Selenistasis: Epistatic Effects of Selenium on Cardiovascular Phenotype


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Clinical Evidence Linking Plasma Selenium to Cardiovascular Disease
2.1. Selenium Level and Risk of Cardiovascular Disease
2.2. Selenium Level and Cardiometabolic Health
2.3. Inconclusive Benefits of Selenium Supplementation
2.4. Is Selenium an Influence on Cardiovascular Health?
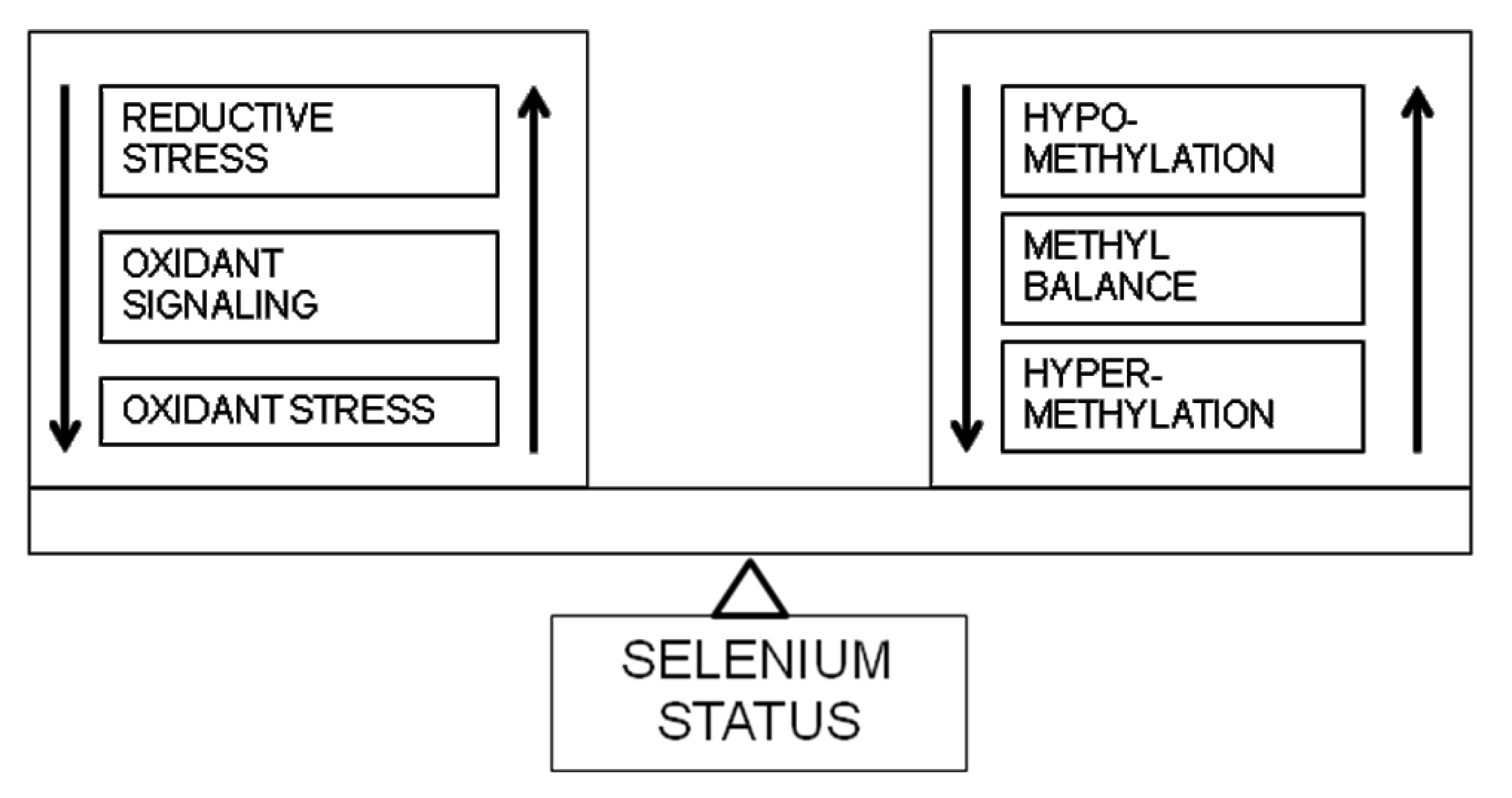
3. Biphasic Effects of Selenium Status
3.1. Major Biologic Effects of Selenium
3.1.1. Anti-Oxidant Effects
3.1.2. Cell Proliferation and Survival
3.1.3. Immune and Anti-Infective Mechanisms
3.1.4. Effects on Matrix Metabolism
3.1.5. Epigenetic Mechanisms
3.2. Both Selenium Deficiency and Selenium Supplementation Induce Similar Phenotypes
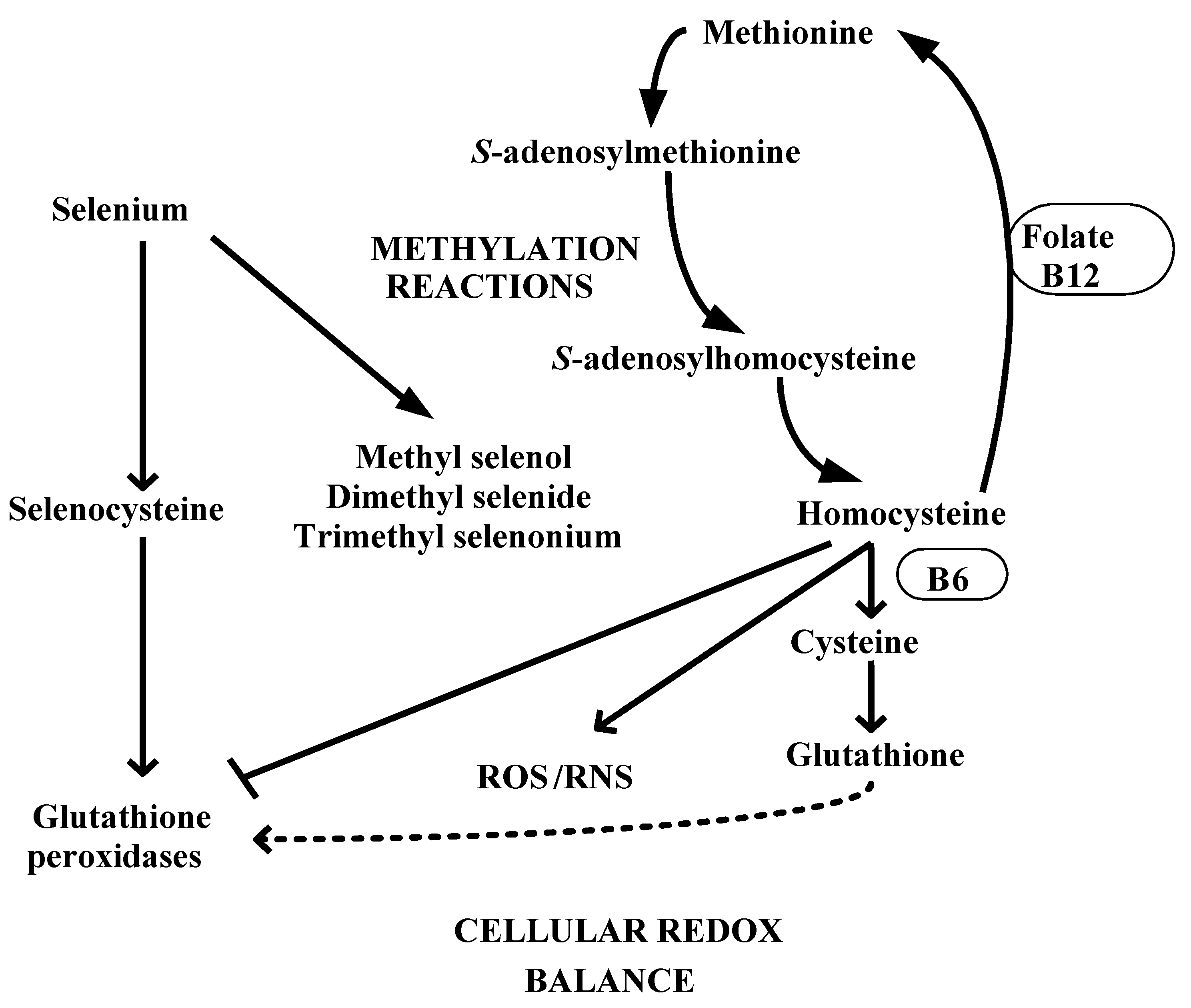
4. Selenium and Redox-Methylation Balance
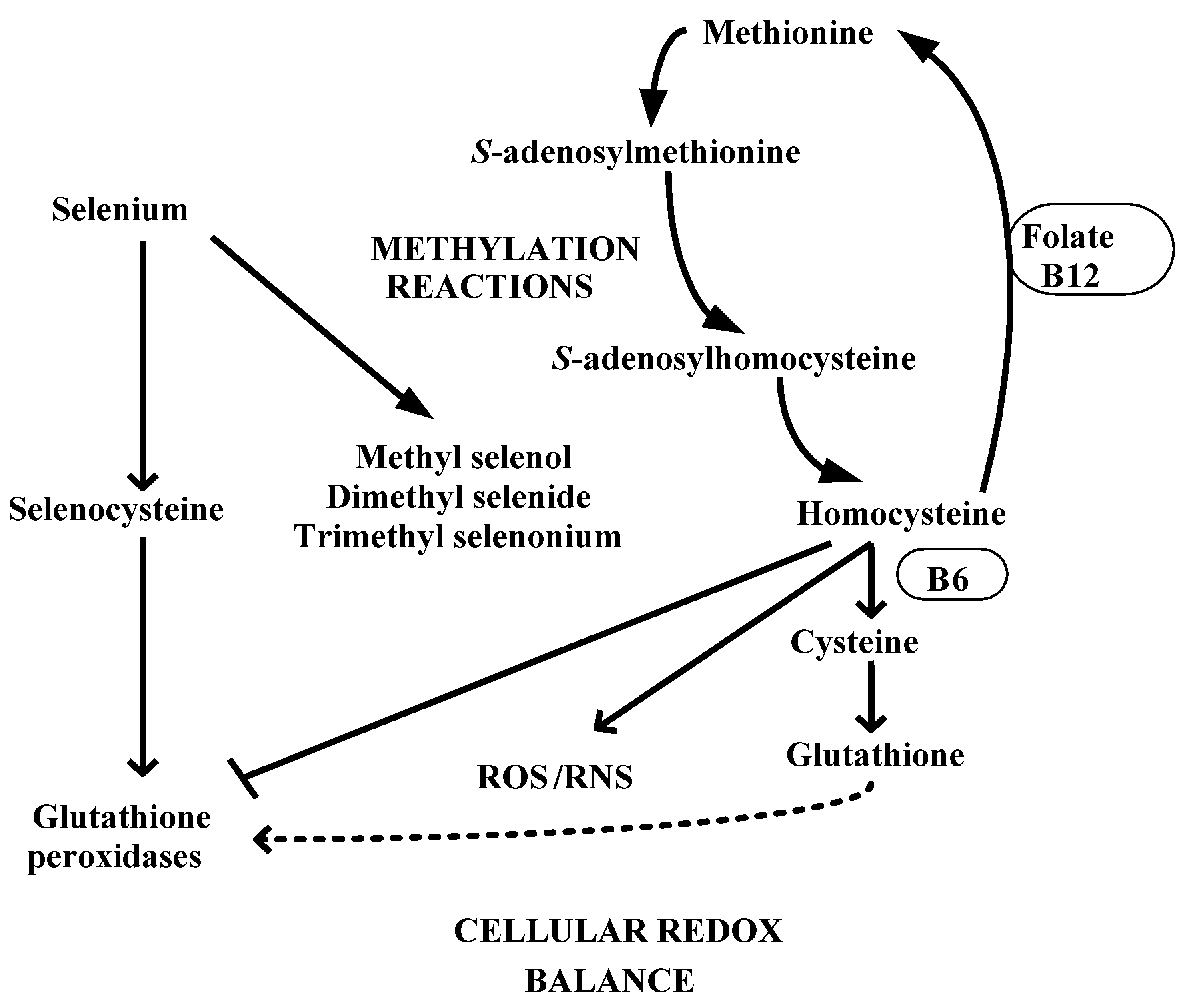
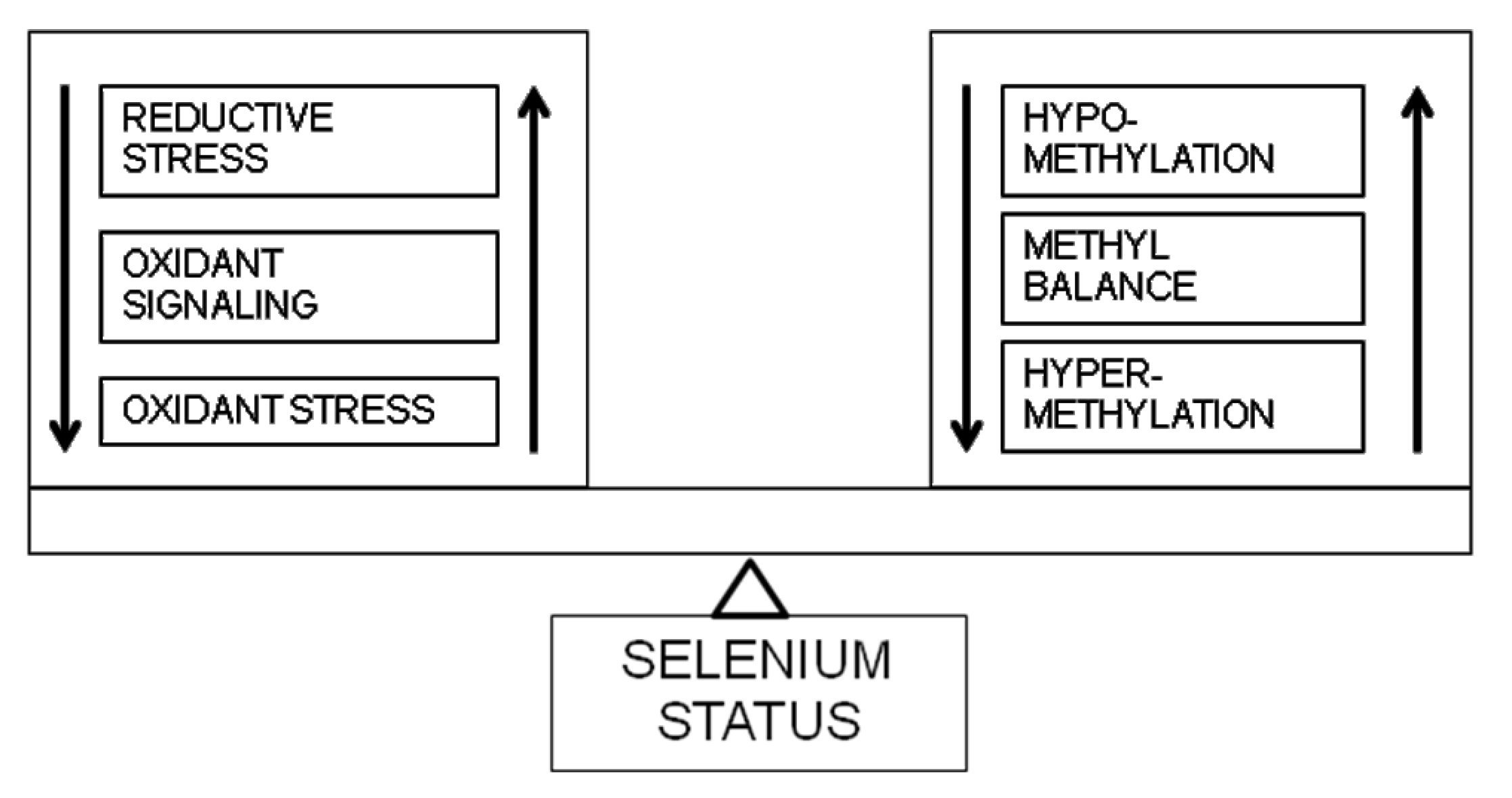
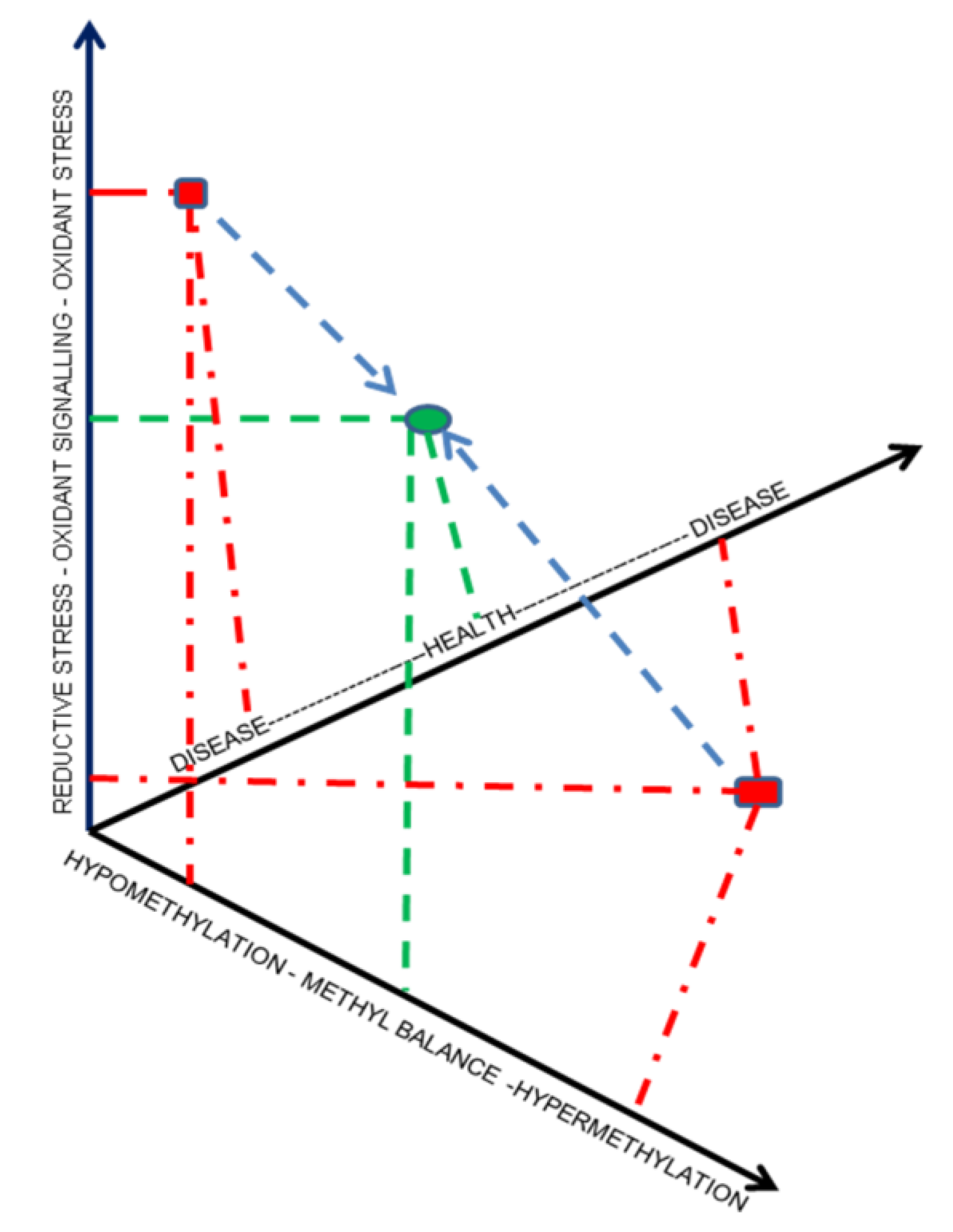
5. Selenistasis—A Framework for Selenium in Health and Disease
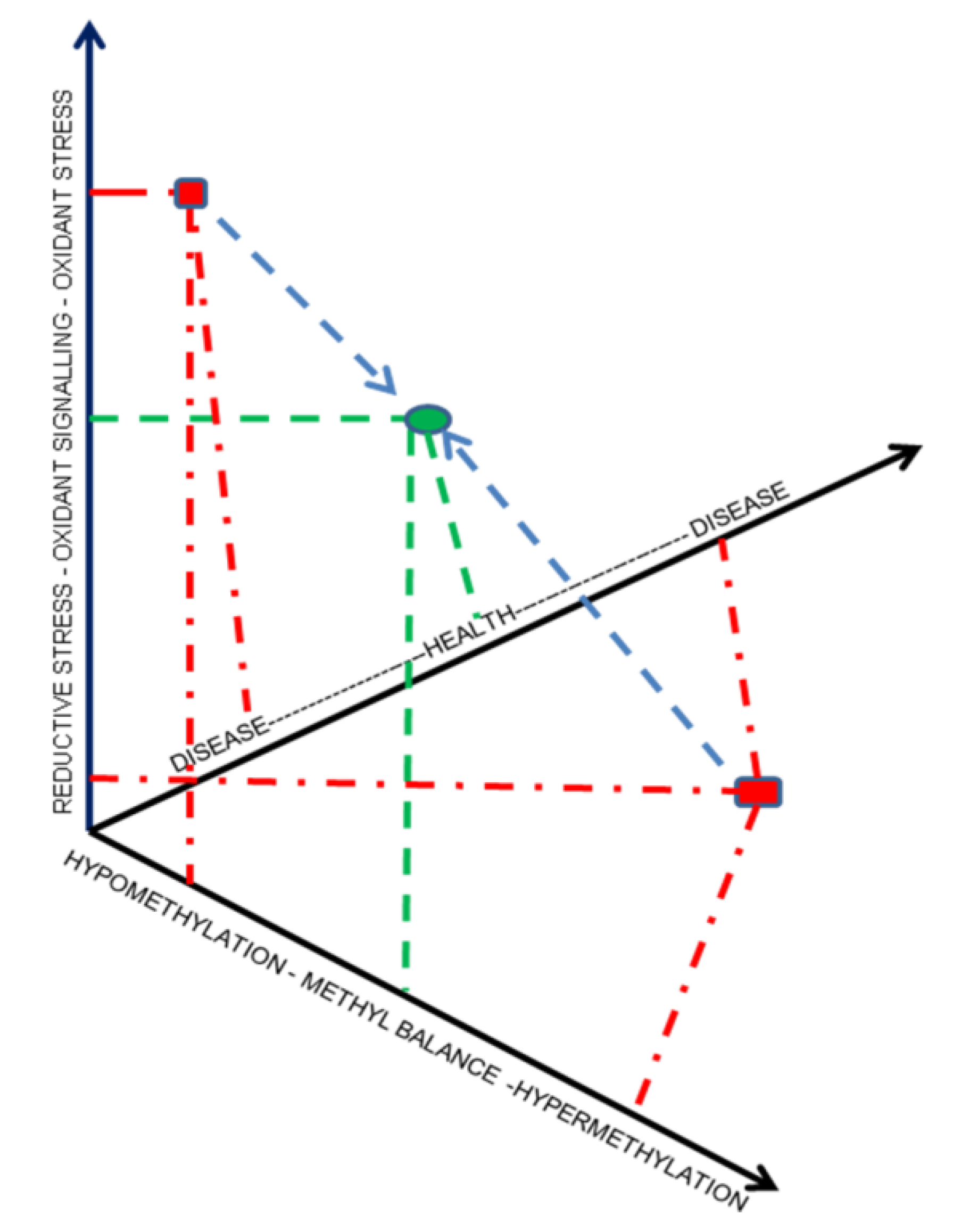
6. Systems Biology—A Novel and Necessary Approach to Evaluating Selenistasis
7. Conclusions
Acknowledgments
Conflict of Interest
References
- Stadtman, T.C. Selenoproteins—Tracing the role of a trace element in protein function. PLoS Biol. 2005, 3, e421. [Google Scholar]
- Salonen, J.T.; Alfthan, G.; Huttunen, J.K.; Pikkarainen, J.; Puska, P. Association between cardiovascular death and myocardial infarction and serum selenium in a matched-pair longitudinal study. Lancet 1982, 2, 175–179. [Google Scholar]
- Flores-Mateo, G.; Navas-Acien, A.; Pastor-Barriuso, R.; Guallar, E. Selenium and coronary heart disease: A meta-analysis. Am. J. Clin. Nutr. 2006, 84, 762–773. [Google Scholar]
- Salvini, S.; Hennekens, C.H.; Morris, J.S.; Willett, W.C.; Stampfer, M.J. Plasma levels of the antioxidant selenium and risk of myocardial infarction among U.S. physicians. Am. J. Cardiol. 1995, 76, 1218–1221. [Google Scholar] [CrossRef]
- Bleys, J.; Navas-Acien, A.; Guallar, E. Serum selenium levels and all-cause, cancer, and cardiovascular mortality among us adults. Arch. Intern. Med. 2008, 168, 404–410. [Google Scholar] [CrossRef]
- Ge, K.; Yang, G. The epidemiology of selenium deficiency in the etiological study of endemic diseases in china. Am. J. Clin. Nutr. 1993, 57, 259S–263S. [Google Scholar]
- Beck, M.A.; Kolbeck, P.C.; Shi, Q.; Rohr, L.H.; Morris, V.C.; Levander, O.A. Increased virulence of a human enterovirus (coxsackievirus B3) in selenium-deficient mice. J. Infect. Dis. 1994, 170, 351–357. [Google Scholar] [CrossRef]
- Beck, M.A.; Kolbeck, P.C.; Rohr, L.H.; Shi, Q.; Morris, V.C.; Levander, O.A. Benign human enterovirus becomes virulent in selenium-deficient mice. J. Med. Virol. 1994, 43, 166–170. [Google Scholar] [CrossRef]
- Beck, M.A.; Shi, Q.; Morris, V.C.; Levander, O.A. Rapid genomic evolution of a non-virulent coxsackievirus B3 in selenium-deficient mice results in selection of identical virulent isolates. Nat. Med. 1995, 1, 433–436. [Google Scholar]
