
Phytochemical Analysis and In Vivo Anticancer Effect of Becium grandiflorum: Isolation and Characterization of a Promising Cytotoxic Diterpene

 , and
, and
Abstract

1. Introduction
2. Materials and Methods
2.1. Material
2.2. Plant Extraction and Fractionation
2.3. GC-MS Analysis of N-Hexane Fraction
2.4. Isolation of the Major Compound from the N-Hexane Fraction
2.5. NMR and LC-ESI-HRMS Analysis of the Major Compound from the N-Hexane Fraction
2.6. Standardization of N-Hexane Fraction Using UPLC-PDA
2.7. In Vitro Studies
2.7.1. Cell Viability and Cytotoxicity Assays
Cell Culture
Cell Viability Assays
Cell Cycle Analysis
Annexin V/Propidium Iodide (PI) Apoptosis Assay
Determination of Levels of Apoptotic Proteins Using ELISA
2.8. In Vivo Experimentation
2.8.1. Experimental Design
2.8.2. Histopathological Examination
2.8.3. Statistical Analysis
2.9. In Silico Disruption of Bax-Bcl2 Interaction by Terpenoid-Induced Bcl2 Conformational Changes
2.9.1. Induced-Fit Docking
2.9.2. Data Analysis and Graphics
3. Results
3.1. GC-MS Analysis of N-Hexane Fraction of B. grandiflorum
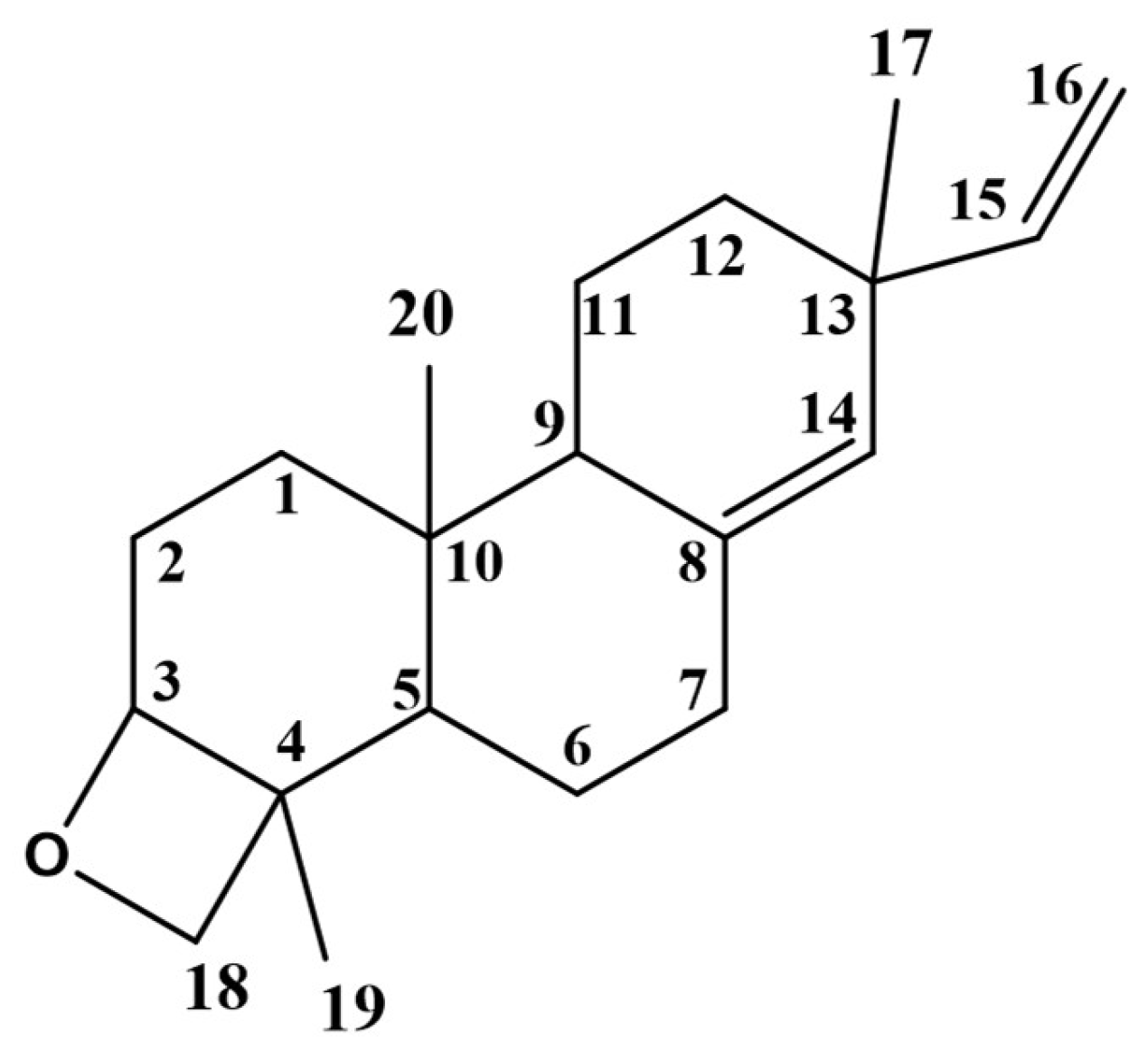
3.2. Isolation and Identification of the Major Compound from N-Hexane Fraction
3.3. UPLC-PDA Standardization of N-Hexane Fraction of B. grandiflorum
3.4. In Vitro Screening of B. grandiflorum
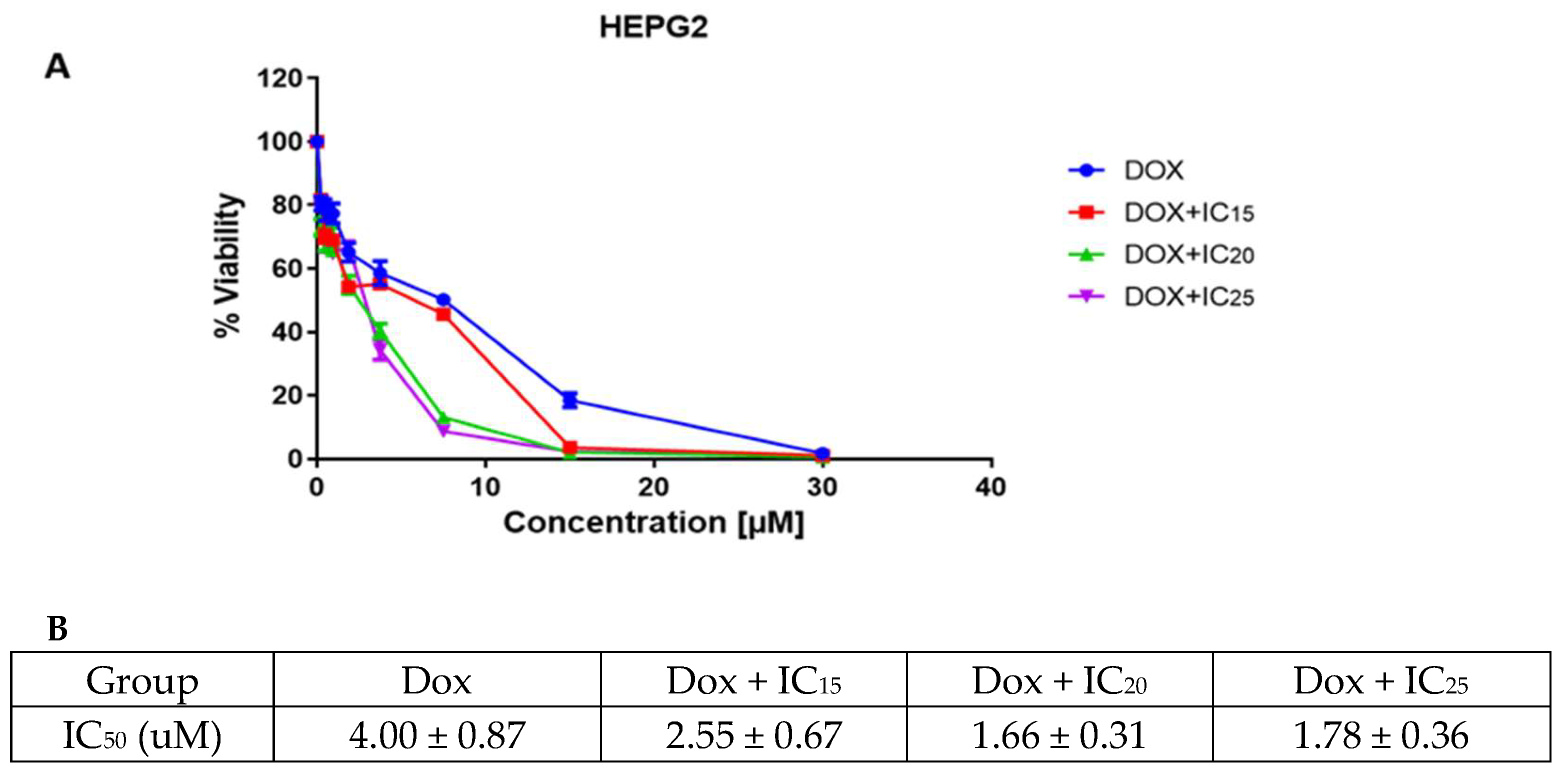
3.4.1. Cell Viability and Cytotoxicity Assessment
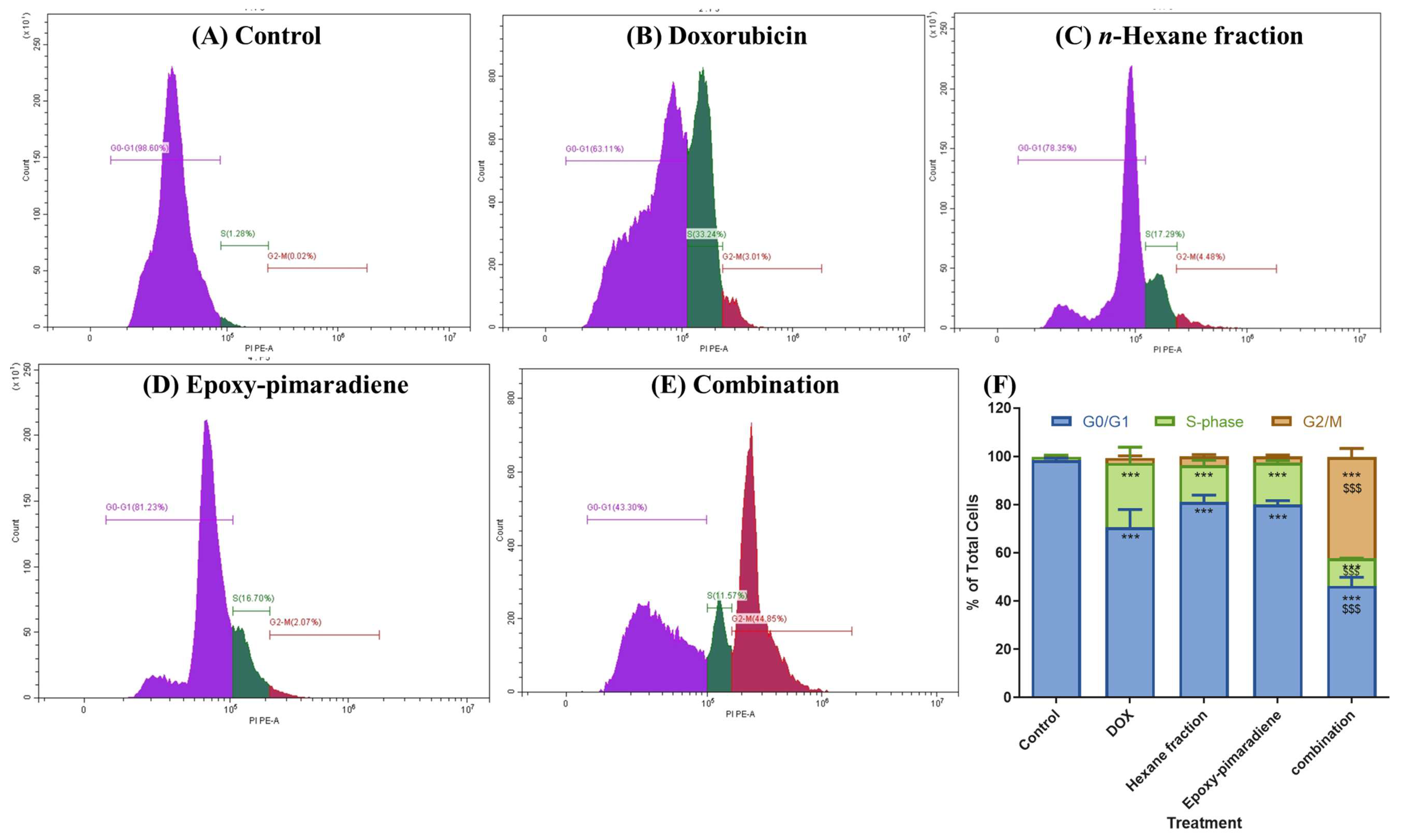
3.4.2. Results of Cell Cycle Analysis
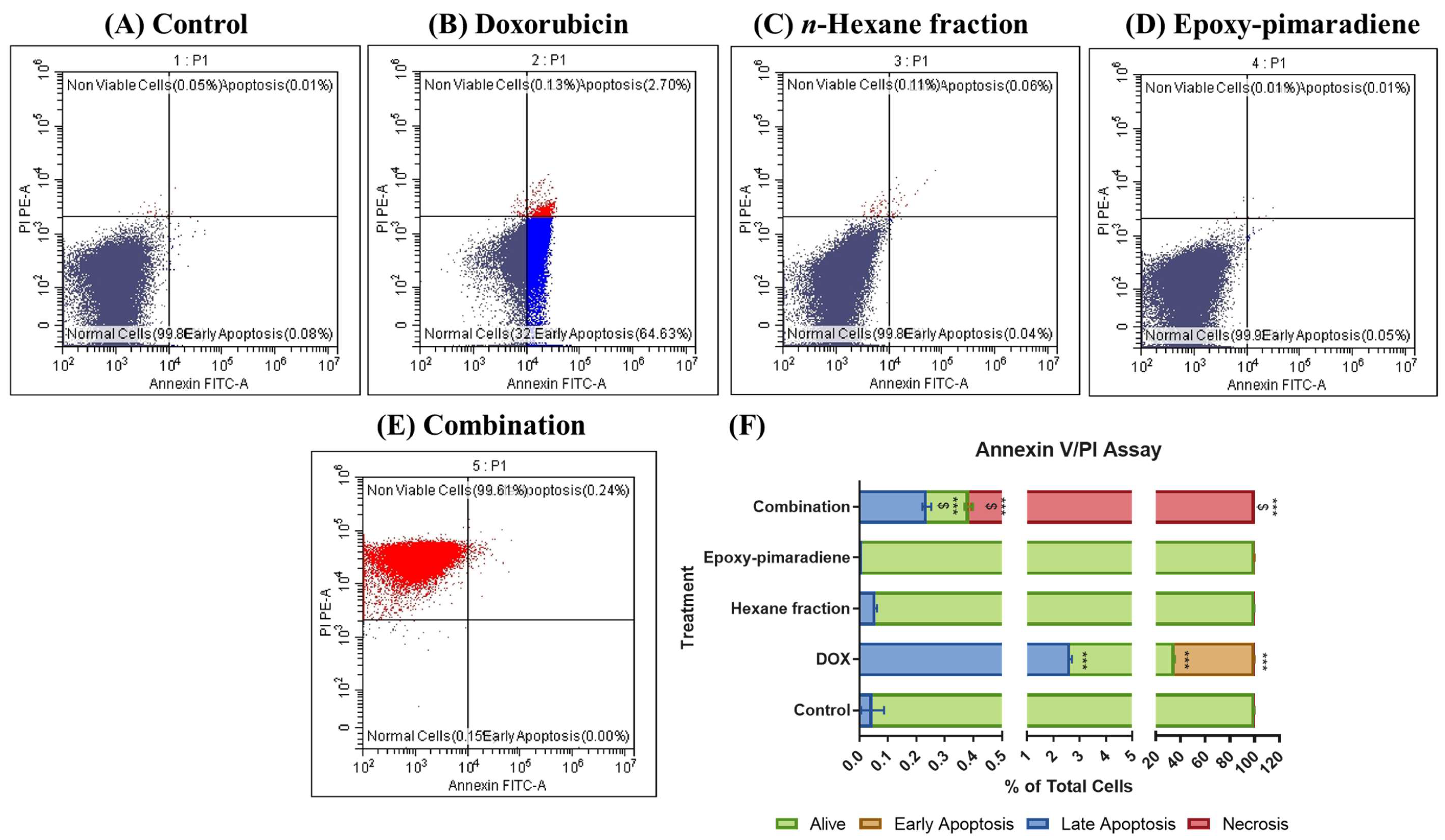
3.4.3. Results of Annexin V/PI Apoptosis Assay
3.4.4. The Effect of N-Hexane Fraction, Epoxy-Pimaradiene, Doxorubicin, and the Combination Therapy on the Key Apoptotic Protein
3.5. In Vivo Anticancer Efficacy of B. grandiflorum on Ehrlich Ascites Carcinoma
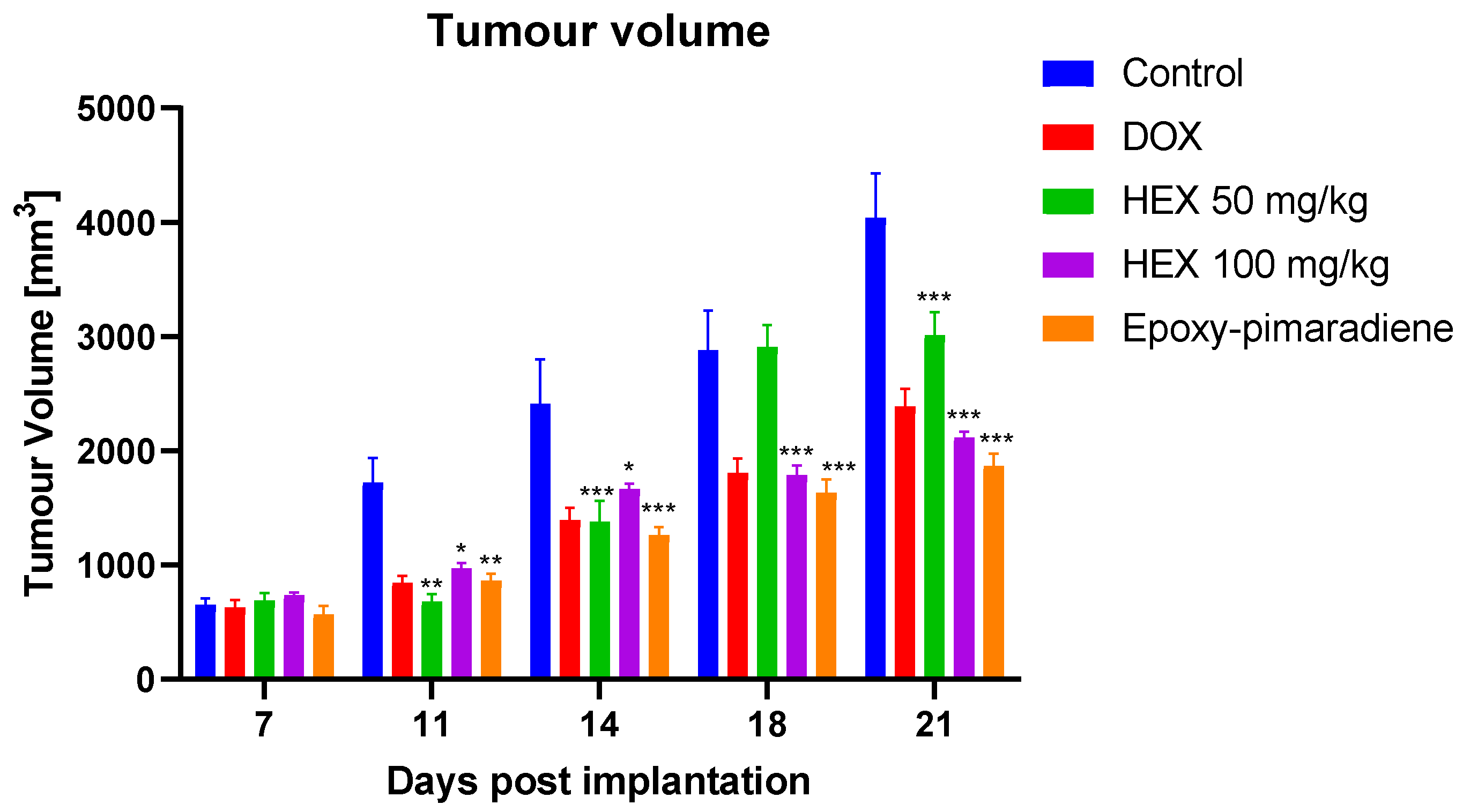
3.5.1. Effect of the Tested Samples on the Tumor Volume
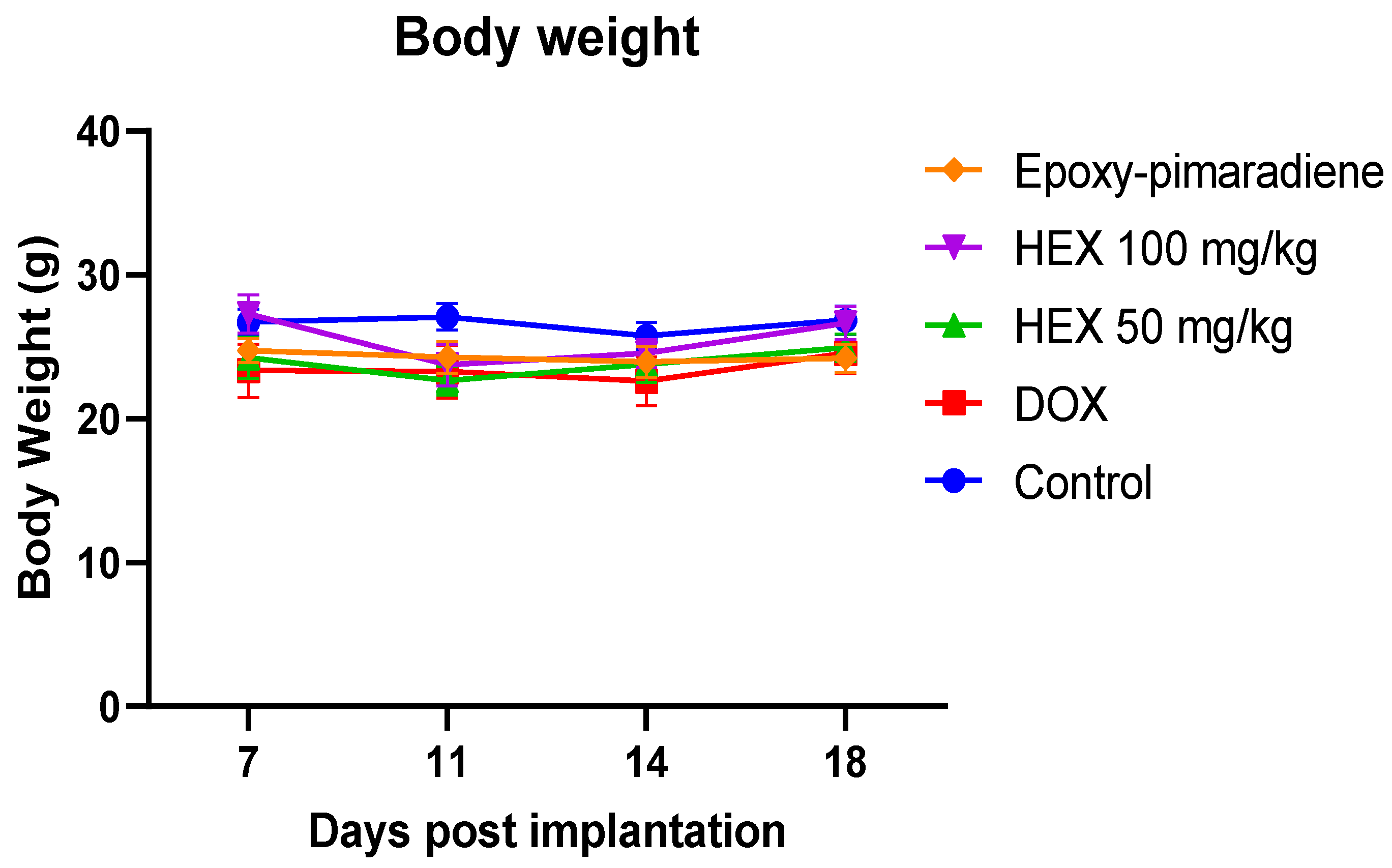
3.5.2. Body Weight of Tumor-Bearing Mice
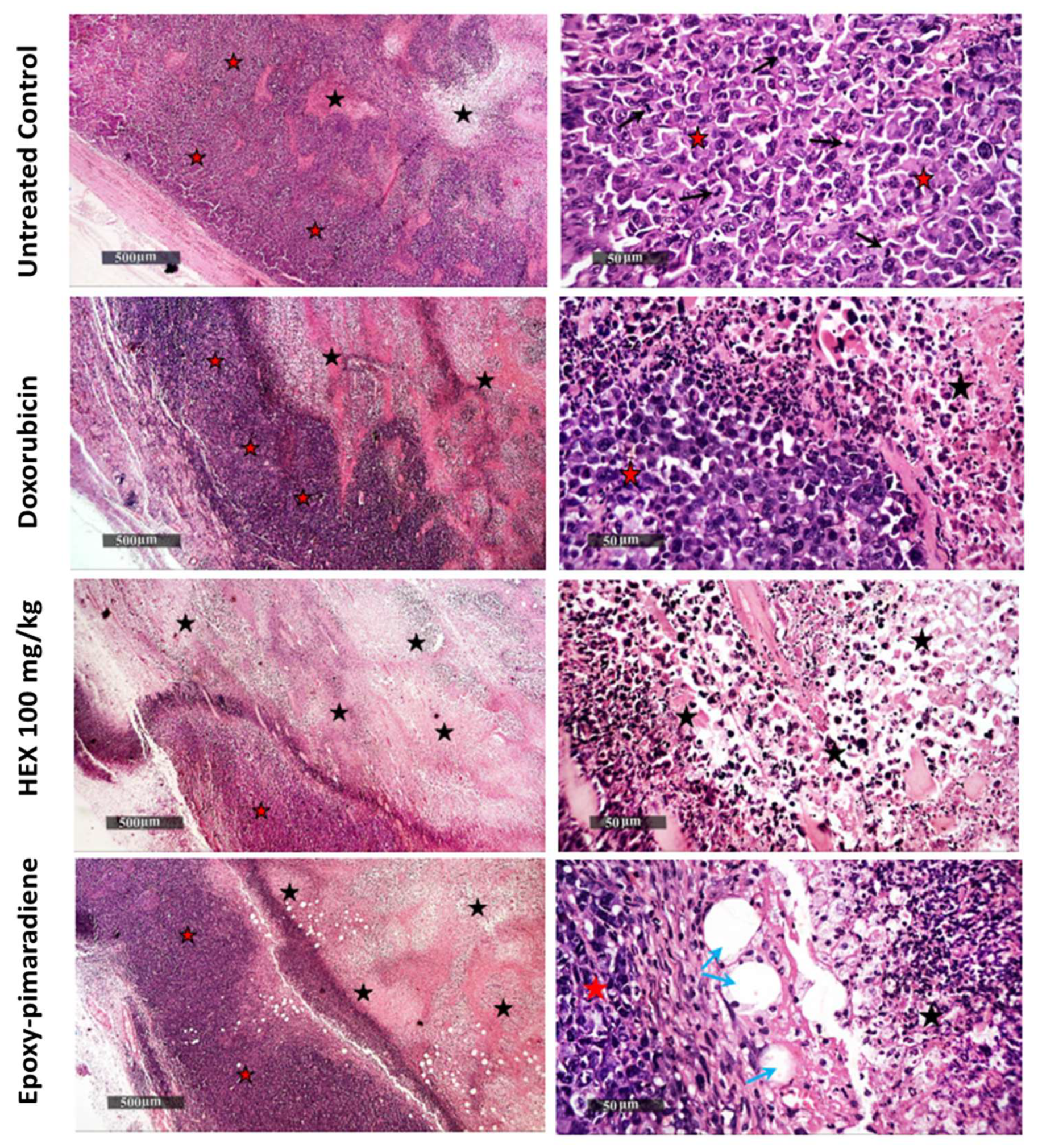
3.5.3. Tumor Histopathological Examination
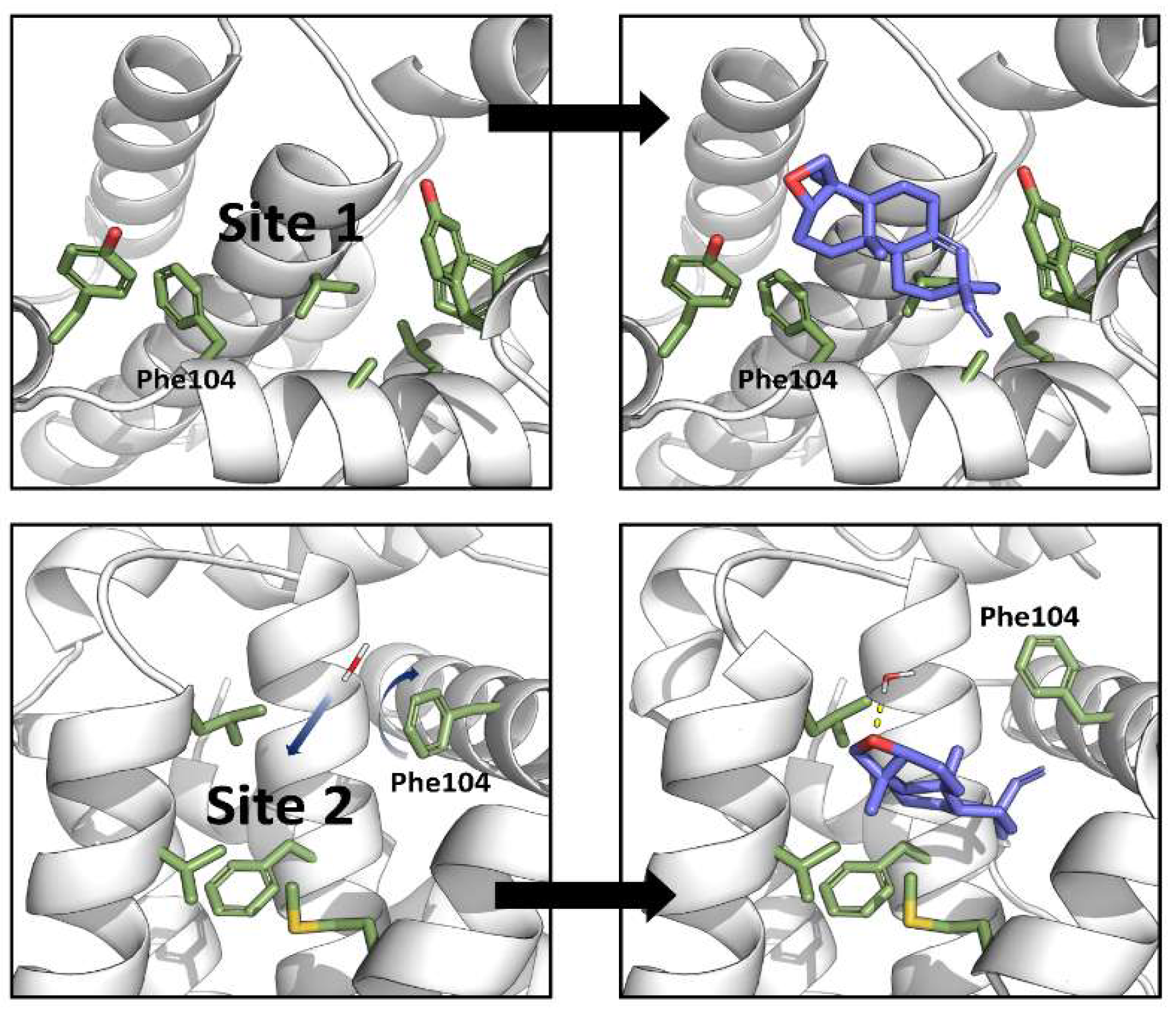
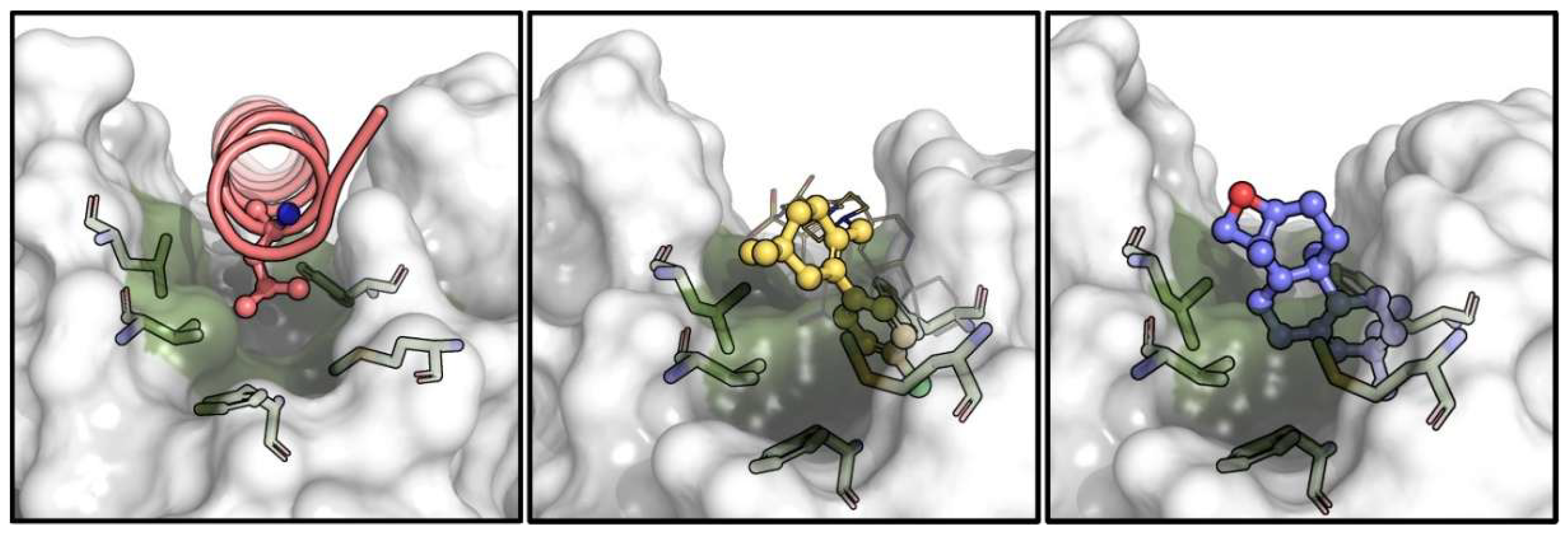
3.6. In Silico Disruption of Bax-Bcl2 Interaction by Terpenoid-Induced Bcl-2 Conformational Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HCT-116 | Human colon cancer cell line |
| MCF-7 | Human breast cancer cell line |
| MDA-MB-231 | Human triple-negative breast cancer cell line |
| HepG2 | Human liver cancer cell line |
| DMEM | Dulbecco’s Modified Eagle Medium |
| BCA | Bicinchoninic Acid |
| BCL-2 | B-Cell Lymphoma 2 |
References
- Uritu, C.M.; Mihai, C.T.; Stanciu, G.-D.; Dodi, G.; Alexa-Stratulat, T.; Luca, A.; Leon-Constantin, M.-M.; Stefanescu, R.; Bild, V.; Melnic, S. Medicinal plants of the family Lamiaceae in pain therapy: A review. Pain Res. Manag. 2018, 2018, 7801543. [Google Scholar] [CrossRef] [PubMed]
