Antioxidant and Anti-Fatigue Constituents of Okra

Abstract
:
1. Introduction
2. Experimental Section
2.1. Plant Material and Extraction
2.2. Chemicals and Reagents
2.3. Animals
2.4. Chemical Analysis of OSD and OSK
2.4.1. Determination of Total Flavonoid (TF) Content
2.4.2. Determination of Total Polyphenol (TP) Content
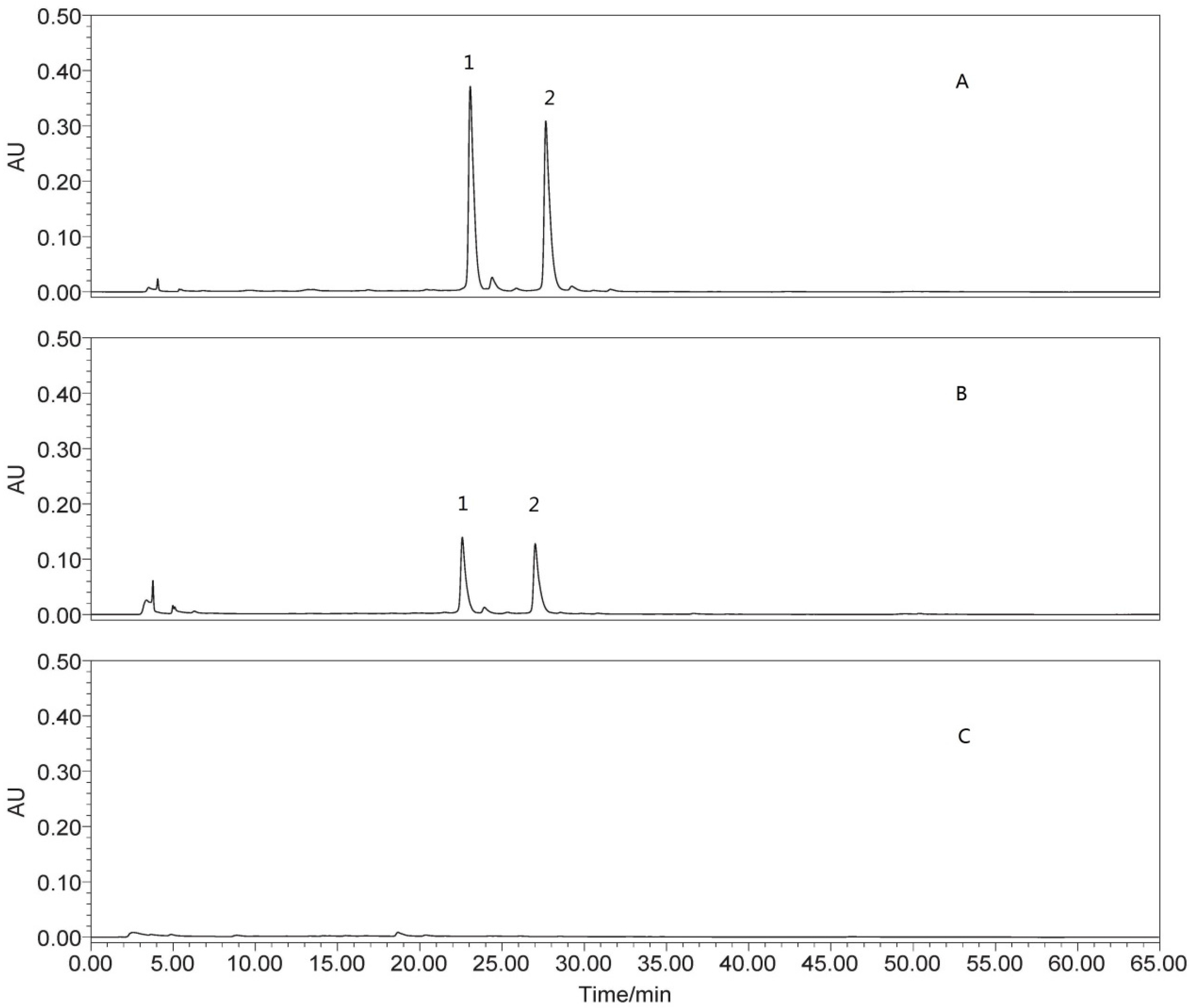
2.4.3. Determination of Isoquercitrin and Quercetin-3-O-Gentiobiose
2.4.4. Determination of Total Polysaccharide (TPS) content
2.5. In Vitro Antioxidant Assays
2.6. Anti-fatigue Effects of OPD, OSK and OSD and Biochemical Analysis
2.6.1. Open-Field Test
2.6.2. Weight-Loaded Swimming Test (WLST)
2.6.3. Analysis of Biochemical Parameters Related to Fatigue
2.7. Statistical Analysis
3. Results
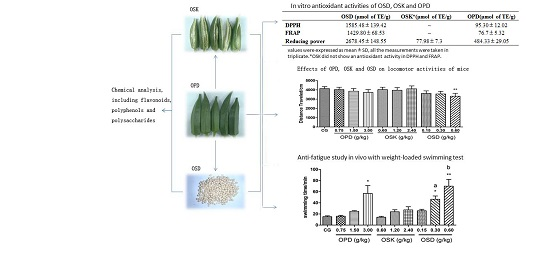
3.1. Chemical Analysis of OSK and OSD

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | OSD | OSK | OPD |
|---|---|---|---|
| TF (%) | 5.35 | ~ | 1.02 |
| TP (%) | 29.5 | 1.25 | 6.73 |
| TPS (%) | 14.8 | 43.06 | 38.65 |
| Isoquercitrin (%) | 2.067 | ~ | 0.395 |
| Quercetin-3-O-gentiobiose (%) | 2.741 | ~ | 0.541 |
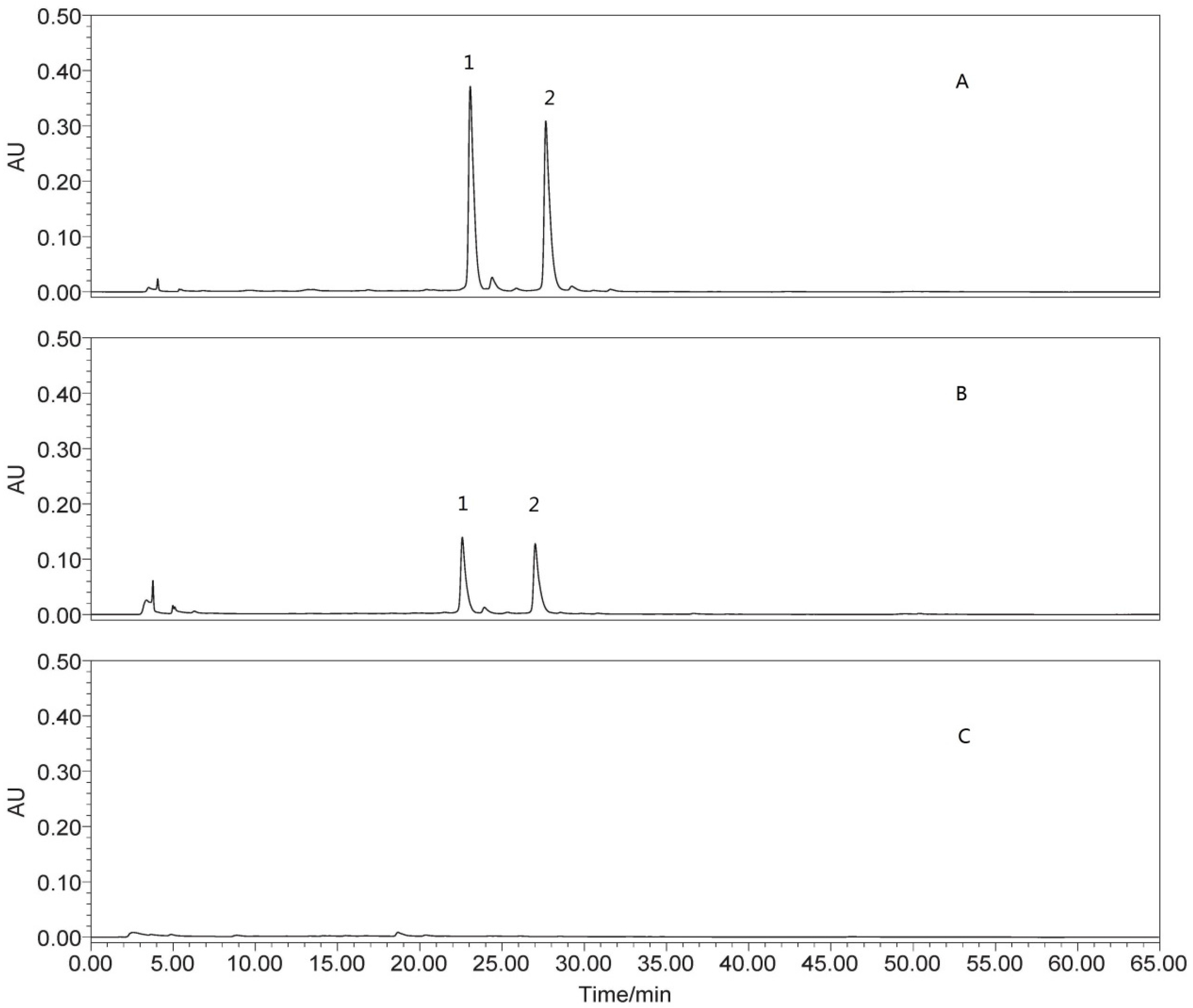
3.2. In Vitro Antioxidant Activity of OPD, OSK and OSD
| OSD (μmol of TE/g) | OSK *(μmol of TE/g) | OPD (μmol of TE/g) | |
|---|---|---|---|
| DPPH | 1585.48 ± 139.42 | ~ | 95.30 ± 12.02 |
| FRAP | 1429.80 ± 68.53 | ~ | 76.7 ± 5.32 |
| Reducing power | 2678.45 ± 148.55 | 77.98 ± 7.3 | 484.33 ± 29.05 |
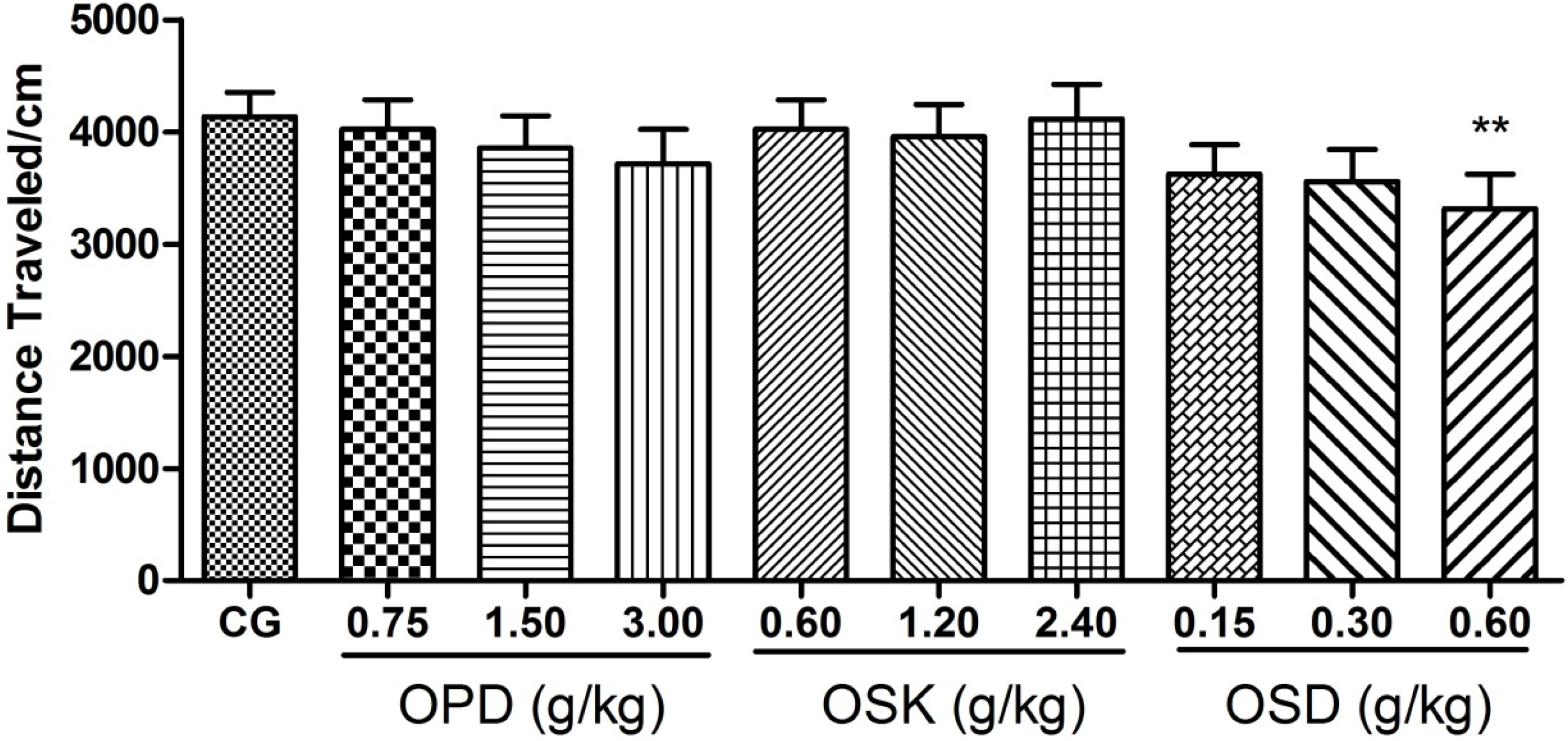
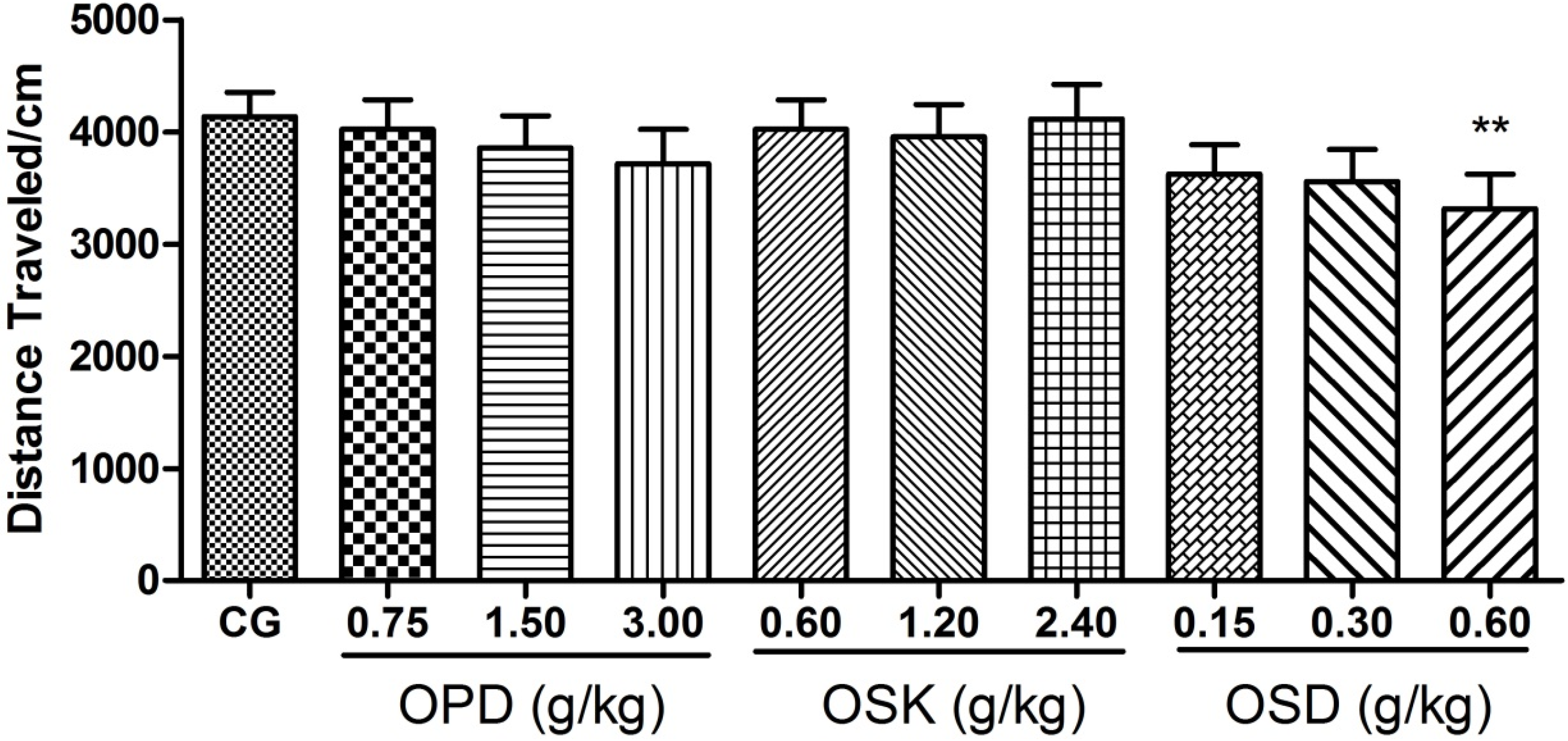
3.3. Effects on Locomotor Activitiy of OPD, OSK and OSD
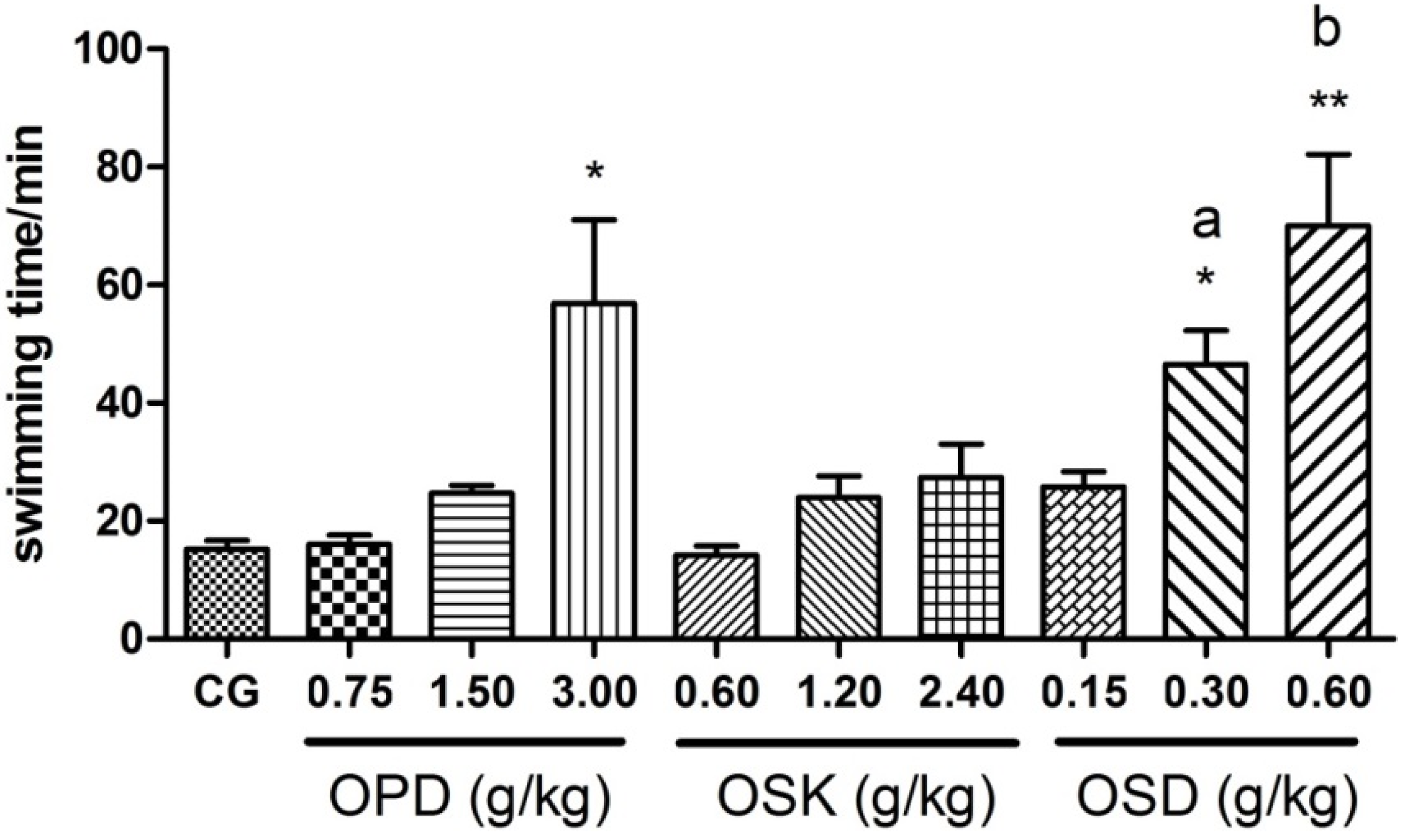
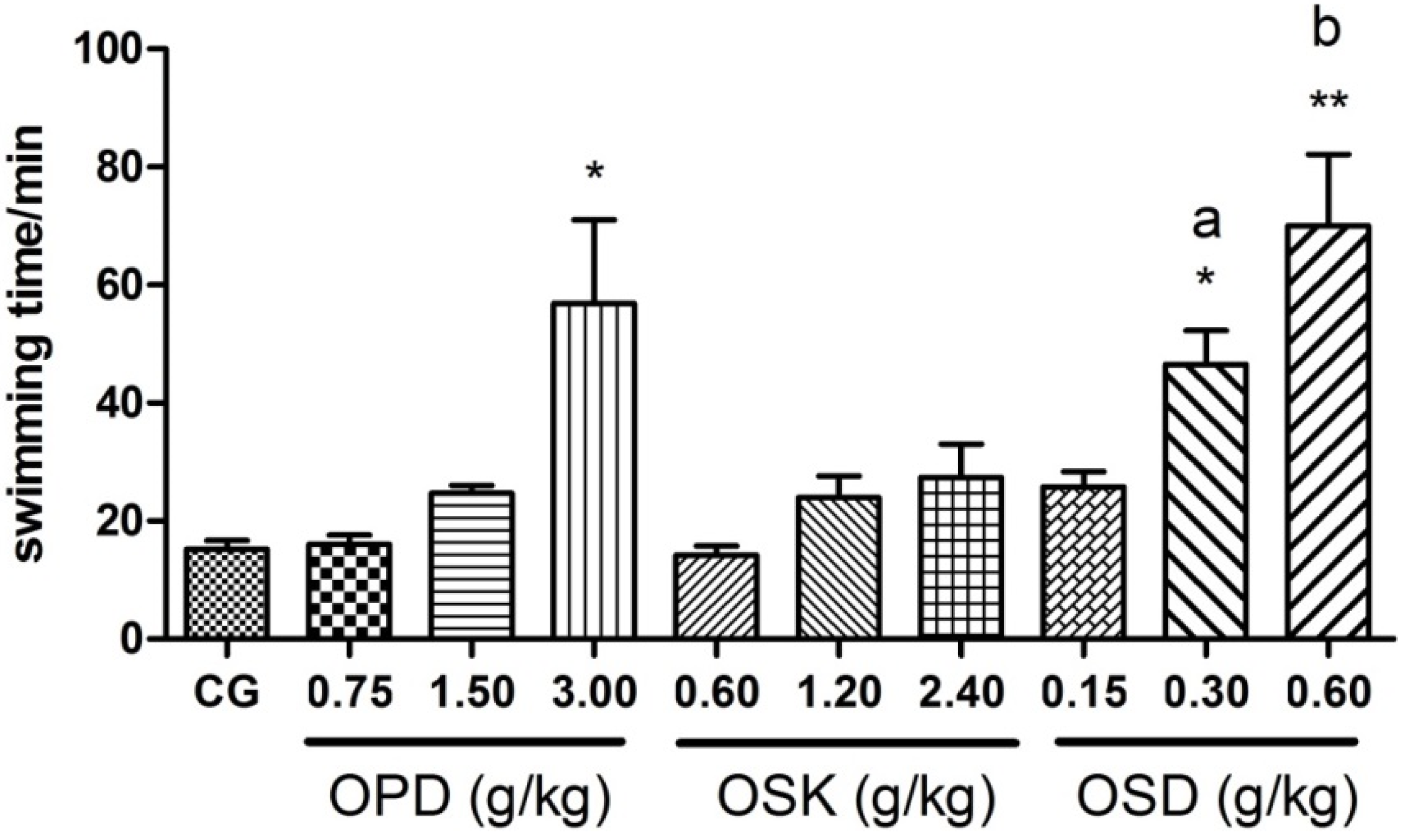
3.4. Effects on the Swimming Exhaustive Time of OPD, OSK and OSD (WLST)
3.5. Effect of OSD on Biochemical Parameters in Mice after Weight-loaded Swimming Test
