An Investigation into the Association between DNA Damage and Dietary Fatty Acid in Men with Prostate Cancer

 ,
, 
Abstract
:1. Introduction
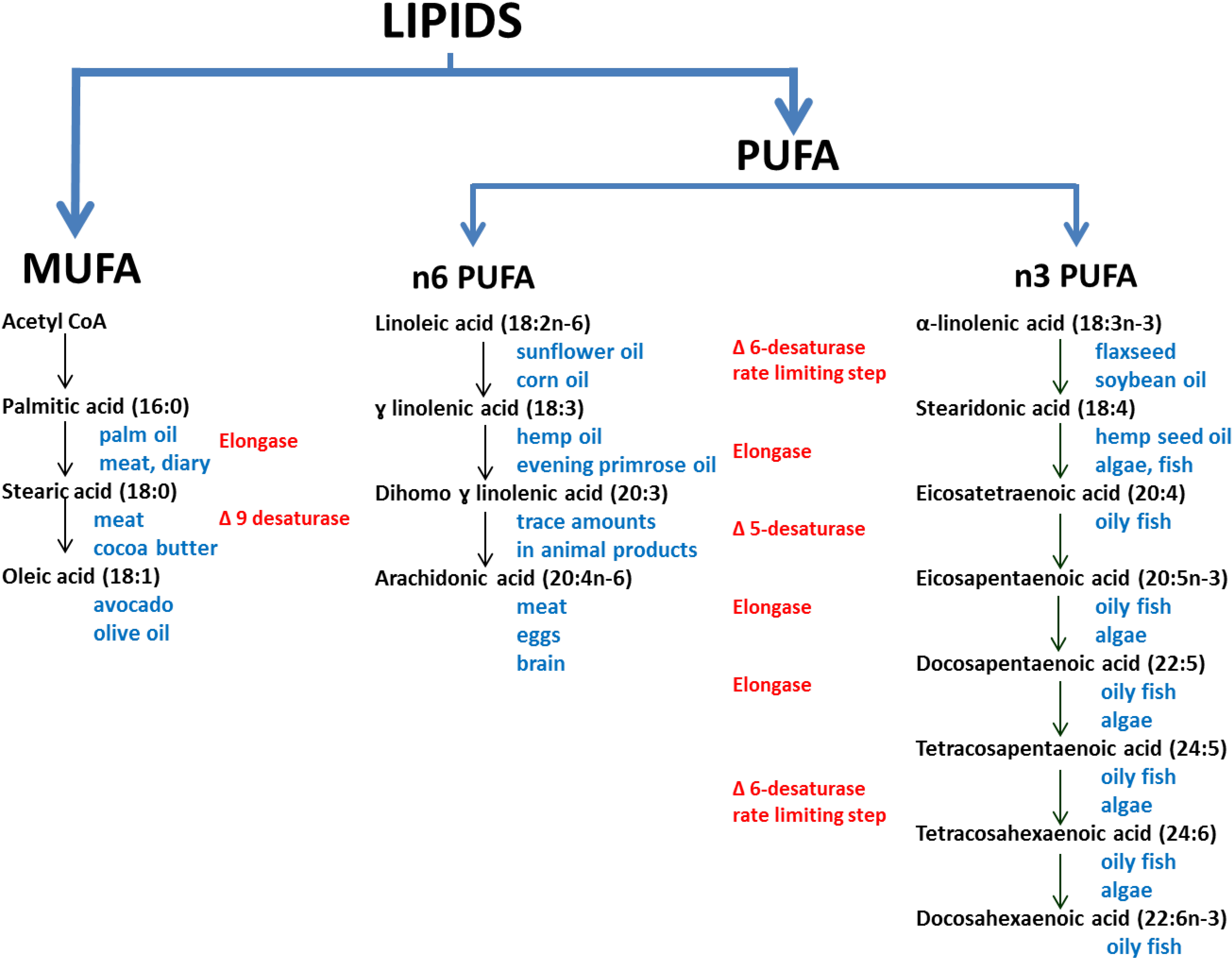
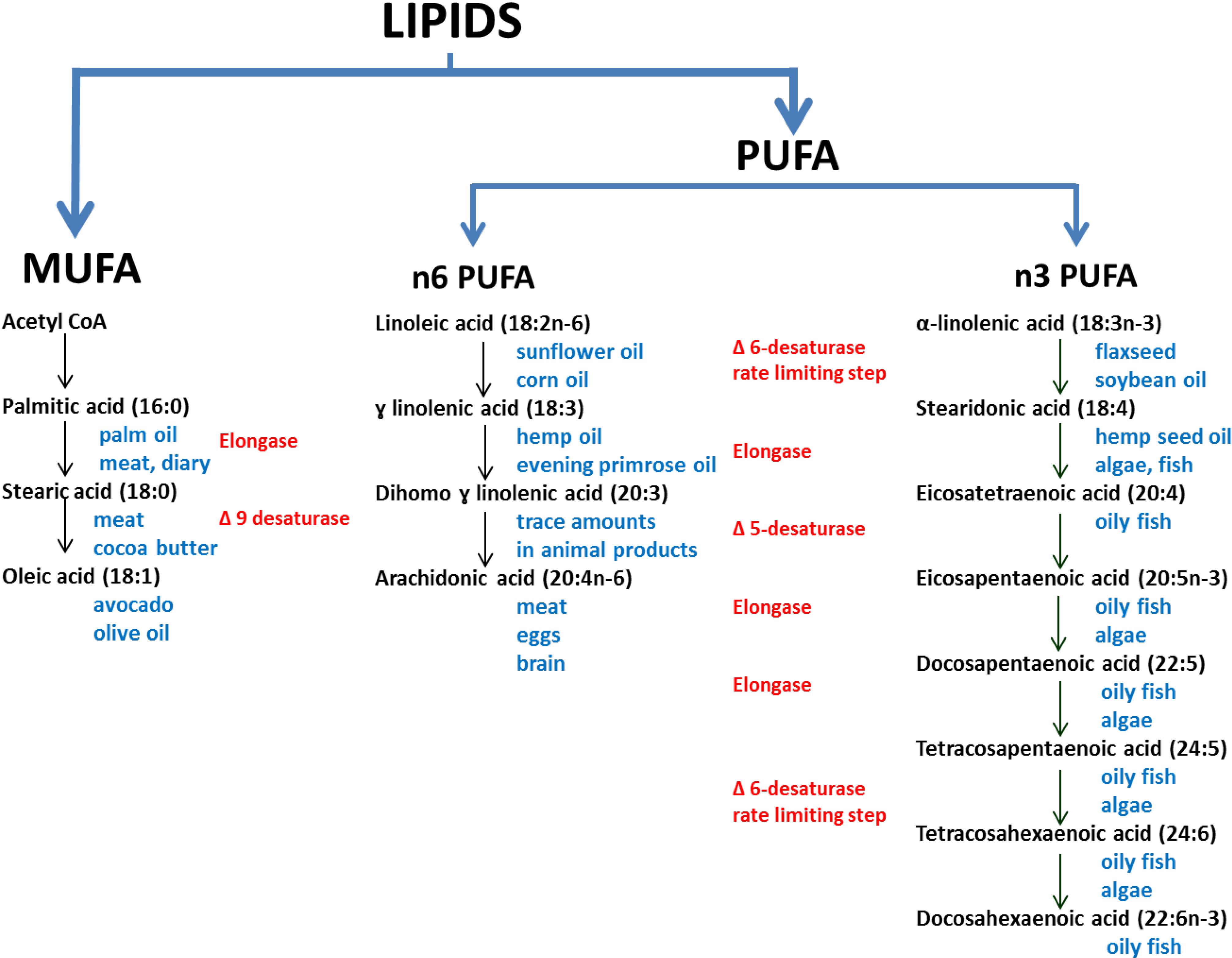
2. Experimental Section
3. Results

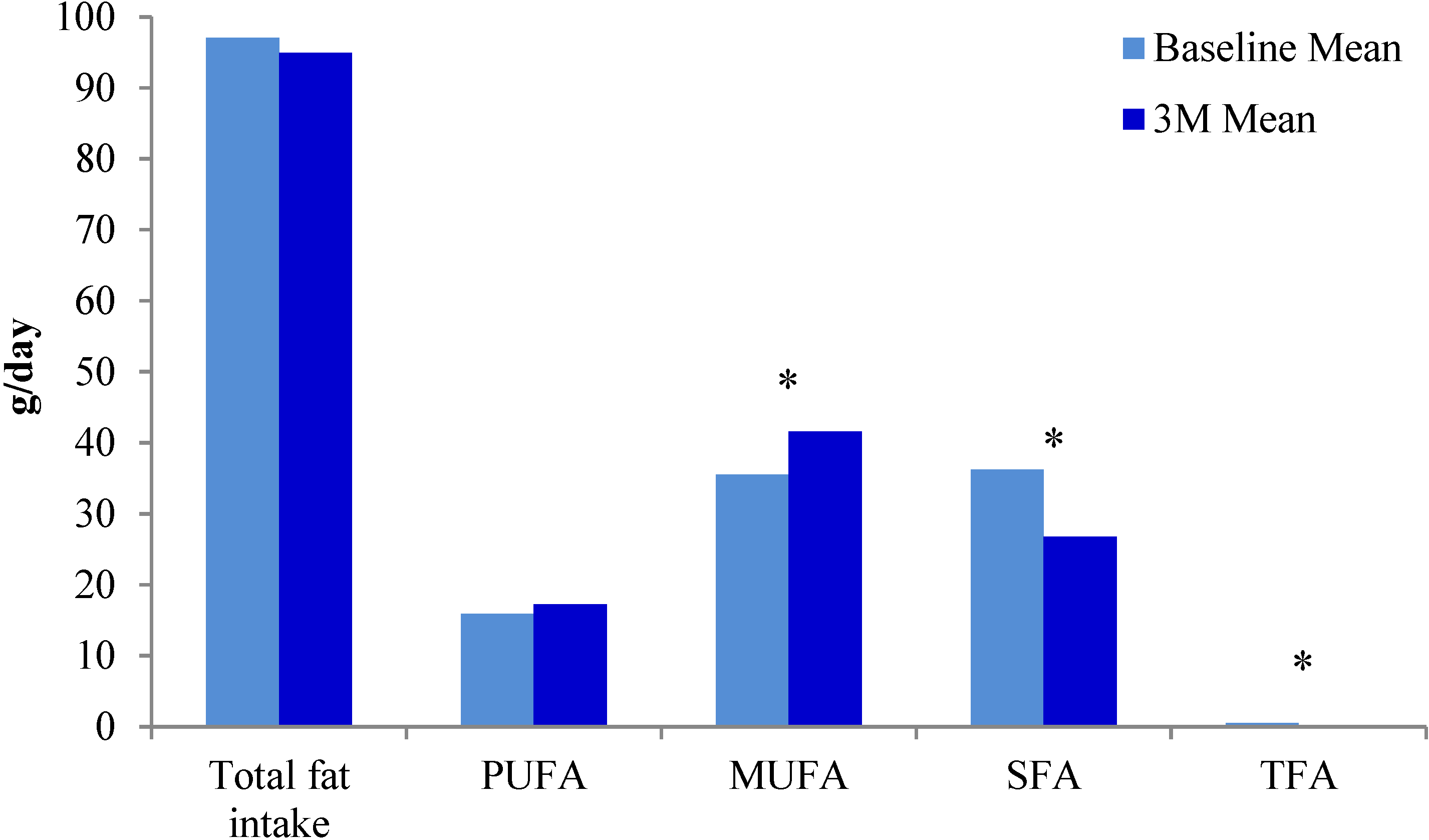
{kind=link}
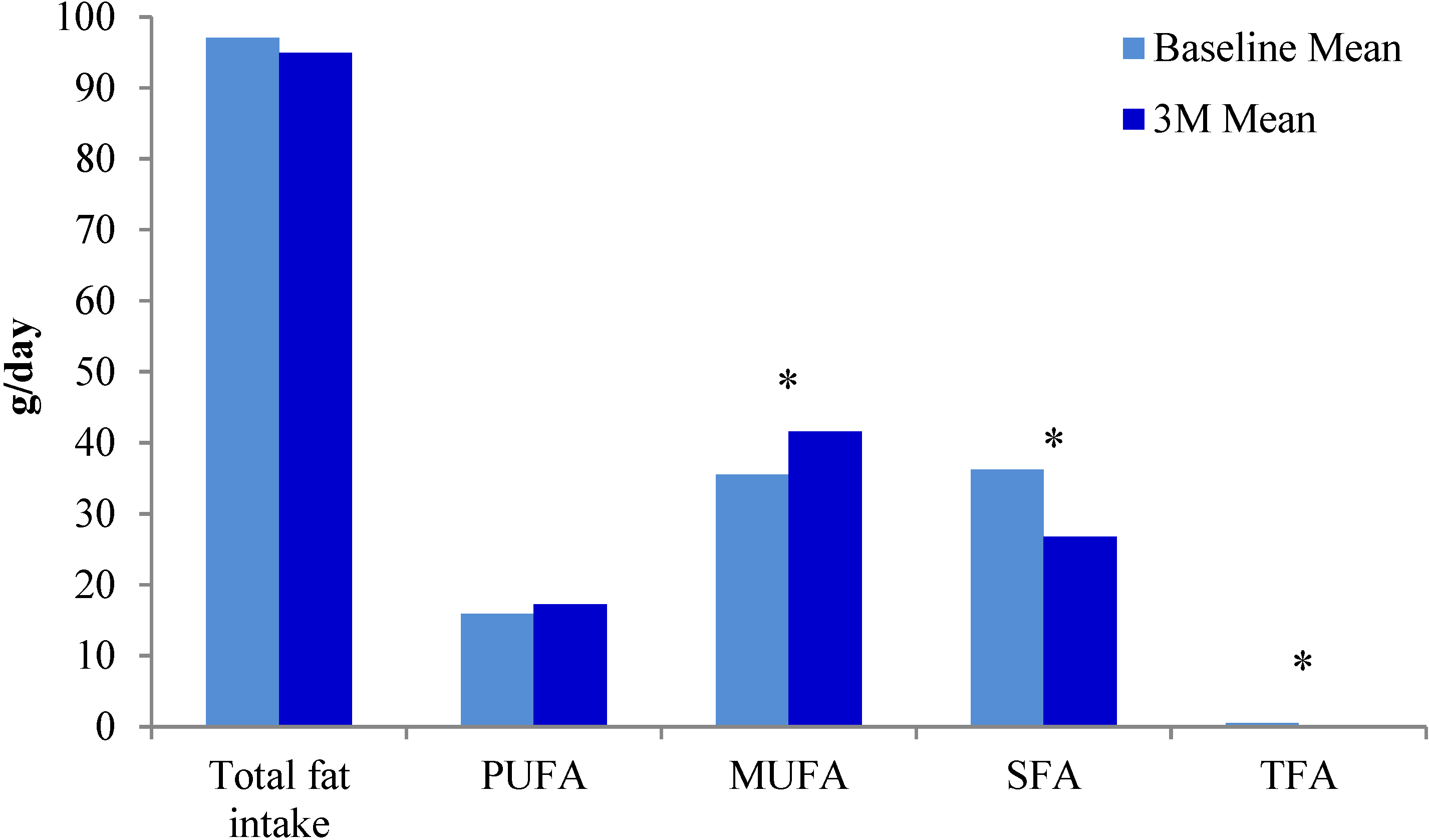
{kind=link}
{kind=link}
| Baseline Characteristics | n | |
|---|---|---|
| Age (years) (range 52–74 years) | 50–59 | 3 |
| 60–69 | 12 | |
| ≥70 | 5 | |
| BMI (kg/m2) (range 23–33 kg/m2) | ||
| ≤19.9 | 0 | |
| 20–24.9 | 4 | |
| 25–29.9 | 12 | |
| ≥30 | 4 | |
| Gleason score * | 3 + 3 | 14 |
| 3 + 4 | 6 | |
| Smoking status | Never | 7 |
| Past | 13 | |
| Present | 0 | |
| Supplements | Omega 3 (from fish oil) | 3 |
| Vitamins | 4 | |
| Treatment type | None | 6 |
| Prostatectomy | 10 | |
| Prostatectomy + ADT + DxR | 1 | |
| Prostatectomy + DxR | 1 | |
| ADT + DxR | 1 | |
| Brachytherapy | 1 | |
| Dietary Component (Unit of Measure) | Mean (SE) | Mean Difference (95% CI) | p | |
|---|---|---|---|---|
| Baseline | Three Months | |||
| Olive oil (mL/day) | 14.5 (3.8) | 28.8 (4.7) | 14.2 (6.8–16.0) | 0.0008 |
| Nuts (Servings/week) | 2.3 (0.5) | 5.1 (0.6) | 2.9 (1.5–4.2) | 0.0003 |
| Butter/cream/margarine (Servings/day) | 2.1 (0.3) | 1.0 (0.3) | −1.1 (−0.6–−1.6) | 0.0002 |
| Dairy products (Servings/week) | 7.4 (0.9) | 4.4 (0.7) | −2.9 (−1.2–−4.7) | 0.0025 |
| Fish (Servings/week) | 1.7 (0.2) | 3.5 (0.5) | 1.8 (0.9–2.7) | 0.0005 |
| Red and processed meat (Servings/week) | 3.9 (0.5) | 1.9 (0.4) | −2.0 (−2.6–−1.3) | 0.0005 |
| Blood Fatty Acids | Mean (SE) | Mean Difference (95% CI) | p | |
|---|---|---|---|---|
| Baseline | Three Months | |||
| Total SFA | 34.7 (0.3) | 33.7 (0.4) | −1.0 (0.4–1.5) | 0.002 |
| 16:0 Palmitic acid | 22.6 (0.3) | 22.3 (0.4) | −0.3 (−0.1–0.7) | 0.161 |
| 18:0 Stearic acid | 10.5 (0.2) | 10.0 (0.2) | −0.5 (0.2–0.9) | 0.002 |
| Total MUFA | 23.4 (0.4) | 23.7 (0.4) | 0.3 (0.4–1.0) | 0.366 |
