
Sugarcane Molasses Polyphenol Extract Attenuates Alcohol-Induced Chronic Liver Damage via Antioxidant, Anti-Inflammatory, and CYP2E1/Keap1/NF-κB Pathway Modulation


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Tests
2.3. Liver Index Analysis
2.4. Biochemical Analysis
2.5. Histological Analysis
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Western Blot
2.8. Statistical Analysis
3. Results
3.1. The Effect of SP Nutritional Intervention on the Body Weights and Liver Index of the ALD Mice
3.2. The Effect of SP Nutritional Intervention on the Serum Transaminase and Lipid Levels in the ALD Mice
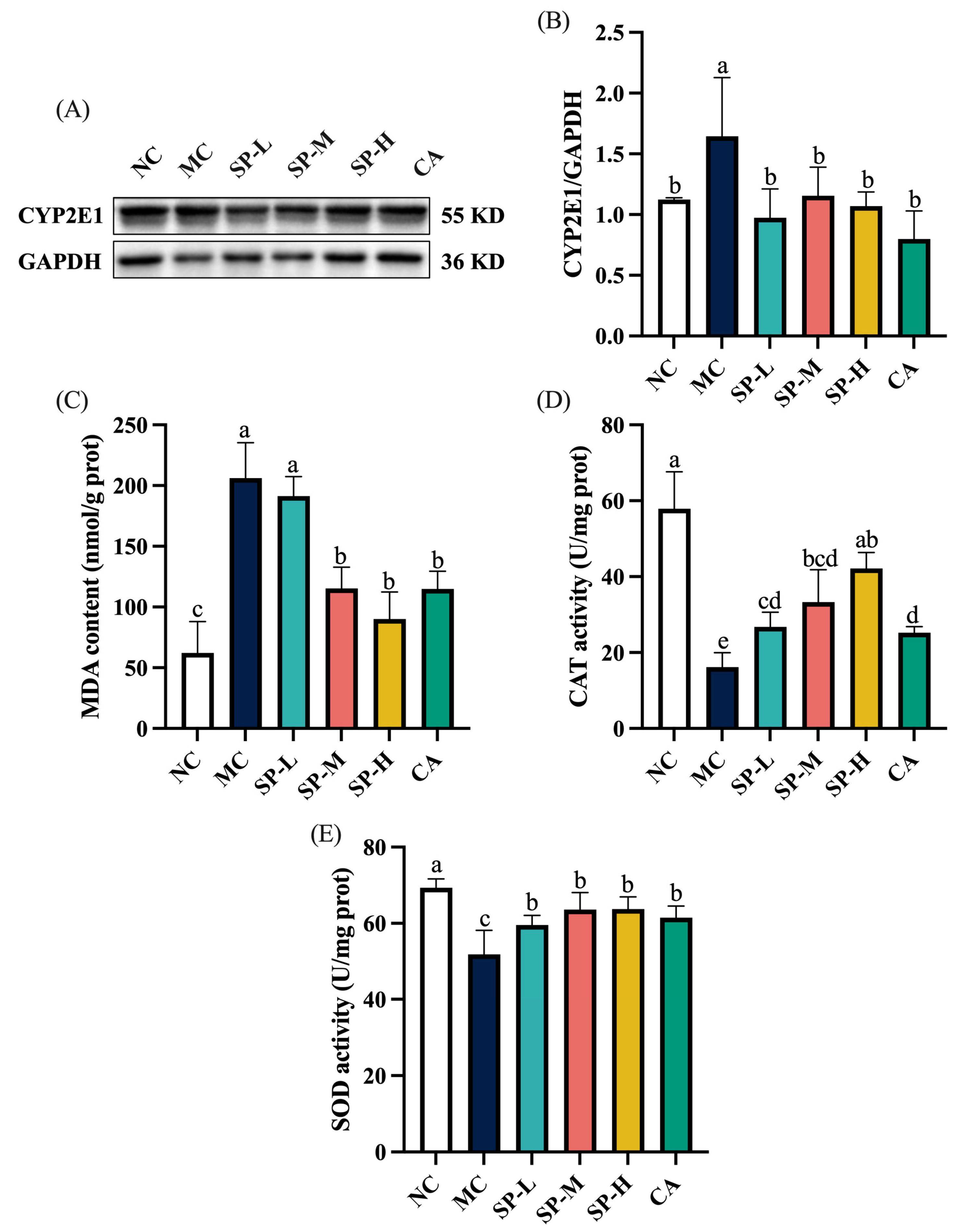
3.3. The Effect of SP Nutritional Intervention on the Liver CYP2E1 Content and Oxidative Stress in the ALD Mice
3.4. The Effect of SP Nutritional Intervention on the Liver Pathology and Inflammatory Factors in the ALD Mice
3.5. The Effect of SP Nutritional Intervention on the Keap1/HO-1 Signaling Pathway in the ALD Mice
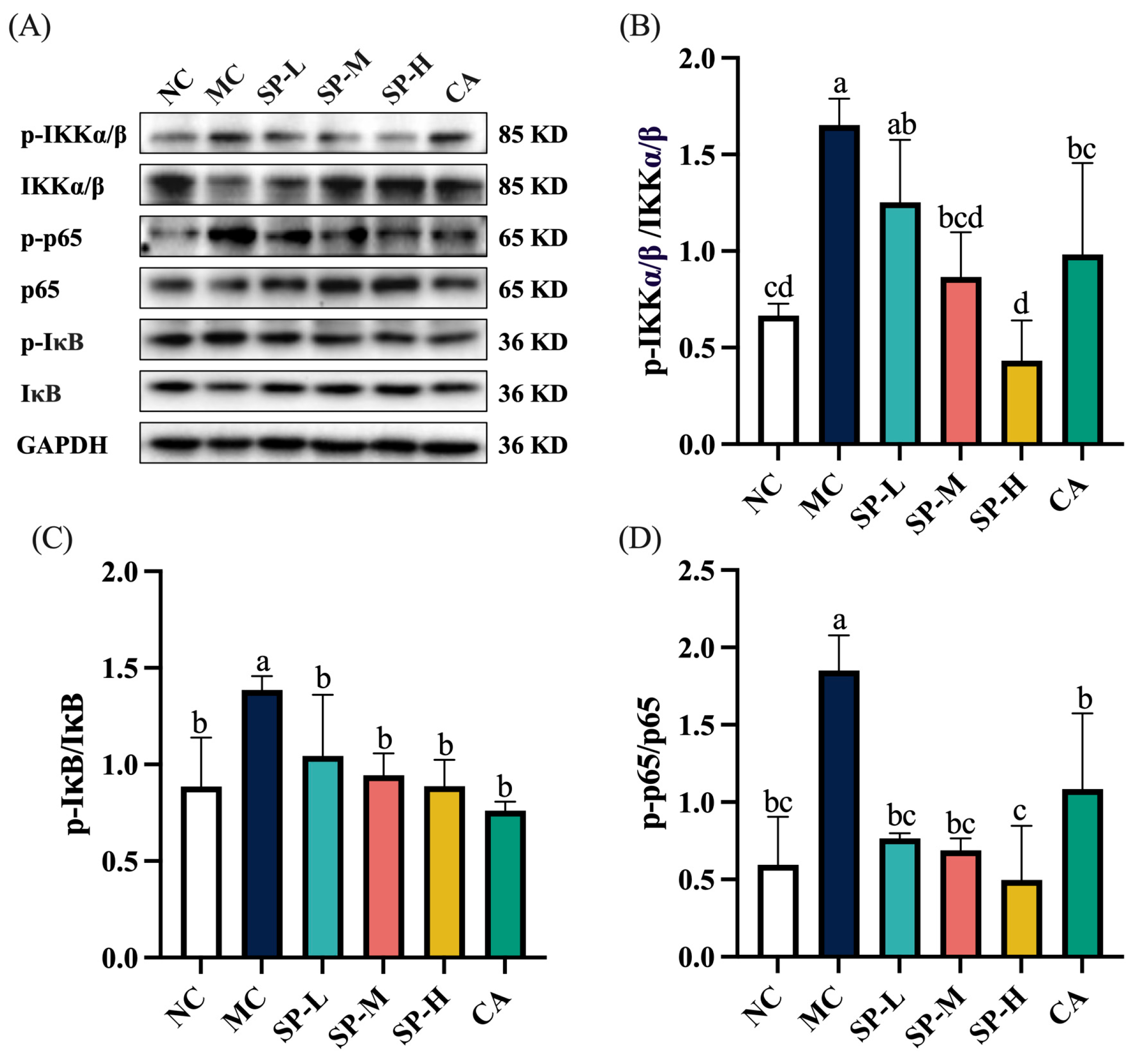
3.6. The Effect of SP Nutritional Intervention on the NF-κB Signaling Pathway in the ALD Mice
4. Discussion
4.1. SP Regulation of the Transaminase Activity and Lipid Accumulation
4.2. SP Modulation of CYP2E1-Mediated Alcohol Metabolism and Oxidative Stress
4.3. SP Influence on the Keap1/HO-1 Signaling Pathway and Antioxidant Defense Mechanisms
4.4. SP Intervention in the Inflammatory Response and Regulation of the NF-κB Signaling Pathway
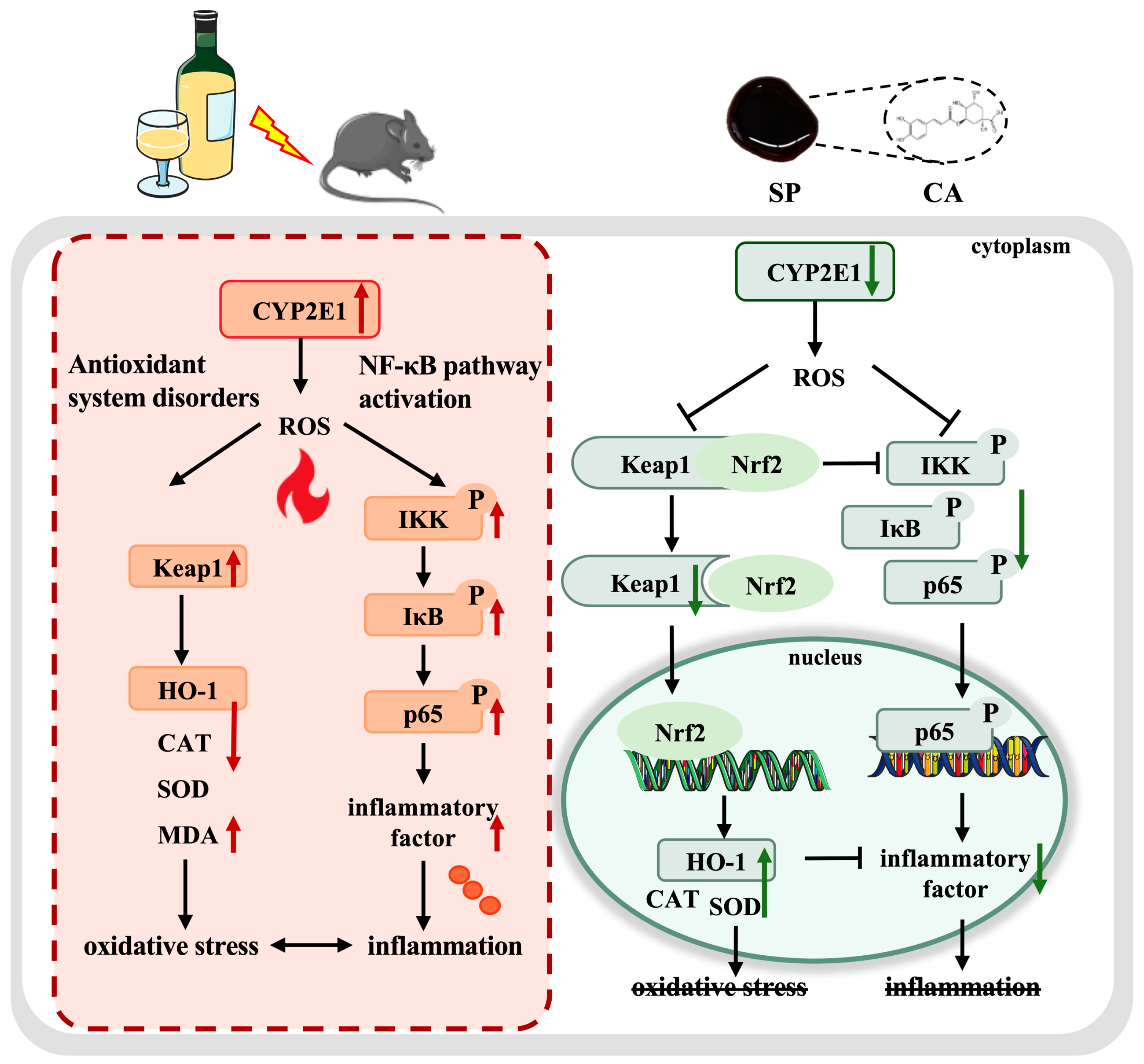
4.5. SP Intervention in Multi-Target Modulation of the CYP2E1/Keap1/NF-κB Signaling Pathway
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seitz, H.K.; Bataller, R.; Cortez-Pinto, H.; Gao, B.; Gual, A.; Lackner, C.; Mathurin, P.; Mueller, S.; Szabo, G.; Tsukamoto, H. Alcoholic liver disease. Nat. Rev. Dis. Primers. 