3.5.1. Effect of OSD on BLA, BUN and HG
| Control Group | OSD Treated Groups | |||
|---|---|---|---|---|
| water | 0.15 (g/kg) | 0.3 (g/kg) | 0.6 (g/kg) | |
| BLA (mmol/L) | 11.24 ± 0.4 | 9.76 ± 0.5 * | 9.46 ± 0.5 * | 8.13 ± 0.4 ** |
| BUN (mmol/L) | 10.72 ± 0.4 | 7.35 ± 0.3 ** | 7.13 ± 0.4 ** | 6.98 ± 0.3 ** |
| HG (mg/g liver) | 9.81 ± 0.9 | 17.3 ± 2.56 * | 18.8 ± 2.43 ** | 14.76 ± 2.02 * |
3.5.2. Effect of OSD on GSH-PX, MDA and SOD
| Control Group | OSD Treated Groups | |||
|---|---|---|---|---|
| water | 0.15 g/kg | 0.3 g/kg | 0.6 g/kg | |
| MDA (nmol/mg protein) | 0.772 ± 0.10 | 0.501 ± 0.02 ** | 0.487 ± 0.03 ** | 0.451 ± 0.03 ** |
| SOD (U/mg protein) | 113 ± 3.5 | 128 ± 4.2 ** | 137 ± 3.7 ** | 159 ± 4.0 ** |
| GSH-PX (U/mg protein) | 63.2 ± 3.6 | 80.4 ± 5.3 * | 86.3 ± 4.8 ** | 98.5 ± 5.6 ** |
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chaudhuri, A.; Behan, P.O. Fatigue in neurological disorders. Lancet 2004, 363, 978–988. [Google Scholar] [CrossRef]
- Belluardo, N.; Westerblad, H.; Mudó, G.; Casabona, A.; Bruton, J.; Caniglia, G.; Pastoris, O.; Grassi, F.; Ibáñez, C.F. Neuromuscular junction disassembly and muscle fatigue in mice lacking neurotrophin-4. Mol. Cell. Neurosci. 2001, 18, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Tharakan, B.; Dhanasekaran, M.; Brown-Borg, H.M.; Manyam, B.V. Trichopus zeylanicus combats fatigue without amphetamine-mimetic activity. Phytother. Res. PTR 2006, 20, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-Z.; Huang, B.-K.; Ye, Q.; Qin, L.-P. Bioactivity-guided fractionation for anti-fatigue property of acanthopanax senticosus. J. Ethnopharmacol. 