| Total n9MUFA | 23.1 (0.4) | 23.4 (0.4) | 0.3 (−0.4–1.0) | 0.380 |
| 18:1ω9 Oleic acid | 22.7 (0.4) | 23.2 (0.4) | 0.5 (−0.2–1.1) | 0.162 |
| Total PUFA | 39.5 (0.5) | 40.3 (0.5) | 0.9 (−0.1–1.8) | 0.079 |
| Total n6PUFA | 32.8 (0.4) | 33.0 (0.5) | 0.2 (−0.7–1.2) | 0.636 |
| 18:2ω6 LA | 19.6 (0.7) | 19.4 (0.9) | −0.2 (−1.7–1.4) | 0.832 |
| 20:4ω6 AA | 9.1 (0.3) | 8.9 (0.3) | −0.2 (−0.7–0.3) | 0.379 |
| Total n3PUFA | 6.6 (0.4) | 7.3 (0.3) | 0.6 (−0.0–1.3) | 0.057 |
| 18:3ω3 LNA | 0.5 (0.0) | 0.6 (0.1) | 0.0 (−0.1–0.2) | 0.689 |
| 20:5ω3 EPA | 1.4 (0.9) | 1.5 (0.7) | 0.1 (−0.2–0.5) | 0.463 |
| 22:6ω3 DHA | 3.0 (0.9) | 3.5 (0.1) | 0.5 (0.2–0.8) | 0.001 |
| EPA + DHA | 4.4 (0.4) | 5.0 (0.2) | 0.6 (0.3–1.2) | 0.042 |
| Modified WBS n3 Index | 6.1 (0.5) | 7.0 (0.3) | 0.9 (0.0–1.7) | 0.043 |
| n6PUFA:n3PUFA | 5.2 (0.3) | 4.7 (0.2) | −0.6 (−1.0–−0.1) | 0.019 |
| AA:EPA | 8.58 (0.9) | 6.9 (0.6) | −1.6 (−3.1–−0.2) | 0.030 |
| Blood Fatty Acids | Dietary Fat Source | Correlation | p |
|---|---|---|---|
| Total n3PUFA | Fish intake | 0.210 | 0.374 |
| Nut intake | 0.341 | 0.141 | |
| Dairy intake | −0.433 | 0.057 | |
| Red meat intake | 0.082 | 0.732 | |
| EPA | Fish intake | 0.172 | 0.468 |
| Nut intake | 0.147 | 0.535 | |
| Dairy intake | −0.580 | 0.007 | |
| Red meat intake | −0.475 | 0.034 | |
| EPA + DHA | Fish intake | 0.123 | 0.605 |
| Nut intake | 0.222 | 0.347 | |
| Dairy intake | −0.609 | 0.004 | |
| Red meat intake | 0.055 | 0.817 | |
| n6PUFA:n3PUFA | Fish intake | 0.192 | 0.418 |
| Nut intake | −0.349 | 0.132 | |
| Dairy intake | −0.147 | 0.537 | |
| Red meat intake | 0.486 | 0.029 | |
| AA:EPA | Fish intake | 0.233 | 0.323 |
| Nut intake | 0.084 | 0.725 | |
| Dairy intake | 0.409 | 0.073 | |
| Red meat intake | −0.029 | 0.904 |
| Outcome of Interest | Dietary Fat Sources | Baseline | Three Months | ||
|---|---|---|---|---|---|
| Correlation | p | Correlation | p | ||
| DNA damage | Olive oil | 0.002 | 0.995 | −0.370 | 0.109 |
| Servings of butter, cream, margarine | 0.278 | 0.235 | 0.456 | 0.043 | |