- Shakya, A.K. Medicinal plants: Future source of new drugs. Int. J. Herb. Med. 2016, 4, 59–64. [Google Scholar] [CrossRef]
- Hamed, A.N.; Attia, E.; Desoukey, S.Y. A review on various classes of secondary metabolites and biological activities of Lamiaceae (Labiatae) (2002–2018). J. Adv. Biomed. Pharm. Sci. 2021, 4, 16–31. [Google Scholar] [CrossRef]
- Raja, R.R. Medicinally potential plants of Labiatae (Lamiaceae) family: An overview. Res. J. Med. Plant 2012, 6, 203–213. [Google Scholar] [CrossRef]
- Michel, J.; Abd Rani, N.Z.; Husain, K. A review on the potential use of medicinal plants from Asteraceae and Lamiaceae plant family in cardiovascular diseases. Front. Pharmacol. 2020, 11, 852. [Google Scholar] [CrossRef]
- Sitarek, P.; Merecz-Sadowska, A.; Śliwiński, T.; Zajdel, R.; Kowalczyk, T. An in vitro evaluation of the molecular mechanisms of action of medical plants from the Lamiaceae family as effective sources of active compounds against human cancer cell lines. Cancers 2020, 12, 2957. [Google Scholar] [CrossRef]
- Berdowska, I.; Zieliński, B.; Matusiewicz, M.; Fecka, I. Modulatory impact of Lamiaceae metabolites on apoptosis of human leukemia cells. Front. Pharmacol. 2022, 13, 867709. [Google Scholar] [CrossRef]
- Hiltunen, R.; Holm, Y. Basil: The genus Ocimum; CRC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
- Gebremeskel, L.; Tuem, K.B.; Teklu, T. Evaluation of antidiabetic effect of ethanolic leaves extract of Becium grandiflorum Lam. (Lamiaceae) in streptozotocin-induced diabetic mice. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 1481. [Google Scholar] [CrossRef]
- Burger, I.; Burger, B.V.; Albrecht, C.F.; Spies, H.S.; Sándor, P. Triterpenoid saponins from Becium grandiflorum var. obovatum. Phytochemistry 1998, 49, 2087–2095. [Google Scholar] [CrossRef]
- Grayer, R.J.; Veitch, N.C. An 8-hydroxylated external flavone and its 8-O-glucoside from Becium grandiflorum. Phytochemistry 1998, 47, 779–782. [Google Scholar] [CrossRef]
- Beshir, K.; Shibeshi, W.; Ejigu, A.; Engidawork, E. In-vivo wound healing activity of 70% ethanol leaf extract of Becium grandiflorum Lam. (Lamiaceae) in mice. Ethiop. Pharm. J. 2016, 32, 117–130. [Google Scholar] [CrossRef]
- Kahsay, M.H. Synthesis and characterization of ZnO nanoparticles using aqueous extract of Becium grandiflorum for antimicrobial activity and adsorption of methylene blue. Appl. Water Sci. 2021, 11, 45. [Google Scholar]
- Reveglia, P.; Cimmino, A.; Masi, M.; Nocera, P.; Berova, N.; Ellestad, G.; Evidente, A. Pimarane diterpenes: Natural source, stereochemical configuration, and biological activity. Chirality 2018, 30, 1115–1134. [Google Scholar] [CrossRef] [PubMed]
- Elhawary, E.A.; Mostafa, N.M.; Labib, R.M.; Singab, A.N.J.P. Metabolomic profiles of essential oils from selected rosa varieties and their antimicrobial activities. Plants 2021, 10, 1721. [Google Scholar] [CrossRef]
