Rice and Bean Targets for Biofortification Combined with High Carotenoid Content Crops Regulate Transcriptional Mechanisms Increasing Iron Bioavailability

Abstract
:1. Introduction
2. Experimental Section
2.1. Sample
2.2. Staple Food Crop Flours Preparation
2.3. Determination of Iron and Zinc

{kind=link}
{kind=link}
{kind=link}
| Pontal Bean | Rice | Pumpkin | Sweet Potato | |
|---|---|---|---|---|
| Moisture (g·100 g−1) | 10.7 a ± 0.28 | 7.35 d ± 0.06 | 9.99 a,b ± 0.55 | 9.92 b ± 0.06 |
| Ash (g·100 g−1) | 3.14 b ± 0.03 | 0.34 e ± 0.02 | 6.38 a ± 0.07 | 2.27 d ± 0.06 |
| Lipids (g·100 g−1) | 1.37 b ± 0.3 | 0.13 c ± 0.13 | 1.46 b ± 0.14 | 1.55 e ± 0.34 |
| Protein (g·100 g−1) | 18.86 b ± 0.08 | 8.83 d ± 0.18 | 15.86 c ± 0.24 | 2.63 e ± 0.12 |
| Total dietary fiber (g·100 g−1) | 26.69 a ± 0.45 | 1.08 c ± 0.1 | 15.02 b ± 0.03 | 15.31 b ± 0.31 |
| Soluble fiber | 7.04 ± 1.27 | 0.47 | 5.10 ± 0.25 | 4.89 ± 0.63 |
| Insoluble fiber | 19.64 ± 0.92 | 0.61 | 9.92 ± 0.23 | 10.42 ± 0.38 |
| Carbohydrates (g·100 g−1) | 48.87 b,c ± 0.73 | 82.48 a ± 0.05 | 52.19 b,c ± 0.34 | 69.62 a,c ± 0.56 |
| Total phenolic (mg·de·EqGA/g) | 1.33 b ± 0.15 | 0.06 d ± 0.01 | 2.41 a ± 0.12 | 1.51 b ± 0.07 |
| Carotenoids (mg/100 g) | nd | nd | 308.84 a ± 1.98 | 127.11 b ± 0.06 |
| Minerals | ||||
| Iron (mg/100 g) | 7.52 ± 0.1 | 3.9 ± 0.03 | 2.09 ± 0.18 | 3.33 ± 0.06 |
| Zinc (mg/100 g) | 3.11 ± 0.01 | 1.73 ± 0.06 | 1.71 ± 001 | 1.8 ± 0.05 |
| Phytic acid (g/100 g) | 0.51 a ± 0.02 | 0.20 b ± 0.03 | 0.03 c ± 0.32 | 0.10 c ± 0.1 |
| Molar Ratio | ||||
| Phytate/iron | 5.78 | 4.38 | 1.26 | 2.54 |
| Zinc/iron | 0.35 | 0.37 | 0.72 | 0.46 |
2.4. Determination of Carotenoids
2.5. Phytate and Phenolic Compounds
2.6. Animals and Diets
2.7. Blood Tests
| Standard Diet without Iron | Standard Diet with Iron (Ferrous Sulfate) | Pontal Bean | Pontal Bean and Rice | Pontal Bean and Pumpkin | Pontal Bean and Sweet Potato | Pontal Bean, Rice and Pumpkin | Pontal Bean, Rice and Sweet Potato | |
|---|---|---|---|---|---|---|---|---|
| Ingredients (1 kg of Diet) | ||||||||
| Ferrous Sulfate (mg) | - | 120.98 | - | - | - | - | - | - |
| Common Bean (g) | - | - | 163.73 | 100.84 | 156.36 | 155.16 | 91.41 | 88.05 |
| Rice (g) | - | - | - | 100.84 | - | - | 91.41 | 88.05 |
| Pumpkin (g) | - | - | - | - | 25.56 | - | 25.56 | - |
| Sweet Potato (g) | - | - | - | - | - | 18.85 | - | 18.85 |
| Albumin (g) | 200.00 | 200.00 | 173.44 | 178.11 | 170.18 | 174.98 | 177.65 | 180.93 |
