Effect of Fructooligosaccharides Fraction from Psacalium decompositum on Inflammation and Dyslipidemia in Rats with Fructose-Induced Obesity

 ,
, 

Abstract
:1. Introduction
2. Experimental Section
2.1. Plant Material
2.2. Isolation of FOS-Fraction from the Roots of P. decompositum
2.3. HPTLC Procedures
2.4. Nuclear Magnetic Resonance (NMR) Analysis of the FOS Fraction
2.5. Animals and Diets
2.6. Oral Glucose Tolerance Test (OGTT)
2.7. Biochemical Parameters
2.8. Cytokines Quantification
2.9. Statistical Analysis
3. Results

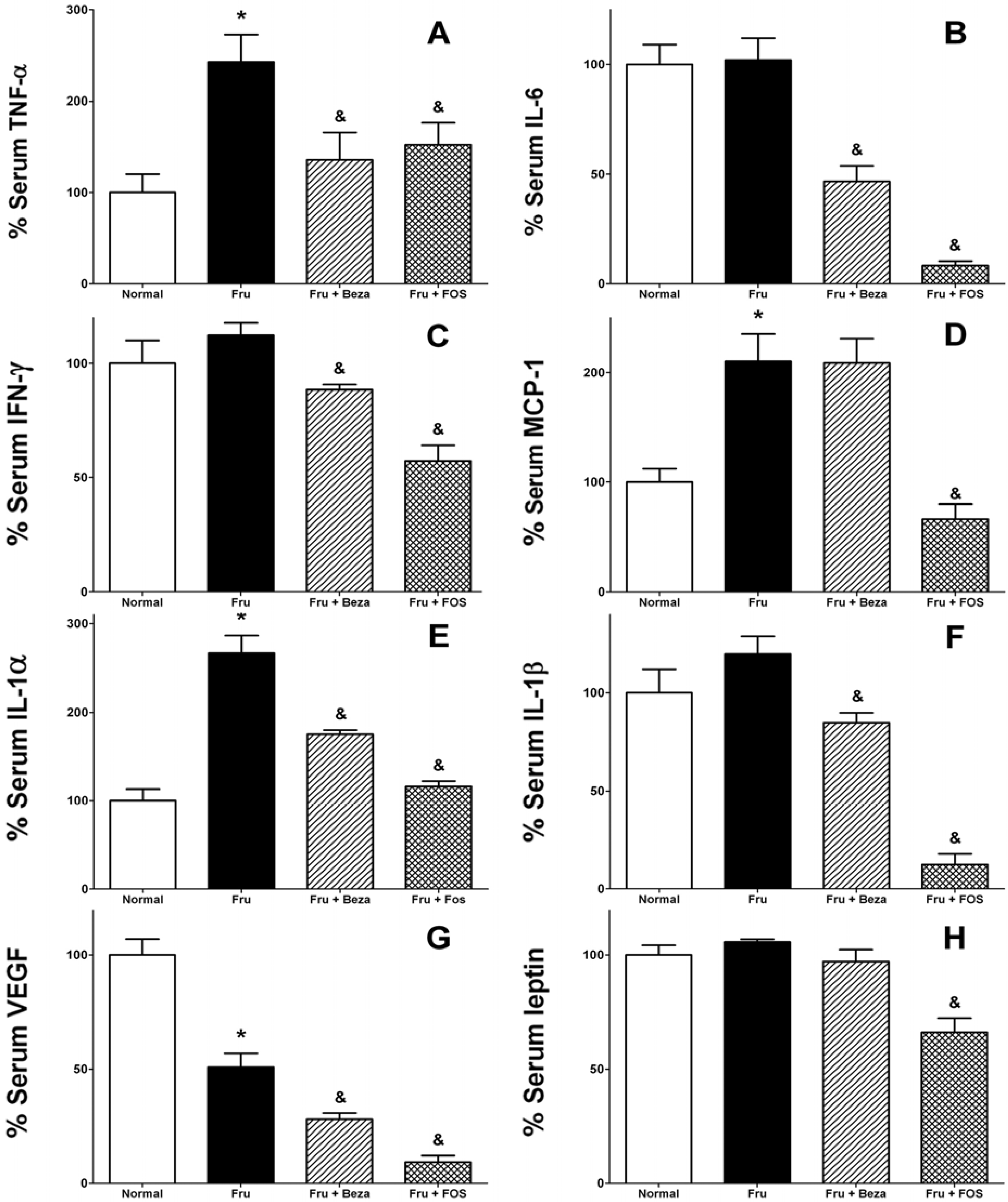{kind=link}
| Normal | Fru | |
|---|---|---|
| Body weight (g) | 307.8 ± 5.6 | 378.5 ± 10.2 * |
| Lee index | 0.28 ± 0.001 | 0.30 ± 0.002 * |
| Glycaemia in fasting (mg/dL) | 101.8 ± 2.5 | 94.6 ± 4.1 |
| AUC of glucose tolerance test (mg/dL/120 min) | 16,800 ± 984 | 18,000 ± 696 |
| Chol (mg/dL) | 100 ± 0.0 | 100 ± 0.0 |
| TG (mg/dL) | 70 ± 0.00 | 189 ± 21.8 * |
| ALT (UI/L) | 5.08 ± 2.6 | 19.3 ± 8.0 * |
| AST (UI/L) | 10.6 ± 2.8 | 12.3 ± 8.5 |
| Normal | Fru | Fru + Beza | Fru + FOS | |
|---|---|---|---|---|
| Final body weight (g) | 490 ± 27.8 | 610 ± 9.1 * | 463 ± 46.0 & | 472 ± 15.8 & |
| Lee index | 0.295 ± 0.003 | 0.33 ± 0.000 * | 0.303 ± 0.002 & | 0.303 ± 0.002 & |
| Glycaemia in fasting (mg/dL) | 96 ± 2.4 | 106.7 ± 2.9 | 100.7 ± 2.8 | 99 ± 2.6 |
| AUC of glucose tolerance test mg/dL/120 min | 16,560 ± 612 | 19,920 ± 1128 * | 18,000 ± 984 | 18,360 ± 816 |
| Chol (mg/dL) | 106 ± 4.0 | 128.5 ± 1.5 * | 101.7 ± 1.7 & | 102.7 ± 2.7 & |
| TG (mg/dL) | 79.5 ± 7.6 | 205.2 ± 34.4 * | 82.5 ± 11.8 & | 95.5 ± 19.6 & |
| ALT (UI/L) | 3.6 ± 0.8 | 30.2 ± 3.4 * | 18.8 ± 7.9 | 12 ± 5.0 |
| AST (UI/L) | 3.2 ± 0.5 | 38.7 ± 5.1 * | 22.7 ± 10.2 | 15.7 ± 7.3 |
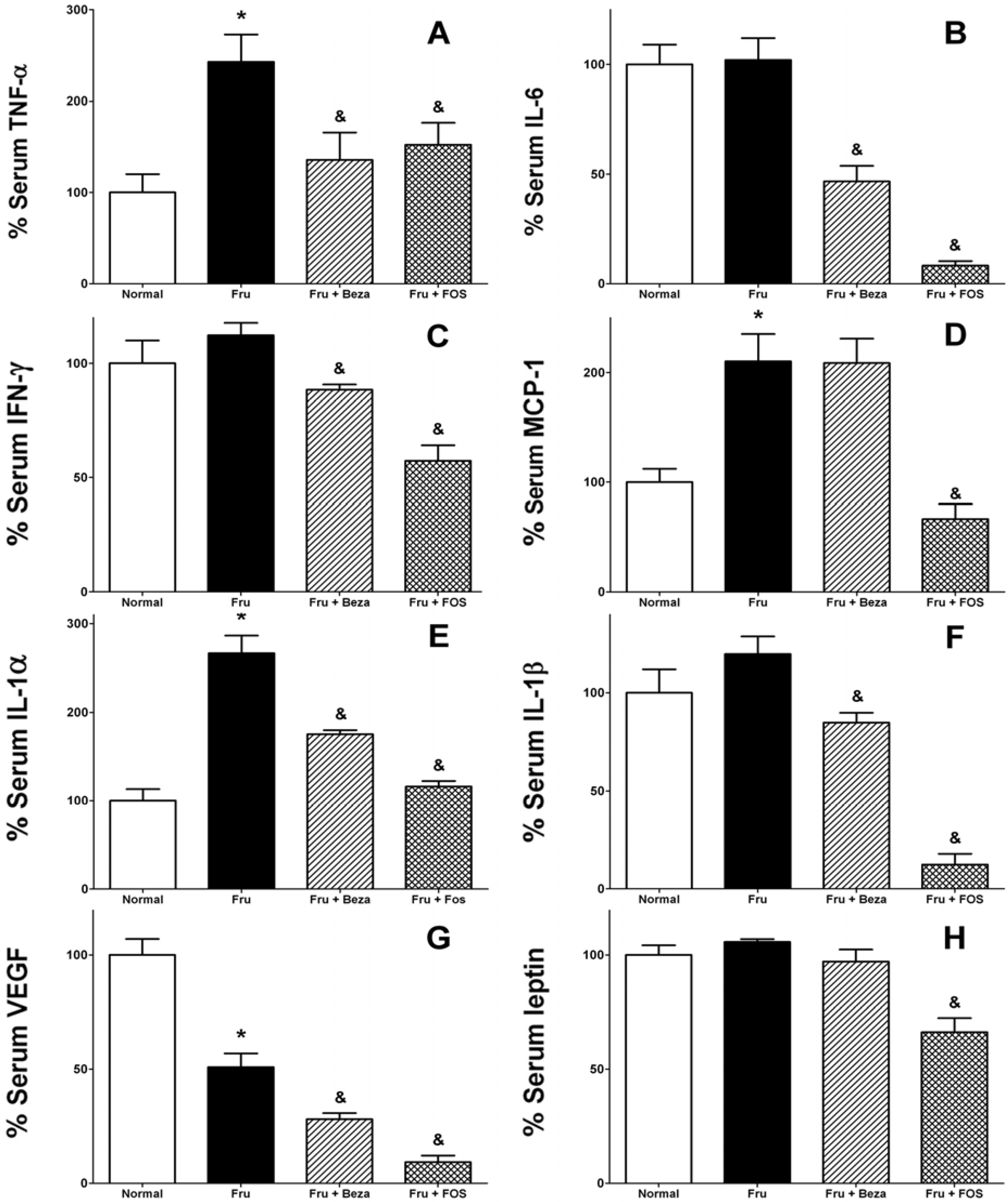
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Mendoza-Cruz, M.; Castro-Lara, D.; Mera-Ovando, L.M.; Chávez-Torrijos, E.K.; Bye-Boettler, R. Plantas Medicinales Utilizadas por los Rarámuri de la Sierra Tarahumra, Chihuahua, Publicación de Difusión Núm. 1; Instituto de Biología, Universidad Nacional Autónoma de México: Distrito Federal, Mexico, 2009; pp. 18–20. [Google Scholar]
- Alarcon-Aguilar, F.J.; Roman-Ramos, R.; Jimenez-Estrada, M.; Reyes-Chilpa, R.; Gonzalez-Paredes, B.; Flores-Saenz, J.L. Effect of three Mexican medicinal plants (Asteraceae) on blood glucose levels in healthy mice and rabbits. J. Ethnopharmacol. 1997, 55, 171–177. [Google Scholar] [CrossRef]