- Ibrahim, N.; Abbas, H.; El-Sayed, N.S.; Gad, H.A. Rosmarinus officinalis L. hexane extract: Phytochemical analysis, nanoencapsulation, and in silico, in vitro, and in vivo anti-photoaging potential evaluation. Sci. Rep. 2022, 12, 13102. [Google Scholar] [CrossRef]
- Swilam, N.; Nematallah, K.A. Polyphenols profile of pomegranate leaves and their role in green synthesis of silver nanoparticles. Sci. Rep. 2020, 10, 14851. [Google Scholar] [CrossRef]
- Mahmoud, M.; Abdollah, M.R.; Elsesy, M.E.; Abou El Ella, D.A.; Zada, S.K.; Tolba, M.F. The natural isoflavone Biochanin-A synergizes 5-fluorouracil anticancer activity in vitro and in vivo in Ehrlich solid-phase carcinoma model. Phytother. Res. 2022, 36, 1310–1325. [Google Scholar] [CrossRef]
- Abdollah, M.R.; Ali, A.A.; Elgohary, H.H.; Elmazar, M.M. Antiangiogenic drugs in combination with seaweed fucoidan: A mechanistic in vitro and in vivo study exploring the VEGF receptor and its downstream signaling molecules in hepatic cancer. Front. Pharmacol. 2023, 14, 1108992. [Google Scholar] [CrossRef]
- Ozaslan, M.; Karagoz, I.D.; Kilic, I.H.; Guldur, M.E. Ehrlich ascites carcinoma. Afr. J. Biotechnol. 2011, 10, 2375–2378. [Google Scholar]
- Culling, C.F.A. Handbook of Histopathological and Histochemical Techniques: Including Museum Techniques; Butterworth-Heinemann: Oxford, UK, 2013. [Google Scholar]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Birkinshaw, R.W.; Gong, J.-n.; Luo, C.S.; Lio, D.; White, C.A.; Anderson, M.A.; Blombery, P.; Lessene, G.; Majewski, I.J.; Thijssen, R. Structures of BCL-2 in complex with venetoclax reveal the molecular basis of resistance mutations. Nat. Commun. 2019, 10, 2385. [Google Scholar] [CrossRef] [PubMed]
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput.-Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Sherman, W.; Day, T.; Jacobson, M.P.; Friesner, R.A.; Farid, R. Novel procedure for modeling ligand/receptor induced fit effects. J. Med. Chem. 2006, 49, 534–553. [Google Scholar] [CrossRef] [PubMed]
- Roozbehani, N.; Golfeshan, F.; Pakshir, K.; Doorandishan, M.; Jassbi, A.R.; Mosaddad, S.A. Chemical composition and effectiveness of Ocimum basilicum L. extracts on the adhesion of Candida albicans and C. dubliniensis on acrylic surfaces of removable orthodontic appliances. Biointerface Res. Appl. Chem. 2021, 11, 9477–9489. [Google Scholar] [CrossRef]
- Mustafa, A.A.; El-Kamali, H.H. Chemical Composition of Ocimum americanum In Sudan. Res. Pharm. Health Sci. 2019, 5, 172–178. [Google Scholar] [CrossRef]
- Bromann, K.; Viljanen, K.; Moreira, V.; Yli-Kauhaluoma, J.; Ruohonen, L.; Nakari-Setälä, T. Isolation and purification of ent-pimara-8 (14), 15-diene from engineered Aspergillus nidulans by accelerated solvent extraction combined with HPLC. Anal. Methods 2014, 6, 1227–1234. [Google Scholar] [CrossRef]