| Dextrinized starch (g) | 132.00 | 132.00 | 132.00 | 132.00 | 132.00 | 132.00 | 132.00 | 132.00 |
| Sucrose (g) | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Soybean Oil (mL) | 70.00 | 70.00 | 67.71 | 68.62 | 67.85 | 67.87 | 69.88 | 69.88 |
| Microcrystalline cellulose (g) | 50.00 | 50.00 | 10.98 | 24.96 | 9.33 | 10.42 | 24.73 | 24.70 |
| Mineral Mix without iron (g) | 35.00 | 35.00 | 35.00 | 35.00 | 35.00 | 35.00 | 35.00 | 35.00 |
| Vitamin Mix (g) | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| l-cystine (g) | 3.00 | 3.00 | 3.00 | 3.00 | 2.73 | 3.00 | 3.00 | 3.00 |
| Choline Bitartrate (g) | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 |
| Corn starch (g) | 397.50 | 397.50 | 337.89 | 309.32 | 333.10 | 337.20 | 305.41 | 302.83 |
| Nutritional Composition | ||||||||
| Total calories (Kcal) | 3830.8 | 3830.8 | 3989.47 | 4105.9 | 4013.1 | 4028.6 | 4100.4 | 4093.5 |
| Caloric density (Kcal/g) | 3.83 | 3.83 | 3.98 | 4.1 | 4.01 | 4.02 | 4.1 | 4.09 |
| Vitamin A (mg/kg) | 1.20 | 1.20 | 1.20 | 1.20 | 5.70 | 5.70 | 5.70 | 5.70 |
| Iron (mg/kg) | 0.30 | 20.4 * | 23.7 ± 0.81 * | 19.7 ± 0.68 * | 26.3 ± 4.7 * | 22.5 ± 0.09 * | 23.9 ± 3.62 * | 22.7 ± 0.82 * |
| Phytate (g/100 g) | nd | nd | 0.83 | 0.72 | 0.805 | 0.81 | 0.63 | 0.66 |
| Phytate: iron molar ratio | nd | nd | 29.81 a ± 1.01 | 30.79 a ± 1.07 | 30.25 a ± 0.12 | 26.89 a ± 4.8 | 22.88 a ± 3.46 | 24.86 a ± 0.9 |
| Zinc: iron molar ratio | - | - | 0.18 a ± 0.006 | 0.21 a ± 0.007 | 0.2 a ± 0.0008 | 0.17 a ± 0.03 | 0.17 a ± 0.02 | 0.18 a ± 0.006 |
| Genes | Oligonucleotide (5’–3’) | |
|---|---|---|
| Forward | Reverse | |
| GAPDH | AGGTTGTCTCCTGTCACTTC | CTGTTGCTGTAGCCATATTC |
| DMT-1 | CTGATTTACAGTCTGGAGCAG | CACTTCAGCAAGGTGCAA |
| DcytB | TGCAGACGCAGAGTTAAGCA | CCGTGAAGTATACCGGCTCC |
| Ferroportin | TTCCGCACTTTTCGAGATGG | TACAGTCGAAGCCCAGGACCGT |
| Hephaestin | GGCACAGTTACAGGGCAGAT | AGTAACGTGGCAGTGCATCA |
| Ferritin | CAGCCGCCTTACAAGTCTCT | ATGGAGCTAACCGCGAAGAC |
| Transferrin | AGCTGCCACCTGAGAACATC | CGCACGCCCTTTATTCATGG |
2.8. Iron Bioavailability
2.9. Extraction of mRNA and Expression of Proteins Involved in Iron Metabolism
2.10. Determination of Gene Expression of Proteins Involved in Iron Metabolism by Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
2.11. Plasma Total Antioxidant Capacity
2.12. Statistical Analysis
3. Results
3.1. The Effect of Combinations of Staple Food Crops on the Bioavailability of Iron
| Fe Intake | Vitamin A Intake | HG | %HRE | RBV-HRE | |
|---|---|---|---|---|---|