- Alarcon-Aguilar, F.J.; Jimenez-Estrada, M.; Reyes-Chilpa, R.; Gonzalez-Paredes, B.; Contreras, C.C.; Roman-Ramos, R. Hypoglycemic activity of root water decoction, sesquiterpenoids, and one polysaccharide fraction from Psacalium decompositum in mice. J. Ethnopharmacol. 2000, 69, 207–215. [Google Scholar] [CrossRef]
- Jimenez-Estrada, M.; Merino-Aguilar, H.; Lopez-Fernandez, A.; Rojano-Vilchis, N.A.; Roman-Ramos, R.; Alarcon-Aguilar, F.J. Chemical characterization and evaluation of the hypoglycemic effect of fructooligosaccharides from Psacalium decompositum. J. Complement. Integr. Med. 2011, 8, 1413–1423. [Google Scholar]
- De Roover, J.; Vandenbranden, K.; van Laere, A.; van den Ende, W. Drought induces fructan synthesis and 1-SST (sucrose: sucrose fructosyltransferase) in roots and leaves of chicory seedlings (Chicorium intybus L.). Planta 2000, 210, 808–814. [Google Scholar] [CrossRef]
- Vereyken, I.J.; Chupin, V.; Demel, R.A.; Smeekens, S.C.M.; de Kruijff, B. Fructans insert between the headgroups of phospholipids. Biochim. Biophys. Acta 2001, 1510, 307–320. [Google Scholar] [CrossRef]
- L’homme, C.L.; Puigserver, A.; Biagini, A. Effect of food processing on the degradation of fructooligosaccharides in fruit. Food Chem. 2003, 82, 533–537. [Google Scholar] [CrossRef]
- Yasuda, A.; Inoue, K.; Sanbongi, C.; Yanagisawa, R.; Ichinose, T.; Tanaka, M.; Yoshikawa, T.; Takano, H. Dietary supplementation with fructooligosaccharides attenuates allergic peritonitis in mice. Biochem. Biophys. Res. Commun. 2012, 422, 546–550. [Google Scholar] [CrossRef]
- Kaume, L.; Gilbert, W.; Gadang, V.; Devareddy, L. Dietary supplementation of fructooligosaccharides reduces hepatic steatosis associated with insulin resistance in obese Zucker rats. J. Funct. Foods Health Dis. 2011, 1, 199–213. [Google Scholar]
- Nakamura, Y.; Natsume, M.; Yasuda, A.; Ishizaka, M.; Kawahata, K.; Koga, J. Fructooligosaccharides suppress high-fat diet-induced fat accumulation in C57BL/6J mice. BioFactors 2011. [Google Scholar] [CrossRef]
- Malaguarnera, M.; Vacante, M.; Antic, T.; Giordano, M.; Chisari, G.; Acquaviva, R.; Mastrojeni, S.; Malaguarnera, G.; Mistretta, A.; Li Volti, G.; et al. Bifidobacterium longum with fructo-oligosaccharides in patients with non alcoholic steatohepatitis. Dig. Dis. Sci. 2012, 57, 545–553. [Google Scholar] [CrossRef]
