
1,8-Cineole Alleviates Hippocampal Oxidative Stress in CUMS Mice via the PI3K/Akt/Nrf2 Pathway
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals
2.3. CUMS Depression Model
2.4. Sucrose Preference Test (SPT) and Open-Field Test (OFT)
2.5. Forced Swimming Test (FST)
2.6. HE Staining
2.7. Dihydroethyl Chloride (DHE) Assay
2.8. Network Pharmacology
2.9. Molecular Docking
2.10. MDA and SOD Assays
2.11. Western Blot
2.12. Cell Modeling and Treatment
2.13. Cell Viability Assay
2.14. Small Interfering RNA (siRNA) Silencing
2.15. Statistical Analysis
3. Results
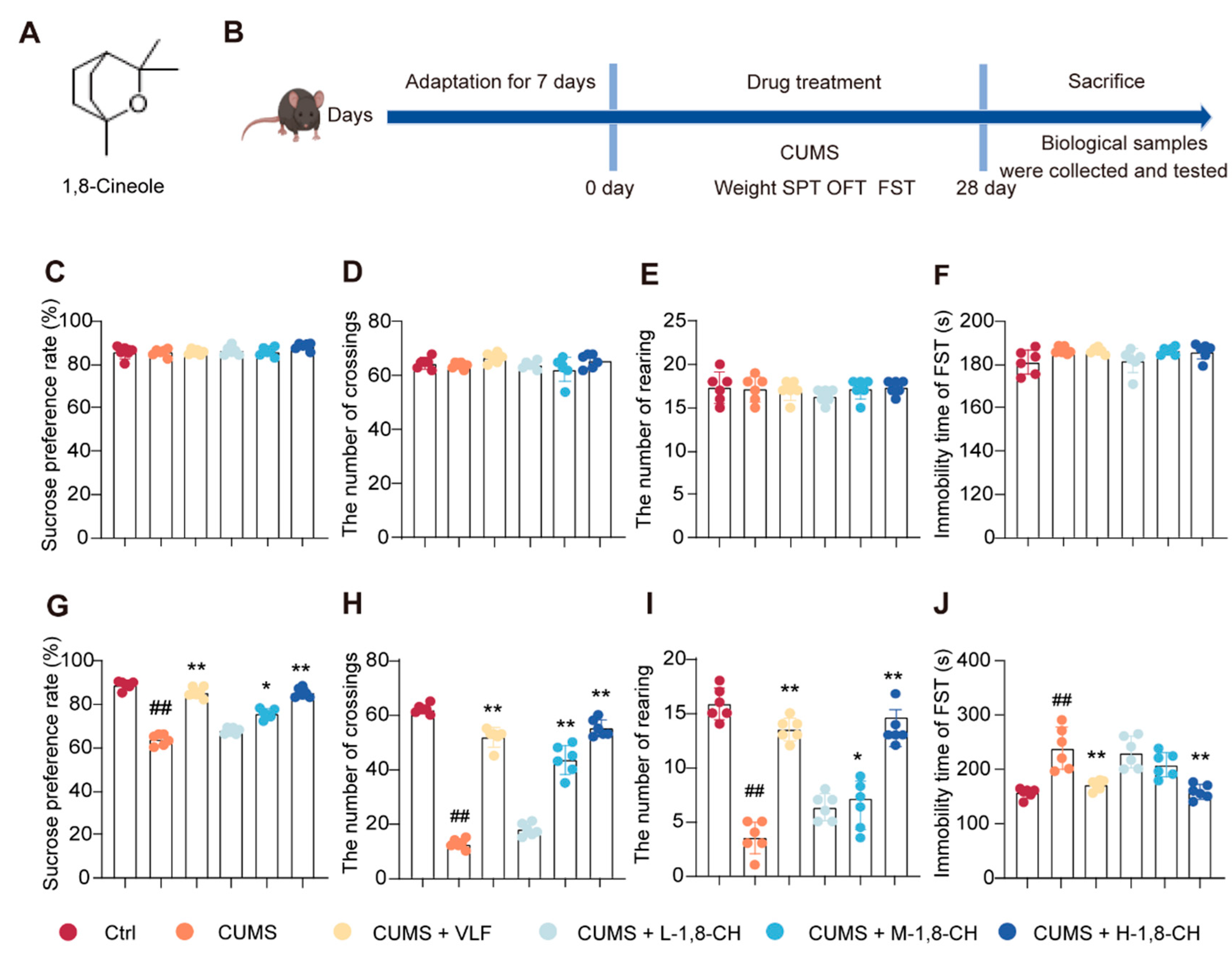
3.1. 1,8-CH Significantly Mitigates Depressive-like Behaviors in CUMS-Induced Mice
3.2. 1,8-CH Alleviates Hippocampal Tissue Damage and Oxidative Stress in CUMS Mice
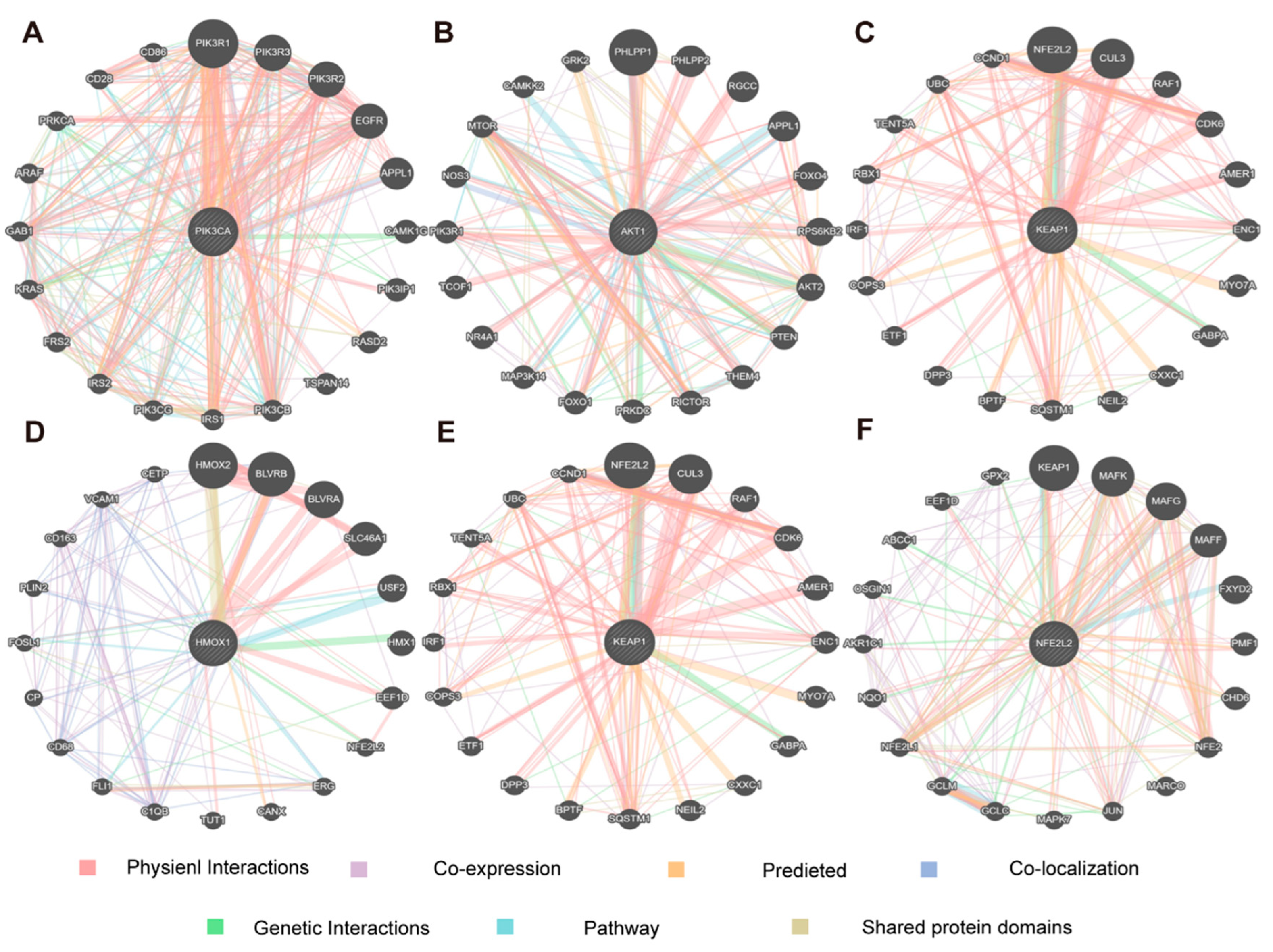
3.3. Network Pharmacology Analysis of 1,8-CH Antidepressant
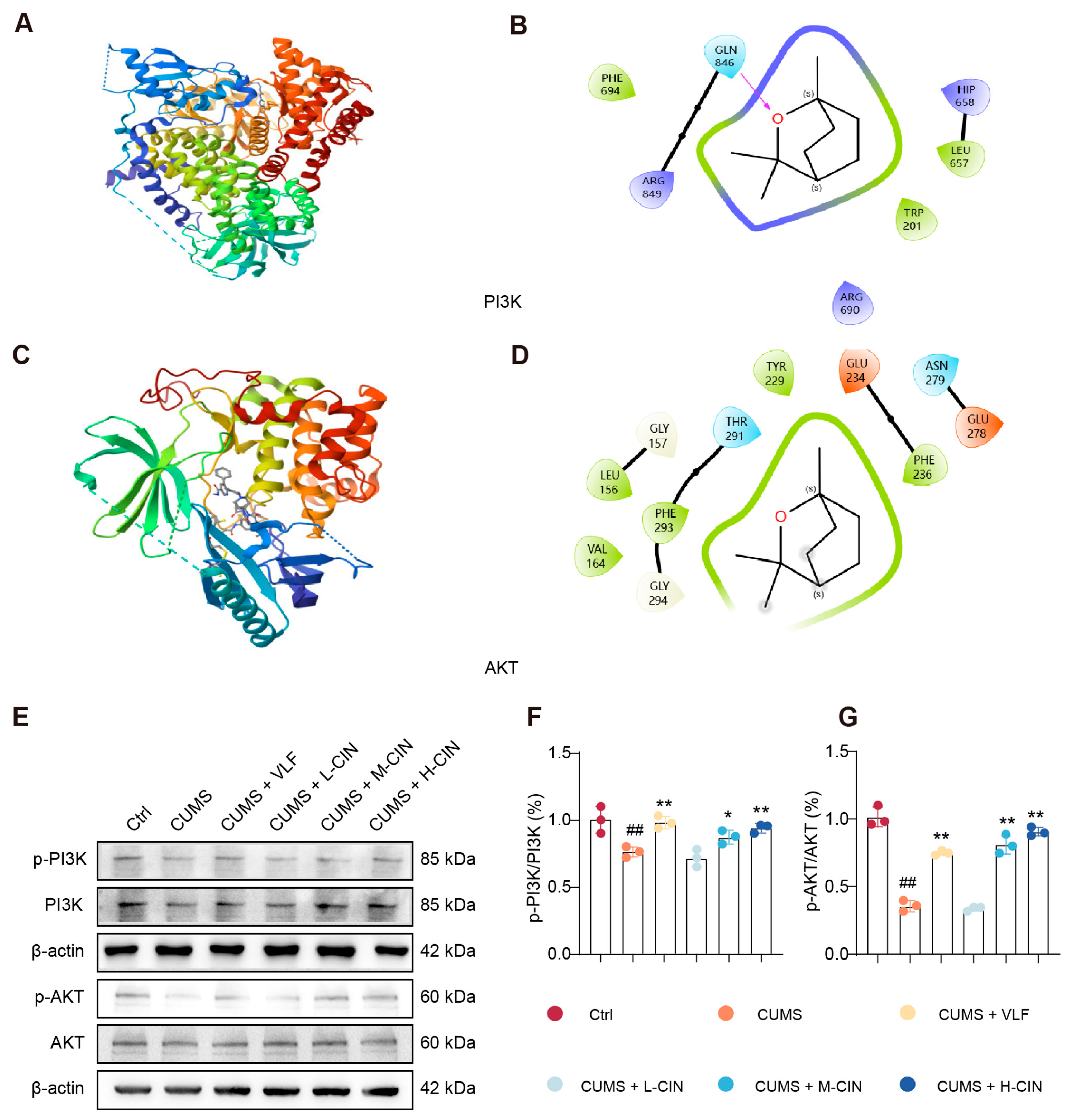
3.4. 1,8-CH Activates the PI3K-Akt Signaling Pathway in the Hippocampus of CUMS-Induced Mice
3.5. 1,8-CH Activates the Nrf2/Keap1/HO-1 Signaling Pathway in the Hippocampus of CUMS-Induced Mice
3.6. Inhibition and Knockdown of Nrf2 Abolishes the Antioxidant Effects of 1,8-CH in CORT-Induced PC12 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sorlí, J.V.; de la Cámara, E.; Fernández-Carrión, R.; Asensio, E.M.; Portolés, O.; Ortega-Azorín, C.; Pérez-Fidalgo, A.; Villamil, L.V.; Fitó, M.; Barragán, R.; et al. Depression and Accelerated Aging: The Eveningness Chronotype and Low Adherence to the Mediterranean Diet Are Associated with Depressive Symptoms in Older Subjects. Nutrients 2024, 17, 104. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Ji, C.; Zheng, J.; Zhou, S.; Tian, J.; Han, Y.; Qin, X. Effects of Xiaoyao San on exercise capacity and liver mitochondrial metabolomics in rat depression model. Chin. Herb. Med. 2024, 16, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Smith, J. Daily briefing: Gentle brain stimulation could treat depression at home. Nature 2024. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhao, Y.; Qin, X.; Tian, J. Deciphering relationship between depression and microbial molecules based on multi-omics: A case study of Chaigui Granules. Chin. Herb. Med. 2024, 16, 612–621. [Google Scholar] [CrossRef]
- Poulsen, N.B.; Lambert, M.N.T.; Jeppesen, P.B. The Effect of Plant Derived Bioactive Compounds on Inflammation: A Systematic Review and Meta-Analysis. Mol. Nutr. Food Res. 2020, 64, e2000473. [Google Scholar] [CrossRef]
- Park, J.; Lee, H.J. Specific Foods Associated with Depressive Symptoms among Young Adults and Their Bioactive Effects. Nutrients 2024, 16, 1818. [Google Scholar] [CrossRef]
- Xia, C.X.; Gao, A.X.; Zhu, Y.; Dong, T.T.; Tsim, K.W. Flavonoids from Seabuckthorn (Hippophae rhamnoides L.) restore CUMS-induced depressive disorder and regulate the gut microbiota in mice. Food Funct. 2023, 14, 7426–7438. [Google Scholar] [CrossRef]
- Okereke, O.I.; Vyas, C.M.; Mischoulon, D.; Chang, G.; Cook, N.R.; Weinberg, A.; Bubes, V.; Copeland, T.; Friedenberg, G.; Lee, I.M.; et al. Effect of Long-term Supplementation with Marine Omega-3 Fatty Acids vs Placebo on Risk of Depression or Clinically Relevant Depressive Symptoms and on Change in Mood Scores: A Randomized Clinical Trial. JAMA 2021, 326, 2385–2394. [Google Scholar] [CrossRef]