- Beck, M.A. Increased virulence of coxsackievirus B3 in mice due to vitamin E or selenium deficiency. J. Nutr. 1997, 127, 966S–970S. [Google Scholar]
- Lei, C.; Niu, X.; Ma, X.; Wei, J. Is selenium deficiency really the cause of keshan disease? Environ. Geochem. Health 2011, 33, 183–188. [Google Scholar] [CrossRef]
- Lee, S.R.; Bar-Noy, S.; Kwon, J.; Levine, R.L.; Stadtman, T.C.; Rhee, S.G. Mammalian thioredoxin reductase: Oxidation of the C-terminal cysteine/selenocysteine active site forms a thioselenide, and replacement of selenium with sulfur markedly reduces catalytic activity. Proc. Natl. Acad. Sci. USA 2000, 97, 2521–2526. [Google Scholar] [CrossRef]
- Dworkin, B.M.; Antonecchia, P.P.; Smith, F.; Weiss, L.; Davidian, M.; Rubin, D.; Rosenthal, W.S. Reduced cardiac selenium content in the acquired immunodeficiency syndrome. J. Parenter. Enter. Nutr. 1989, 13, 644–647. [Google Scholar]
- Oster, O.; Prellwitz, W.; Kasper, W.; Meinertz, T. Congestive cardiomyopathy and the selenium content of serum. Clin. Chim. Acta 1983, 128, 125–132. [Google Scholar] [CrossRef]
- De Lorgeril, M.; Salen, P.; Accominotti, M.; Cadau, M.; Steghens, J.P.; Boucher, F.; de Leiris, J. Dietary and blood antioxidants in patients with chronic heart failure: Insights into the potential importance of selenium in heart failure. Eur. J. Heart Fail. 2001, 3, 661–669. [Google Scholar] [CrossRef]
- Arroyo, M.; Laguardia, S.P.; Bhattacharya, S.K.; Nelson, M.D.; Johnson, P.L.; Carbone, L.D.; Newman, K.P.; Weber, K.T. Micronutrients in African-Americans with decompensated and compensated heart failure. Transl. Res. 2006, 148, 301–308. [Google Scholar] [CrossRef]
- Inoko, M.; Konishi, T.; Matsusue, S.; Kobashi, Y. Midmural fibrosis of left ventricle due to selenium deficiency. Circulation 1998, 98, 2638–2639. [Google Scholar]
- Lockitch, G.; Taylor, G.P.; Wong, L.T.; Davidson, A.G.; Dison, P.J.; Riddell, D.; Massing, B. Cardiomyopathy associated with nonendemic selenium deficiency in a caucasian adolescent. Am. J. Clin. Nutr. 1990, 52, 572–577. [Google Scholar]
- Reeves, W.C.; Marcuard, S.P.; Willis, S.E.; Movahed, A. Reversible cardiomyopathy due to selenium deficiency. J. Parenter. Enter. Nutr. 1989, 13, 663–665. [Google Scholar]
- Laclaustra, M.; Navas-Acien, A.; Stranges, S.; Ordovas, J.M.; Guallar, E. Serum selenium concentrations and diabetes in U.S. Adults: National Health and Nutrition Examination Survey (NHANES) 2003–2004. Environ. Health Perspect. 2009, 117, 1409–1413. [Google Scholar]
- Laclaustra, M.; Stranges, S.; Navas-Acien, A.; Ordovas, J.M.; Guallar, E. Serum selenium and serum lipids in US adults: National Health and Nutrition Examination Survey (NHANES) 2003–2004. Atherosclerosis 2010, 210, 643–648. [Google Scholar] [CrossRef]
- Laclaustra, M.; Navas-Acien, A.; Stranges, S.; Ordovas, J.M.; Guallar, E. Serum selenium concentrations and hypertension in the US population. Circ. Cardiovasc. Qual. Outcomes 2009, 2, 369–376. [Google Scholar] [CrossRef]