- Sun, H.-D.; Huang, S.-X.; Han, Q.-B. Diterpenoids from Isodon species and their biological activities. Nat. Prod. Rep. 2006, 23, 673–698. [Google Scholar] [CrossRef]
- Khodapasand, E.; Jafarzadeh, N.; Farrokhi, F.; Kamalidehghan, B.; Houshmand, M. Is Bax/Bcl-2 ratio considered as a prognostic marker with age and tumor location in colorectal cancer? Iran. Biomed. J. 2015, 19, 69–75. [Google Scholar] [CrossRef]
- Andreu-Fernández, V.; Sancho, M.; Genovés, A.; Lucendo, E.; Todt, F.; Lauterwasser, J.; Funk, K.; Jahreis, G.; Pérez-Payá, E.; Mingarro, I. Bax transmembrane domain interacts with prosurvival Bcl-2 proteins in biological membranes. Proc. Natl. Acad. Sci. USA 2017, 114, 310–315. [Google Scholar] [CrossRef]
- Ku, B.; Liang, C.; Jung, J.U.; Oh, B.-H. Evidence that inhibition of BAX activation by BCL-2 involves its tight and preferential interaction with the BH3 domain of BAX. Cell Res. 2011, 21, 627–641. [Google Scholar] [CrossRef]
- Hart, P.C.; Rajab, I.M.; Alebraheem, M.; Potempa, L.A. C-Reactive protein and cancer—Diagnostic and therapeutic insights. Front. Immunol. 2020, 11, 595835. [Google Scholar] [CrossRef] [PubMed]
- Kenchegowda, M.; Rahamathulla, M.; Hani, U.; Begum, M.Y.; Guruswamy, S.; Osmani, R.A.M.; Gowrav, M.P.; Alshehri, S.; Ghoneim, M.M.; Alshlowi, A. Smart nanocarriers as an emerging platform for cancer therapy: A review. Molecules 2021, 27, 146. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, H.; Zhang, Y.; Lu, X.; Wang, B.; Liu, X. Bioactive pimarane-type diterpenes from marine organisms. Chem. Biodivers. 2018, 15, e1700276. [Google Scholar] [CrossRef]
- Ye, K.; Ai, H.-l. Pimarane Diterpenes from Fungi. Pharmaceuticals 2022, 15, 1291. [Google Scholar] [CrossRef]
- Kongwaen, P.; Boonsombat, J.; Thongnest, S.; Ruchisansakun, S.; Mahidol, C.; Ruchirawat, S. Cytotoxic Isopimarane Diterpenoids from Kaempferia koratensis Rhizomes. Rev. Bras. De Farmacogn. 2023, 33, 415–421. [Google Scholar] [CrossRef]
- de Oliveira, A.d.S.S.; Conrado, G.G.; Grazzia, N.; Miguel, D.C.; Júnior, G.C.F.; Garcia, V.L. In Vitro Cytotoxic and Leishmanicidal Activity of Isolated and Semisynthetic ent-Pimaranes from Aldama arenaria. Planta Medica 2022, 88, 1163–1174. [Google Scholar] [CrossRef]
- Gelain, D.P.; Pasquali, M.A.d.B.; Caregnato, F.F.; Castro, M.A.A.; Moreira, J.C.F. Retinol induces morphological alterations and proliferative focus formation through free radical-mediated activation of multiple signaling pathways. Acta Pharmacol. Sin. 2012, 33, 558–567. [Google Scholar] [CrossRef]
- Garattini, E.; Bolis, M.; Garattini, S.K.; Fratelli, M.; Centritto, F.; Paroni, G.; Zanetti, A.; Pagani, A.; Fisher, J.N.; Zambelli, A. Retinoids and breast cancer: From basic studies to the clinic and back again. Cancer Treat. Rev. 2014, 40, 739–749. [Google Scholar] [CrossRef]
- Blowman, K.; Magalhães, M.; Lemos, M.; Cabral, C.; Pires, I. Anticancer properties of essential oils and other natural products. Evid.-Based Complement. Altern. Med. 2018, 2018, 3149362. [Google Scholar] [CrossRef]
