Advances in Nutritional Research on Regulatory T-Cells


Abstract
:1. Introduction to T-Cell Immunology

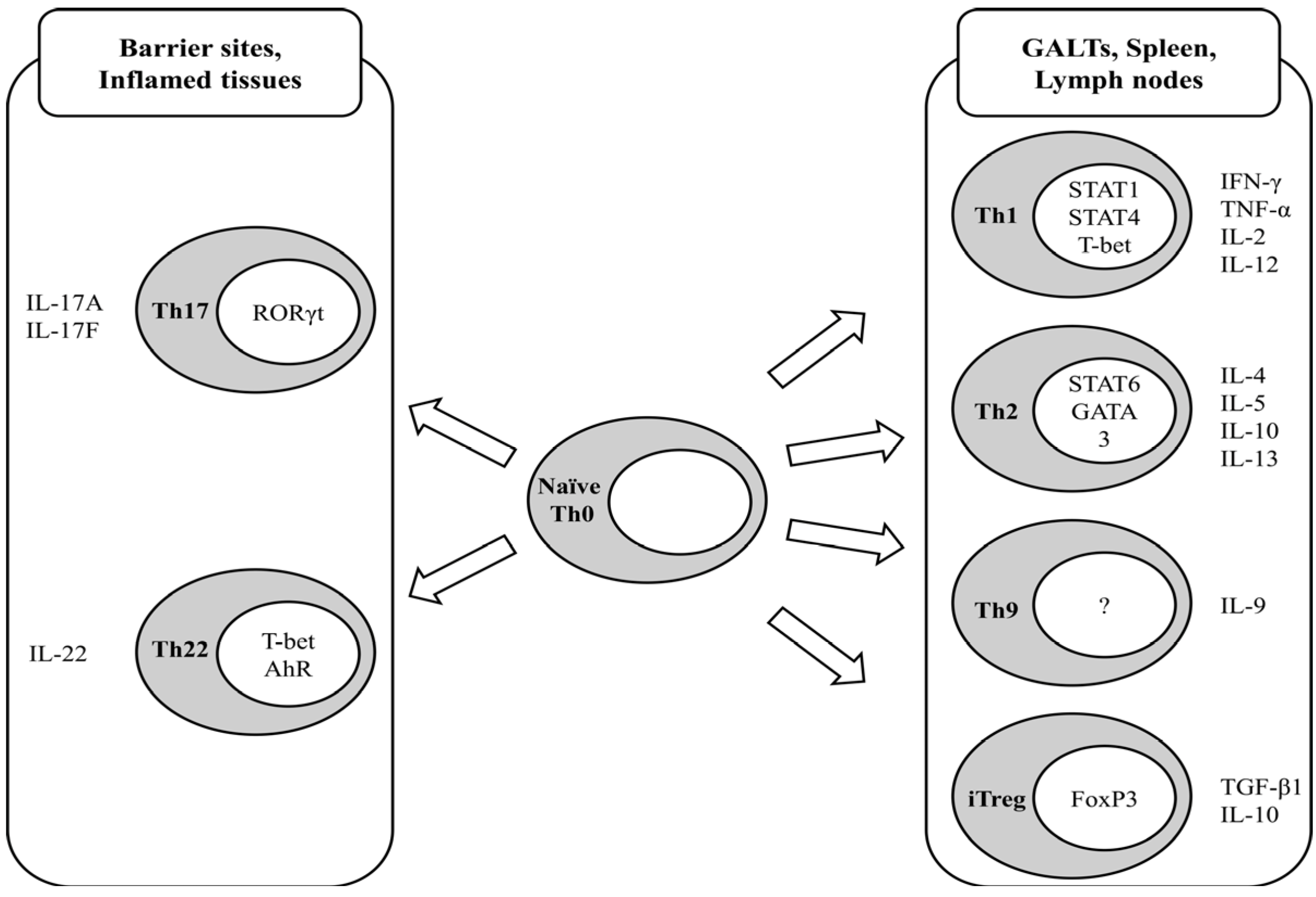
2. Regulatory T-Cells
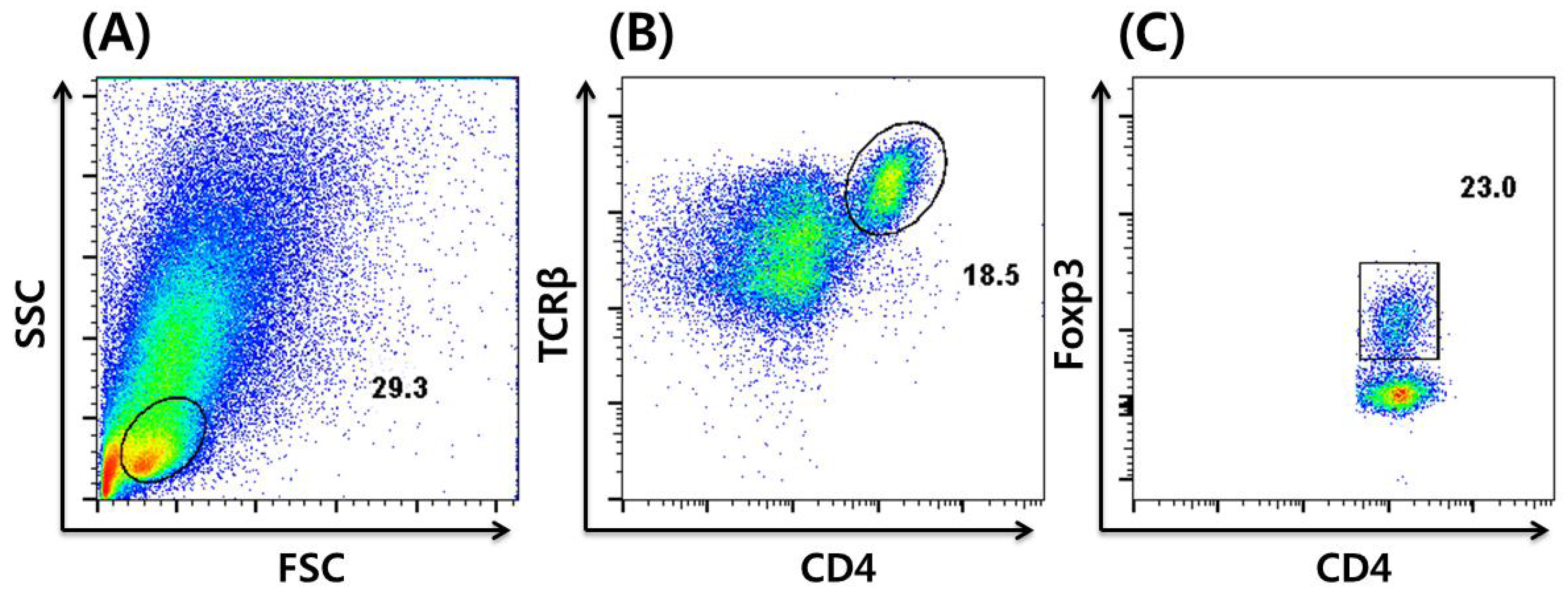
3. In vitro Induction and Intracellular Staining of iTregs
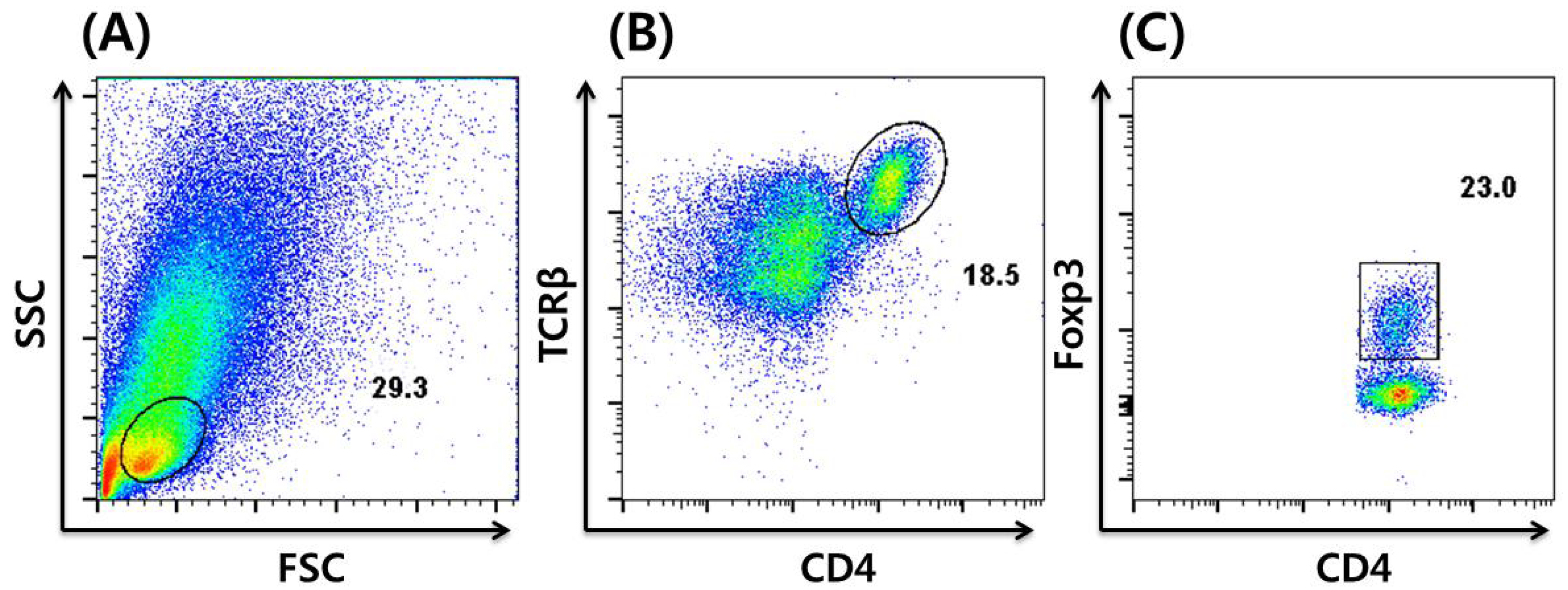
4. Dietary Regulation of Tregs
4.1. Vitamins
4.2. Dietary Fatty Acids
4.3. Dietary Polyphenols
5. Conclusions

{kind=link}
{kind=link}
| Dietary component | Function | Reference |
|---|---|---|
| Vitamin A | Increased Treg population in human biopsy culture from ulcerative colitis patients in vitro. | [26] |
| Amelioration of TNBS-induced murine colitis model in vivo. | ||
| Vitamin B9 | Essential for murine Treg survival ex vivo. | [30] |
| Increased Treg population in murine small intestine in vivo. | ||
| Omega-3 PUFA | Enhanced development of murine Tregs in vivo. | [37] |
| Increased Tregs in a murine transplant model in vivo. | [38] | |
| Increased ability of murine dendritic cells to convert Tregs in vitro. | [39] | |
| Decreased regulatory function of murine Tregs ex vivo. | [40] | |
| Polyphenols | ||
| Naringenin | Increased murine Treg population in vivo. | [44] |
| EGCG 1 | Increased murine Treg population in vivo. | [47] |
| FGM 2 | Increased human Treg population in vivo. | [48] |
Acknowledgments
Conflicts of Interest
References
- Jager, A.; Kuchroo, V.K. Effector and regulatory T-cell subsets in autoimmunity and tissue inflammation. Scand. J. Immunol. 2010, 72, 173–184. [Google Scholar] [CrossRef]
- Stout, R.D.; Bottomly, K. Antigen-specific activation of effector macrophages by IFN-gamma producing (TH1) T cell clones. Failure of IL-4-producing (TH2) T cell clones to activate effector function in macrophages. J. Immunol. 1989, 142, 760–765. [Google Scholar]