| Servings of fish | 0.202 | 0.393 | 0.510 | 0.829 | |
| Servings of red meat | 0.066 | 0.783 | 0.576 | 0.007 | |
| Blood Fatty Acid | Total MUFA | 0.200 | 0.3988 | −0.565 | 0.009 |
| Total n9MUFA | 0.211 | 0.371 | −0.561 | 0.010 | |
| Oleic acid | 0.220 | 0.352 | −0.514 | 0.020 | |
| Total n6PUFA | −0.116 | 0.627 | 0.536 | 0.015 | |
| Total n3PUFA | −0.314 | 0.178 | −0.224 | 0.342 | |
| n6PUFA:n3PUFA ratio | 0.330 | 0.155 | 0.507 | 0.023 | |

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cancer: New Registrations and Deaths 2010. Available online: http://www.health.govt.nz/publication/cancer-new-registrations-and-deaths-2010 (accessed on 22 August 2014).
- Bishop, K.S.; Chi, H.-J.K.; Han, D.Y.; Ferguson, L.R. Prostate cancer prevention in the developing world—What are we waiting for? Curr. Pharmacogenomics Pers. Med. 2012, 10, 70–86. [Google Scholar] [CrossRef]
- Itsiopoulos, C.; Hodge, A.; Kaimakamis, M. Can the Mediterranean diet prevent prostate cancer? Mol. Nutr. Food Res. 2009, 53, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C.; John, S.M.; Schmitz, G. Over-stimulation of insulin/IGF-1 signaling by western diet may promote diseases of civilization: Lessons learnt from Laron syndrome. Nutr. Metab. 2011, 8, 41. [Google Scholar] [CrossRef]
- Serra-Majem, L.; de la Cruz, J.N.; Ribas, L.; Salleras, L. Mediterranean diet and health: Is all the secret in olive oil? Pathophysiol. Haemost. Thromb. 2003, 33, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Breen, A.P.; Murphy, J.A. Reactions of oxyl radicals with DNA. Free Radic. Biol. Med. 1995, 18, 1033–1077. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, T.; Wakabayashi, K.; Nakagama, H.; Nagao, M. Heterocyclic amines: Mutagens/carcinogens produced during cooking of meat and fish. Cancer Sci. 2004, 95, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Norrish, A.E.; Ferguson, L.R.; Knize, M.G.; Felton, J.S.; Sharpe, S.J.; Jackson, R.T. Heterocyclic amine content of cooked meat and risk of prostate cancer. J. Natl. Cancer Inst. 1999, 91, 2038–2044. [Google Scholar] [CrossRef] [PubMed]