2011, 133, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowska, T.; Chalder, T.; Hirsch, S.R.; Wallace, P.; Wright, D.J.; Wessely, S.C. Population based study of fatigue and psychological distress. BMJ 1994, 308, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Li, G. The Fatigue Situation of 2823 Healthy People to Check-up and Its Correlation with Physical Indicators; Beijing University of Chinese Medicine: Beijing, China, 2013. [Google Scholar]
- Uehata, T. Karoshi, death by overwork. Nippon Rinsho Jpn. J. Clin. Med. 2005, 63, 1249–1253. [Google Scholar]
- Layzer, R.B. Muscle metabolism during fatigue and work. Bailliere’s Clin. Endocrinol. Metab. 1990, 4, 441–459. [Google Scholar] [CrossRef]
- Hultman, E.; Bergstrom, M.; Spriet, L.L.; Söderlund, K. Energy Metabolism and Fatigue; Human Kinetics: Champaign, IL, USA, 1990. [Google Scholar]
- Glaister, M. Multiple sprint work: Physiological responses, mechanisms of fatigue and the influence of aerobic fitness. Sports Med. 2005, 35, 757–777. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, S.; Mizuma, H.; Tanaka, M.; Jin, G.; Tahara, T.; Mizuno, K.; Yamato, M.; Okuyama, K.; Eguchi, A.; Akimoto, K.; et al. Thiamine tetrahydrofurfuryl disulfide improves energy metabolism and physical performance during physical-fatigue loading in rats. Nutr. Res. 2009, 29, 867–872. [Google Scholar] [CrossRef] [PubMed]
- McCully, K.K.; Authier, B.; Olive, J.; Clark, B.J., 3rd. Muscle fatigue: The role of metabolism. Can. J. Appl. Physiol. 2002, 27, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.T. Fatigue. In Encyclopedia of the Neurological Sciences, 2nd ed.; Aminoff, M.J., Daroff, R.B., Eds.; Academic Press: Oxford, UK, 2014; pp. 276–280. [Google Scholar]