| FS | 6.75 a ± 0.19 | 0.39 b ± 0.01 | 6.9 A ± 1.95 | 76.92 B ± 0.4 | - |
| PB | 5.13 b,c ± 0.28 | 0.39 b ± 0.02 | 2.6 B,b ± 1.33 | 60.71 C,b ± 0.15 | 0.79 b ± 0.2 |
| PB + R | 4.28 e ± 0.33 | 0.39 b ± 0.03 | 3.84 B,a ± 1.03 | 87.52A,a ± 0.16 | 1.14 a ± 0.21 |
| PB + P | 4.6 d,e ± 0.38 | 1.29 a ± 0.11 | 4.85 A,a ± 1.16 | 86.75 A,a ± 0.12 | 1.13 a ± 0.16 |
| PB + SP | 5.37 b ± 0.67 | 1.28 a ± 0.16 | 5.77 A,a ± 2.6 | 86.72 A,a ± 0.24 | 1.13 a ± 0.32 |
| PB + R + P | 5.2 b,c ± 0.15 | 1.37 a ± 0.04 | 4.72 A,a ± 1.56 | 81.65 A,a ± 0.12 | 1.06 a,b ± 0.15 |
| PB + R + SP | 4.81 c,d ± 0.37 | 1.33 a ± 0.1 | 4.97 A,b ± 1.85 | 85.72 A,a ± 0.24 | 1.05 a,b ± 0.33 |
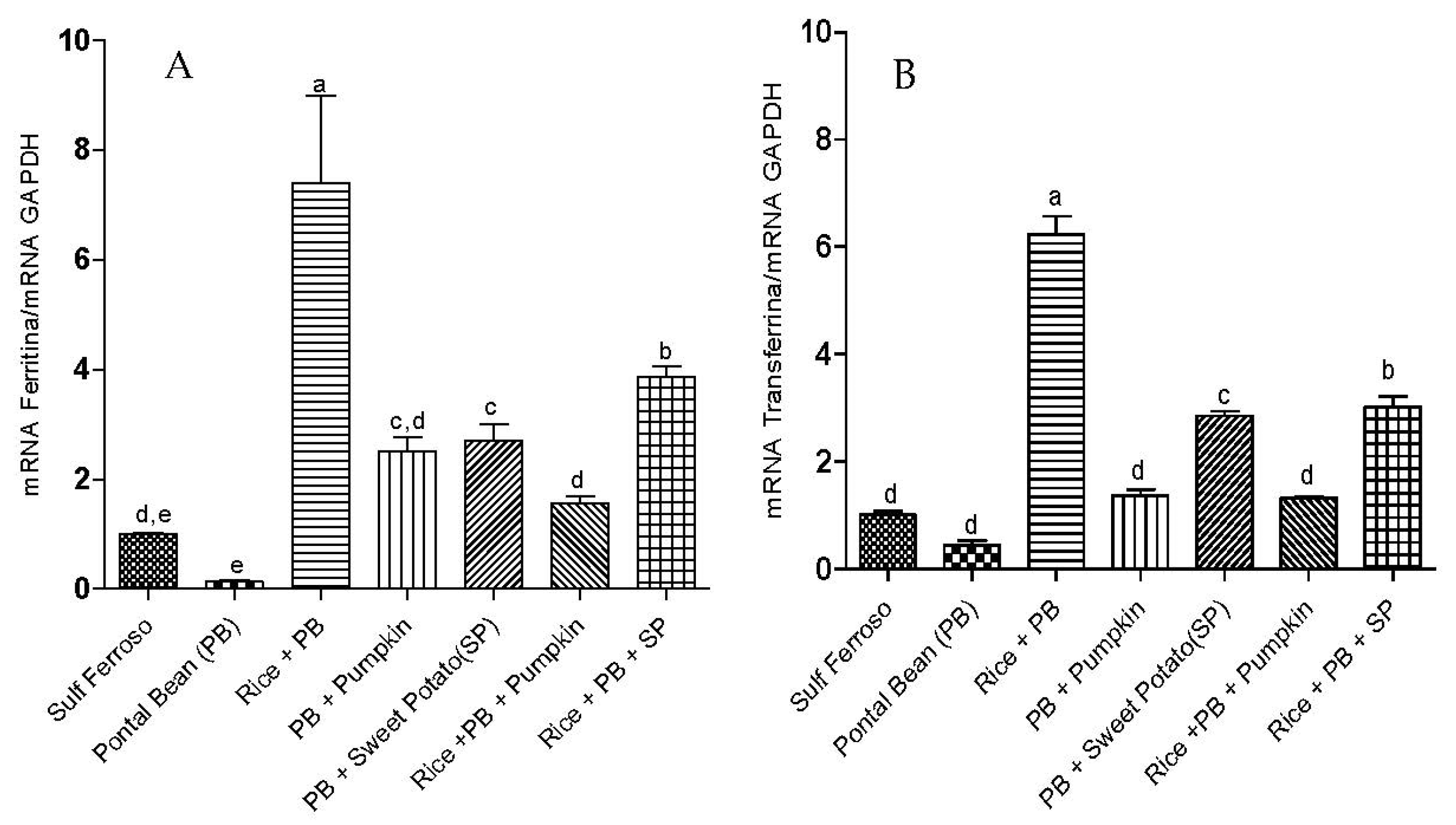
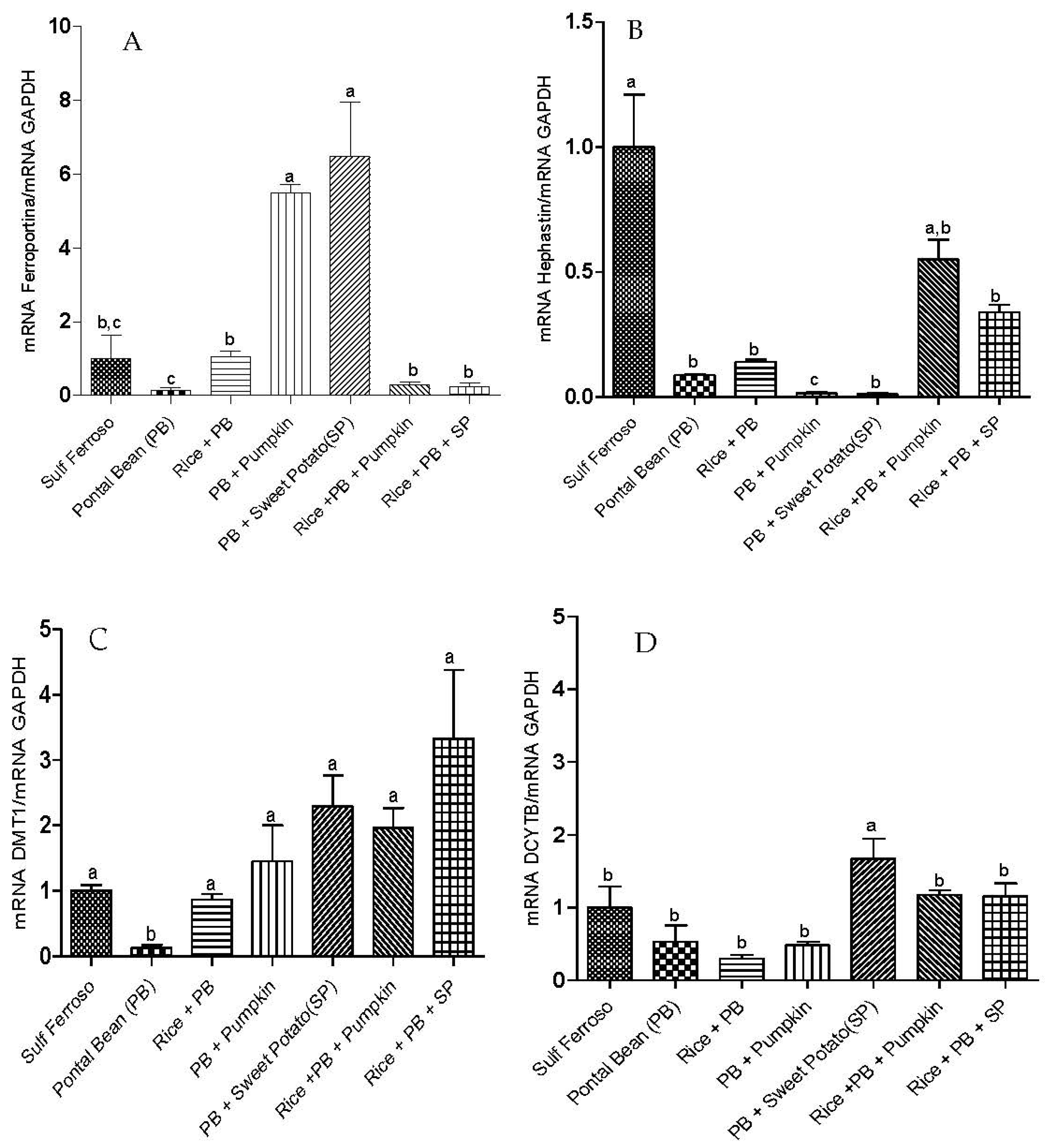
3.2. Gene Expression of Proteins Involved in Iron Metabolism
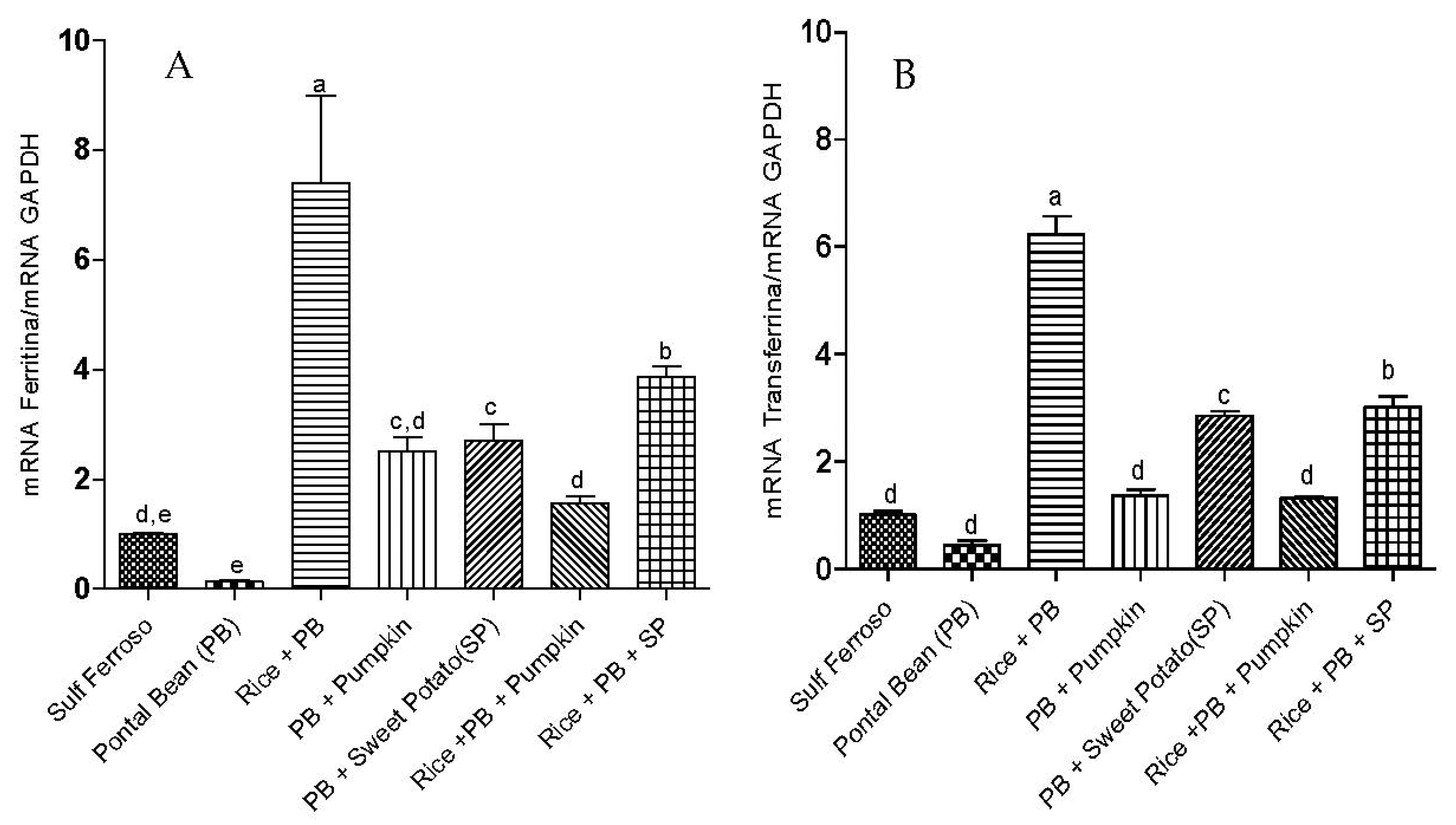
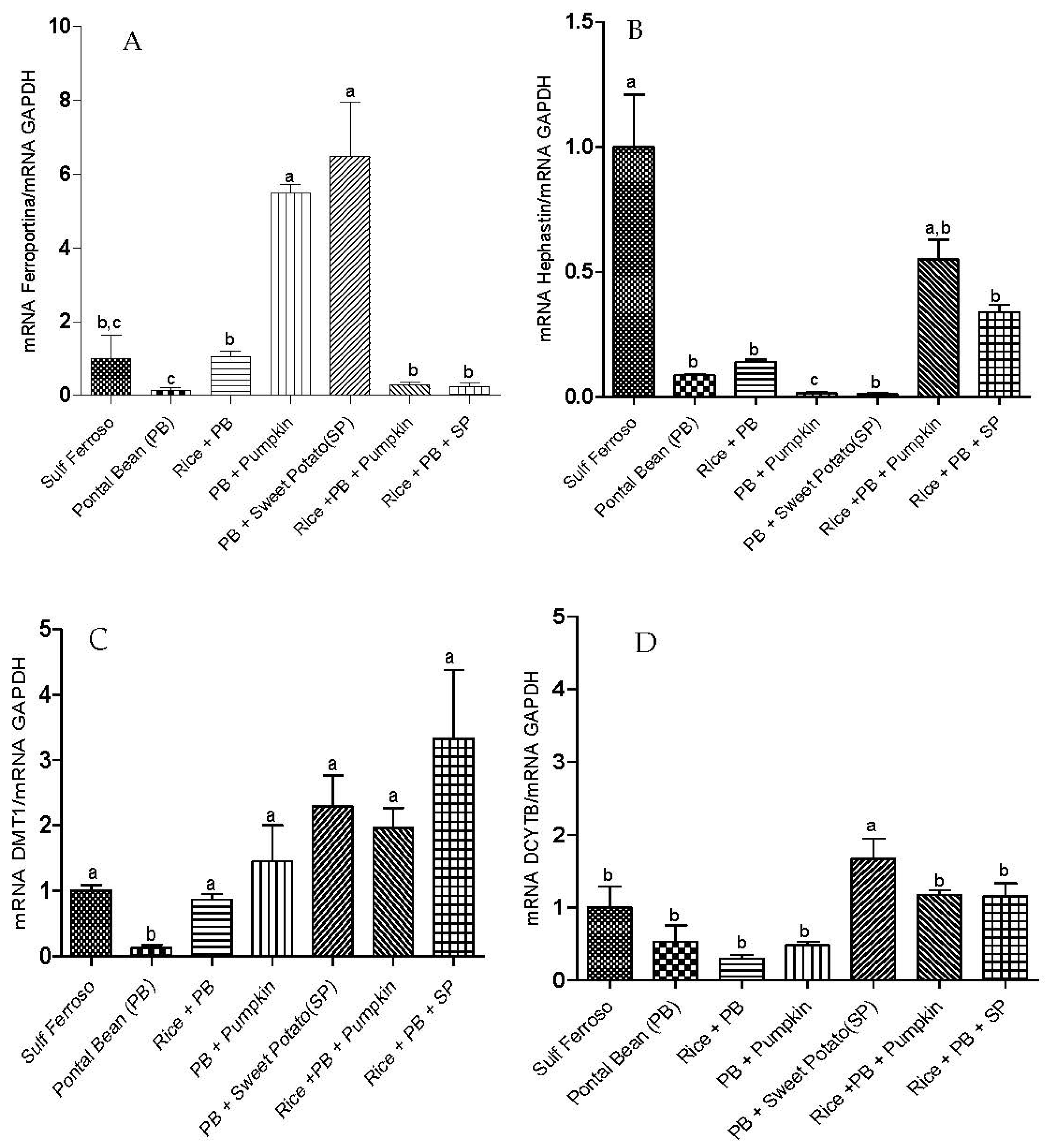
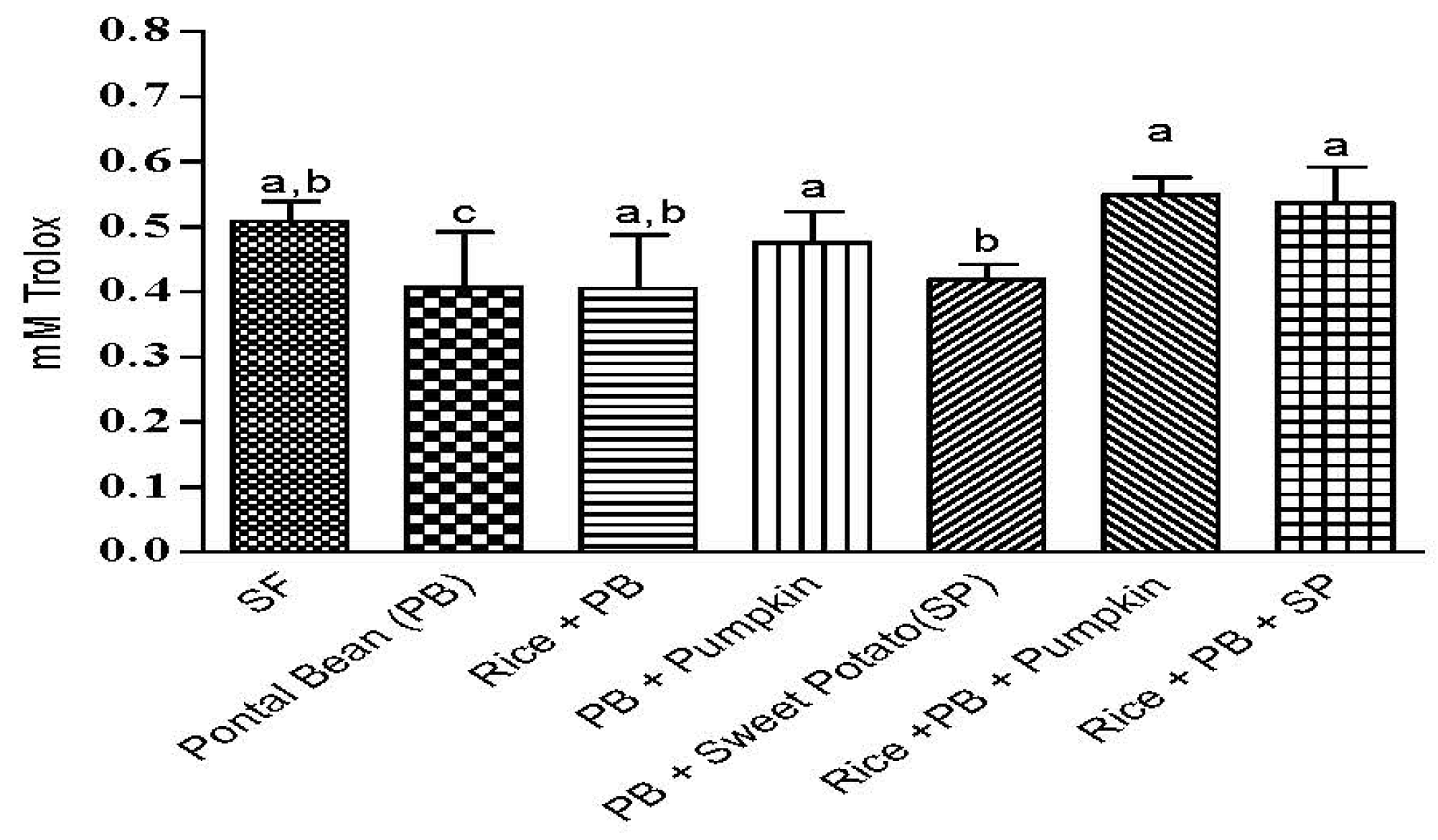
3.3 The Effect of the Combinations of Staple Food Crops on Plasma Total Antioxidant Capacity
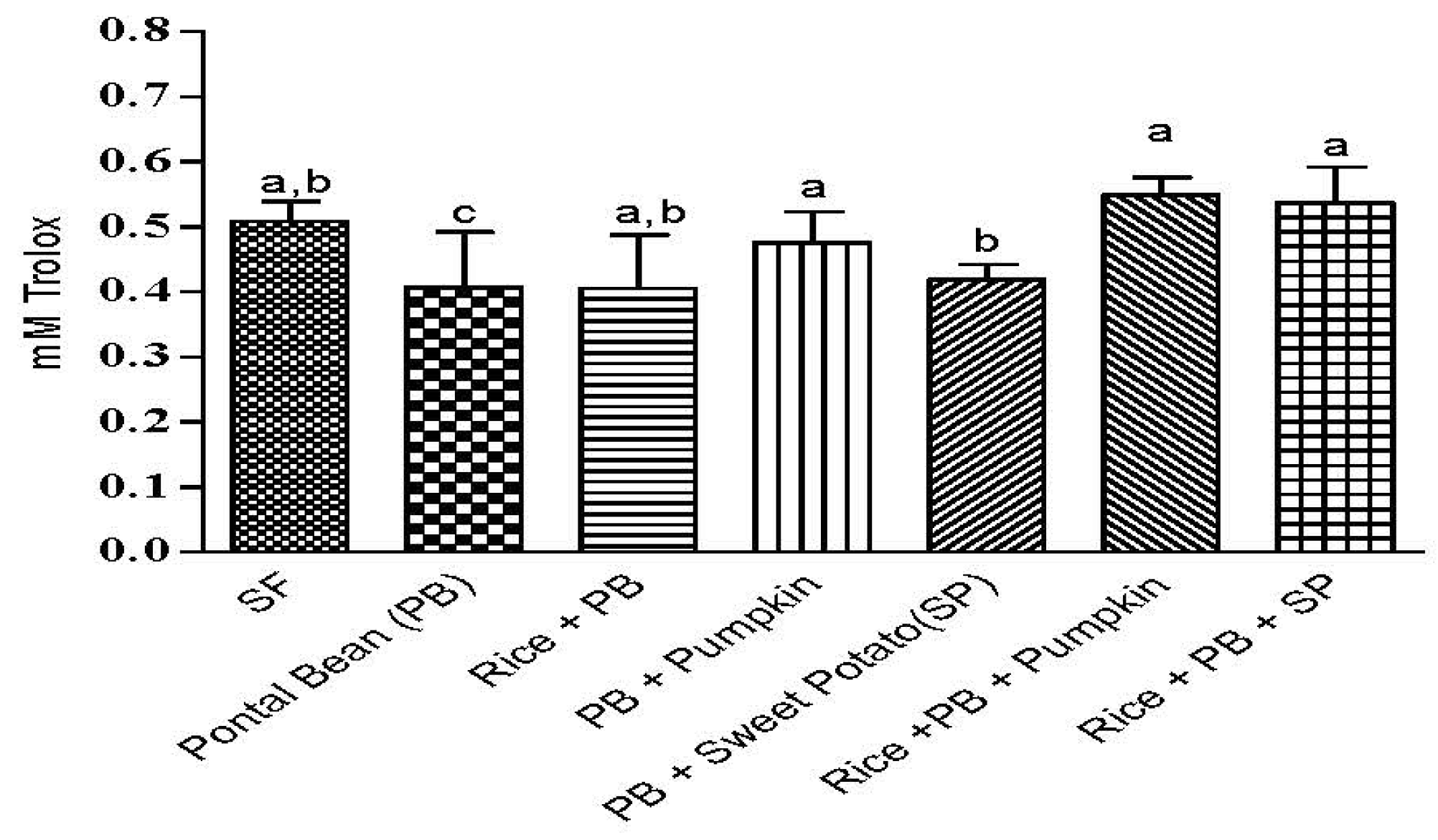
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Muthayya, S.; Rah, J.H.; Sugimoto, J.D.; Roos, F.F.; Kraemer, K.; Black, R.E. The global hidden hunger indices and maps: An advocacy tool for action. PLoS ONE 2013, 8, e67860. [Google Scholar] [CrossRef] [PubMed]
- WHO. Worldwide Prevalence of Anaemia 1993–2005; WHO Global Database on Anaemia; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Bouis, H.E.; Hotz, C.; Mcclafferty, B.; Meenakshi, J.V.; Pfeiffer, W.H. Biofortification: A new tool to reduce micronutrient malnutrition. Food Nut. Bull. 2011, 11, 31–40. [Google Scholar]
- La Frano, M.R.; de Moura, F.F.; Boy, E.; Lönnerdal, B.; Burri, B.J. Bioavailability of iron, zinc, and provitamin A carotenoids in biofortified staple crops. Nutr. Rev. 2014, 72, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Casgrain, A.; Collings, R.; Harvey, L.J.; Boza, J.J.; Fairweather-Tait, S.J. Micronutrient bioavailability research priorities. Am. J. Clin. Nutr. 2010, 91, 1423–1429. [Google Scholar] [CrossRef] [PubMed]
- Alférez, M.J.; Díaz-Castro, J.; López-Aliaga, I.; Rodríguez-Ferrer, M.; Pérez-Sánchez, L.J.; Campos, M.S. Development of nutritional iron deficiency in growing male rats: Haematological parameters, iron bioavailability and oxidative defence. Br. J. Nutr. 2011, 105, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Dragsted, L.O. Biomarkers of exposure to vitamins A, C and E and their relation to lipid and protein oxidation markers. Eur. J. Nutr. 2008, 47, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Roman, S.D. Antioxidant systems and oxidative stress in the testes. Oxid. Med. Cell. Longev. 