2018, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.Z.; Chandimali, N.; Han, Y.H.; Lee, D.H.; Kim, J.S.; Kim, S.U.; Kim, T.D.; Jeong, D.K.; Sun, H.N.; Lee, D.S.; et al. Pathogenesis, Early Diagnosis, and Therapeutic Management of Alcoholic Liver Disease. Int. J. Mol. Sci. 2019, 20, 2712. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, H.F.J. Alcohol and Human Health: What Is the Evidence? Annu. Rev. Food Sci. Technol. 2020, 11, 1–21. [Google Scholar] [CrossRef]
- Leung, T.M.; Nieto, N. CYP2E1 and oxidant stress in alcoholic and non-alcoholic fatty liver disease. J. Hepatol. 2013, 58, 395–398. [Google Scholar] [CrossRef]
- Lai, W.; Zhang, J.; Sun, J.; Min, T.; Bai, Y.; He, J.; Cao, H.; Che, Q.; Guo, J.; Su, Z. Oxidative stress in alcoholic liver disease, focusing on proteins, nucleic acids, and lipids: A review. Int. J. Biol. Macromol. 2024, 278, 134809. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Yang, Y.M.; Cho, Y.E.; Hwang, S. Crosstalk between Oxidative Stress and Inflammatory Liver Injury in the Pathogenesis of Alcoholic Liver Disease. Int. J. Mol. Sci. 2022, 23, 774. [Google Scholar] [CrossRef]
- Wu, S.; Liao, X.; Zhu, Z.; Huang, R.; Chen, M.; Huang, A.; Zhang, J.; Wu, Q.; Wang, J.; Ding, Y. Antioxidant and anti-inflammation effects of dietary phytochemicals: The Nrf2/NF-KB signalling pathway and upstream factors of Nrf2. Phytochemistry 2022, 204, 113429. [Google Scholar] [CrossRef]
- Lingappan, K. NF-kappaB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Tao, K.X.; Peng, H.; Bi, X.; Wang, X.Y.; Tian, Y.; Zhang, H.; Sun, Y. Effect of Tetrastigma hemsleyanum leaves and Extract supplementation on liver metabolomics and the gut microbiota in alcohol-induced liver injury. Food Biosci. 2024, 58, 103617. [Google Scholar] [CrossRef]
