Strawberry Phytochemicals Inhibit Azoxymethane/Dextran Sodium Sulfate-Induced Colorectal Carcinogenesis in Crj: CD-1 Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagent Kits
2.2. Lyophilized Strawberries
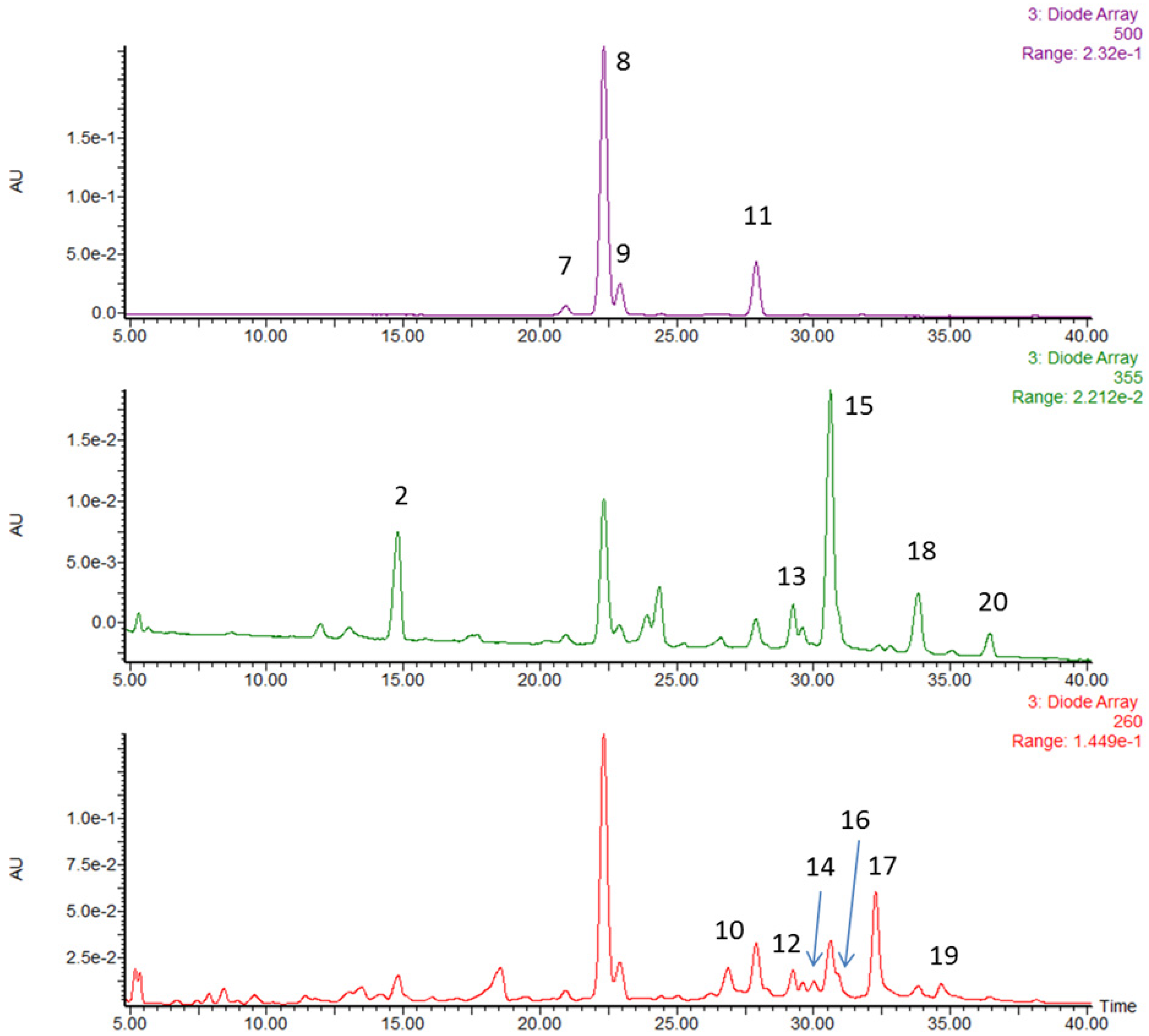
2.3. HPLC-MS/MS Analysis of Strawberry Phytochemicals
2.3.1. Extraction
2.3.2. Identification
2.3.3. Quantification
2.4. Experimental Procedure
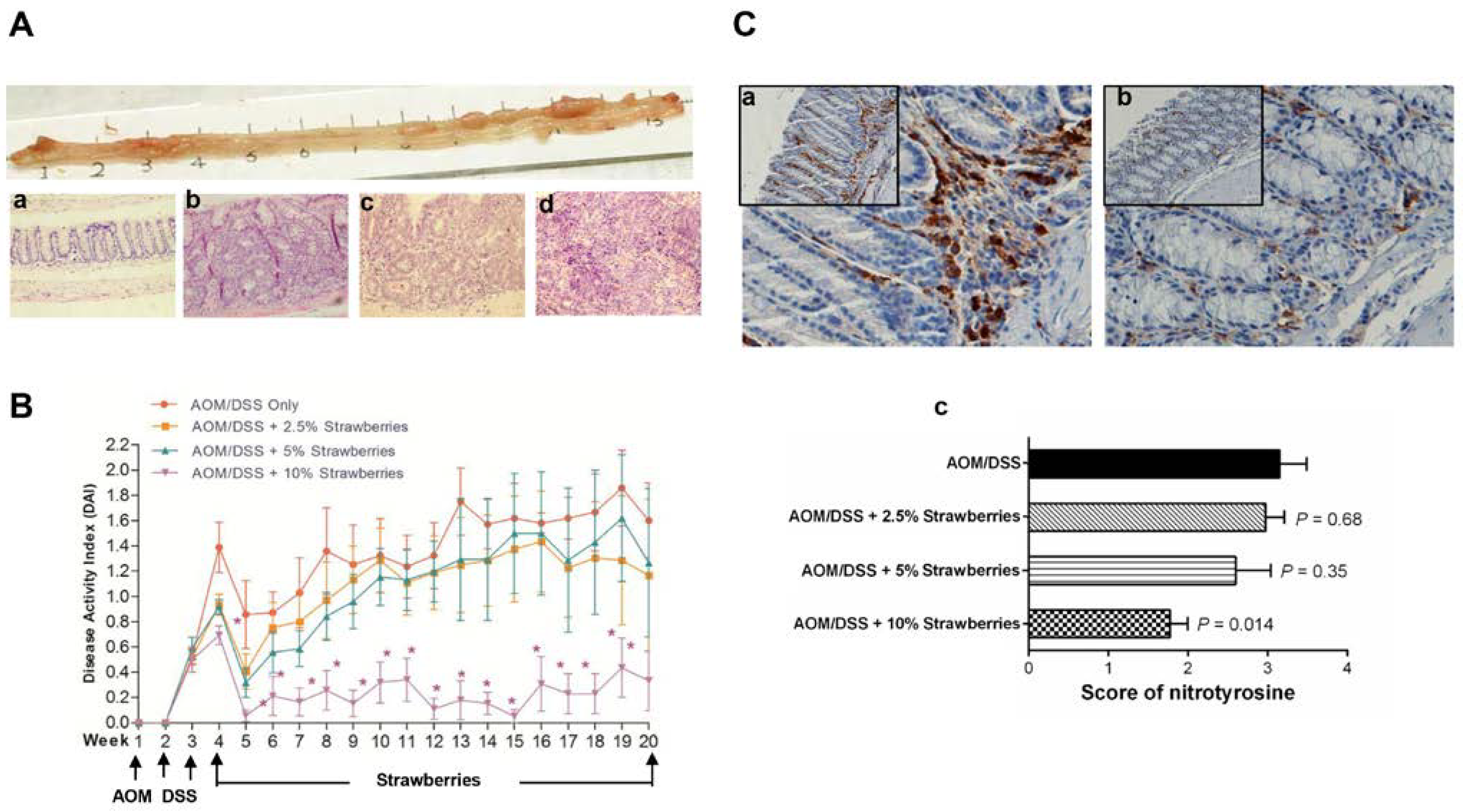
2.5. Evaluation of Histological Grade

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment | Diet | Tumor Incidence (%) | Tumor Multiplicity (Mean ± SE) | Histologic Grade (Mean ± SE; %) | |||
|---|---|---|---|---|---|---|---|---|
| Normal | Dysplasia | Adenoma | Adenocarcinoma | |||||
| 1 | None | AIN-76A | 0 | 0 | - | - | - | - |
| 2 | AOM a + DSS b | AIN-76A | 100 | 3.9 ± 1.5 | 20.2 ± 3.7 | 29.7 ± 3.2 | 20.8 ± 8.1 | 28.8 ± 12.1 |
| 3 | AOM + DSS | 2.5% Straw c | 64 | 3.8 ± 1.4 | 38.5 ± 9.1 f | 46.0 ± 2.3 f | 10.6 ± 6.4 g | 4.7 ± 4.6 g |
| 4 | AOM + DSS | 5.0% Straw | 75 | 3.0 ± 1.1 | 40.1 ± 4.0 f | 50.0 ± 5.0 f | 6.2 ± 5.0 g | 4.7 ± 4.7 g |
| 5 | AOM + DSS | 10.0% Straw | 44 d | 1.8 ± 1.0 e | 45.7 ± 4.0 f | 48.7 ± 1.3 f | 2.8 ±2.8 g | 2.8 ± 2.8 g |
2.6. Modified Disease Activity Index
2.7. Immunohistochemistry
2.8. Real-Time Polymerase Chain Reaction
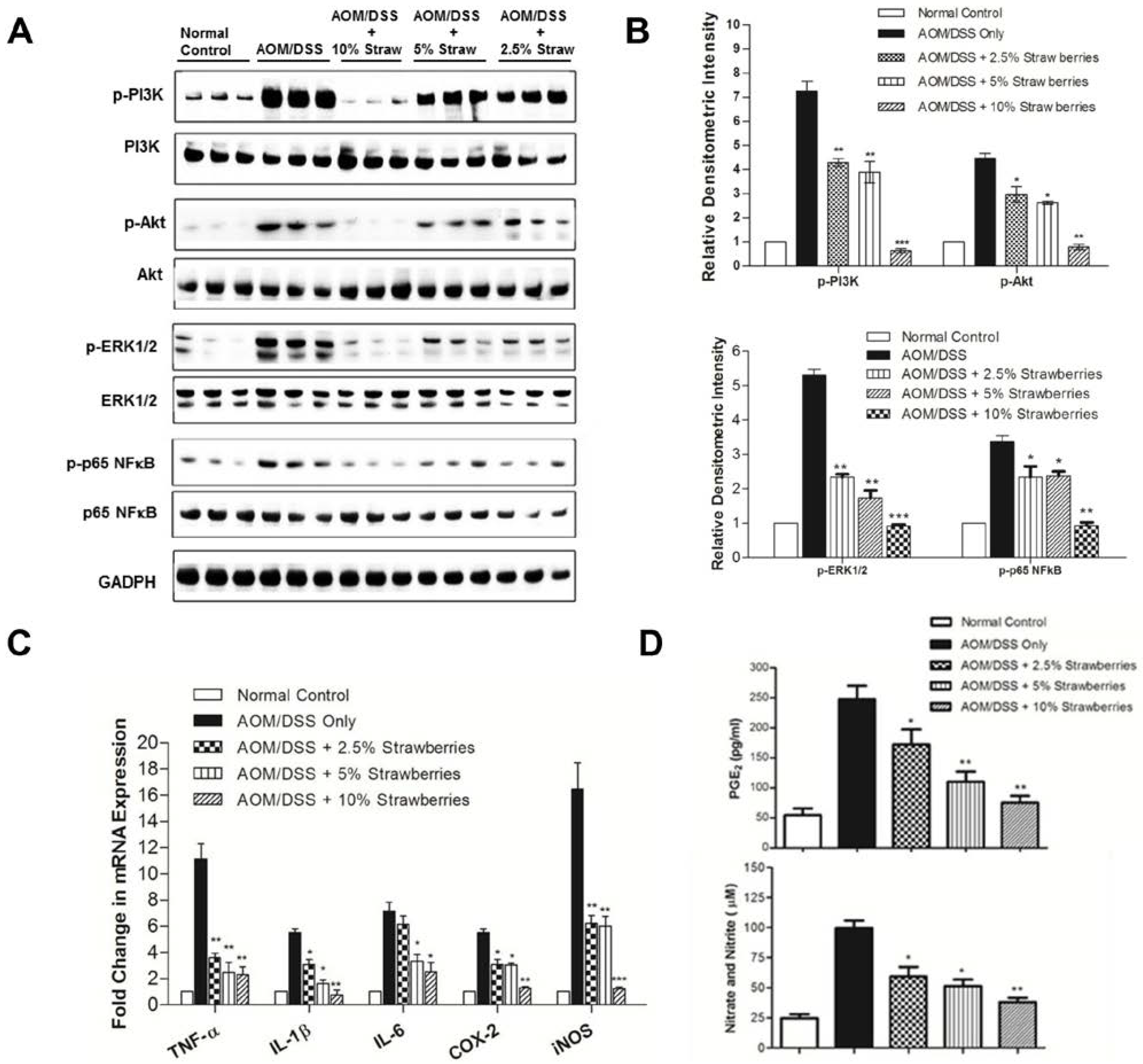
2.9. Western Blot Analysis
2.10. Prostaglandin E2 Measurement and Nitrate/Nitrite Colorimetric Assay
2.11. Statistical Analysis
3. Results
3.1. Characterization and Quantification of Phytochemicals in Strawberries
3.2. General Observations
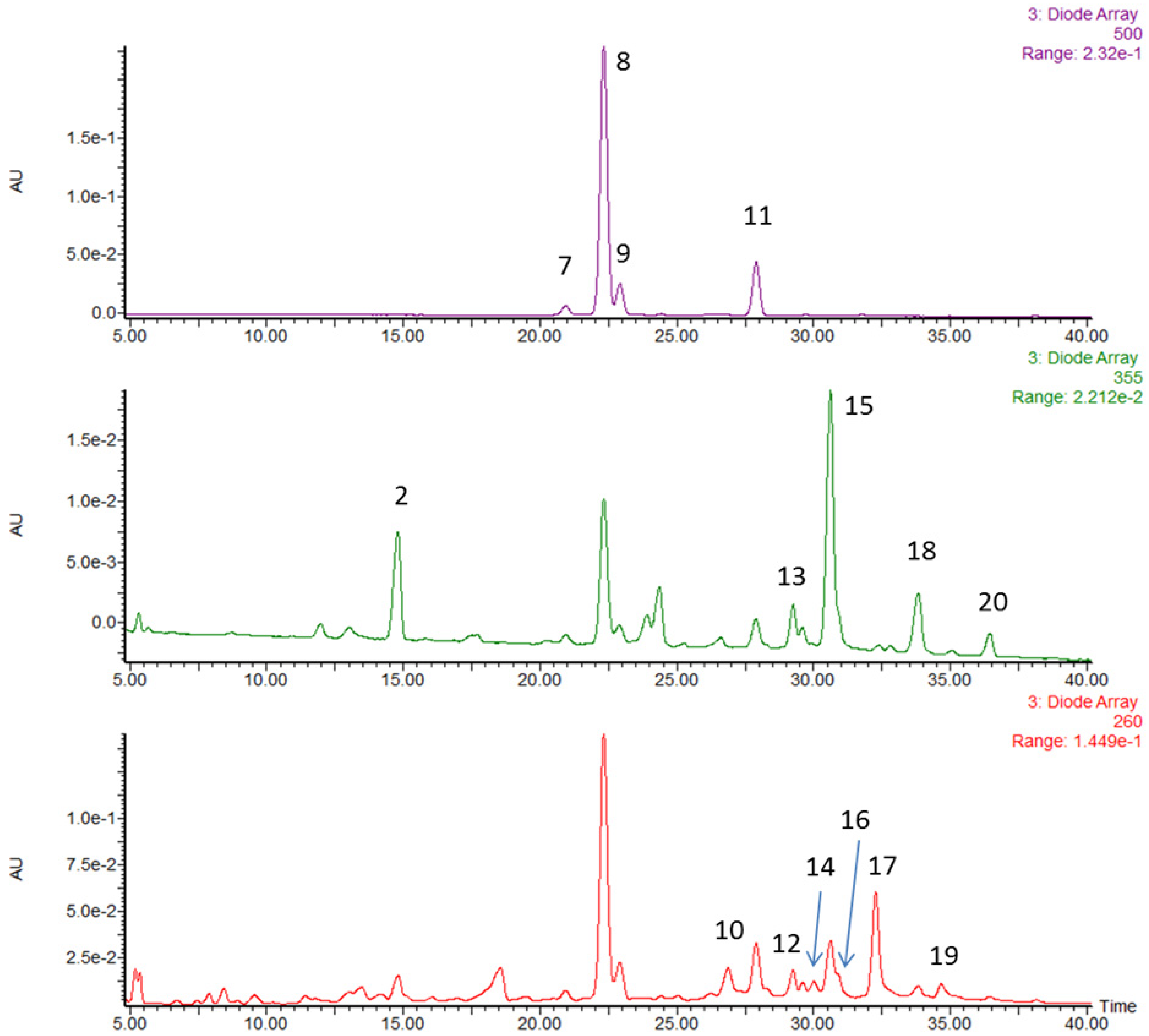
| Peak No. | RT (min) | [M–H]− MSel a | [M–H]− MSe2 b | Peak Assignment | mg (100 g)−1 | % by Weight |
|---|---|---|---|---|---|---|
| 1 | 13.0 | 783.1 | 301.0 | ellagitannin | - | - |
| 2 | 14.8 | 325.1 | 145 | coumaroyl hexoside | - | - |
| 3 | 15.2 | 577.1 | 289 | procyanidin dimer | - | - |
| 4 | 16.1 | 289.1 | - | catechin | - | - |
| 5 | 17.1 | 577.1 | 289 | procyanidin dimer | - | - |
| 6 | 18.0 | 633.1 | 301.0 | ellagitannin | - | - |
| Anthocyanins | ||||||
| 7 | 20.9 | 447.1 | 285 | cyanidin glucoside | 14.9 | 1.7 |
| 8 | 22.3 | 431.1 | 269 | pelargonidin glucoside | 367.7 | 41.1 |