2008, 1, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.B.; Biebinger, R.; Rohner, F.; Dib, A.; Zeder, C.; Hurrell, R.F.; Chaouki, N. Vitamin A supplementation in children with poor vitamin A and iron status increases erythropoietin and hemoglobin concentrations without changing total body iron. Am. J. Clin. Nutr. 2006, 84, 580–586. [Google Scholar] [PubMed]
- Arruda, S.F.; de Almeida Siqueira, E.M.; de Valência, F.F. Vitamin A deficiency increases hepcidin expression and oxidative stress in rat. Nutrition 2009, 25, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Mehdad, A.; Siqueira, E.M.; Arruda, S.F. Effect of vitamin A deficiency on iron bioavailability. Ann. Nutr. Metab. 2009, 57, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Gondim, S.S.R.; Diniz, A.D.S.; Cagliari, M.P.P.; Araújo, E.D.S.; Queiroz, D.D.; Paiva, A.D.A. Relationship among hemoglobin levels, serum retinol level and nutritional status in children aged 6 to 59 months from the state of Paraíba, Brazil. Rev. Nutr. 2012, 25, 441–449. [Google Scholar] [CrossRef]
- Glahn, R.P.; Lee, O.A.; Yeung, A.; Goldman, M.I.; Miller, D.D. Caco-2 cell ferritin formation predicts nonradiolabeled food iron availability in an in vitro digestion/Caco-2 cell culture model. J. Nutr. 1998, 128, 1555–1561. [Google Scholar] [PubMed]
- Tako, E.; Hoekenga, O.A.; Kochian, L.V.; Glahn, R.P. High bioavailablilty iron maize (Zea mays L.) developed through molecular breeding provides more absorbable iron in vitro (Caco-2 model) and in vivo (Gallus gallus). Nutr. J. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- De Moura, N.C.; Canniatti-Brazaca, S.G. Avaliação da disponibilidade de ferro de feijão comum. Ciênc. Tecnol. Aliment. 2006, 26, 270–276. [Google Scholar] [CrossRef]
- Ariza-Nieto, M.; Blair, M.W.; Welch, R.M.; Glahn, R.P. Screening of iron bioavailability patterns in eight bean (Phaseolus vulgaris L.) genotypes using the Caco-2 cell in vitro model. J. Agric. Food Chem. 2007, 55, 7950–7956. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Blair, M.W.; Glahn, R.P. Biofortified red mottled beans (Phaseolus vulgaris L.) in a maize and bean diet provide more bioavailable iron than standard red mottled beans: Studies in poultry (Gallus gallus) and an in vitro digestion/Caco-2 model. Nutr. J. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- Glahn, R.P.; Cheng, Z.; Fukai, S.; Rerkasem, B.; Huang, L. The bioavailability of iron fortified in whole grain parboiled rice. Food Chem. 2009, 112, 982–986. [Google Scholar] [CrossRef]