- Xu, T.; Hu, S.; Liu, Y.; Sun, K.; Luo, L.; Zeng, L. Hawk Tea Flavonoids as Natural Hepatoprotective Agents Alleviate Acute Liver Damage by Reshaping the Intestinal Microbiota and Modulating the Nrf2 and NF-κB Signaling Pathways. Nutrients 2022, 14, 3662. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Wang, Z.; He, Z.; Wang, Z.; Chen, Q.; Qin, F.; Zeng, M.; Chen, J. Pine pollen extract alleviates ethanol-induced oxidative stress and apoptosis in HepG2 cells via MAPK signaling. Food Chem. Toxicol. 2023, 171, 113550. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, B.; Meng, X.; Zhang, Y.; Xuan, F.; Li, L.; Li, S.; Xu, Y.; Wang, Y. Dietary polyphenols alleviate alcoholic liver injury: Potential mechanisms, product development status, and future research directions. Phytochem. Rev. 2025, 1–16. [Google Scholar] [CrossRef]
- Zhao, L.; Mehmood, A.; Yuan, D.D.; Usman, M.; Murtaza, M.A.; Yaqoob, S.; Wang, C.T. Protective Mechanism of Edible Food Plants against Alcoholic Liver Disease with Special Mention to Polyphenolic Compounds. Nutrients 2021, 13, 1612. [Google Scholar] [CrossRef]
- Kataoka, S.; Mukai, Y.; Takebayashi, M.; Kudo, M.; Acuram, U.R.; Kurasaki, M.; Sato, S. Melinjo (Gnetum gnemon) extract intake during lactation stimulates hepatic AMP-activated protein kinase in offspring of excessive fructose-fed pregnant rats. Reprod. Biol. 2016, 16, 165–173. [Google Scholar] [CrossRef]
- Lee, Y.J.; Hsu, J.D.; Lin, W.L.; Kao, S.H.; Wang, C.J. Upregulation of caveolin-1 by mulberry leaf extract and its major components, chlorogenic acid derivatives, attenuates alcoholic steatohepatitis via inhibition of oxidative stress. Food Funct. 2017, 8, 397–405. [Google Scholar] [CrossRef]
- Gao, Y.; Zong, Z.H.; Xia, W.; Fang, X.J.; Liu, R.L.; Wu, W.J.; Mu, H.L.; Han, Y.C.; Xiao, S.Y.; Gao, H.Y.; et al. Hepatoprotective effect of water bamboo shoot (Zizania latifolia) extracts against acute alcoholic liver injury in a mice model and screening of bioactive phytochemicals. Food Front. 2023, 4, 1362–1371. [Google Scholar] [CrossRef]