- Andrade, M.A.; Braga, M.A.; Cesar, P.H.; Trento, M.V.C.; Espósito, M.A.; Silva, L.F.; Marcussi, S. Anticancer properties of essential oils: An overview. Curr. Cancer Drug Targets 2018, 18, 957–966. [Google Scholar] [CrossRef]
- Angelini, P.; Tirillini, B.; Akhtar, M.S.; Dimitriu, L.; Bricchi, E.; Bertuzzi, G.; Venanzoni, R. Essential oil with anticancer activity: An overview. Anticancer Plants Nat. Prod. Biotechnol. Implements 2018, 2, 207–231. [Google Scholar] [CrossRef]
- Israels, E.D.; Israels, L.G. The cell cycle. Oncologist 2000, 5, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Matthews, H.K.; Bertoli, C.; de Bruin, R.A. Cell cycle control in cancer. Nat. Rev. Mol. Cell Biol. 2022, 23, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.J.; Reddy, E.P. CDK4: A key player in the cell cycle, development, and cancer. Genes Cancer 2012, 3, 658–669. [Google Scholar] [CrossRef]
- Massagué, J. G1 cell-cycle control and cancer. Nature 2004, 432, 298–306. [Google Scholar] [CrossRef]
- Lindqvist, A.; van Zon, W.; Karlsson Rosenthal, C.; Wolthuis, R.M.F. Cyclin B1–Cdk1 activation continues after centrosome separation to control mitotic progression. PLoS Biol. 2007, 5, e123. [Google Scholar] [CrossRef]
- Salanci, Š.; Vilková, M.; Martinez, L.; Mirossay, L.; Michalková, R.; Mojžiš, J. The induction of G2/M phase cell cycle arrest and apoptosis by the chalcone derivative 1C in sensitive and resistant ovarian cancer cells is associated with ROS generation. Int. J. Mol. Sci. 2024, 25, 7541. [Google Scholar] [CrossRef]
- Johnson, J.J.; Syed, D.N.; Suh, Y.; Heren, C.R.; Saleem, M.; Siddiqui, I.A.; Mukhtar, H. Disruption of androgen and estrogen receptor activity in prostate cancer by a novel dietary diterpene carnosol: Implications for chemoprevention. Cancer Prev. Res. 2010, 3, 1112–1123. [Google Scholar] [CrossRef]
- Yin, R.; Li, T.; Tian, J.X.; Xi, P.; Liu, R.H. Ursolic acid, a potential anticancer compound for breast cancer therapy. Crit. Rev. Food Sci. Nutr. 2018, 58, 568–574. [Google Scholar] [CrossRef]
- Hasan, M.R.; Alotaibi, B.S.; Althafar, Z.M.; Mujamammi, A.H.; Jameela, J. An Update on the Therapeutic Anticancer Potential of Ocimum sanctum L.: “Elixir of Life”. Molecules 2023, 28, 1193. [Google Scholar] [CrossRef]
- Wang, H.; Ye, Y.; Chu, J.-H.; Zhu, G.-Y.; Li, Y.-W.; Fong, D.W.; Yu, Z.-L. Oridonin induces G2/M cell cycle arrest and apoptosis through MAPK and p53 signaling pathways in HepG2 cells. Oncol. Rep. 2010, 24, 647–651. [Google Scholar]
- Li, J.; Wu, Y.; Wang, D.; Zou, L.; Fu, C.; Zhang, J.; Leung, G.P.-H. Oridonin synergistically enhances the anti-tumor efficacy of doxorubicin against aggressive breast cancer via pro-apoptotic and anti-angiogenic effects. Pharmacol. Res. 2019, 146, 104313. [Google Scholar] [CrossRef] [PubMed]