- Filler, K.; Lyon, D.; Bennett, J.; McCain, N.; Elswick, R.; Lukkahatai, N.; Saligan, L.N. Association of mitochondrial dysfunction and fatigue: A review of the literature. BBA Clin. 2014, 1, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Liu, H.-L.; Fang, J.; Yu, C.-H.; Xiong, Y.-K.; Yuan, K. Anti-fatigue and vasoprotective effects of quercetin-3-O-gentiobiose on oxidative stress and vascular endothelial dysfunction induced by endurance swimming in rats. Food Chem. Toxicol. 2014, 68, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.H. Human muscle function and fatigue. Ciba Found. Symp. 1981, 82, 1–18. [Google Scholar] [PubMed]
- Gupta, A.; Vij, G.; Sharma, S.; Tirkey, N.; Rishi, P.; Chopra, K. Curcumin, a polyphenolic antioxidant, attenuates chronic fatigue syndrome in murine water immersion stress model. Immunobiology 2009, 214, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Chen, R.; Wang, X.S.; Shen, B.; Yue, W.; Wu, Q. Antioxidant and anti-fatigue activities of phenolic extract from the seed coat of euryale ferox salisb. And identification of three phenolic compounds by lc-esi-ms/ms. Molecules 2013, 18, 11003–11021. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.-Q.; Guo, Y.; Xu, D.-H.; Huang, Y.-S.; Yuan, K.; Lv, Z.-Q. Antioxidant and anti-fatigue effects of anthocyanins of mulberry juice purification (MJP) and mulberry marc purification (MMP) from different varieties mulberry fruit in china. Food Chem. Toxicol. 2013, 59, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.S.L.; Naveen, S.; Singsit, D.; Naika, M.; Khanum, F. Anti-fatigue effects of polyphenols extracted from pomegranate peel. Int. J. Integr. Biol. 2011, 11, 69–72. [Google Scholar]
- Messing, J.; Thole, C.; Niehues, M.; Shevtsova, A.; Glocker, E.; Boren, T.; Hensel, A. Antiadhesive properties of Abelmoschus esculentus (Okra) immature fruit extract against helicobacter pylori adhesion. PLoS ONE 2014, 9, e84836. [Google Scholar] [CrossRef] [PubMed]