- Stranges, S.; Laclaustra, M.; Ji, C.; Cappuccio, F.P.; Navas-Acien, A.; Ordovas, J.M.; Rayman, M.; Guallar, E. Higher selenium status is associated with adverse blood lipid profile in British adults. J. Nutr. 2010, 140, 81–87. [Google Scholar] [CrossRef]
- Rayman, M.P.; Stranges, S.; Griffin, B.A.; Pastor-Barriuso, R.; Guallar, E. Effect of supplementation with high-selenium yeast on plasma lipids: A randomized trial. Ann. Intern. Med. 2011, 154, 656–665. [Google Scholar]
- Nawrot, T.S.; Staessen, J.A.; Roels, H.A.; Den Hond, E.; Thijs, L.; Fagard, R.H.; Dominiczak, A.F.; Struijker-Boudier, H.A. Blood pressure and blood selenium: A cross-sectional and longitudinal population study. Eur. Heart J. 2007, 28, 628–633. [Google Scholar]
- Akbaraly, T.N.; Arnaud, J.; Rayman, M.P.; Hininger-Favier, I.; Roussel, A.M.; Berr, C.; Fontbonne, A. Plasma selenium and risk of dysglycemia in an elderly french population: Results from the prospective epidemiology of vascular ageing study. Nutr. Metab. 2010, 7, 21. [Google Scholar] [CrossRef]
- Stranges, S.; Sieri, S.; Vinceti, M.; Grioni, S.; Guallar, E.; Laclaustra, M.; Muti, P.; Berrino, F.; Krogh, V. A prospective study of dietary selenium intake and risk of type 2 diabetes. BMC Public Health 2010, 10, 564. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Frustaci, A.; Sabbioni, E.; Fortaner, S.; Farina, M.; del Torchio, R.; Tafani, M.; Morgante, E.; Ciriolo, M.R.; Russo, M.A.; Chimenti, C. Selenium- and zinc-deficient cardiomyopathy in human intestinal malabsorption: Preliminary results of selenium/zinc infusion. Eur. J. Heart Fail. 2012, 14, 202–210. [Google Scholar] [CrossRef]
- Witte, K.K.; Nikitin, N.P.; Parker, A.C.; von Haehling, S.; Volk, H.D.; Anker, S.D.; Clark, A.L.; Cleland, J.G. The effect of micronutrient supplementation on quality-of-life and left ventricular function in elderly patients with chronic heart failure. Eur. Heart J. 2005, 26, 2238–2244. [Google Scholar]
- Lippman, S.M.; Klein, E.A.; Goodman, P.J.; Lucia, M.S.; Thompson, I.M.; Ford, L.G.; Parnes, H.L.; Minasian, L.M.; Gaziano, J.M.; Hartline, J.A.; et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: The selenium and vitamin E cancer prevention trial (SELECT). JAMA 2009, 301, 39–51. [Google Scholar]
- Stranges, S.; Marshall, J.R.; Natarajan, R.; Donahue, R.P.; Trevisan, M.; Combs, G.F.; Cappuccio, F.P.; Ceriello, A.; Reid, M.E. Effects of long-term selenium supplementation on the incidence of type 2 diabetes: A randomized trial. Ann. Intern. Med. 2007, 147, 217–223. [Google Scholar]
- Combs, G.F., Jr.; Watts, J.C.; Jackson, M.I.; Johnson, L.K.; Zeng, H.; Scheett, A.J.; Uthus, E.O.; Schomburg, L.; Hoeg, A.; Hoefig, C.S.; et al. Determinants of selenium status in healthy adults. Nutr. J. 2011, 10, 75. [Google Scholar] [CrossRef]
- Uthus, E.O.; Yokoi, K.; Davis, C.D. Selenium deficiency in Fisher-344 rats decreases plasma and tissue homocysteine concentrations and alters plasma homocysteine and cysteine redox status. J. Nutr. 2002, 132, 1122–1128. [Google Scholar]