- John, E.M.; Stern, M.C.; Sinha, R.; Koo, J. Meat consumption, cooking practices, meat mutagens, and risk of prostate cancer. Nutr. Cancer 2011, 63, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.D.; Corral, R.; Catsburg, C.; Lewinger, J.-P.; Koo, J.; John, E.M.; Ingles, S.; Stern, M.C. Red meat and poultry, cooking practices, genetic susceptibility and risk of prostate cancer: Results from the California Collaborative Prostate Cancer Study. Carcinogenesis 2012, 33, 2108–2118. [Google Scholar] [CrossRef]
- Van Hemelrijck, M.; Rohrmann, S.; Steinbrecher, A.; Kaaks, R.; Teucher, B.; Linseisen, J. Heterocyclic aromatic amine (HCA) intake and prostate cancer risk: Effect modification by genetic variants. Nutr. Cancer 2012, 64, 704–713. [Google Scholar]
- Zemel, M.B.; Sun, X. Dietary calcium and dairy products modulate oxidative and inflammatory stress in mice and humans. J. Nutr. 2008, 138, 1047–1052. [Google Scholar] [PubMed]
- Ganmaa, D.; Li, X.M.; Wang, J.; Qin, L.Q.; Wang, P.Y.; Sato, A. Incidence and mortality of testicular and prostatic cancers in relation to world dietary practices. Int. J. Cancer 2002, 98, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Torfadottir, J.E.; Steingrimsdottir, L.; Mucci, L.; Aspelund, T.; Kasperzyk, J.L.; Olafsson, O.; Fall, K.; Tryggvadottir, L.; Harris, T.B.; Launer, L.; et al. Milk intake in early life and risk of advanced prostate cancer. Am. J. Epidemiol. 2012, 175, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, A.; Kasperzyk, J.L.; Kenfield, S.A.; Richman, E.L.; Chan, J.M.; Willett, W.C.; Stampfer, M.J.; Mucci, L.A.; Giovannucci, E.L. Milk and dairy consumption among men with prostate cancer and risk of metastases and prostate cancer death. Cancer Epidemiol. Biomark. Prev. 2012, 21, 428–436. [Google Scholar] [CrossRef]