- Tongjaroenbuangam, W.; Ruksee, N.; Chantiratikul, P.; Pakdeenarong, N.; Kongbuntad, W.; Govitrapong, P. Neuroprotective effects of quercetin, rutin and okra (Abelmoschus esculentus linn.) in dexamethasone-treated mice. Neurochem. Int. 2011, 59, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zhang, Y.; Sun, Q.; Yu, L.; Li, M.; Zheng, B.; Wu, X.; Yang, B.; Li, Y.; Huang, C. Extract of Okra lowers blood glucose and serum lipids in high-fat diet-induced obese C57Bl/6 mice. J. Nutr. Biochem. 2014, 25, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, G.; Ren, D.; Yang, S.T. Hypolipidemic activity of Okra is mediated through inhibition of lipogenesis and upregulation of cholesterol degradation. Phytother. Res. PTR 2014, 28, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Yu, W.; Li, Y.; Prasad, N.; Tang, Z. Antioxidant activity of extract and its major constituents from okra seed on rat hepatocytes injured by carbon tetrachloride. BioMed Res. Int. 2014, 2014, 341291. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jin, Z.; Mao, P.; Jin, J.; Huang, J.; Yang, M. Study on anti-fatigue effect of okra extracts. Chin. J. Mod. Appl. Pharm. 2012, 29, 4. [Google Scholar] [CrossRef]
- Arapitsas, P. Identification and quantification of polyphenolic compounds from okra seeds and skins. Food Chem. 2008, 110, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Sengkhamparn, N.; Sagis, L.M.C.; de Vries, R.; Schols, H.A.; Sajjaanantakul, T.; Voragen, A.G.J. Physicochemical properties of pectins from okra (Abelmoschus esculentus (L.) moench). Food Hydrocoll. 2010, 24, 35–41. [Google Scholar] [CrossRef]
- Karakoltsidis, P.A.; Constantinides, S.M. Okra seeds: A new protein source. J. Agric. Food Chem. 1975, 23, 1204–1207. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Jia, X.; Shi, J.; Xu, Y.; Jing, L.; Jia, L. Two new pentacyclic triterpenes from Abelmoschus esculentus. Helv. Chim. Acta 2013, 96, 533–537. [Google Scholar] [CrossRef]