- Uthus, E.O.; Ross, S.A.; Davis, C.D. Differential effects of dietary selenium (Se) and folate on methyl metabolism in liver and colon of rats. Biol. Trace Elem. Res. 2006, 109, 201–214. [Google Scholar]
- Davis, C.D.; Uthus, E.O. Dietary folate and selenium affect dimethylhydrazine-induced aberrant crypt formation, global DNA methylation and one-carbon metabolism in rats. J. Nutr. 2003, 133, 2907–2914. [Google Scholar]
- Metes-Kosik, N.; Luptak, I.; Dibello, P.M.; Handy, D.E.; Tang, S.S.; Zhi, H.; Qin, F.; Jacobsen, D.W.; Loscalzo, J.; Joseph, J. Both selenium deficiency and modest selenium supplementation lead to myocardial fibrosis in mice via effects on redox-methylation balance. Mol. Nutr. Food Res. 2012, 56, 1812–1824. [Google Scholar]
- Salbe, A.D.; Levander, O.A. Comparative toxicity and tissue retention of selenium in methionine-deficient rats fed sodium selenate or L-selenomethionine. J. Nutr. 1990, 120, 207–212. [Google Scholar]
- Xiang, N.; Zhao, R.; Zhong, W. Sodium selenite induces apoptosis by generation of superoxide via the mitochondrial-dependent pathway in human prostate cancer cells. Cancer Chemother. Pharmacol. 2009, 63, 351–362. [Google Scholar]
- Misra, S.; Niyogi, S. Selenite causes cytotoxicity in rainbow trout (Oncorhynchus mykiss) hepatocytes by inducing oxidative stress. Toxicol. In Vitro 2009, 23, 1249–1258. [Google Scholar] [CrossRef]
- Alanne, M.; Kristiansson, K.; Auro, K.; Silander, K.; Kuulasmaa, K.; Peltonen, L.; Salomaa, V.; Perola, M. Variation in the selenoprotein s gene locus is associated with coronary heart disease and ischemic stroke in two independent finnish cohorts. Hum. Genet. 2007, 122, 355–365. [Google Scholar] [CrossRef]
- Bailey, R.L.; Gahche, J.J.; Lentino, C.V.; Dwyer, J.T.; Engel, J.S.; Thomas, P.R.; Betz, J.M.; Sempos, C.T.; Picciano, M.F. Dietary supplement use in the united states, 2003–2006. J. Nutr. 2011, 141, 261–266. [Google Scholar] [CrossRef]
- Bermano, G.; Nicol, F.; Dyer, J.A.; Sunde, R.A.; Beckett, G.J.; Arthur, J.R.; Hesketh, J.E. Tissue-specific regulation of selenoenzyme gene expression during selenium deficiency in rats. Biochem. J. 1995, 311, 425–430. [Google Scholar]
- Labunskyy, V.M.; Lee, B.C.; Handy, D.E.; Loscalzo, J.; Hatfield, D.L.; Gladyshev, V.N. Both maximal expression of selenoproteins and selenoprotein deficiency can promote development of type 2 diabetes-like phenotype in mice. Antioxid. Redox Signal. 2011, 14, 2327–2336. [Google Scholar] [CrossRef]
- Carlson, B.A.; Moustafa, M.E.; Sengupta, A.; Schweizer, U.; Shrimali, R.; Rao, M.; Zhong, N.; Wang, S.; Feigenbaum, L.; Lee, B.J.; et al. Selective restoration of the selenoprotein population in a mouse hepatocyte selenoproteinless background with different mutant selenocysteine tRNAs lacking UM34. J. Biol. Chem. 2007, 282, 32591–32602. [Google Scholar]