- Giovannucci, E.; Rimm, E.B.; Colditz, G.A.; Stampfer, M.J.; Ascherio, A.; Chute, C.G.; Willett, W.C. A prospective study of dietary fat and risk of prostate cancer. J. Natl. Cancer Inst. 1993, 85, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Salem, S.; Salahi, M.; Mohseni, M.; Ahmadi, H.; Mehrsai, A.; Jahani, Y.; Pourmand, G. Major dietary factors and prostate cancer risk: A prospective multicenter case-control study. Nutr. Cancer 2011, 63, 21–27. [Google Scholar] [PubMed]
- Pelser, C.; Mondul, A.M.; Hollenbeck, A.R.; Park, Y. Dietary fat, fatty acids, and risk of prostate cancer in the NIH-AARP diet and health study. Cancer Epidemiol. Biomark. Prev. 2013, 22, 697–707. [Google Scholar] [CrossRef]
- Ohwaki, K.; Endo, F.; Kachi, Y.; Hattori, K.; Muraishi, O.; Nishikitani, M.; Yano, E. Relationship between dietary factors and prostate-specific antigen in healthy men. Urol. Int. 2012, 89, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Erkkilä, A.; de Mello, V.D.F.; Risérus, U.; Laaksonen, D.E. Dietary fatty acids and cardiovascular disease: An epidemiological approach. Prog. Lipid Res. 2008, 47, 172–187. [Google Scholar] [CrossRef] [PubMed]
- White, H.M.; Richert, B.T.; Latour, M.A. Impacts of Nutrition and Environmental Stressors on Lipid Metabolism. In Lipid Metabolism; Baeze, R.V., Ed.; InTech: Rijeka, Croatia, 2013; Chapter 10. [Google Scholar] [CrossRef]
- Fatty Acids: Methylene-Interrupted Double Bonds. Available online: http://lipidlibrary.aocs.org/Lipids/fa_poly/index.htm (accessed on 22 August 2014).
- Lenihan-Geels, G.; Bishop, K.S.; Ferguson, L.R. Alternative sources of omega-3 fats: Can we find a sustainable substitute for fish? Nutrients 2013, 5, 1301–1315. [Google Scholar] [CrossRef] [PubMed]
- Plourde, M.; Cunnane, S.C. Extremely limited synthesis of long chain polyunsaturates in adults: Implications for their dietary essentiality and use as supplements. Appl. Physiol. Nutr. Metab. 2007, 32, 619–634. [Google Scholar] [CrossRef] [PubMed]
- Plourde, M.; Fortier, M.; Vandal, M.; Tremblay-Mercier, J.; Freemantle, E.; Begin, M.; Pifferi, F.; Cunnane, S.C. Unresolved issues in the link between docosahexaenoic acid and Alzheimer’s disease. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 301–308. [Google Scholar] [CrossRef]
