosis inducing effects of Kuding tea polyphenols in human buccal squamous cell carcinoma cell line BcaCD885. Nutrients 2014, 6, 3084–3100. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Kim, S.Y.; Park, K.Y. Bamboo salt has in vitro anticancer activity in HCT-116 cells and exerts anti-metastatic effects in vivo. J. Med. Food 2013, 16, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Matsumiya, T.; Hayakari, R.; Narita, N.; Ito, R.; Kon, T.; Kubota, K.; Sakaki, H.; Yoshida, H.; Imaizumi, T.; Kobayashi, W.; et al. Role of type I- and type II-interferon in expression of melanoma differentiation-associated gene-5 in HSC-3 oral squamous carcinoma cells. Biomed. Res. 2014, 35, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Contreras, R.; Scougall-Vilchis, R.J.; Contreras-Bulnes, R.; Kanda, Y.; Nakajima, H.; Sakagami, H. Effects of TiO2 nano glass ionomer cements against normal and cancer oral cells. Vivo 2014, 28, 895–907. [Google Scholar]
- Yamamura, T.; Matsumoto, N.; Matsue, Y.; Okudera, M.; Nishikawa, Y.; Abiko, Y.; Komiyama, K. Sodium butyrate, a histone deacetylase inhibitor, regulates lymphangiogenic factors in oral cancer cell line HSC-3. Anticancer Res. 2014, 34, 1701–1708. [Google Scholar] [PubMed]
- Baddela, V.S.; Baufeld, A.; Yenuganti, V.R.; Vanselow, J.; Singh, D. Suitable housekeeping genes for normalization of transcript abundance analysis by real-time RT-PCR in cultured bovine granulosa cells during hypoxia and differential cell plating density. Reprod. Biol. Endocrinol. 2014, 12. [Google Scholar] [CrossRef]
- Zhang, R.; Yang, D.; Zhou, C.; Cheng, K.; Liu, Z.; Chen, L.; Fang, L.; Xie, P. β-actin as a loading control for plasma-based Western blot analysis of major depressive disorder patients. Anal. Biochem. 2012, 427, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.W.; Julien, O.; Unger, E.K.; Shah, N.M.; Wells, J.A. Turning on caspases with genetics and small molecules. Methods Enzymol. 2014, 544, 179–213. [Google Scholar] [PubMed]
- Zhao, Y.; Lei, M.; Wang, Z.; Qiao, G.; Yang, T.; Zhang, J. TCR-induced, PKC-θ-mediated NF-κB activation is regulated by a caspase-8-caspase-9-caspase-3 cascade. Biochem. Biophys. Res. Commun. 2014, 450, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Tian, J.; Wu, H.; Liu, Y.; Ren, J.; Zheng, S.; Zhang, C.; Yang, C.; Li, Y.; Wang, S. Ginsenoside rb1 protects neonatal rat cardiomyocytes from hypoxia/ischemia induced apoptosis and inhibits activation of the mitochondrial apoptotic pathway. Evid. Based Complement. Alternat. Med. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Tsagarakis, N.J.; Drygiannakis, I.; Batistakis, A.G.; Kolios, G.; Kouroumalis, E.A. A concentration-dependent effect of ursodeoxycholate on apoptosis and caspases activities of HepG2 hepatocellular carcinoma cells. Eur. J. Pharmacol. 2010, 640, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Shan, L.J.; Liu, Y.J.; Chen, D.; Xiao, X.G.; Li, Y. Ursodeoxycholic acid induces apoptosis of hepatocellular carcinoma cells in vitro. J. Dig. Dis. 2014, 15, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Braun, C.R.; Bird, G.H.; Walensky, L.D. Photoreactive stapled peptides to identify and characterize BCL-2 family interaction sites by mass spectrometry. Method. Enzymol. 2014, 544, 25–48. [Google Scholar]
- Choi, H.J.; Yee, S.B.; Park, S.E.; Im, E.; Jung, J.H.; Chung, H.Y.; Choi, Y.H.; Kim, N.D. Petrotetrayndiol A induces cell cycle arrest and apoptosis in SK-MEL-2 human melanoma cells through cytochrome c-mediated activation of caspases. Cancer Lett. 2006, 232, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Park, J.A.; Lee, K.Y.; Oh, Y.J.; Kim, K.W.; Lee, S.K. Activation of caspase-3 protease via a Bcl-2-insensitive pathway during the process of ginsenoside Rh2-induced apoptosis. Cancer Lett. 1997, 121, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Silke, J.; Vucic, D. IAP family of cell death and signaling regulators. Method. Enzymol. 2014, 545, 35–65. [Google Scholar]