- Kumar, T.; Wang, J.G.; Xu, C.H.; Lu, X.; Mao, J.; Lin, X.Q.; Kong, C.Y.; Li, C.J.; Li, X.J.; Tian, C.Y.; et al. Genetic Engineering for Enhancing Sugarcane Tolerance to Biotic and Abiotic Stresses. Plants 2024, 13, 1739. [Google Scholar] [CrossRef]
- Hewawansa, U.H.A.J.; Houghton, M.J.; Barber, E.; Costa, R.J.S.; Kitchen, B.; Williamson, G. Flavonoids and phenolic acids from sugarcane: Distribution in the plant, changes during processing, and potential benefits to industry and health. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13307. [Google Scholar] [CrossRef]
- Kassa, M.G.; Asemu, A.M.; Belachew, M.T.; Satheesh, N.; Abera, B.D.; Teferi, D.A. Review on the application, health usage, and negative effects of molasses. CyTA J. Food. 2024, 22, 2321984. [Google Scholar] [CrossRef]
- Wang, J.; Wang, M.; Zhang, C.; Li, W.; Zhang, T.; Zhou, Y.; Flavel, M.; Xi, Y.; Li, H.; Liu, X. Protective effects of sugarcane polyphenol against UV-B-induced photoaging in Balb/c mouse skin: Antioxidant, anti-inflammatory, and anti-glycosylation Effects. J. Food Sci. 2024, 89, 3048–3063. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J.; Li, W.; Zhang, C.; Xi, Y.; Zhou, Y.; Liu, X.; Li, H. Protective effects of polyphenol-rich sugarcane molasses extract against AGEs-induced glycosylation damage in L929 cells. Food Biosci. 2025, 63, 105775. [Google Scholar] [CrossRef]
- Wang, W.; Xu, C.; Wang, Q.; Hussain, M.A.; Wang, C.; Hou, J.; Jiang, Z. Protective Effect of Polyphenols, Protein, Peptides, and Polysaccharides on Alcoholic Liver Disease: A Review of Research Status and Molecular Mechanisms. J. Agric. Food Chem. 2023, 71, 5861–5883. [Google Scholar] [CrossRef]
- Zogona, D.; Zongo, A.W.S.; Elkhedir, A.E.; Salah, M.; Tao, M.; Li, R.; Wu, T.; Xu, X. Red raspberry supplementation mitigates alcohol-induced liver injury associated with gut microbiota alteration and intestinal barrier dysfunction in mice. Food Funct. 2023, 14, 1209–1226. [Google Scholar] [CrossRef]