- Barcelo-Coblijn, G.; Murphy, E.J. Alpha-linolenic acid and its conversion to longer chain n-3 fatty acids: Benefits for human health and a role in maintaining tissue n-3 fatty acid levels. Prog. Lipid Res. 2009, 48, 355–374. [Google Scholar] [CrossRef] [PubMed]
- Richman, E.; Kenfield, S.A.; Chavarro, J.E.; Stampfer, M.J.; Giovannucci, E.L.; Willett, W.C.; Chan, J.M. Fat intake after diagnosis and risk of lethal prostate cancer and all-cause mortality. JAMA 2013, 173, 1–8. [Google Scholar]
- Blein, S.; Berndt, S.; Joshi, A.D.; Campa, D.; Ziegler, R.G.; Riboli, E.; Cox, D.G.; Gaudet, M.M.; Stevens, V.L.; Diver, W.R.; et al. Factors associated with oxidative stress and cancer risk in the breast and prostate cancer cohort consortium. Free Radic. Res. 2014, 48, 380–386. [Google Scholar] [CrossRef]
- Lockett, K.L.; Hall, M.C.; Clark, P.E.; Chuang, S.C.; Robinson, B.; Lin, H.Y.; Su, L.J.; Hu, J.J. DNA damage levels in prostate cancer cases and controls. Carcinogenesis 2006, 27, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, R.; Pandey, M.; Bhaskaran, N.; Maclennan, G.T.; Fu, P.; Ponsky, L.E.; Gupta, S. Protection against oxidative DNA damage and stress in human prostate by glutathione S-transferase P1. Mol. Carcinog. 2014, 53, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Wasson, G.R.; McKelvey-Martin, V.J.; Downes, C.S. The use of the comet assay in the study of human nutrition and cancer. Mutagenesis 2008, 23, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; McNutt, M.A.; Zhu, W.G. The comet assay: A sensitive method for detecting DNA damage in individual cells. Methods 2009, 48, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Erdrich, S.; Bishop, K. The Modified Mediterranean Diet for Men: Recipe Collection. Available online: https://cdn.auckland.ac.nz/assets/fmhs/sms/nutrition/pcd/docs/recipes.pdf (accessed on 16 June 2014).