- Heinzel, F.P.; Sadick, M.D.; Mutha, S.S.; Locksley, R.M. Production of interferon gamma, interleukin 2, interleukin 4, and interleukin 10 by CD4+ lymphocytes in vivo during healing and progressive murine leishmaniasis. Proc. Natl. Acad. Sci. USA 1991, 88, 7011–7015. [Google Scholar] [CrossRef]
- Chen, Z.; Lin, F.; Gao, Y.; Li, Z.; Zhang, J.; Xing, Y.; Deng, Z.; Yao, Z.; Tsun, A.; Li, B. FOXP3 and RORgammat: Transcriptional regulation of Treg and Th17. Int. Immunopharmacol. 2011, 11, 536–542. [Google Scholar] [CrossRef]
- Chang, Q.; Wang, Y.K.; Zhao, Q.; Wang, C.Z.; Hu, Y.Z.; Wu, B.Y. Th17 cells are increased with severity of liver inflammation in patients with chronic hepatitis C. J. Gastroenterol. Hepatol. 2012, 27, 273–278. [Google Scholar] [CrossRef]
- Murdaca, G.; Colombo, B.M.; Puppo, F. The role of Th17 lymphocytes in the autoimmune and chronic inflammatory diseases. Intern. Emerg. Med. 2011, 6, 487–495. [Google Scholar] [CrossRef]
- Caprioli, F.; Pallone, F.; Monteleone, G. Th17 immune response in IBD: A new pathogenic mechanism. J. Crohn’s Colitis 2008, 2, 291–295. [Google Scholar] [CrossRef]
- Seiderer, J.; Elben, I.; Diegelmann, J.; Glas, J.; Stallhofer, J.; Tillack, C.; Pfennig, S.; Jurgens, M.; Schmechel, S.; Konrad, A.; et al. Role of the novel Th17 cytokine IL-17F in inflammatory bowel disease (IBD): Upregulated colonic IL-17F expression in active Crohn’s disease and analysis of the IL17F p.His161Arg polymorphism in IBD. Inflamm. Bowel Dis. 2008, 14, 437–445. [Google Scholar] [CrossRef]
- Basu, R.; O’Quinn, D.B.; Silberger, D.J.; Schoeb, T.R.; Fouser, L.; Ouyang, W.; Hatton, R.D.; Weaver, C.T. Th22 cells are an important source of IL-22 for host protection against enteropathogenic bacteria. Immunity 2012, 37, 1061–1075. [Google Scholar] [CrossRef]
- Dardalhon, V.; Awasthi, A.; Kwon, H.; Galileos, G.; Gao, W.; Sobel, R.A.; Mitsdoerffer, M.; Strom, T.B.; Elyaman, W.; Ho, I.C.; et al. IL-4 inhibits TGF-beta-induced Foxp3+ T cells and, together with TGF-beta, generates IL-9+ IL-10+ Foxp3(-) effector T cells. Nat. Immunol. 2008, 9, 1347–1355. [Google Scholar] [CrossRef]