- Zheng, K.Y.; Gao, B.; Wang, H.J.; He, J.G.; Chen, H.S.; Hu, Z.L.; Long, L.H.; Chen, J.G.; Wang, F. Melatonin Ameliorates Depressive-Like Behaviors in Ovariectomized Mice by Improving Tryptophan Metabolism via Inhibition of Gut Microbe Alistipes Inops. Adv. Sci. 2024, 11, e2309473. [Google Scholar] [CrossRef]
- Noor, A.A.M. Exploring the Therapeutic Potential of Terpenoids for Depression and Anxiety. Chem. Biodivers. 2024, 21, e202400788. [Google Scholar] [CrossRef]
- Agatonovic-Kustrin, S.; Kustrin, E.; Gegechkori, V.; Morton, D.W. Anxiolytic Terpenoids and Aromatherapy for Anxiety and Depression. Adv. Exp. Med. Biol. 2020, 1260, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.A.; Rao, V.S.N. Possible role of mast cells in cineole-induced scratching behavior in mice. Food Chem. Toxicol. 2002, 40, 1453–1457. [Google Scholar] [CrossRef] [PubMed]
- Kako, H.; Fukumoto, Y.; Kobayashi, Y.; Yokogoshi, H. Effects of direct exposure of green odour components on dopamine release from rat brain striatal slices and PC12 cells. Brain Res. Bull. 2008, 75, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Yasin, M.; Younis, A.; Javed, T.; Akram, A.; Ahsan, M.; Shabbir, R.; Ali, M.M.; Tahir, A.; El-Ballat, E.M.; Sheteiwy, M.S.; et al. River Tea Tree Oil: Composition, Antimicrobial and Antioxidant Activities, and Potential Applications in Agriculture. Plants 2021, 10, 2105. [Google Scholar] [CrossRef]