- Moustafa, M.E.; Carlson, B.A.; El-Saadani, M.A.; Kryukov, G.V.; Sun, Q.A.; Harney, J.W.; Hill, K.E.; Combs, G.F.; Feigenbaum, L.; Mansur, D.B.; et al. Selective inhibition of selenocysteine tRNA maturation and selenoprotein synthesis in transgenic mice expressing isopentenyladenosine-deficient selenocysteine tRNA. Mol. Cell. Biol. 2001, 21, 3840–3852. [Google Scholar] [CrossRef]
- Leopold, J.A.; Loscalzo, J. Oxidative risk for atherothrombotic cardiovascular disease. Free Radic. Biol. Med. 2009, 47, 1673–1706. [Google Scholar]
- De Lorgeril, M.; Salen, P. Selenium and antioxidant defenses as major mediators in the development of chronic heart failure. Heart Fail. Rev. 2006, 11, 13–17. [Google Scholar] [CrossRef]
- Toufektsian, M.C.; Boucher, F.; Pucheu, S.; Tanguy, S.; Ribuot, C.; Sanou, D.; Tresallet, N.; de Leiris, J. Effects of selenium deficiency on the response of cardiac tissue to ischemia and reperfusion. Toxicology 2000, 148, 125–132. [Google Scholar] [CrossRef]
- Rakotovao, A.; Tanguy, S.; Toufektsian, M.C.; Berthonneche, C.; Ducros, V.; Tosaki, A.; de Leiris, J.; Boucher, F. Selenium status as determinant of connexin-43 dephosphorylation in ex vivo ischemic/reperfused rat myocardium. J. Trace Elem. Med. Biol. 2005, 19, 43–47. [Google Scholar] [CrossRef]
- Xia, Y.M.; Hill, K.E.; Burk, R.F. Effect of selenium deficiency on hydroperoxide-induced glutathione release from the isolated perfused rat heart. J. Nutr. 1985, 115, 733–742. [Google Scholar]
- Ji, L.L.; Stratman, F.W.; Lardy, H.A. Antioxidant enzyme response to selenium deficiency in rat myocardium. J. Am. Coll. Nutr. 1992, 11, 79–86. [Google Scholar]
- Doroshow, J.H.; Locker, G.Y.; Myers, C.E. Enzymatic defenses of the mouse heart against reactive oxygen metabolites: Alterations produced by doxorubicin. J. Clin. Investig. 1980, 65, 128–135. [Google Scholar] [CrossRef]
- Dimitrov, N.V.; Hay, M.B.; Siew, S.; Hudler, D.A.; Charamella, L.J.; Ullrey, D.E. Abrogation of adriamycin-induced cardiotoxicity by selenium in rabbits. Am. J. Pathol. 1987, 126, 376–383. [Google Scholar]
- Nakano, E.; Takeshige, K.; Toshima, Y.; Tokunaga, K.; Minakami, S. Oxidative damage in selenium deficient hearts on perfusion with adriamycin: Protective role of glutathione peroxidase system. Cardiovasc. Res. 1989, 23, 498–504. [Google Scholar] [CrossRef]
- Boucher, F.; Coudray, C.; Tirard, V.; Barandier, C.; Tresallet, N.; Favier, A.; de Leiris, J. Oral selenium supplementation in rats reduces cardiac toxicity of adriamycin during ischemia and reperfusion. Nutrition 1995, 11, 708–711. [Google Scholar]
- Dursun, N.; Taskin, E.; Yerer Aycan, M.B.; Sahin, L. Selenium-mediated cardioprotection against adriamycin-induced mitochondrial damage. Drug Chem. Toxicol. 2011, 34, 199–207. [Google Scholar]
- Van Vleet, J.F.; Ferrans, V.J.; Weirich, W.E. Cardiac disease induced by chronic adriamycin administration in dogs and an evaluation of vitamin E and selenium as cardioprotectants. Am. J. Pathol. 1980, 99, 13–42. [Google Scholar]
- Hermansen, K.; Wassermann, K. The effect of vitamin E and selenium on doxorubicin (adriamycin) induced delayed toxicity in mice. Acta Pharmacol. Toxicol. 1986, 58, 31–37. [Google Scholar]