- Veldhoen, M.; Uyttenhove, C.; van Snick, J.; Helmby, H.; Westendorf, A.; Buer, J.; Martin, B.; Wilhelm, C.; Stockinger, B. Transforming growth factor-beta ‘reprograms’ the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nat. Immunol. 2008, 9, 1341–1346. [Google Scholar] [CrossRef]
- Kaplan, M.H. Th9 cells: Differentiation and disease. Immunol. Rev. 2013, 252, 104–115. [Google Scholar] [CrossRef]
- Baker, P.J. Homeostatic control of antibody responses: A model based on the recognition of cell-associated antibody by regulatory T cells. Transplant. Rev. 1975, 26, 3–20. [Google Scholar]
- Baumgart, M.; Tompkins, F.; Leng, J.; Hesse, M. Naturally occurring CD4+Foxp3+ regulatory T cells are an essential, IL-10-independent part of the immunoregulatory network in Schistosoma mansoni egg-induced inflammation. J. Immunol. 2006, 176, 5374–5387. [Google Scholar]
- Yuan, X.; Malek, T.R. Cellular and molecular determinants for the development of natural and induced regulatory T cells. Hum. Immunol. 2012, 73, 773–782. [Google Scholar] [CrossRef]
- Thornton, A.M.; Korty, P.E.; Tran, D.Q.; Wohlfert, E.A.; Murray, P.E.; Belkaid, Y.; Shevach, E.M. Expression of Helios, an Ikaros transcription factor family member, differentiates thymic-derived from peripherally induced Foxp3+ T regulatory cells. J. Immunol. 2010, 184, 3433–3441. [Google Scholar] [CrossRef]
- Himmel, M.E.; MacDonald, K.G.; Garcia, R.V.; Steiner, T.S.; Levings, M.K. Helios+ and Helios− cells coexist within the natural FOXP3+ T regulatory cell subset in humans. J. Immunol. 2013, 190, 2001–2008. [Google Scholar] [CrossRef]
- Curotto de Lafaille, M.A.; Lafaille, J.J. Natural and adaptive foxp3+ regulatory T cells: More of the same or a division of labor? Immunity 2009, 30, 626–635. [Google Scholar]