- Galisteo, M.; Duarte, J.; Zarzuelo, A. Effects of dietary fibers on disturbances clustered in the metabolic syndrome. J. Nutr. Biochem. 2008, 19, 71–84. [Google Scholar] [CrossRef]
- Rutledge, A.C.; Adeli, K. Fructose and the metabolic syndrome: Pathophysiology and molecular mechanisms. Nutr. Rev. 2007, 65, S13–S23. [Google Scholar] [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Esposito, K.; Nappo, F.; Marfella, R.; Giugliano, G.; Giugliano, F.; Ciotola, M.; Quagliaro, L.; Ceriello, A.; Giugliano, D. Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: Role of oxidative stress. Circulation 2002, 106, 2067–2072. [Google Scholar] [CrossRef]
- Esposito, K.; Pontillo, A.; Giugliano, F.; Giugliano, G.; Marfella, R.; Nicoletti, G.; Giugliano, D. Association of low interleukin-10 levels with the metabolic syndrome in obese women. J. Clin. Endocrinol. Meta. 2003, 88, 1055–1058. [Google Scholar] [CrossRef]
- Wexler, D.J.; Hu, F.B.; Manson, J.E.; Rifai, N.; Meigs, J.B. Mediating effects of inflammatory biomarkers on insulin resistance associated with obesity. Obes. Res. 2005, 13, 1772–1783. [Google Scholar] [CrossRef]
- Blancas-Flores, G.; Almanza-Pérez, J.C.; López-Roa, R.I.; Alarcón-Aguilar, F.J.; García-Macedo, R.; Cruz, M. La obesidad como un proceso inflamatorio. Bol. Med. Hosp. Infant. Mex. 2010, 67, 88–97. [Google Scholar]
- Alarcon-Aguilar, F.J.; Jimenez-Estrada, M.; Reyes-Chilpa, R.; Roman-Ramos, R. Hypoglycemic effect of extracts and fractions from Psacalium decompositum in healthy and alloxan-diabetic mice. J. Ethnopharmacol. 2000, 72, 21–27. [Google Scholar] [CrossRef]
- Tai, M.M. A mathematical model for the determination of total area under glucose tolerance and other metabolic curves. Diabetes Care 1994, 17, 152–154. [Google Scholar]
- Figlewicz, D.P.; Ioannou, G.; Bennett-Jay, J.; Kittleson, S.; Savard, C.; Roth, C.L. Effect of moderate intake of sweeteners on metabolic health in the rat. Physiol. Behav. 2009, 98, 618–624. [Google Scholar] [CrossRef]
- Elliott, S.S.; Keim, N.L.; Stern, J.S.; Teff, K.; Havel, P.J. Fructose, weight gain, and the insulin resistance syndrome. Am. J. Clin. Nutr. 2002, 76, 911–922. [Google Scholar]