- Lymbury, R.S.; Marino, M.J.; Perkins, A.V. Effect of dietary selenium on the progression of heart failure in the ageing spontaneously hypertensive rat. Mol. Nutr. Food Res. 2010, 54, 1436–1444. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Turowski, S.G.; San Martin, I.D.; Rajput, A.; Rustum, Y.M.; Hoffman, R.M.; Seshadri, M. Magnetic resonance and fluorescence-protein imaging of the anti-angiogenic and anti-tumor efficacy of selenium in an orthotopic model of human colon cancer. Anticancer Res. 2011, 31, 387–393. [Google Scholar]
- Liu, J.G.; Zhao, H.J.; Liu, Y.J.; Wang, X.L. Effect of selenium-enriched malt on VEGF and several relevant angiogenic cytokines in diethylnitrosamine-induced hepatocarcinoma rats. J. Trace Elem. Med. Biol. 2010, 24, 52–57. [Google Scholar]
- Wang, Z.; Hu, H.; Li, G.; Lee, H.J.; Jiang, C.; Kim, S.H.; Lu, J. Methylseleninic acid inhibits microvascular endothelial G1 cell cycle progression and decreases tumor microvessel density. Int. J. Cancer 2008, 122, 15–24. [Google Scholar]
- Raines, A.M.; Sunde, R.A. Selenium toxicity but not deficient or super-nutritional selenium status vastly alters the transcriptome in rodents. BMC Genomics 2011, 12, 26. [Google Scholar]
- Bajpai, S.; Mishra, M.; Kumar, H.; Tripathi, K.; Singh, S.K.; Pandey, H.P.; Singh, R.K. Effect of selenium on connexin expression, angiogenesis, and antioxidant status in diabetic wound healing. Biol. Trace Elem. Res. 2011, 144, 327–338. [Google Scholar]
- Wang, Z.; Jiang, C.; Ganther, H.; Lu, J. Antimitogenic and proapoptotic activities of methylseleninic acid in vascular endothelial cells and associated effects on PI3K-AKT, ERK, JNK and p38 MAPK signaling. Cancer Res. 2001, 61, 7171–7178. [Google Scholar]
- Tang, R.; Liu, H.; Wang, T.; Huang, K. Mechanisms of selenium inhibition of cell apoptosis induced by oxysterols in rat vascular smooth muscle cells. Arch. Biochem. Biophys. 2005, 441, 16–24. [Google Scholar]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar]
- Rivera, M.T.; de Souza, A.P.; Moreno, A.H.; Xavier, S.S.; Gomes, J.A.; Rocha, M.O.; Correa-Oliveira, R.; Neve, J.; Vanderpas, J.; Araujo-Jorge, T.C. Progressive Chagas cardiomyopathy is associated with low selenium levels. Am. J. Trop. Med. Hyg. 2002, 66, 706–712. [Google Scholar]
- Davis, C.D.; Brooks, L.; Calisi, C.; Bennett, B.J.; McElroy, D.M. Beneficial effect of selenium supplementation during murine infection with trypanosoma cruzi. J. Parasitol. 1998, 84, 1274–1277. [Google Scholar]
- De Souza, A.P.; de Oliveira, G.M.; Vanderpas, J.; de Castro, S.L.; Rivera, M.T.; Araujo-Jorge, T.C. Selenium supplementation at low doses contributes to the decrease in heart damage in experimental trypanosoma cruzi infection. Parasitol. Res. 2003, 91, 51–54. [Google Scholar]
- Stoedter, M.; Renko, K.; Hog, A.; Schomburg, L. Selenium controls the sex-specific immune response and selenoprotein expression during the acute-phase response in mice. Biochem. J. 2010, 429, 43–51. [Google Scholar]