- Toda, A.; Piccirillo, C.A. Development and function of naturally occurring CD4+CD25+ regulatory T cells. J. Leukoc. Biol. 2006, 80, 458–470. [Google Scholar] [CrossRef]
- Tanoue, T.; Honda, K. Induction of Treg cells in the mouse colonic mucosa: A central mechanism to maintain host-microbiota homeostasis. Semin. Immunol. 2012, 24, 50–57. [Google Scholar] [CrossRef]
- Kurzweil, V.; Tarangelo, A.; Oliver, P.M. Gastrointestinal microbiota do not significantly contribute to T cell activation or GI inflammation in Ndfip1-cKO mice. PLoS One 2012, 7, e34478. [Google Scholar] [CrossRef]
- Khounlotham, M.; Kim, W.; Peatman, E.; Nava, P.; Medina-Contreras, O.; Addis, C.; Koch, S.; Fournier, B.; Nusrat, A.; Denning, T.L.; et al. Compromised intestinal epithelial barrier induces adaptive immune compensation that protects from colitis. Immunity 2012, 37, 563–573. [Google Scholar] [CrossRef]
- Koch, U.; Radtke, F. Mechanisms of T cell development and transformation. Annu. Rev. Cell Dev. Biol. 2011, 27, 539–562. [Google Scholar] [CrossRef]
- Fantini, M.C.; Dominitzki, S.; Rizzo, A.; Neurath, M.F.; Becker, C. In vitro generation of CD4+ CD25+ regulatory cells from murine naive T cells. Nat. Protoc. 2007, 2, 1789–1794. [Google Scholar] [CrossRef]
- Maden, M. Retinoid signalling in the development of the central nervous system. Nat. Rev. Neurosci. 2002, 3, 843–853. [Google Scholar] [CrossRef]
- Bai, A.; Lu, N.; Guo, Y.; Liu, Z.; Chen, J.; Peng, Z. All-trans retinoic acid down-regulates inflammatory responses by shifting the Treg/Th17 profile in human ulcerative and murine colitis. J. Leukoc. Biol. 2009, 86, 959–969. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Y.; Liu, Q.; Zhong, W.; Xia, Z. All-trans retinoic acid attenuates airway inflammation by inhibiting Th2 and Th17 response in experimental allergic asthma. BMC Immunol. 2013, 14, 28. [Google Scholar] [CrossRef]