| 9 | 22.9 | 577.2 | 269 | pelargonidin rutinoside | 55.3 | 6.2 |
| 11 | 27.9 | 473 | 269 | pelargonidin malonyl glucoside | 83.9 | 9.4 |
| Total | 58.4 | |||||
| Ellagitannin/ellagic acid/ellagic acid derivatives | ||||||
| 10 | 26.9 | 935.1 | 301.0 | ellagitannin | 64.1 | 7.2 |
| 12 | 28.3 | 935.1 | 301.0 | ellagitannin | 11.4 | 1.3 |
| 13 | 29.3 | 447.1 | 301.0 | ellagic acid rhamnoside | 23.1 | 2.6 |
| 14 | 30 | 935.1 | 301.0 | ellagitannin | 23.1 | 2.6 |
| 16 | 30.7 | 301.0 | - | ellagic acid | 7.3 | 0.8 |
| 17 | 32.3 | (934.1)2− | 301.0 | agrimoniin | 144.5 | 16.2 |
| 19 | 34.6 | (1401.6)2− | 301.0 | lambertianin | 20.3 | 2.3 |
| Total | 16.9 | |||||
| Flavonols | ||||||
| 15 | 30.6 | 477.1 | 301.0 | quercetin hexuronide | 58.8 | 6.6 |
| 18 | 33.8 | 447.1 | 285 | kaempferol glucoside | 14.5 | 1.6 |
| 461.1 | kaempferol hexuronide | |||||
| 20 | 36.4 | 489.1 | 285 | kaempferol malonyl hexoside | 5.1 | 0.6 |
| Total | 10.5 | |||||
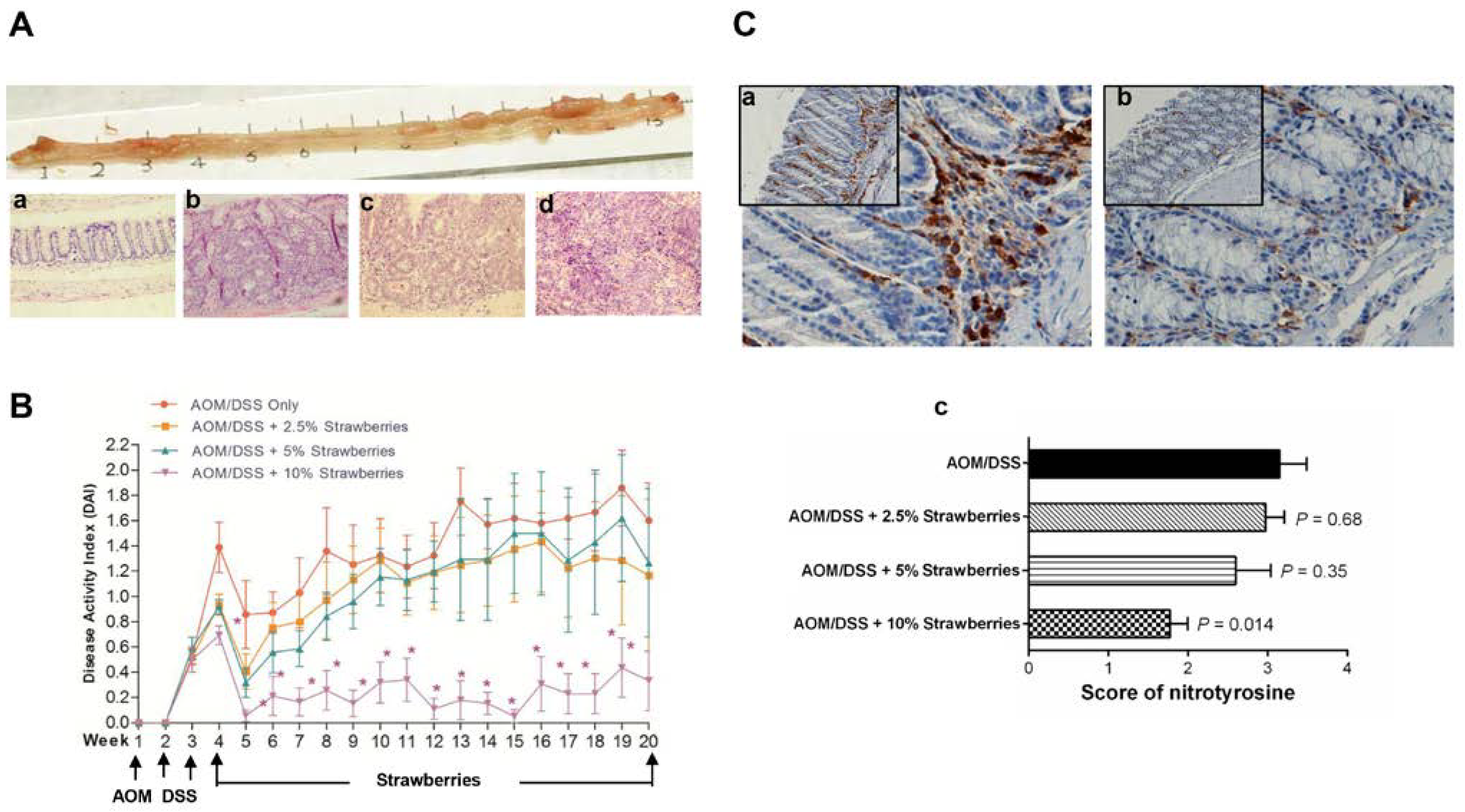
3.3. Strawberries Inhibit Tumor Development and Reduce the Disease Activity Index
3.4. Strawberries Reduce Nitrotyrosine Production
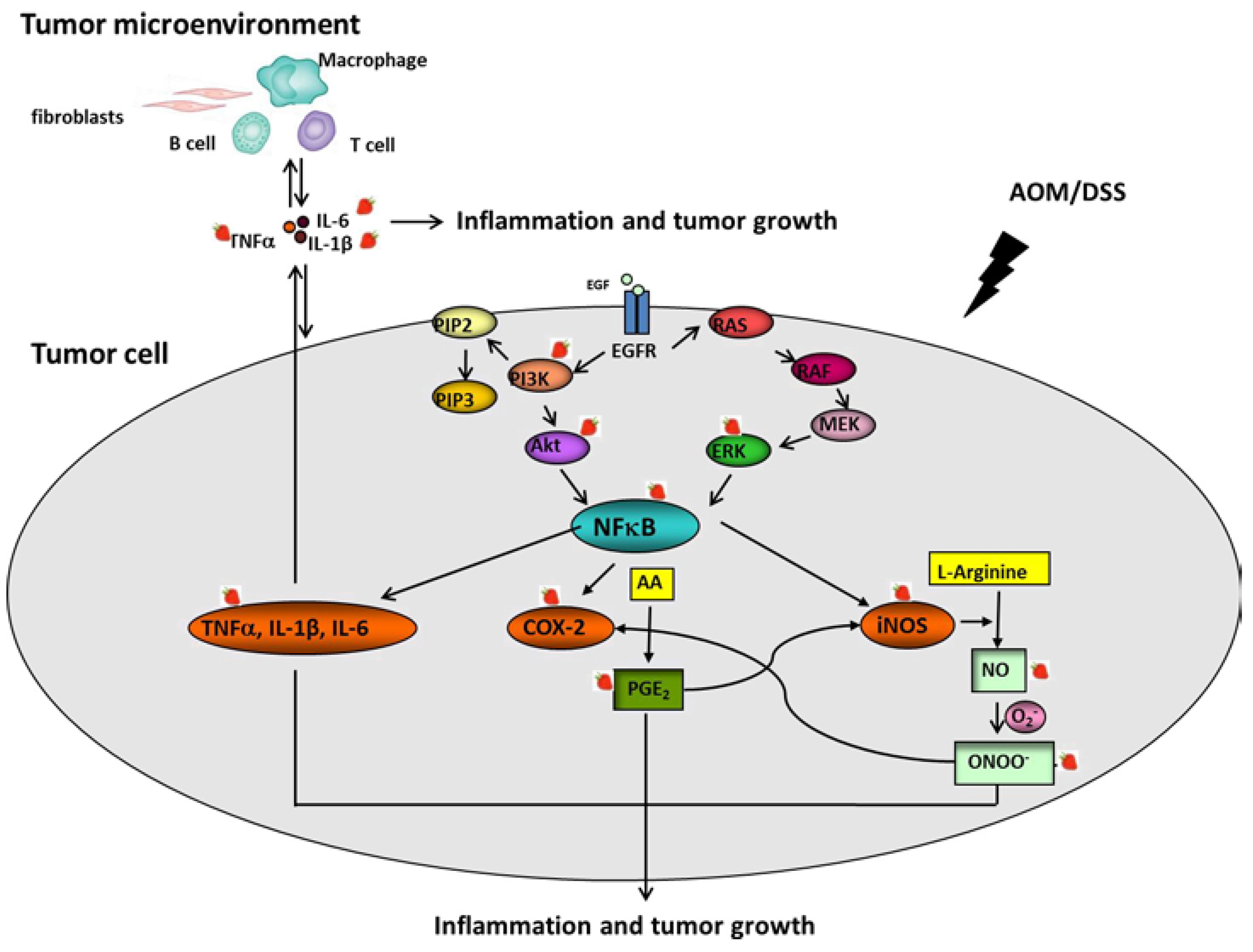
3.5. Strawberries Decrease Phosphorylation of PI3K, Akt, ERK and NFκB
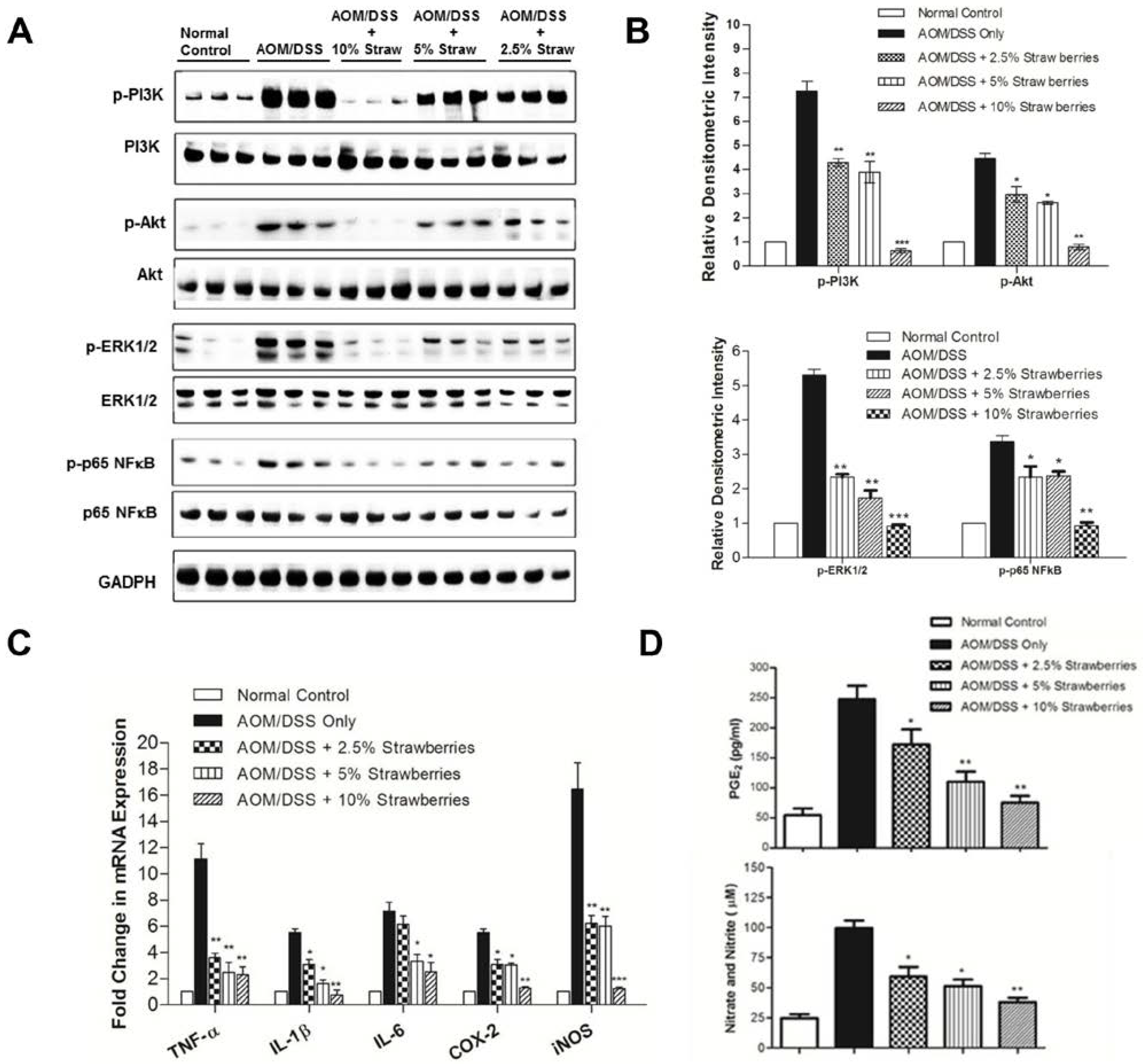
3.6. Strawberries Down-Regulate Expression of Proinflammatory Mediators
3.7. Strawberries Reduce PGE2 and Total Nitrite Productions
4. Discussion
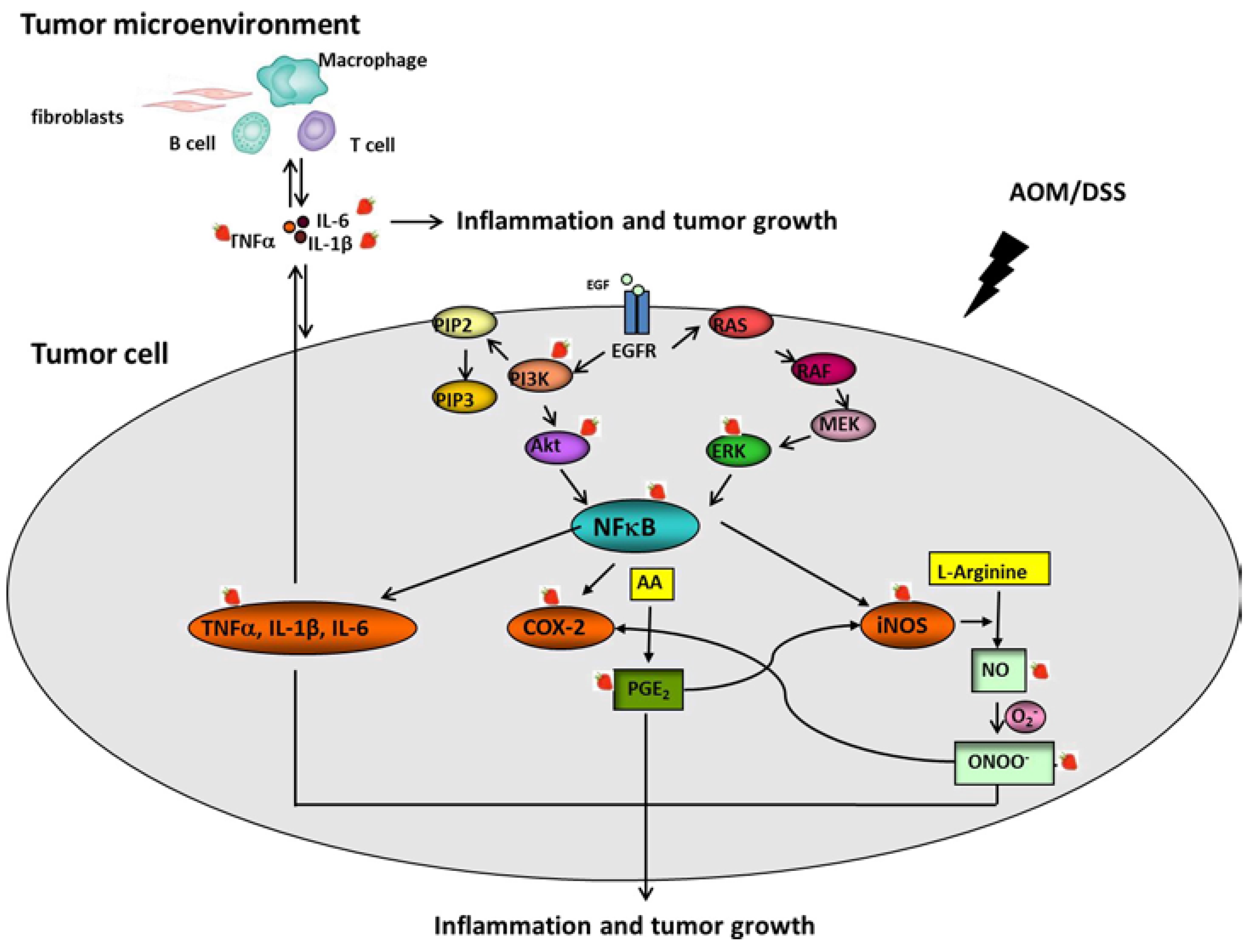
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- American Cancer Society. Cancer Facts and Figures; American Cancer Society: Atlanta, GA, USA, 2012; p. 10. [Google Scholar]
- World Cancer Research Fund/American Institute for Cancer Research. Food, Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective; AICR: Washington, DC, USA, 2007; pp. 1–537. [Google Scholar]
- Azer, S.A. Overview of molecular pathways in inflammatory bowel disease associated with colorectal cancer development. Eur. J. Gastroenterol. Hepatol. 2013, 25, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Feagins, L.A.; Souza, R.F.; Spechler, S.J. Carcinogenesis in IBD: Potential targets for the prevention of colorectal cancer. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Tulipani, S.; Alvarez-Suarez, J.M.; Quiles, J.L.; Mezzetti, B.; Battino, M. The strawberry: Composition, nutritional quality, and impact on human health. Nutrition 2012, 28, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Nguyen, A.; Betts, N.M.; Lyons, T.J. Strawberry as a functional food: An evidence-based review. Crit. Rev. Food Sci. Nutr. 2014, 54, 790–806. [Google Scholar] [CrossRef] [PubMed]
- Casto, B.C.; Knobloch, T.J.; Galioto, R.L.; Yu, Z.; Accurso, B.T.; Warner, B.M. Chemoprevention of oral cancer by lyophilized strawberries. Anticancer Res. 2013, 33, 4757–4566. [Google Scholar] [PubMed]
- Somasagara, R.R.; Hegde, M.; Chiruvella, K.K.; Musini, A.; Choudhary, B.; Raghavan, S.C. Extracts of strawberry fruits induce intrinsic pathway of apoptosis in breast cancer cells and inhibits tumor progression in mice. PLoS One 2012, 7, e47021. [Google Scholar] [CrossRef] [PubMed]
- Balansky, R.; Ganchev, G.; Iltcheva, M.; Kratchanova, M.; Denev, P.; Kratchanov, C.; Polasa, K.; D’Agostini, F.; Steele, V.E.; de Flora, S. Inhibition of lung tumor development by berry extracts in mice exposed to cigarette smoke. Int. J. Cancer 2012, 131, 1991–1997. [Google Scholar] [CrossRef] [PubMed]
- Carlton, P.S.; Kresty, L.A.; Siglin, J.C.; Morse, M.A.; Lu, J.; Morgan, C.; Stoner, G.D. Inhibition of N-nitrosomethylbenzylamine-induced tumorigenesis in the rat esophagus by dietary freeze-dried strawberries. Carcinogenesis 2001, 22, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Yan, F.; Qian, J.; Guo, M.; Zhang, H.; Tang, X.; Chen, F.; Stoner, G.D.; Wang, X. Randomized phase II trial of lyophilized strawberries in patients with dysplastic precancerous lesions of the esophagus. Cancer Prev. Res. 2012, 5, 41–50. [Google Scholar] [CrossRef]
- Yarom, N.; Jonker, D.J. The role of the epidermal growth factor receptor in the mechanism and treatment of colorectal cancer. Discov. Med. 2011, 11, 95–105. [Google Scholar] [PubMed]
- Wang, M.Y.; Lu, K.V.; Zhu, S.; Dia, E.Q.; Vivanco, I.; Shackleford, G.M.; Cavenee, W.K.; Mellinghoff, I.K.; Cloughesy, T.F.; Sawyers, C.L.; et al. Mammalian target of rapamycin inhibition promotes response to epidermal growth factor receptor kinase inhibitors in PTEN-deficient and PTEN-intact glioblastoma cells. Cancer Res. 2006, 66, 7864–7869. [Google Scholar] [CrossRef] [PubMed]