- Hui, K.K.; Kanungo, A.K.; Elia, A.J.; Henderson, J.T. Caspase-3 deficiency reveals a physiologic role for Smac/DIABLO in regulating programmed cell death. Cell Death Differ. 2011, 18, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Shin, D.Y.; Lee, Y.W.; Cho, C.K.; Kim, G.Y.; Kim, W.J.; Yoo, H.S.; Choi, Y.H. Apoptotic and anti-metastatic effects of the whole skin of Venenum bufonis in A549 human lung cancer cells. Int. J. Oncol. 2012, 40, 1210–1219. [Google Scholar] [PubMed]
- Deng, L.; Lu, Y.; Zhao, X.; Sun, Y.; Shi, Y.; Fan, H.; Liu, C.; Zhou, J.; Nie, Y.; Wu, K.; Fan, D.; Guo, X. Ran GTPase protein promotes human pancreatic cancer proliferation by deregulating the expression of survivin and cell cycle proteins. Biochem. Biophys. Res. Commun. 2013, 440, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.B.; Cao, Y.F.; Wang, Y. Effect of Neiyi Kangfu suppository on expressions of cytochrome C and survivin in ectopic and eutopic endometrium in rats with endometriosis. Chin. J. Integr. Tradi. West. Med. 2008, 28, 139–141. [Google Scholar] [CrossRef]
- Kearns, M.T.; Barthel, L.; Bednarek, J.M.; Yunt, Z.X.; Henson, P.M.; Janssen, W.J. Fas ligand-expressing lymphocytes enhance alveolar macrophage apoptosis in the resolution of acute pulmonary inflammation. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, 62–70. [Google Scholar] [CrossRef]
- Guo, C.L.; Yang, X.H.; Cheng, W.; Xu, Y.; Li, J.B.; Sun, Y.X.; Bi, Y.M.; Zhang, L.N.; Wang, Q.C. Expression of Fas/FasL in CD8+ T and CD3+ Foxp3+ Treg cells—Relationship with apoptosis of circulating CD8+ T cells in hepatocellular carcinoma patients. Asian Pac. J. Cancer Prev. 2014, 15, 2613–2618. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, X.; He, D.; Jin, X.; Guo, P. Metformin sensitizes human bladder cancer cells to TRAIL-induced apoptosis through mTOR/S6K1-mediated downregulation of c-FLIP. Anticancer Drug. 2014, 25, 887–897. [Google Scholar] [CrossRef]
- Chen, Z.; Sangwan, V.; Banerjee, S.; Chugh, R.; Dudeja, V.; Vickers, S.M.; Saluja, A.K. Triptolide sensitizes pancreatic cancer cells to TRAIL-induced activation of the death receptor pathway. Cancer Lett. 2014, 348, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, M.S.; Huang, Y. Death receptor activation complexes: It takes two to activate TNF receptor 1. Cell Cycle 2003, 2, 550–552. [Google Scholar] [PubMed]
- Chaudhary, P.M.; Eby, M.T.; Jasmin, A.; Kumar, A.; Liu, L.; Hood, L. Activation of the NF-kappaB pathway by caspase 8 and its homologs. Oncogene 2000, 19, 4451–4460. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Sun, P.; Qian, Y.; Suo, H.Y. D. candidum has in vitro anticancer effects in HCT-116 cancer cells and exerts in vivo anti-metastatic effects in mice. Nutr. Res. Pract. 2014, 8, 487–493. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, L.; Zhao, X.; Liu, W.; Deng, J.; Tan, X.; Qiu, L. Anticancer Effect of Ursodeoxycholic Acid in Human Oral Squamous Carcinoma HSC-3 Cells through the Caspases. Nutrients 2015, 7, 3200-3218. https://doi.org/10.3390/nu7053200
Pang L, Zhao X, Liu W, Deng J, Tan X, Qiu L. Anticancer Effect of Ursodeoxycholic Acid in Human Oral Squamous Carcinoma HSC-3 Cells through the Caspases. Nutrients. 2015; 7(5):3200-3218. https://doi.org/10.3390/nu7053200
Chicago/Turabian StylePang, Liang, Xin Zhao, Weiwei Liu, Jiang Deng, Xiaotong Tan, and Lihua Qiu. 2015. "Anticancer Effect of Ursodeoxycholic Acid in Human Oral Squamous Carcinoma HSC-3 Cells through the Caspases" Nutrients 7, no. 5: 3200-3218. https://doi.org/10.3390/nu7053200
APA StylePang, L., Zhao, X., Liu, W., Deng, J., Tan, X., & Qiu, L. (2015). Anticancer Effect of Ursodeoxycholic Acid in Human Oral Squamous Carcinoma HSC-3 Cells through the Caspases. Nutrients, 7(5), 3200-3218. https://doi.org/10.3390/nu7053200