- Park, M.K.; Jhun, J.Y.; Lee, S.Y.; Oh, H.J.; Park, M.J.; Byun, J.K.; Yoon, B.Y.; Park, E.M.; Lee, D.G.; Kwok, S.K.; et al. Retinal attenuates inflammatory arthritis by reciprocal regulation of IL-17-producing T cells and Foxp3(+) regulatory T cells and the inhibition of osteoclastogenesis. Immunol. Lett. 2012, 148, 59–68. [Google Scholar] [CrossRef]
- Chang, J.; Thangamani, S.; Kim, M.H.; Ulrich, B.; Morris, S.M., Jr.; Kim, C.H. Retinoic acid promotes the development of Arg1-expressing dendritic cells for the regulation of T-cell differentiation. Eur. J. Immunol. 2013, 43, 967–978. [Google Scholar] [CrossRef]
- Kunisawa, J.; Hashimoto, E.; Ishikawa, I.; Kiyono, H. A pivotal role of vitamin B9 in the maintenance of regulatory T cells in vitro and in vivo. PLoS One 2012, 7, e32094. [Google Scholar]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2012, 75, 645–662. [Google Scholar]
- Cabre, E.; Manosa, M.; Gassull, M.A. Omega-3 fatty acids and inflammatory bowel diseases—A systematic review. Br. J. Nutr. 2012, 107 (Suppl. 2), 240–252. [Google Scholar] [CrossRef]
- Maroufyan, E.; Kasim, A.; Ebrahimi, M.; Loh, T.C.; Bejo, M.H.; Zerihun, H.; Hosseni, F.; Goh, Y.M.; Farjam, A.S. Omega-3 polyunsaturated fatty acids enrichment alters performance and immune response in infectious bursal disease challenged broilers. Lipids Health Dis. 2012, 11, 15. [Google Scholar] [CrossRef]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef]
- Sorokovoi, K.V.; Pogozheva, A.V.; Sergeeva, K.V. Effect of diet including omega-3 polyunsaturated fatty acids on the status of T-cell immunity in patients with ischemic heart disease and hyperproteinemia. Vopr. Pitan. 1997, 5, 6–8. [Google Scholar]
- Terada, S.; Takizawa, M.; Yamamoto, S.; Ezaki, O.; Itakura, H.; Akagawa, K.S. Suppressive mechanisms of EPA on human T cell proliferation. Microbiol. Immunol. 2001, 45, 473–481. [Google Scholar]