- Sobhy, M.; Ali, S.S.; Cui, H.; Lin, L.; El-Sapagh, S. Exploring the potential of 1,8-cineole from cardamom oil against food-borne pathogens: Antibacterial mechanisms and its application in meat preservation. Microb. Pathog. 2023, 184, 106375. [Google Scholar] [CrossRef]
- Bezerra, D.S.; Delmondes, G.D.; Lopes, M.J.P.; Araujo, I.M.; Leite, G.M.D.; Barbosa, M.D.; Barbosa, R.; Alves, A.F.; Medeiros, C.R.; Florencio, S.G.L.; et al. Eucalyptol prevents pilocarpine-induced seizure and neuronal damage in mice, through the cholinergic, monoaminergic and antioxidant pathways. Food Biosci. 2023, 53, 102824. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Wang, J.; Jin, X.F.; Gao, P.; Zhao, Y.M.; Yin, M.J.; Ma, X.; Xin, Z.Y.; Zhao, Y.M.; Zhou, X.H.; et al. 1,8-Cineole Alleviates OGD/R-Induced Oxidative Damage and Restores Mitochondrial Function by Promoting the Nrf2 Pathway. Biol. Pharm. Bull. 2023, 46, 1371–1384. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Fu, Y.; Fu, D.; Zhen, D.; Xing, A.; Chen, Y.; Gong, G.; Wei, C. 1, 8-cineole protects against ISO-induced heart failure by inhibiting oxidative stress and ER stress in vitro and in vivo. Eur. J. Pharmacol. 2021, 910, 174472. [Google Scholar] [CrossRef]
- Bahrami, T.; Yaghmaei, P.; Yousofvand, N. The effects of Ibuprofen and 1, 8- cineol on anxiety and spatial memory in hyperammonemic rats. Metab. Brain Dis. 2023, 38, 613–620. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Zhang, R.; Gao, J.; Wu, M.; Wang, J.; Sheng, J.; Sun, P. The Potential Mechanism of Alpiniae oxyphyllae Fructus Against Hyperuricemia: An Integration of Network Pharmacology, Molecular Docking, Molecular Dynamics Simulation, and In Vitro Experiments. Nutrients 2024, 17, 71. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, X.; Liu, Q.; Wang, Y. Network pharmacology, molecular docking, and untargeted metabolomics reveal molecular mechanisms of multi-targets effects of Qingfei Tongluo Plaster improving respiratory syncytial virus pneumonia. Chin. Herb. Med. 2024, 16, 638–655. [Google Scholar] [CrossRef] [PubMed]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology 1987, 93, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Hei, X.; Xie, M.; Xu, J.; Li, J.; Liu, T. β-Asarone Exerts Antioxidative Effects on H(2)O(2)-Stimulated PC12 Cells by Activating Nrf2/HO-1 Pathway. Neurochem. Res. 2020, 45, 1953–1961. [Google Scholar] [CrossRef]
- Xu, L.; Anwyl, R.; Rowan, M.J. Behavioural stress facilitates the induction of long-term depression in the hippocampus. Nature 1997, 387, 497–500. [Google Scholar] [CrossRef]
- Xia, W.; Xing, W.; Jin, P.; Zhang, C.; Fu, W.; Wu, H.; Zhu, W.; Xu, D. Odorant for Relieving Post-Traumatic Stress Disorder-Like Symptom such as Relieving Anxiety and Depression, Enhancing Autonomous Movement, Exploration Ability, and Improving Short-Term Learning and Memory Ability, Comprises Linalyl Acetate and 1,8-Cineole. CN118697726-A. Available online: https://webofscience.clarivate.cn/wos/alldb/summary/8af96498-4c09-4ca0-aded-548d5e3684cf-0151f96a5e/relevance/1 (accessed on 12 March 2025).