- Strader, A.D.; Woods, S.C. Gastrointestinal hormones and food intake. Gastroenterology 2005, 128, 175–191. [Google Scholar] [CrossRef]
- De Moura, R.F.; Ribeiro, C.; de Oliveira, J.A.; Stevanato, E.; de Mello, M.A. Metabolic syndrome signs in Wistar rats submitted to different high-fructose ingestion protocols. Br. J. Nutr. 2009, 101, 1178–1184. [Google Scholar] [CrossRef]
- Lindqvist, A.; Baelemans, A.; Erlanson-Albertsson, C. Effects of sucrose, glucose and fructose on peripheral and central appetite signals. Regul. Pept. 2008, 150, 26–32. [Google Scholar] [CrossRef]
- Abdullah, M.M.; Riediger, N.N.; Chen, Q.; Zhao, Z.; Azordegan, N.; Xu, Z.; Fischer, G.; Othman, R.A.; Pierce, G.N.; Tappia, P.S.; et al. Effects of long-term consumption of a high-fructose diet on conventional cardiovascular risk factors in Sprague-Dawley rats. Mol. Cell Biochem. 2009, 327, 247–256. [Google Scholar] [CrossRef]
- Bocarsly, M.E.; Powell, E.S.; Avena, N.M.; Hoebel, B.G. High-fructose corn syrup causes characteristics of obesity in rats: Increased body weight, body fat and triglyceride levels. Pharmacol. Biochem. Behav. 2010, 97, 101–106. [Google Scholar] [CrossRef]
- Alzamendi, A.; Giovambattista, A.; Raschia, A.; Madrid, V.; Gaillard, R.C.; Rebolledo, O.; Gagliardino, J.J.; Spinedi, E. Fructose-rich diet-induced abdominal adipose tissue endocrine dysfunction in normal male rats. Endocrine 2009, 35, 227–232. [Google Scholar] [CrossRef]
- Kusminski, C.M.; da Silva, N.F.; Creely, S.J.; Fisher, F.M.; Harte, A.L.; Baker, A.R.; Kumar, S.; McTernan, P.G. The in vitro effects of resistin on the innate immune signaling pathway in isolated human subcutaneous adipocytes. J. Clin. Endocrinol. MeTab. 2007, 92, 270–276. [Google Scholar] [CrossRef]
- Astolphi, R.D.; Curbete, M.M.; Colombo, N.H.; Shirakashi, D.J.; Chiba, F.Y.; Prieto, A.K.; Cintra, L.T.; Bomfim, S.R.; Ervolino, E.; Sumida, D.H. Periapical lesions decrease insulin signal and cause insulin resistance. J. Endod. 2013, 39, 648–652. [Google Scholar] [CrossRef]
- Veličković, N.; Djordjevic, A.; Vasiljević, A.; Bursać, B.; Milutinović, D.V.; Matić, G. Tissue-specific regulation of inflammation by macrophage migration inhibitory factor and glucocorticoids in fructose-fed Wistar rats. Br. J. Nutr. 2013, 110, 456–465. [Google Scholar] [CrossRef]