- Nakano, H.; Shindo, M.; Sakon, S.; Nishinaka, S.; Mihara, M.; Yagita, H.; Okumura, K. Differential regulation of IkappaB kinase alpha and beta by two upstream kinases, NF-kappaB-inducing kinase and mitogen-activated protein kinase/ERK kinase kinase-1. Proc. Natl. Acad. Sci. USA 1998, 95, 3537–3542. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Xu, J.; Zhang, X.H.; Qiu, B.Y.; Peng, L.; Zhang, M.; Gan, H.T. PI3K/Akt signaling pathway is involved in the pathogenesis of ulcerative colitis. Inflamm. Res. 2011, 60, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Greten, F.R.; Eckmann, L.; Greten, T.F.; Park, J.M.; Li, Z.W.; Egan, L.J.; Kagnoff, M.F.; Karin, M. IKKbeta links inflammation and tumorigenesis in a mouse model of colitis-associated cancer. Cell 2004, 118, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Morisaki, T.; Sasaki, N.; Nakano, K.; Mibu, R.; Tanaka, M.; Katano, M. Increased nuclear factor-κB activation in human colorectal carcinoma and its correlation with tumor progression. Anticancer Res. 2004, 24, 675–681. [Google Scholar] [PubMed]
- Chen, T.; Hwang, H.; Rose, M.E.; Nines, R.G.; Stoner, G.D. Chemopreventive properties of black raspberries in N-nitrosomethylbenzylamine-induced rat esophageal tumorigenesis: Down-regulation of cyclooxygenase-2, inducible nitric oxide synthase, and c-Jun. Cancer Res. 2006, 66, 2853–2859. [Google Scholar] [CrossRef] [PubMed]
- Stoner, G.D. Foodstuffs for preventing cancer: The preclinical and clinical development of berries. Cancer Prev. Res. 2009, 2, 187–194. [Google Scholar] [CrossRef]
- Gu, J.; Ahn-Jarvis, J.H.; Riedl, K.M.; Schwartz, S.J.; Clinton, S.K.; Vodovotz, Y. Characterization of black raspberry functional food products for cancer prevention human clinical trials. J. Agric. Food Chem. 2014, 62, 3997–4006. [Google Scholar] [CrossRef] [PubMed]
- Giusti, M.M.; Rodríguez-Saona, L.E.; Wrolstad, R.E. Molar absorptivity and color characteristics of acylated and non-acylated pelargonidin-based anthocyanins. J. Agric. Food Chem. 1999, 47, 4631–4637. [Google Scholar] [CrossRef] [PubMed]