- Jia, L.; Zhong, L.J.; Li, H.F.; Jing, L.L. Chemical constituents in water fraction of Abelmoschus esculentus. Chin. Tradit. Herb. Drugs 2011, 42, 2186–2188. [Google Scholar]
- Jia, L.; Guo, M.; Li, D.; Jing, L. Chemical constituents from petroleum ether portion of Abelmoschus esculentus II. Zhongguo Zhongyao Zazhi 2011, 36, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Li, H.F.; Jing, L.L. Chemical constituents in n-butanol extract of Abelmoschus esculentus L. Chin. Tradit. Herb. Drugs 2010, 41, 1771–1773. [Google Scholar]
- Jia, L.; Li, D.; Jing, L.L.; Guo, M.M. Studies on the chemical constituents from petroleum ether portion of Abelmoschus esculentus. J. Chin. Med. Mater. 2010, 33, 1262–1265. [Google Scholar]
- Kobori, M.; Takahashi, Y.; Akimoto, Y.; Sakurai, M.; Matsunaga, I.; Nishimuro, H.; Ippoushi, K.; Oike, H.; Ohnishi-Kameyama, M. Chronic high intake of quercetin reduces oxidative stress and induces expression of the antioxidant enzymes in the liver and visceral adipose tissues in mice. J. Funct. Foods 2015, 15, 551–560. [Google Scholar] [CrossRef]
- Nogueira, L.; Ramirez-Sanchez, I.; Perkins, G.A.; Murphy, A.; Taub, P.R.; Ceballos, G.; Villarreal, F.J.; Hogan, M.C.; Malek, M.H. (-)-epicatechin enhances fatigue resistance and oxidative capacity in mouse muscle. J. Physiol. 2011, 589, 4615–4631. [Google Scholar] [CrossRef] [PubMed]
- Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the care and use of laboratory animals, 8th ed.; The National Academiec Press: Washington, DC, USA, 2011. [Google Scholar]
- Liao, H.; Dong, W.; Shi, X.; Liu, H.; Yuan, K. Analysis and comparison of the active components and antioxidant activities of extracts from Abelmoschus esculentus L. Pharmacogn. Mag. 2012, 8, 156–161. [Google Scholar] [PubMed]