- Senn, J.J.; Klover, P.J.; Nowak, I.A.; Zimmers, T.A.; Koniaris, L.G.; Furlanetto, R.W.; Mooney, R.A. Suppressor of cytokine signaling-3 (SOCS-3), a potential mediator of interleukin-6-dependent insulin resistance in hepatocytes. J. Biol. Chem. 2003, 278, 13740–13746. [Google Scholar]
- Tataranni, P.A.; Ortega, E. A burning question: Does an adipokine-induced activation of the immune system mediate the effect of overnutrition on type 2 diabetes? Diabetes 2005, 54, 917–927. [Google Scholar] [CrossRef]
- Kim, J.H.; Bachmann, R.A.; Chen, J. Interleukin-6 and insulin resistance. Vitam. Horm. 2009, 80, 613–633. [Google Scholar] [CrossRef]
- Pahan, K. Lipid-lowering drugs. Cell. Mol. Life Sci. 2006, 63, 1165–1178. [Google Scholar] [CrossRef]
- Ranjbaran, H.; Sokol, S.I.; Gallo, A.; Eid, R.E.; Iakimov, A.O.; D’Alessio, A.; Kapoor, J.R.; Akhtar, S.; Howes, C.J.; Aslan, M.; et al. An inflammatory pathway of IFN-gamma production in coronary atherosclerosis. J. Immunol. 2007, 178, 592–604. [Google Scholar]
- Guerre-Millo, M. Adipose tissue and adipokines: For better or worse. Diabetes Metab. 2004, 30, 13–19. [Google Scholar] [CrossRef]
- Hernández, C.; González, V.; Fuentes, A.; Cesar, R.; González, B.; García, E. Citocinas proinflamatorias en la infección de tejidos blandos de pacientes diabéticos. Rev. Med. Inst. Mex. Seguro Soc. 2004, 42, 227–233. [Google Scholar]
- Esposito, K.; Giugliano, D. Diet and inflammation: A link to metabolic and cardiovascular diseases. Eur. Heart J. 2006, 27, 15–20. [Google Scholar] [CrossRef]
- Maachi, M.; Pieroni, L.; Bruckert, E.; Jardel, C.; Fellahi, S.; Hainque, B.; Capeau, J.; Bastard, J.P. Systemic low-grade inflammation is related to both circulating and adipose tissue TNFα, leptin and IL-6 levels in obese women. Int. J. Obes. Relat. Metab. Disord. 2004, 28, 993–997. [Google Scholar] [CrossRef]
- Medina, E.A.; Erickson, K.L.; Stanhope, K.L.; Havel, P.J. Evidence that tumor necrosis factor-alpha-induced hyperinsulinemia prevents decreases of circulating leptin during fasting in rats. Metabolism 2002, 51, 1104–1110. [Google Scholar] [CrossRef]
- Bastard, J.P.; Maachi, M.; van Nhieu, J.T.; Jardel, C.; Bruckert, E.; Grimaldi, A.; Robert, J.J.; Capeau, J.; Hainque, B. Adipose tissue IL-6 content correlates with resistance to insulin activation of glucose uptake both in vivo and in vitro. J. Clin. Endocrinol. Metab. 2002, 87, 2084–2089. [Google Scholar] [CrossRef]