- Wihadmadyatami, H.; Karnati, S.; Hening, P.; Tjahjono, Y.; Maharjanti, F.; Kusindarta, D.L.; Triyono, T. Ethanolic extract Ocimum sanctum Linn. induces an apoptosis in human lung adenocarcinoma (A549) cells. Heliyon 2019, 5, e02772. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, X.; Wei, L.; Pan, Y.; Huang, S.; Wang, H.; Begonia, G.B.; Ekunwe, S.I. Antioxidant properties and chemical constituents of ethanolic extract and its fractions of Ocimum gratissimum. Med. Chem. Res. 2013, 22, 1124–1130. [Google Scholar] [CrossRef]
- Strzyz, P. Pulling the apoptotic trigger for necrosis. Nat. Rev. Mol. Cell Biol. 2017, 18, 72. [Google Scholar] [CrossRef]
- Chan, F.K.-M.; Moriwaki, K.; De Rosa, M.J. Detection of necrosis by release of lactate dehydrogenase activity. In Immune Homeostasis: Methods and Protocols; Springer: New York, NY, USA, 2013; pp. 65–70. [Google Scholar]
- Van Noorden, C.J.F. The history of Z-VAD-FMK, a tool for understanding the significance of caspase inhibition. Acta Histochem. 2001, 103, 241–251. [Google Scholar]
- Dhandayuthapani, S.; Azad, H.; Rathinavelu, A. Apoptosis induction by Ocimum sanctum extract in LNCaP prostate cancer cells. J. Med. Food 2015, 18, 776–785. [Google Scholar] [CrossRef]
- Chen, H.-M.; Lee, M.-J.; Kuo, C.-Y.; Tsai, P.-L.; Liu, J.-Y.; Kao, S.-H. Ocimum gratissimum aqueous extract induces apoptotic signalling in lung adenocarcinoma cell A549. Evid.-Based Complement. Altern. Med. 2010, 2011, 739093. [Google Scholar] [CrossRef]
- Ferreira, P.M.P.; Militão, G.C.G.; Lima, D.J.B.; de Jesus Costa, N.D.; da Conceição Machado, K.; Dos Santos, A.G.; Cavalheiro, A.J.; da Silva Bolzani, V.; Silva, D.H.S.; Pessoa, C. Morphological and biochemical alterations activated by antitumor clerodane diterpenes. Chem.-Biol. Interact. 2014, 222, 112–125. [Google Scholar] [CrossRef]
- Al-Lihaibi, S.S.; Alarif, W.M.; Abdel-Lateff, A.; Ayyad, S.-E.N.; Abdel-Naim, A.B.; El-Senduny, F.F.; Badria, F.A. Three new cembranoid-type diterpenes from Red Sea soft coral Sarcophyton glaucum: Isolation and antiproliferative activity against HepG2 cells. Eur. J. Med. Chem. 2014, 81, 314–322. [Google Scholar] [CrossRef]
- de Abrantes, R.A.; Batista, T.M.; Mangueira, V.M.; de Sousa, T.K.; Ferreira, R.C.; Moura, A.P.G.; Abreu, L.S.; Alves, A.F.; Velozo, E.S.; Batista, L.M. Antitumor and antiangiogenic effects of Tonantzitlolone B, an uncommon diterpene from Stillingia loranthacea. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2022, 395, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Shubin, A.V.; Demidyuk, I.V.; Komissarov, A.A.; Rafieva, L.M.; Kostrov, S.V. Cytoplasmic vacuolization in cell death and survival. Oncotarget 2016, 7, 55863. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, B.; Masi, M.; Maddau, L.; De Lorenzi, M.; Dam, R.; Cimmino, A.; y Banuls, L.M.; Andolfi, A.; Kiss, R.; Mathieu, V. Evaluation of in vitro anticancer activity of sphaeropsidins A–C, fungal rearranged pimarane diterpenes, and semisynthetic derivatives. Phytochem. Lett. 2012, 5, 770–775. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak # | Component | Molecular Formula | Molecular Weight | RT (Min) | Relative Percentile (%) | KI | |