- Mao, J.; Tan, L.; Tian, C.; Wang, W.; Zhang, H.; Zhu, Z.; Li, Y. Research progress on rodent models and its mechanisms of liver injury. Life Sci. 2024, 337, 122343. [Google Scholar] [CrossRef]
- Xia, T.; Zhang, B.; Li, S.; Fang, B.; Duan, W.; Zhang, J.; Song, J.; Wang, M. Vinegar extract ameliorates alcohol-induced liver damage associated with the modulation of gut microbiota in mice. Food Funct. 2020, 11, 2898–2909. [Google Scholar] [CrossRef]
- Cederbaum, A.I.; Lu, Y.; Wang, X.; Wu, D. Synergistic toxic interactions between CYP2E1, LPS/TNFα, and JNK/p38 MAP kinase and their implications in alcohol-induced liver injury. Adv. Exp. Med. Biol. 2015, 815, 145–172. [Google Scholar]
- Wang, C.; Ma, C.; Gong, L.; Guo, Y.; Fu, K.; Zhang, Y.; Zhou, H.; Li, Y. Macrophage Polarization and Its Role in Liver Disease. Front. Immunol. 2021, 12, 803037. [Google Scholar] [CrossRef]
- Gao, B.; Ahmad, M.F.; Nagy, L.E.; Tsukamoto, H. Inflammatory pathways in alcoholic steatohepatitis. J. Hepatol. 2019, 70, 249–259. [Google Scholar] [CrossRef]
- Cai, Z.; Song, L.H.; Qian, B.J.; Xu, W.; Ren, J.K.; Jing, P.; Oey, I. Understanding the effect of anthocyanins extracted from purple sweet potatoes on alcohol-induced liver injury in mice. Food Chem. 2018, 245, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Sun, Z.Q.; Wang, X.L.; Li, M.J.; Zhou, B.R.; Zhang, X.S. Solanum nigrum L. berries extract ameliorated the alcoholic liver injury by regulating gut microbiota, lipid metabolism, inflammation, and oxidative stress. Food Res. Int. 2024, 188, 114489. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Pan, J.H.; Kim, S.H.; Lee, J.H.; Park, J.-W. Chlorogenic acid ameliorates alcohol-induced liver injuries through scavenging reactive oxygen species. Biochimie 2018, 150, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lee, J.S.; Lee, I.H.; Hong, J.T. Therapeutic potency of fermented field water-dropwort (Oenanthe javanica (Blume) DC.) in ethanol-induced liver injury. RSC Adv. 2020, 10, 1544–1551. [Google Scholar] [CrossRef]