- Molvig, J.; Pociot, F.; Worsaae, H.; Wogensen, L.D.; Baek, L.; Christensen, P.; Mandrup-Poulsen, T.; Andersen, K.; Madsen, P.; Dyerberg, J.; et al. Dietary supplementation with omega-3-polyunsaturated fatty acids decreases mononuclear cell proliferation and interleukin-1 beta content but not monokine secretion in healthy and insulin-dependent diabetic individuals. Scand. J. Immunol. 1991, 34, 399–410. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 fatty acids in health and disease and in growth and development. Am. J. Clin. Nutr. 1991, 54, 438–463. [Google Scholar]
- Curtis, C.L.; Rees, S.G.; Little, C.B.; Flannery, C.R.; Hughes, C.E.; Wilson, C.; Dent, C.M.; Otterness, I.G.; Harwood, J.L.; Caterson, B. Pathologic indicators of degradation and inflammation in human osteoarthritic cartilage are abrogated by exposure to n-3 fatty acids. Arthritis Rheum. 2002, 46, 1544–1553. [Google Scholar] [CrossRef]
- Han, S.C.; Kang, G.J.; Ko, Y.J.; Kang, H.K.; Moon, S.W.; Ann, Y.S.; Yoo, E.S. Fermented fish oil suppresses T helper 1/2 cell response in a mouse model of atopic dermatitis via generation of CD4+CD25+Foxp3+ T cells. BMC Immunol. 2012, 13, 44. [Google Scholar] [CrossRef]
- Iwami, D.; Nonomura, K.; Shirasugi, N.; Niimi, M. Immunomodulatory effects of eicosapentaenoic acid through induction of regulatory T cells. Int. Immunopharmacol. 2011, 11, 384–389. [Google Scholar] [CrossRef]
- Kong, W.; Yen, J.H.; Ganea, D. Docosahexaenoic acid prevents dendritic cell maturation, inhibits antigen-specific Th1/Th17 differentiation and suppresses experimental autoimmune encephalomyelitis. Brain Behav. Immun. 2011, 25, 872–882. [Google Scholar] [CrossRef]
- Yessoufou, A.; Ple, A.; Moutairou, K.; Hichami, A.; Khan, N.A. Docosahexaenoic acid reduces suppressive and migratory functions of CD4+CD25+ regulatory T-cells. J. Lipid Res. 2009, 50, 2377–2388. [Google Scholar] [CrossRef]
- Wang, H.K.; Yeh, C.H.; Iwamoto, T.; Satsu, H.; Shimizu, M.; Totsuka, M. Dietary flavonoid naringenin induces regulatory T cells via an aryl hydrocarbon receptor mediated pathway. J. Agric. Food Chem. 2012, 60, 2171–2178. [Google Scholar] [CrossRef]