- Rabe, K.; Lehrke, M.; Parhofer, K.G.; Broedl, U.C. Adipokines and insulin resistance. Mol. Med. 2008, 14, 741–751. [Google Scholar]
- Halle, M.; Persson, P.B. Role of leptin and leptin receptor in inflammation. Am. J. Physiol. 2003, 284, R760–R762. [Google Scholar]
- Martin, S.S.; Qasim, A.; Reilly, M.P. Leptin resistance: A possible interface of inflammation and metabolism in obesity-related cardiovascular disease. J. Am. Coll. Cardiol. 2008, 52, 1201–1210. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Merino-Aguilar, H.; Arrieta-Baez, D.; Jiménez-Estrada, M.; Magos-Guerrero, G.; Hernández-Bautista, R.J.; Susunaga-Notario, A.D.C.; Almanza-Pérez, J.C.; Blancas-Flores, G.; Román-Ramos, R.; Alarcón-Aguilar, F.J. Effect of Fructooligosaccharides Fraction from Psacalium decompositum on Inflammation and Dyslipidemia in Rats with Fructose-Induced Obesity. Nutrients 2014, 6, 591-604. https://doi.org/10.3390/nu6020591
Merino-Aguilar H, Arrieta-Baez D, Jiménez-Estrada M, Magos-Guerrero G, Hernández-Bautista RJ, Susunaga-Notario ADC, Almanza-Pérez JC, Blancas-Flores G, Román-Ramos R, Alarcón-Aguilar FJ. Effect of Fructooligosaccharides Fraction from Psacalium decompositum on Inflammation and Dyslipidemia in Rats with Fructose-Induced Obesity. Nutrients. 2014; 6(2):591-604. https://doi.org/10.3390/nu6020591
Chicago/Turabian StyleMerino-Aguilar, Héctor, Daniel Arrieta-Baez, Manuel Jiménez-Estrada, Gil Magos-Guerrero, René Javier Hernández-Bautista, Ana Del Carmen Susunaga-Notario, Julio Cesar Almanza-Pérez, Gerardo Blancas-Flores, Rubén Román-Ramos, and Francisco Javier Alarcón-Aguilar. 2014. "Effect of Fructooligosaccharides Fraction from Psacalium decompositum on Inflammation and Dyslipidemia in Rats with Fructose-Induced Obesity" Nutrients 6, no. 2: 591-604. https://doi.org/10.3390/nu6020591
APA StyleMerino-Aguilar, H., Arrieta-Baez, D., Jiménez-Estrada, M., Magos-Guerrero, G., Hernández-Bautista, R. J., Susunaga-Notario, A. D. C., Almanza-Pérez, J. C., Blancas-Flores, G., Román-Ramos, R., & Alarcón-Aguilar, F. J. (2014). Effect of Fructooligosaccharides Fraction from Psacalium decompositum on Inflammation and Dyslipidemia in Rats with Fructose-Induced Obesity. Nutrients, 6(2), 591-604. https://doi.org/10.3390/nu6020591