- Lv, X.C.; Wu, Q.; Cao, Y.J.; Lin, Y.C.; Guo, W.L.; Rao, P.F.; Zhang, Y.Y.; Chen, Y.T.; Ai, L.Z.; Ni, L. Ganoderic acid A from Ganoderma lucidum protects against alcoholic liver injury through ameliorating the lipid metabolism and modulating the intestinal microbial composition. Food Funct. 2022, 13, 5820–5837. [Google Scholar] [CrossRef]
- Simón, J.; Casado-Andrés, M.; Goikoetxea-Usandizaga, N.; Serrano-Maciá, M.; Martínez-Chantar, M.L. Nutraceutical Properties of Polyphenols against Liver Diseases. Nutrients 2020, 12, 3517. [Google Scholar] [CrossRef]
- Hou, R.L.; Liu, X.; Wu, X.P.; Zheng, M.F.; Fu, J.S. Therapeutic effect of natural melanin from edible fungus Auricularia auricula on alcohol-induced liver damage in vitro and in vivo. Food Sci. Hum. Wellness. 2021, 10, 514–522. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, Z.; Yang, M.; Wang, Y.; Cao, J.; Khan, A.; Liu, Y.; Cheng, G. Protective effects of E Se tea extracts against alcoholic fatty liver disease induced by high fat/alcohol diet: In vivo biological evaluation and molecular docking study. Phytomedicine 2022, 101, 154113. [Google Scholar] [CrossRef]
- Wang, W.; Shang, H.; Li, J.; Ma, Y.; Xu, C.; Ma, J.; Hou, J.; Jiang, Z. Four Different Structural Dietary Polyphenols, Especially Dihydromyricetin, Possess Superior Protective Effect on Ethanol-Induced ICE-6 and AML-12 Cytotoxicity: The Role of CYP2E1 and Keap1-Nrf2 Pathways. J. Agric. Food Chem. 2023, 71, 1518–1530. [Google Scholar] [CrossRef]