- Gasperotti, M.; Masuero, D.; Vrhovsek, U.; Guella, G.; Mattivi, F. Profiling and accurate quantification of Rubus ellagitannins and ellagic acid conjugates using direct UPLC-Q-TOF HDMS and HPLC-DAD analysis. J. Agric. Food Chem. 2010, 58, 4602–4616. [Google Scholar] [CrossRef] [PubMed]
- Riddell, R.H.; Goldman, H.; Ransohoff, D.F.; Appelman, H.D.; Fenoglio, C.M.; Haggitt, R.C.; Ahren, C.; Correa, P.; Hamilton, S.R.; Morson, B.C. Dysplasia in inflammatory bowel disease: Standardized classification with provisional clinical applications. Hum. Pathol. 1983, 14, 931–968. [Google Scholar] [CrossRef] [PubMed]
- Pascal, R.R. Dysplasia and early carcinoma in inflammatory bowel disease and colorectal carcinomas. Hum. Pathol. 1994, 25, 1160–1171. [Google Scholar] [CrossRef] [PubMed]
- Boivin, G.P.; Washington, K.; Yang, K.; Ward, J.M.; Pretlow, T.P.; Russell, R.; Besselsen, D.G.; Godfrey, V.L.; Doetschman, T.; Dove, W.F.; et al. Pathology of mouse models of intestinal cancer: Consensus report and recommendations. Gastroenterology 2003, 124, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Ghia, J.E.; Blennerhassett, P.; Kumar-Ondiveeran, H.; Verdu, E.-F.; Collins, S.M. The vagus nerve: A tonic inhibitory influence associated with inflammatory bowel disease in a murine model. Gastroenterology 2006, 131, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Yasui, Y.; Suzuki, R.; Miyamoto, S.; Tsukamoto, T.; Sugie, S.; Kohno, H.; Tanaka, T. A lipophilic statin, pitavastatin, suppresses inflammation-associated mouse colon carcinogenesis. Int. J. Cancer 2007, 121, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Määttä-Riihinen, K.R.; Kamal-Eldin, A.; Mattila, P.H.; González-Paramás, A.M.; Törrönen, A.M. Distribution and contents of phenolic compounds in eighteen Scandinavian berry species. J. Agric. Food Chem. 2004, 52, 4477–4486. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and human health: Effects beyond antioxidant activity. J. Agric. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef] [PubMed]