- Zhu, Z.; Li, N. Study on the content determination method of okra polysaccharide. Jiangsu Agric. Sci. 2012, 40, 2. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Luo, J.; Li, L.; Kong, L. Preparative separation of phenylpropenoid glycerides from the bulbs of lilium lancifolium by high-speed counter-current chromatography and evaluation of their antioxidant activities. Food Chem. 2012, 131, 1056–1062. [Google Scholar] [CrossRef]
- Dang, H.; Chen, Y.; Liu, X.; Wang, Q.; Wang, L.; Jia, W.; Wang, Y. Bed nucleus of the stria terminalis: Anatomy, physiology, functions bed nucleus. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 1417–1424. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Mai, W.L.; Li, Y.H.; Chen, S.G.; Wang, L.W.; Feng, Z.Q.; Zhu, Y.F.; Liu, X.M. Establishment of compute-based image-processing system for animal’s locomotor activity and sedative verification by kaixin powder. Chin. Tradit. Herb. Drugs 2009, 40, 1773–1779. [Google Scholar]
- Qi, B.; Liu, L.; Zhang, H.; Zhou, G.-X.; Wang, S.; Duan, X.-Z.; Bai, X.-Y.; Wang, S.-M.; Zhao, D.-Q. Anti-fatigue effects of proteins isolated from panax quinquefolium. J. Ethnopharmacol. 2014, 153, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Yu, K.Q.; Liu, Y.Y.; Ouyang, M.Z.; Yan, M.H.; Luo, R.; Zhao, X.S. Anti-fatigue activity of polysaccharides extract from radix rehmanniae preparata. Int. J. Biol. Macromol. 2012, 50, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, J.; Tang, G. A study on the effect of anti-fatigue of Okra. Chin. J. Mod. Appl. Pharm. 2003, 20, 2. [Google Scholar] [CrossRef]
- Contarteze, R.V.; Manchado Fde, B.; Gobatto, C.A.; de Mello, M.A. Stress biomarkers in rats submitted to swimming and treadmill running exercises. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.Y. Experimental methodology of pharmacology; People’s Medical Publishing House: Beijing, China, 2002. [Google Scholar]