- Haas, J.D.; Beard, J.L.; Murray-Kolb, L.E.; del Mundo, A.M.; Felix, A.; Gregorio, G.B. Iron-biofortified rice improves the iron stores of nonanemic Filipino women. Nutr. J. 2005, 135, 2823–2830. [Google Scholar]
- Vaz-Tostes, M.G.; Verediano, T.A.; Mejia, E.G.; Costa, N.M.B. Evaluation of iron and zinc bioavailability of beans targeted for biofortification using in vitro and in vivo models and their effect on the nutritional status of preschool children. J. Sci. Food Agric. 2015. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Cárdenas, L.; Leonel, A.J.; Costa, N.M.B. Efeito do processamento doméstico sobre o teor de nutrientese de fatores antinutricionais de diferentes cultivares de feijão comum. Ciênc. Tecnol. Aliment. 2008, 28, 200–213. [Google Scholar] [CrossRef]
- Silva, R.G.V. Caracterização Físico-Quimica de Farinha de Batata-Doce Para Produtos de Panificação. Dissertação (Mestrado), Universidade Estadual do Sudoeste da Bahia, Bahia, Brasil, 2010. [Google Scholar]
- Gomes, J.C. Análise de Alimentos; Universidade Federal de Viçosa: Viçosa, Brasil, 1996; p. 126. [Google Scholar]
- Pires, P.C.C.; Candido, F.G.; Cardoso, L.M.; Costa, N.; Martino, H.S.; Pinheiro-Sant’Ana, H.M. Comparison of mineral and trace element contents between organically and conventionally grown fruit. Fruits 2015, 70, 29–36. [Google Scholar] [CrossRef]
- Rodriguez, D.B.; Raymundo, L.C.; Lee, T.C.; Simpson, K.L.; Chichester, C.O. Carotenoid pigment changes in ripening Momordica charantia fruits. Ann. Bot. 1976, 40, 615–624. [Google Scholar]
- Sant’Ana, H.M.P.; Stringheta, P.C.; Brandão, S.C.C.; de Azeredo, R.M.C. Carotenoid retention and vitamin A value in carrot (Daucus carota L.) prepared by food service. Food Chem. 1998, 61, 145–151. [Google Scholar] [CrossRef]
- Latta, M.; Eskin, M. A simple and rapid colorimetric method for phytate determination. J. Agric. Food Chem. 1980, 28, 1313–1315. [Google Scholar] [CrossRef]
- Ellis, R.; Morris, R. Appropriate resin selections for rapid phytate analysis by ion-exchange chromatograpy. Cereal Chem. 1986, 63, 58–59. [Google Scholar]
- Singleton, V.L.; Rudolf, O.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folil-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- AOAC—Association of Official Analytical Chemistry. Official Methods of Analysis, 19th ed.; Official methods of analysis of the AOAC international: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [PubMed]