- Forbes-Hernández, T.Y.; Giampieri, F.; Gasparrini, M.; Mazzoni, L.; Quiles, J.L.; Alvarez-Suarez, J.M.; Battino, M. The effects of bioactive compounds from plant foodson mitochondrial function: A focus on apoptotic mechanisms. Food Chem. Toxicol. 2014, 68, 154–182. [Google Scholar] [CrossRef] [PubMed]
- Kubotaa, M.; Hosoyaa, T.; Fukumotob, S.; Miyagic, T.; Kumazawaa, S. Anti-melanogenic compounds in Rubus croceacanthus. J. Berry Res. 2014, 4, 127–135. [Google Scholar]
- Schantz, M.; Moh, C.; Baum, M.; Richling, E. Antioxidative efficiency of an anthocyaninrich bilberry extract in the human colon tumor cell lines Caco-2 and HT-29. J. Berry Res. 2010, 1, 25–33. [Google Scholar]
- Zikri, N.N.; Riedl, K.M.; Wang, L.S.; Lechner, J.; Schwartz, S.J.; Stoner, G.D. Blackraspberry components inhibit proliferation, induce apoptosis, and modulate gene expression in rat esophageal epithelial cells. Nutr. Cancer 2009, 6, 816–826. [Google Scholar] [CrossRef]
- Long, N.; Suzuki, S.; Sato, S.; Naiki-Ito, A.; Sakatani, K.; Shirai, T.; Takahashi, S. Purple corn color inhibition of prostate carcinogenesis by targeting cell growth pathways. Cancer Sci. 2013, 104, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Filipiak, K.; Hidalgo, M.; Silvan, J.M.; Fabre, B.; Carbajo, R.J.; Pineda-Lucena, A.; Ramos, A.; de Pascual-Teresa, B.; de Pascual-Teresa, S. Dietary gallic acid and anthocyanin cytotoxicity on human fibrosarcoma HT1080 cells. A study on the mode of action. Food Funct. 2014, 5, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Bazylko, A.; Piwowarski, J.P.; Filipek, A.; Bonarewicz, J.; Tomczyk, M. In vitro antioxidant and anti-inflammatory activities of extracts from Potentilla recta and its main ellagitannin, agrimoniin. J. Ethnopharmacol. 2013, 149, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, M.; Nieminen, R.; Asmawi, M.Z.; Vuorela, P.; Vapaatalo, H.; Moilanen, E. Effects of flavonoids on prostaglandin E2 production and on COX-2 and mPGES-1 expressions in activated macrophages. Planta Med. 2011, 77, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; Latimer, C.; Allsopp, P.; Ternan, N.G.; McMullan, G.; McDougall, G.J.; Stewart, D.; Crozier, A.; Rowland, I.; Gill, C.I. In Vitro and in Vivo models of colorectal cancer: Antigenotoxic activity of berries. J. Agric. Food Chem. 2014, 62, 3852–3866. [Google Scholar] [CrossRef] [PubMed]