- Kucharz, E.J.; Olczyk, K. Influence of chronic intoxication with selenium on collagen and elastin content in tissues of rat. Toxicol. Lett. 1993, 68, 295–299. [Google Scholar]
- Ding, M.; Potter, J.J.; Liu, X.; Torbenson, M.S.; Mezey, E. Selenium supplementation decreases hepatic fibrosis in mice after chronic carbon tetrachloride administration. Biol. Trace Elem. Res. 2010, 133, 83–97. [Google Scholar]
- Chen, J.; Chu, Y.; Cao, J.; Wang, W.; Liu, J.; Wang, J. Effects of T-2 toxin and selenium on chondrocyte expression of matrix metalloproteinases (MMP-1, MMP-13), α2-macroglobulin (α2M) and timps. Toxicol. In Vitro 2011, 25, 492–499. [Google Scholar]
- Yoon, S.O.; Kim, M.M.; Chung, A.S. Inhibitory effect of selenite on invasion of HT1080 tumor cells. J. Biol. Chem. 2001, 276, 20085–20092. [Google Scholar] [CrossRef]
- Carlson, B.A.; Yoo, M.H.; Sano, Y.; Sengupta, A.; Kim, J.Y.; Irons, R.; Gladyshev, V.N.; Hatfield, D.L.; Park, J.M. Selenoproteins regulate macrophage invasiveness and extracellular matrix-related gene expression. BMC Immunol. 2009, 10, 57. [Google Scholar]
- Handy, D.E.; Castro, R.; Loscalzo, J. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease. Circulation 2011, 123, 2145–2156. [Google Scholar]
- Cox, R.; Goorha, S. A study of the mechanism of selenite-induced hypomethylated DNA and differentiation of friend erythroleukemic cells. Carcinogenesis 1986, 7, 2015–2018. [Google Scholar]
- Xiang, N.; Zhao, R.; Song, G.; Zhong, W. Selenite reactivates silenced genes by modifying DNA methylation and histones in prostate cancer cells. Carcinogenesis 2008, 29, 2175–2181. [Google Scholar]
- Zeng, H.; Yan, L.; Cheng, W.H.; Uthus, E.O. Dietary selenomethionine increases exon-specific DNA methylation of the p53 gene in rat liver and colon mucosa. J. Nutr. 2011, 141, 1464–1468. [Google Scholar]
- Davis, C.D.; Uthus, E.O.; Finley, J.W. Dietary selenium and arsenic affect DNA methylation in vitro in Caco-2 cells and in vivo in rat liver and colon. J. Nutr. 2000, 130, 2903–2909. [Google Scholar]
- Davis, C.D.; Uthus, E.O. Dietary selenite and azadeoxycytidine treatments affect dimethylhydrazine-induced aberrant crypt formation in rat colon and DNA methylation in HT-29 cells. J. Nutr. 2002, 132, 292–297. [Google Scholar]
- Novoselov, S.V.; Calvisi, D.F.; Labunskyy, V.M.; Factor, V.M.; Carlson, B.A.; Fomenko, D.E.; Moustafa, M.E.; Hatfield, D.L.; Gladyshev, V.N. Selenoprotein deficiency and high levels of selenium compounds can effectively inhibit hepatocarcinogenesis in transgenic mice. Oncogene 2005, 24, 8003–8011. [Google Scholar]
- Joseph, J.; Joseph, L. Hyperhomocysteinemia and cardiovascular disease: New mechanisms beyond atherosclerosis. Metab. Syndr. Relat. Disord. 2003, 1, 97–104. [Google Scholar]
- Hill, K.E.; Burk, R.F. Effect of selenium deficiency and vitamin E deficiency on glutathione metabolism in isolated rat hepatocytes. J. Biol. Chem. 1982, 257, 10668–10672. [Google Scholar]