- Della Lucia, C.M.; Tostes, M.G.V.; Silveira, C.M.; Bordalo, L.A.; Rodrigues, F.C.; Pinheiro-Sant’Ana, H.M.; Martino, H.S.; Costa, N.M. Iron bioavailability in Wistar rats fed with fortified rice by Ultra Rice technology with or without addition of yacon flour (Smallanthus sonchifolius). Arch. Lat. Am. Nutr. 2013, 63, 64–73. [Google Scholar]
- Mwanri, L.; Worsley, A.; Ryan, P.; Masika, J. Supplemental vitamin A improves anemia and growth in anemic school children in Tanzania. J. Nutr. 2000, 130, 2691–2696. [Google Scholar] [PubMed]
- Hernández, M.; Sousa, V.; Moreno, Á.; Villapando, S.; López-Alarcón, M. Iron bioavailability and utilization in rats are lower from lime-treated corn flour than from wheat flour when they are fortified with different sources of iron. J. Nutr. 2003, 133, 154–159. [Google Scholar] [PubMed]
- Petry, N.; Egli, I.; Gahutu, J.B.; Tugirimana, P.L.; Boy, E.; Hurrell, R. Stable iron isotope studies in Rwandese women indicate that the common bean has limited potential as a vehicle for iron biofortification. J. Nutr. 2012, 142, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Bloem, M.W. The anemia of vitamin A deficiency: Epidemiology and pathogenesis. Eur. J. Clin. Nutr. 2002, 56, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Oates, P.S. The role of hepcidin and ferroportin in iron absorption. Histol. Histopathol. 2007, 22, 791–804. [Google Scholar] [PubMed]
- Hurrell, R.F.; Juillerat, M.A.; Reddy, M.B.; Lynch, S.R.; Dassenko, S.A.; Cook, J.D. Soy protein, phytate, and iron absorption in humans. Am. J. Clin. Nutr. 1992, 56, 573–578. [Google Scholar] [PubMed]
- Anton, A.A.; Ross, K.A.; Beta, T.; Fulcher, R.G.; Arntfield, S.D. Effect of pre-dehulling treatments on some nutritional and physical properties of navy and pinto beans (Phaseolus vulgaris L.). LWT Food Sci. Tech. 2008, 41, 771–778. [Google Scholar] [CrossRef]
- Petry, N.; Egli, I.; Campion, B.; Nielsen, E.; Hurrell, R. Genetic reduction of phytate in common bean (Phaseolus vulgaris L.) seeds increases iron absorption in young women. J. Nutr. 2013, 143, 1219–1224. [Google Scholar] [CrossRef] [PubMed]
- Garcia-CasaL, M.N.; Layrisse, M. Food iron absorption: Role of vitamin A. Arch. Latinoam. Nutr. 1998, 48, 191–196. [Google Scholar] [PubMed]