- Martinez-Gonzalez, M.A.; Garcia-Arellano, A.; Toledo, E.; Salas-Salvado, J.; Buil-Cosiales, P.; Corella, D.; Covas, M.I.; Schroder, H.; Aros, F.; Gomez-Gracia, E.; et al. A 14-item Mediterranean diet assessment tool and obesity indexes among high-risk subjects: The PREDIMED trial. PLoS One 2012. [Google Scholar] [CrossRef]
- Dhillon, V.S.; Thomas, P.; Fenech, M. Comparison of DNA damage and repair following radiation challenge in buccal cells and lymphocytes using single-cell gel electrophoresis. Int. J. Radiat. Biol. 2004, 80, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Olive, P.L.; Banath, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Karunasinghe, N.; Ryan, J.; Tuckey, J.; Masters, J.; Jamieson, M.; Clarke, L.C.; Marshall, J.R.; Ferguson, L.R. DNA stability and serum selenium levels in a high-risk group for prostate cancer. Cancer Epidemiol. Biomark. Prev. 2004, 13, 391–397. [Google Scholar]
- Ferguson, L.R.; Han, D.Y.; Fraser, A.G.; Huebner, C.; Lam, W.J.; Morgan, A.R.; Duan, H.; Karunasinghe, N. Genetic factors in chronic inflammation: Single nucleotide polymorphisms in the STAT-JAK pathway, susceptibility to DNA damage and Crohn’s disease in a New Zealand population. Mutat. Res. 2010, 690, 108–115. [Google Scholar] [CrossRef]
- Rise, P.; Eligini, S.; Ghezzi, S.; Colli, S.; Galli, C. Fatty acid composition of plasma, blood cells and whole blood: Relevance for the assessment of the fatty acid status in humans. Prostaglandins Leukot. Essent. Fat. Acids 2007, 76, 363–369. [Google Scholar] [CrossRef]
- Sun, Q.; Ma, J.; Campos, H.; Hankinson, S.E.; Hu, F.B. Comparison between plasma and erythrocyte fatty acid content as biomarkers of fatty acid intake in US women. Am. J. Clin. Nutr. 2007, 86, 74–81. [Google Scholar] [PubMed]
- Katan, M.B.; Deslypere, J.P.; van Birgelen, A.P.; Penders, M.; Zegwaard, M. Kinetics of the incorporation of dietary fatty acids into serum cholesteryl esters, erythrocyte membranes, and adipose tissue: An 18-month controlled study. J. Lipid Res. 1997, 38, 2012–2022. [Google Scholar] [PubMed]
- Katan, M.B.; Grundy, S.M.; Willett, W.C. Should a low-fat, high-carbohydrate diet be recommended for everyone? Beyond low-fat diets. N. Engl. J. Med. 1997, 337, 563–566; discussion 566–567. [Google Scholar] [PubMed]
- Harris, W.S. The omega-3 index: Clinical utility for therapeutic intervention. Curr. Cardiol. Rep. 2010, 12, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Hall, E.; Nelson, E.B.; Ryan, A.S. Validation of a rapid measure of blood PUFA levels in humans. Lipids 2008, 43, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Bibus, D.M.; Lipid Technologies, LLC, Austin, MN, USA. Personal communication, 2014.