- Hill, K.E.; Burk, R.F.; Lane, J.M. Effect of selenium depletion and repletion on plasma glutathione and glutathione-dependent enzymes in the rat. J. Nutr. 1987, 117, 99–104. [Google Scholar]
- Hassoun, B.S.; Palmer, I.S.; Dwivedi, C. Selenium detoxification by methylation. Res. Commun. Mol. Pathol. Pharmacol. 1995, 90, 133–142. [Google Scholar]
- Rajasekaran, N.S.; Connell, P.; Christians, E.S.; Yan, L.J.; Taylor, R.P.; Orosz, A.; Zhang, X.Q.; Stevenson, T.J.; Peshock, R.M.; Leopold, J.A.; et al. Human αB-crystallin mutation causes oxido-reductive stress and protein aggregation cardiomyopathy in mice. Cell 2007, 130, 427–439. [Google Scholar] [CrossRef]
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857. [Google Scholar]
- Wang, X.D.; Vatamaniuk, M.Z.; Wang, S.K.; Roneker, C.A.; Simmons, R.A.; Lei, X.G. Molecular mechanisms for hyperinsulinaemia induced by overproduction of selenium-dependent glutathione peroxidase-1 in mice. Diabetologia 2008, 51, 1515–1524. [Google Scholar] [CrossRef]
- Yan, X.; Pepper, M.P.; Vatamaniuk, M.Z.; Roneker, C.A.; Li, L.; Lei, X.G. Dietary selenium deficiency partially rescues type 2 diabetes-like phenotypes of glutathione peroxidase-1-overexpressing male mice. J. Nutr. 2012, 142, 1975–1982. [Google Scholar]
- Cockerham, C.C. An extension of the concept of partitioning hereditary variance for analysis of covariances among relatives when epistasis is present. Genetics 1954, 39, 859–882. [Google Scholar]
- Loscalzo, J.; Barabasi, A.L. Systems biology and the future of medicine. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 619–627. [Google Scholar] [CrossRef]
- Van Ommen, B.; Fairweather-Tait, S.; Freidig, A.; Kardinaal, A.; Scalbert, A.; Wopereis, S. A network biology model of micronutrient related health. Br. J. Nutr. 2008, 99, S72–S80. [Google Scholar]
- Van Ommen, B.; El-Sohemy, A.; Hesketh, J.; Kaput, J.; Fenech, M.; Evelo, C.T.; McArdle, H.J.; Bouwman, J.; Lietz, G.; Mathers, J.C.; et al. The micronutrient genomics project: A community-driven knowledge base for micronutrient research. Genes Nutr. 2010, 5, 285–296. [Google Scholar] [CrossRef]
- Wikipathway Web site. Available online: http://wikipathways.org/index.php/Pathway:WP15 (accessed on 18 January 2013).
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Joseph, J.; Loscalzo, J. Selenistasis: Epistatic Effects of Selenium on Cardiovascular Phenotype. Nutrients 2013, 5, 340-358. https://doi.org/10.3390/nu5020340
Joseph J, Loscalzo J. Selenistasis: Epistatic Effects of Selenium on Cardiovascular Phenotype. Nutrients. 2013; 5(2):340-358. https://doi.org/10.3390/nu5020340
Chicago/Turabian StyleJoseph, Jacob, and Joseph Loscalzo. 2013. "Selenistasis: Epistatic Effects of Selenium on Cardiovascular Phenotype" Nutrients 5, no. 2: 340-358. https://doi.org/10.3390/nu5020340
APA StyleJoseph, J., & Loscalzo, J. (2013). Selenistasis: Epistatic Effects of Selenium on Cardiovascular Phenotype. Nutrients, 5(2), 340-358. https://doi.org/10.3390/nu5020340