- Zackular, J.P.; Baxter, N.T.; Iverson, K.D.; Sadler, W.D.; Petrosino, J.F.; Chen, G.Y.; Schloss, P.D. The gut microbiome modulates colon tumorigenesis. Mbio 2013, 4, e00692–13. [Google Scholar] [CrossRef] [PubMed]
- Clapper, M.L.; Cooper, H.S.; Chang, W.C. Dextran sulfate sodium-induced colitis-associated neoplasia: A promising model for the development of chemopreventive interventions. Acta Pharmacol. Sin. 2007, 28, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Yamada, Y.; Sugie, S.; Mori, H. A novel inflammation-related mouse colon carcinogenesis model induced by azoxymethane and dextran sodium sulfate. Cancer Sci. 2003, 94, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Corn, B.W.; Kovner, F.; Bek, S.; Wexler, I.; Lifschit, S.B.; Seger, R. ERK signaling in colorectal cancer: A preliminary report on the expression of phosphorylated ERK and the effects of radiation therapy. Am. J. Clin. Oncol. 2008, 31, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Pikarsky, E.; Porat, R.M.; Stein, I.; Abramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben-Neriah, Y. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, Z.; Wang, L.; Zhang, X. NF-kappaB signaling pathway, inflammation and colorectal cancer. Cell. Mol. Immunol. 2009, 6, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Taketo, M.M. COX-2 and colon cancer. Inflamm. Res. 1998, 47, S112–S116. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, M.; LaRusso, N.F.; Gores, G.J. Nitric oxide in gastrointestinal epithelial cell carcinogenesis: Linking inflammation to oncogenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G626–G634. [Google Scholar] [PubMed]
- Kaviani, A.; Ohta, M.; Itani, R.; Sander, F.; Tarnawski, A.S.; Sarfeh, I.J. Tumor necrosis factor-alpha regulates inducible nitric oxide synthase gene expression in the portal hypertensive gastric mucosa of the rat. J. Gastrointest. Surg. 1997, 1, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Sands, B.E.; Tremaine, W.J.; Sandborn, W.J.; Rutgeerts, P.J.; Hanauer, S.B.; Mayer, L.; Targan, S.R.; Podolsky, D.K. Infliximab in the treatment of severe, steroid-refractory ulcerative colitis: A