- Petry, N.; Egli, I.; Zeder, C.; Walczyk, T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Citelli, M.; Bittencourt, L.L.; Silva, S.V.; Pierucci, A.P.T.; Pedrosa, C. Vitamin A modulates the expression of genes involved in iron bioavailability. Biol. Trace Elem. Res. 2012, 149, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Brasse-LagneL, C.; Karim, Z.; Letteron, P.; Bekri, S.; Bado, A.; Beaumont, C. Intestinal DMT1 cotransporter is down-regulated by hepcidin via proteasome internalization and degradation. Gastroenterology 2011, 140, 1261–1271. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Beebe, S.E.; Spenser Reed, S.; Hart, J.J.; Glahn, R.P. Polyphenolic compounds appear to limit the nutritional benefit of biofortified higher iron black bean (Phaseolus vulgaris L.). Nutr. J. 2014, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Tako, E.; Reed, S.M.; Budiman, J.; Jonathan, J.H.; Glahn, R.P. Higher iron pearl millet (Pennisetum glaucum L.) provides more absorbable iron that is limited by increased polyphenolic content. Nutr. J. 2015, 14. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, Y.; Yokoyama, K.; Yamamoto, H.; Nakayama, M.; Hosoya, T. Do serum hepcidin-25 levels correlate with oxidative stress in patients with chronic kidney disease not receiving dialysis? Clin. Nephrol. 2012, 78, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Mena, N.P.; Esparza, A.; Tapia, V.; Valdés, P.; Nunez, M.T. Hepcidin inhibits apical iron uptake in intestinal cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G192–G198. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, B.; Platel, K.; Srinivasan, K. Double fortification of sorghum (Sorghum bicolor L. Moench) and finger millet (Eleucine coracana L. Gaertn) flours with iron and zinc. J. Cereal Sci. 2012, 55, 195–201. [Google Scholar] [CrossRef]
- Groto, H.Z.W. Fisiologia e metabolismo do ferro. Rev. Bras. Hematol. Hemoter. 2010, 32, 8–17. [Google Scholar] [CrossRef]
- He, W.L.; Feng, Y.; Wei, Y.Y.; Yang, X.E.; Shi, C.H.; He, Z.L.; Stoffella, P.J. Differential iron-bioavailability with relation to nutrient compositions in polished rice among selected Chinese genotypes using Caco-2 cell culture model. Int. J. Food Sci. Nutr. 2013, 64, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, Y.; Deng, X.; Xu, Y.; Gao, Z.; Li, H. Iron overload-induced rat liver injury: Involvement of protein tyrosine nitration and the effect of baicalin. Eur. J. Pharm. 