- Korobkova, E.A. Effect of Natural Polyphenols on CYP Metabolism: Implications for Diseases. Chem. Res. Toxicol. 2015, 28, 1359–1390. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Tan, F.; Li, C.; Li, W.F.; Liao, W.; Li, Q.; Qin, G.H.; Liu, W.W.; Zhao, X. White Peony (Fermented Camellia sinensis) Polyphenols Help Prevent Alcoholic Liver Injury via Antioxidation. Antioxidants 2019, 8, 524. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Yang, C.; Ren, D.; Shao, H.; Zhao, Y.; Yang, X. Fu brick tea alleviates alcoholic liver injury by modulating the gut microbiota-liver axis and inhibiting the hepatic TLR4/NF-κB signaling pathway. Food Funct. 2022, 13, 9391–9406. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Fu, J.Q.; Li, L.; Chen, C.J.; Wang, H.H.; Hou, Y.Y.; Xu, Y.Y.; Pi, J.B. Nrf2 in alcoholic liver disease. Toxicol. Appl. Pharmacol. 2018, 357, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.M.; Wang, M.Y.; Ji, C.C.; Qi, X.; Cui, C.B. Regulation of Hazelnut Leaf Polyphenols on Oxidative Stress in Chronic Alcoholic Liver Injury. Efood 2025, 6, e70053. [Google Scholar] [CrossRef]
- Buko, V.; Zavodnik, I.; Budryn, G.; Zaklos-Szyda, M.; Belonovskaya, E.; Kirko, S.; Zyzelewicz, D.; Zakrzeska, A.; Bakunovich, A.; Rusin, V.; et al. Chlorogenic Acid Protects against Advanced Alcoholic Steatohepatitis in Rats via Modulation of Redox Homeostasis, Inflammation, and Lipogenesis. Nutrients 2021, 13, 4155. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, H.H.L.; Kim, Y.; Lee, H.B.; Lee, E.; Park, J.H.; Park, H.Y. Corchorus olitorius L. ameliorates alcoholic liver disease by regulating gut-liver axis. J. Funct. Foods. 2021, 85, 104648. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.B.; Zhang, Z.Q.; Zhang, H.Y.; Hu, H.B. Targeting NF-κB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Wang, H.H.; Gong, W.Y.; Gao, J.X.; Cheng, W.X.; Hu, Y.D.; Hu, C.Q. Effects of vitamin D deficiency on chronic alcoholic liver injury. Free Radic. Biol. Med. 2024, 224, 220–231. [Google Scholar] [CrossRef]