|---|---|---|---|---|---|---|---|
| Obsd. | Lit. | ||||||
| 1 | Hexahydrofarnesyl acetone | C18H36O | 268 | 31.686 | 0.23 | 1841 | 1842 |
| 2 | 1-Eicosene | C20H40 | 280 | 33.288 | 0.36 | 1923 | 1986 |
| 3 | Ethyl palmitate | C18H36O2 | 284 | 34.74 | 0.86 | 1995 | 1995 |
| 4 | Ethyl linolenate | C20H34O2 | 306 | 38.122 | 0.61 | 2173 | 2171 |
| 5 | Unidentified | - | - | 38.475 | 0.42 | 2192 | - |
| 6 | 3- α-Hydroxy-manool | C20H34O2 | 306 | 39.35 | 10.92 | 2241 | 2273 |
| 7 | 4-Epidehydroabietol | C20H30O | 286 | 39.625 | 0.34 | 2257 | 2326 |
| 8 | Pimara-8(14),15-dien-3-ol | C20H32O | 288 | 39.945 | 14.02 | 2275 | 2253 |
| 9 | ent-Kaurenol | C20H32O | 288 | 40.240 | 3.83 | 2291 | 2302 |
| 10 | Isopimarol | C20H32O | 288 | 40.805 | 3.51 | 2324 | 2305 |
| 11 | Verticillol | C20H34O | 290 | 41.050 | 0.82 | 2338 | 2106 |
| 12 | Unidentified | - | - | 41.145 | 0.18 | 2344 | - |
| 13 | Retinol | C20H30O | 286 | 42.370 | 5.36 | 2417 | 2453 |
| 14 | Kauren-19-oic acid | C20H30O2 | 302 | 42.815 | 0.53 | 2444 | 2358 |
| 15 | Unidentified | - | - | 43.030 | 1.51 | 2457 | - |
| 16 | Retinol | C20H30O | 286 | 43.195 | 1.03 | 2467 | 2453 |
| 17 | Unidentified | - | - | 43.590 | 0.47 | 2491 | - |
| 18 | Communic acid | C20H30O2 | 302 | 43.785 | 0.75 | 2503 | 2404 |
| 19 | Unidentified | - | - | 44.205 | 0.7 | 2530 | - |
| 20 | Sandaracopimaric acid | C20H30O2 | 302 | 44.455 | 1.73 | 2546 | 2282 |
| 21 | 18-epoxy-pimara- 8(14),15-diene | C20H30O | 286 | 44.665 | 27.90 | 2559 | - |
| 22 | Pimara-8(14),15-dien-3-one | C20H30O | 286 | 45.265 | 6.14 | 2597 | 2279 |
| 23 | Unidentified | - | - | 45.710 | 0.57 | 2626 | - |
| 24 | Unidentified | - | - | 46.995 | 0.29 | 2712 | - |
| 25 | Unidentified | - | - | 47.155 | 0.57 | 2722 | - |
| 26 | Unidentified | - | - | 47.365 | 0.45 | 2737 | - |
| 27 | Sesamin | C20H18O6 | 354 | 53.605 | 3.94 | 3193 | 3150 |
| 28 | Unidentified | - | - | 54.520 | 2.52 | 3258 | - |
| 29 | Unidentified | - | - | 54.965 | 0.43 | 3288 | - |
| 30 | 2-methyloctacosane | C29H60 | 408 | 55.130 | 1.59 | 3300 | 2866 |
| 31 | Unidentified | - | - | 55.380 | 0.7 | 3315 | - |
| 32 | δ7,25-Stigmastadienol | C29H48O | 412 | 55.755 | 2.8 | 3338 | 3325 |
| 33 | Unidentified | - | - | 55.900 | 1.69 | 3346 | - |
| 34 | Gamma.-Sitosterol | C29H50O | 414 | 56.045 | 0.4 | 3355 | 3351 |
| 35 | Unidentified | - | - | 56.395 | 1.18 | 3376 | - |
| 36 | Unidentified | - | - | 56.555 | 0.65 | 3386 | - |
| Total identified % | 87.67 | ||||||
| Number of identified components | 21 | ||||||
| Hydrocarbons % | 1.95 | ||||||
| Sesquiterpenes % | 0.23 | ||||||
| Oxygenated diterpenes % | 70.49 | ||||||