- Zanwar, A.A.; Badole, S.L.; Shende, P.S.; Hegde, M.V.; Bodhankar, S.L. Antioxidant role of catechin in health and disease. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 267–271. [Google Scholar]
- Jadhav, S.B.; Singhal, R.S. Laccase–gum Arabic conjugate for preparation of water-soluble oligomer of catechin with enhanced antioxidant activity. Food Chem. 2014, 150, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Liudong, F.; Feng, Z.; Daoxing, S.; Xiufang, Q.; Xiaolong, F.; Haipeng, L. Evaluation of antioxidant properties and anti-fatigue effect of green tea polyphenols. Sci. Res. Essays 2011, 6, 2624–2629. [Google Scholar]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. In vitro antioxidant activity and in vivo anti-fatigue effect of loach (misgurnus anguillicaudatus) peptides prepared by papain digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, F.; Zhong, Y.; Li, M.; Chang, Q.; Liao, Y.; Liu, X.; Pan, R. Antioxidant and Anti-Fatigue Constituents of Okra. Nutrients 2015, 7, 8846-8858. https://doi.org/10.3390/nu7105435
Xia F, Zhong Y, Li M, Chang Q, Liao Y, Liu X, Pan R. Antioxidant and Anti-Fatigue Constituents of Okra. Nutrients. 2015; 7(10):8846-8858. https://doi.org/10.3390/nu7105435
Chicago/Turabian StyleXia, Fangbo, Yu Zhong, Mengqiu Li, Qi Chang, Yonghong Liao, Xinmin Liu, and Ruile Pan. 2015. "Antioxidant and Anti-Fatigue Constituents of Okra" Nutrients 7, no. 10: 8846-8858. https://doi.org/10.3390/nu7105435
APA StyleXia, F., Zhong, Y., Li, M., Chang, Q., Liao, Y., Liu, X., & Pan, R. (2015). Antioxidant and Anti-Fatigue Constituents of Okra. Nutrients, 7(10), 8846-8858. https://doi.org/10.3390/nu7105435