- Kong, E.Q.Z.; Subramaniyan, V.; Lubau, N.S.A. Uncovering the impact of alcohol on internal organs and reproductive health: Exploring TLR4/NF-kB and CYP2E1/ROS/Nrf2 pathways. Anim. Models Exp. Med. 2024, 7, 444–459. [Google Scholar] [CrossRef]
- Xu, D.W.; Xu, M.; Jeong, S.S.; Qian, Y.H.; Wu, H.L.; Xia, Q.; Kong, X.N. The Role of Nrf2 in Liver Disease: Novel Molecular Mechanisms and Therapeutic Approaches. Front. Pharmacol. 2019, 9, 1428. [Google Scholar] [CrossRef]
- Lee, D.F.; Kuo, H.P.; Liu, M.; Chou, C.K.; Xia, W.; Du, Y.; Shen, J.; Chen, C.T.; Huo, L.; Hsu, M.C.; et al. KEAP1 E3 Ligase-Mediated Downregulation of NF-κB Signaling by Targeting IKKβ. Mol. Cell. 2009, 36, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Zhao, L.; Wang, Y.; Zhang, C.; Li, H. Sugarcane Molasses Polyphenol Extract Attenuates Alcohol-Induced Chronic Liver Damage via Antioxidant, Anti-Inflammatory, and CYP2E1/Keap1/NF-κB Pathway Modulation. Nutrients 2025, 17, 1589. https://doi.org/10.3390/nu17091589
Wang M, Zhao L, Wang Y, Zhang C, Li H. Sugarcane Molasses Polyphenol Extract Attenuates Alcohol-Induced Chronic Liver Damage via Antioxidant, Anti-Inflammatory, and CYP2E1/Keap1/NF-κB Pathway Modulation. Nutrients. 2025; 17(9):1589. https://doi.org/10.3390/nu17091589
Chicago/Turabian StyleWang, Min, Lin Zhao, Yumei Wang, Chengfeng Zhang, and He Li. 2025. "Sugarcane Molasses Polyphenol Extract Attenuates Alcohol-Induced Chronic Liver Damage via Antioxidant, Anti-Inflammatory, and CYP2E1/Keap1/NF-κB Pathway Modulation" Nutrients 17, no. 9: 1589. https://doi.org/10.3390/nu17091589
APA StyleWang, M., Zhao, L., Wang, Y., Zhang, C., & Li, H. (2025). Sugarcane Molasses Polyphenol Extract Attenuates Alcohol-Induced Chronic Liver Damage via Antioxidant, Anti-Inflammatory, and CYP2E1/Keap1/NF-κB Pathway Modulation. Nutrients, 17(9), 1589. https://doi.org/10.3390/nu17091589