2012, 680, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Van der Gaag, M.S.; van den Berg, R.; van den Berg, H.; Schaafsma, G.; Hendriks, H.F. Moderate consumption of beer, red wine and spirits has counteracting effects on plasma antioxidants in middle-aged men. Eur. J. Clin. Nutr. 2000, 54, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Young, J.F.; Dragsted, L.O.; Haraldsdóttir, J.; Daneshvar, B.; Kall, M.A.; Loft, S.; Nilsson, L.; Nielsen, S.E.; Mayer, B.; Skibsted, L.H.; et al. Green tea extract only affects markers of oxidative status postprandially: Lasting antioxidant effect of flavonoid-free diet. Br. J. Nutr. 2002, 87, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, H.; Moreira, E.A.; da Cunha, D.F.; Junqueira-Franco, M.V.; Bernardes, M.M.; Jordão, A.A., Jr. Papel dos nutrientes na peroxidação lipídica e no sistema de defesa antioxidante. Medicina 1998, 31, 31–44. [Google Scholar] [CrossRef]
- Cornelis, P.; Wei, Q.; Andrews, S.C.; Vinckx, T. Iron homeostasis and management of oxidative stress response in bacteria. Metallomics 2011, 3, 540–549. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dias, D.M.; De Castro Moreira, M.E.; Gomes, M.J.C.; Lopes Toledo, R.C.; Nutti, M.R.; Pinheiro Sant’Ana, H.M.; Martino, H.S.D. Rice and Bean Targets for Biofortification Combined with High Carotenoid Content Crops Regulate Transcriptional Mechanisms Increasing Iron Bioavailability. Nutrients 2015, 7, 9683-9696. https://doi.org/10.3390/nu7115488
Dias DM, De Castro Moreira ME, Gomes MJC, Lopes Toledo RC, Nutti MR, Pinheiro Sant’Ana HM, Martino HSD. Rice and Bean Targets for Biofortification Combined with High Carotenoid Content Crops Regulate Transcriptional Mechanisms Increasing Iron Bioavailability. Nutrients. 2015; 7(11):9683-9696. https://doi.org/10.3390/nu7115488
Chicago/Turabian StyleDias, Desirrê Morais, Maria Eliza De Castro Moreira, Mariana Juste Contin Gomes, Renata Celi Lopes Toledo, Marilia Regini Nutti, Helena Maria Pinheiro Sant’Ana, and Hércia Stampini Duarte Martino. 2015. "Rice and Bean Targets for Biofortification Combined with High Carotenoid Content Crops Regulate Transcriptional Mechanisms Increasing Iron Bioavailability" Nutrients 7, no. 11: 9683-9696. https://doi.org/10.3390/nu7115488
APA StyleDias, D. M., De Castro Moreira, M. E., Gomes, M. J. C., Lopes Toledo, R. C., Nutti, M. R., Pinheiro Sant’Ana, H. M., & Martino, H. S. D. (2015). Rice and Bean Targets for Biofortification Combined with High Carotenoid Content Crops Regulate Transcriptional Mechanisms Increasing Iron Bioavailability. Nutrients, 7(11), 9683-9696. https://doi.org/10.3390/nu7115488