- Assini, J.M.; Mulvihill, E.E.; Sutherland, B.G.; Telford, D.E.; Sawyez, C.G.; Felder, S.L.; Chhoker, S.S.; Edwards, J.Y.; Gros, R.; Huff, M.W. Naringenin prevents cholesterol-induced systemic inflammation, metabolic dysregulation and atherosclerosis in Ldlr-/- mice. J. Lipid Res. 2012, 54, 711–724. [Google Scholar]
- Shi, Y.; Dai, J.; Liu, H.; Li, R.R.; Sun, P.L.; Du, Q.; Pang, L.L.; Chen, Z.; Yin, K.S. Naringenin inhibits allergen-induced airway inflammation and airway responsiveness and inhibits NF-kappaB activity in a murine model of asthma. Can. J. Physiol. Pharmacol. 2009, 87, 729–735. [Google Scholar] [CrossRef]
- Wang, J.; Ren, Z.; Xu, Y.; Xiao, S.; Meydani, S.N.; Wu, D. Epigallocatechin-3-gallate ameliorates experimental autoimmune encephalomyelitis by altering balance among CD4+ T-cell subsets. Am. J. Pathol. 2012, 180, 221–234. [Google Scholar] [CrossRef]
- Magrone, T.; Jirillo, E. Influence of polyphenols on allergic immune reactions: Mechanisms of action. Proc. Nutr. Soc. 2012, 71, 316–321. [Google Scholar] [CrossRef]
- Issazadeh-Navikas, S.; Teimer, R.; Bockermann, R. Influence of dietary components on regulatory T cells. Mol. Med. 2012, 18, 95–110. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, W.; Lee, H. Advances in Nutritional Research on Regulatory T-Cells. Nutrients 2013, 5, 4305-4315. https://doi.org/10.3390/nu5114305
Kim W, Lee H. Advances in Nutritional Research on Regulatory T-Cells. Nutrients. 2013; 5(11):4305-4315. https://doi.org/10.3390/nu5114305
Chicago/Turabian StyleKim, Wooki, and Hyungjae Lee. 2013. "Advances in Nutritional Research on Regulatory T-Cells" Nutrients 5, no. 11: 4305-4315. https://doi.org/10.3390/nu5114305
APA StyleKim, W., & Lee, H. (2013). Advances in Nutritional Research on Regulatory T-Cells. Nutrients, 5(11), 4305-4315. https://doi.org/10.3390/nu5114305