- Harris, W.S. The omega-3 index as a risk factor for coronary heart disease. Am. J. Clin. Nutr. 2008, 87, 1997S–2002S. [Google Scholar] [PubMed]
- Gann, P.H.; Hennekens, C.H.; Sacks, F.M.; Grodstein, F.; Giovannucci, E.L.; Stampfer, M.J. Prospective study of plasma fatty acids and risk of prostate cancer. J. Natl. Cancer Inst. 1994, 86, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Folsom, A.R.; Shahar, E.; Eckfeldt, J.H. Plasma fatty acid composition as an indicator of habitual dietary fat intake in middle-aged adults. The Atherosclerosis Risk in Communities (ARIC) Study Investigators. Am. J. Clin. Nutr. 1995, 62, 564–571. [Google Scholar] [PubMed]
- Saadatian-Elahi, M.; Slimani, N.; Chajes, V.; Jenab, M.; Goudable, J.; Biessy, C.; Ferrari, P.; Byrnes, G.; Autier, P.; Peeters, P.H.; et al. Plasma phospholipid fatty acid profiles and their association with food intakes: Results from a cross-sectional study within the European Prospective Investigation into Cancer and Nutrition. Am. J. Clin. Nutr. 2009, 89, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Holub, B.J.; Wlodek, M.; Rowe, W.; Piekarski, J. Correlation of omega-3 levels in serum phospholipid from 2053 human blood samples with key fatty acid ratios. Nutr. J. 2009, 8, 58. [Google Scholar] [CrossRef] [PubMed]
- Alhazmi, A.; Stojanovski, E.; Garg, M.L.; McEvoy, M. Fasting whole blood fatty acid profile and risk of type 2 diabetes in adults: A nested case control study. PLoS One 2014. [Google Scholar] [CrossRef]
- Surette, M.E. Dietary omega-3 PUFA and health: Stearidonic acid-containing seed oils as effective and sustainable alternatives to traditional marine oils. Mol. Nutr. Food Res. 2013, 57, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Leitzmann, M.F.; Stampfer, M.J.; Michaud, D.S.; Augustsson, K.; Colditz, G.C.; Willett, W.C.; Giovannucci, E.L. Dietary intake of n-3 and n-6 fatty acids and the risk of prostate cancer. Am. J. Clin. Nutr. 2004, 80, 204–216. [Google Scholar] [PubMed]
- Collins, A. The comet assay for DNA damage and repair. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Waters, D.J.; Shen, S.; Glickman, L.T.; Cooley, D.M.; Bostwick, D.G.; Qian, J.; Combs, G.F.; Morris, J.S. Prostate cancer risk and DNA damage: Translational significance of selenium supplementation in a canine model. Carcinogenesis 2005, 26, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Mroz, R.M.; Schins, R.P.F.; Li, H.; Jimenez, L.A.; Drost, E.M.; Holownia, A.; MacNee, W.; Donaldson, K. Nanoparticle-driven DNA damage mimics irradiation-related carcinogenesis pathways. Eur. Respir. J. 2008, 31, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Karanika, S.; Karantanos, T.; Li, L.; Corn, P.G.; Thompson, T.C. DNA damage response and prostate cancer: Defects, regulation and therapeutic implications. Oncogene 2014. [Google Scholar] [CrossRef]
- Machowetz, A.; Poulsen, H.E.; Gruendel, S.; Weimann, A.; Fito, M.; Marrugat, J.; de la Torre, R.; Salonen, J.T.; Nyyssonen, K.; Mursu, J.; et al. Effect of olive oils on biomarkers of oxidative DNA stress in Northern and Southern Europeans. FASEB J. 2007, 21, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Colomer, R.; Menendez, J.A. Mediterranean diet, olive oil and cancer. Clin. Transl. Oncol. 2006, 8, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Weinbrenner, T.; Fito, M.; de la Torre, R.; Saez, G.T.; Rijken, P.; Tormos, C.; Coolen, S.; Albaladejo, M.F.; Abanades, S.; Schroder, H.; et al. Olive oils high in phenolic compounds modulate oxidative/antioxidative status in men. J. Nutr. 2004, 134, 2314–2321. [Google Scholar] [PubMed]
- Mitjavila, M.T.; Fandos, M.; Salas-Salvado, J.; Covas, M.I.; Borrego, S.; Estruch, R.; Lamuela-Raventos, R.; Corella, D.; Martinez-Gonzalez, M.A.; Sanchez, J.M.; et al. The Mediterranean diet improves the systemic lipid and DNA oxidative damage in metabolic syndrome individuals. A randomized, controlled, trial. Clin. Nutr. 2013, 32, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Norrish, A.E.; Skeaff, C.M.; Arribas, G.L.; Sharpe, S.J.; Jackson, R.T. Prostate cancer risk and consumption of fish oils: A dietary biomarker-based case-control study. Br. J. Cancer 1999, 81, 1238–1242. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bishop, K.S.; Erdrich, S.; Karunasinghe, N.; Han, D.Y.; Zhu, S.; Jesuthasan, A.; Ferguson, L.R. An Investigation into the Association between DNA Damage and Dietary Fatty Acid in Men with Prostate Cancer. Nutrients 2015, 7, 405-422. https://doi.org/10.3390/nu7010405
Bishop KS, Erdrich S, Karunasinghe N, Han DY, Zhu S, Jesuthasan A, Ferguson LR. An Investigation into the Association between DNA Damage and Dietary Fatty Acid in Men with Prostate Cancer. Nutrients. 2015; 7(1):405-422. https://doi.org/10.3390/nu7010405
Chicago/Turabian StyleBishop, Karen S., Sharon Erdrich, Nishi Karunasinghe, Dug Yeo Han, Shuotun Zhu, Amalini Jesuthasan, and Lynnette R. Ferguson. 2015. "An Investigation into the Association between DNA Damage and Dietary Fatty Acid in Men with Prostate Cancer" Nutrients 7, no. 1: 405-422. https://doi.org/10.3390/nu7010405
APA StyleBishop, K. S., Erdrich, S., Karunasinghe, N., Han, D. Y., Zhu, S., Jesuthasan, A., & Ferguson, L. R. (2015). An Investigation into the Association between DNA Damage and Dietary Fatty Acid in Men with Prostate Cancer. Nutrients, 7(1), 405-422. https://doi.org/10.3390/nu7010405