| Fatty acids % | 1.47 | ||||||
| Sterols % | 3.2 | ||||||
| Lignans % | 3.94 | ||||||
| Others % | 6.39 | ||||||
| C/H Position | δH, Multi. | δC, Type | COSY | HMBC |
|---|---|---|---|---|
| 1 | 1.72, m (1H) 1.14, m (1H) | 36.9, CH2 | ||
| 2 | 1.62–1.72 m (2H) | 27.25 CH2 | ||
| 3 | 3.7 m (1H) | 77.07 CH | ||
| 4 | - | 42.1 C | C-18 | |
| 5 | 1.14, m (1H) | 48.6 CH | ||
| 6 | 1.40–1.45, m (2H) | 22.4 CH2 | ||
| 7 a | 2.26, m (1H) | 35.7 CH2 | ||
| 7 β | 2.05, m (1H) | |||
| 8 | - | 136.9 C | ||
| 9 | 1.75, m (1H) | 50.5 CH | ||
| 10 | - | 37.7 C | ||
| 11 | 1.57–1.52 m (2H) | 18.7 CH2 | ||
| 12 | 1.50–1.44 m (2H) | 34.4 CH2 | ||
| 13 | - | 37.4 C | ||
| 14 | 5.25, s (1H) | 129.04 CH | C-15-C-17 | |
| 15 | 5.78 dd (1 H) J1 10.6 Hz and J2 17.5 Hz | 148.8 CH | H-16 | C-16 |
| 16 a | 4.96, dd (1H) J1= 1.5 Hz and J2 = 17.5 Hz | 110.15 CH2 | H-15 | |
| 16 b | 4.87, dd (1H) J1 = 1.5 Hz and J2 = 10.6 Hz | H-15 | ||
| 17 | 1.05, s (3H) | 25.97 CH3 | ||
| 18 | 3.44, d (1H) 3.7, d (1H) | 72.01 CH2 | H-3 | C-3, C-19 |
| 19 | 0.94, s (3H) | 11.54 CH3 | C-18 | |
| 20 | 0.89, s (3H) | 15.50 CH3 |
| Validation Parameter | Result |
|---|---|
| Linearity (R2) | 0.9998 |
| Limit of detection (LOD) (µg ml−1) | 10.46 |
| Limit of quantification (LOQ) (µg ml−1) | 31.70 |
| Recovery % (mean± SD) | 100.58 ± 1.64 |
| Precision (RSD%) Intra-day Inter-day | 0.07 0.08 |
| Cell Line | IC50 (µg/mL) | ||
|---|---|---|---|
| Hydroalcoholic Extract | N-Hexane Fraction | Epoxy-Pimaradiene | |
| MCF-7 | 43.54 ± 9.19 | 20.29 ± 2.16 | 23.06 ± 2.02 |
| MDA-MB-231 | - | 89.10 ± 10.05 | 123.20 ± 16.21 |
| HCT-116 | 40.51 ± 5.25 | 15.54 ± 1.64 | 10.61 ± 0.61 |
| HepG2 | 19.85 ± 2.34 | 13.75 ± 1.05 | 11.45 ±1.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahim, C.; Abdollah, M.R.A.; Labib, R.M.; Ibrahim, N.; Swilam, N. Phytochemical Analysis and In Vivo Anticancer Effect of Becium grandiflorum: Isolation and Characterization of a Promising Cytotoxic Diterpene. Nutrients 2025, 17, 1164. https://doi.org/10.3390/nu17071164
Fahim C, Abdollah MRA, Labib RM, Ibrahim N, Swilam N. Phytochemical Analysis and In Vivo Anticancer Effect of Becium grandiflorum: Isolation and Characterization of a Promising Cytotoxic Diterpene. Nutrients. 2025; 17(7):1164. https://doi.org/10.3390/nu17071164
Chicago/Turabian StyleFahim, Christeen, Maha R. A. Abdollah, Rola M. Labib, Nehal Ibrahim, and Noha Swilam. 2025. "Phytochemical Analysis and In Vivo Anticancer Effect of Becium grandiflorum: Isolation and Characterization of a Promising Cytotoxic Diterpene" Nutrients 17, no. 7: 1164. https://doi.org/10.3390/nu17071164
APA StyleFahim, C., Abdollah, M. R. A., Labib, R. M., Ibrahim, N., & Swilam, N. (2025). Phytochemical Analysis and In Vivo Anticancer Effect of Becium grandiflorum: Isolation and Characterization of a Promising Cytotoxic Diterpene. Nutrients, 17(7), 1164. https://doi.org/10.3390/nu17071164