pilot study. Inflamm. Bowel Dis. 2001, 7, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.R.; Zhang, J.A.; Zhang, Q.; Permatasari, F.; Xu, Y.; Wu, D.; Yin, Z.Q.; Luo, D. Palmitic acid induces production of proinflammatory cytokines interleukin-6, interleukin-1β, and tumor necrosis factor-α via a NF-κB-dependent mechanism in HaCaT keratinocytes. Med. Inflamm. 2013, 2013, 530429. [Google Scholar]
- Peddareddigari, V.G.; Wang, D.; DuBois, R.N. The Tumor Microenvironment in Colorectal Carcinogenesis. Cancer Microenviron. 2010, 3, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Janakiram, N.B.; Mohammed, A.; Rao, C.V. Role of lipoxins, resolvins, and other bioactive lipids in colon and pancreatic cancer. Cancer Metastasis Rev. 2011, 30, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Half, E.; Arber, N. Colon cancer: Preventive agents and the present status of chemoprevention. Expert Opin. Pharmacother. 2009, 10, 211–219. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, N.; Clinton, S.K.; Liu, Z.; Wang, Y.; Riedl, K.M.; Schwartz, S.J.; Zhang, X.; Pan, Z.; Chen, T. Strawberry Phytochemicals Inhibit Azoxymethane/Dextran Sodium Sulfate-Induced Colorectal Carcinogenesis in Crj: CD-1 Mice. Nutrients 2015, 7, 1696-1715. https://doi.org/10.3390/nu7031696
Shi N, Clinton SK, Liu Z, Wang Y, Riedl KM, Schwartz SJ, Zhang X, Pan Z, Chen T. Strawberry Phytochemicals Inhibit Azoxymethane/Dextran Sodium Sulfate-Induced Colorectal Carcinogenesis in Crj: CD-1 Mice. Nutrients. 2015; 7(3):1696-1715. https://doi.org/10.3390/nu7031696
Chicago/Turabian StyleShi, Ni, Steven K. Clinton, Zhihua Liu, Yongquan Wang, Kenneth M. Riedl, Steven J. Schwartz, Xiaoli Zhang, Zui Pan, and Tong Chen. 2015. "Strawberry Phytochemicals Inhibit Azoxymethane/Dextran Sodium Sulfate-Induced Colorectal Carcinogenesis in Crj: CD-1 Mice" Nutrients 7, no. 3: 1696-1715. https://doi.org/10.3390/nu7031696
APA StyleShi, N., Clinton, S. K., Liu, Z., Wang, Y., Riedl, K. M., Schwartz, S. J., Zhang, X., Pan, Z., & Chen, T. (2015). Strawberry Phytochemicals Inhibit Azoxymethane/Dextran Sodium Sulfate-Induced Colorectal Carcinogenesis in Crj: CD-1 Mice. Nutrients, 7(3), 1696-1715. https://doi.org/10.3390/nu7031696