- Porres-Martínez, M.; González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. In vitro neuroprotective potential of the monoterpenes α-pinene and 1,8-cineole against H2O2-induced oxidative stress in PC12 cells. Z. Naturforschung Sect. C J. Biosci. 2016, 71, 191–199. [Google Scholar] [CrossRef]
- Sun, R.; Liang, Y.; Zhu, S.; Yin, Q.; Bian, Y.; Ma, H.; Zhao, F.; Yin, G.; Tang, D. Homotherapy-for-heteropathy of Bupleurum Chinense DC.-Scutellaria baicalensis Georgi in treating depression and colorectal cancer: A network pharmacology and animal model approach. J. Ethnopharmacol. 2024, 328, 118038. [Google Scholar] [CrossRef]
- Si, L.; Xiao, L.; Xie, Y.; Xu, H.; Yuan, G.; Xu, W.; Wang, G. Social isolation after chronic unpredictable mild stress perpetuates depressive-like behaviors, memory deficits and social withdrawal via inhibiting ERK/KEAP1/NRF2 signaling. J. Affect. Disord. 2023, 324, 576–588. [Google Scholar] [CrossRef]
- Li, Z.; Liang, S.; Cui, X.; Shen, C.; Xu, Z.; Chen, W.; Wu, M.; Liang, C.; Liu, J.; Huang, J.; et al. Network pharmacology- and molecular docking-based investigation on the mechanism of action of Si-ni San in the treatment of depression combined with anxiety and experimental verification in adolescent rats. Front. Psychiatry 2024, 15, 1414242. [Google Scholar] [CrossRef]
- Jenniskens, J.C.A.; Offermans, K.; Simons, C.; Samarska, I.; Fazzi, G.E.; van der Meer, J.R.M.; Smits, K.M.; Schouten, L.J.; Weijenberg, M.P.; Grabsch, H.I.; et al. Energy balance-related factors in childhood and adolescence and risk of colorectal cancer based on KRAS, PIK3CA, and BRAF mutations and MMR status. Mol. Carcinog. 2022, 61, 1099–1115. [Google Scholar] [CrossRef]
- Ghareghomi, S.; Moosavi-Movahedi, F.; Saso, L.; Habibi-Rezaei, M.; Khatibi, A.; Hong, J.; Moosavi-Movahedi, A.A. Modulation of Nrf2/HO-1 by Natural Compounds in Lung Cancer. Antioxidants 2023, 12, 735. [Google Scholar] [CrossRef]
- Liu, Y.; Mi, Y.; Wang, Y.; Meng, Q.; Xu, L.; Liu, Y.; Zhou, D.; Wang, Y.; Liang, D.; Li, W.; et al. Loureirin C inhibits ferroptosis after cerebral ischemia reperfusion through regulation of the Nrf2 pathway in mice. Phytomedicine 2023, 113, 154729. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.O.; Sala-Newby, G.B.; Ladak, S.; Angelini, G.D.; Caputo, M.; Suleiman, M.S.; Evans, P.C.; George, S.J.; Zakkar, M. Nrf2-Keap-1 imbalance under acute shear stress induces inflammatory response in venous endothelial cells. Perfusion 2022, 37, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Chang, W.; Qi, J.; Chen, G.; Li, N. Lung protective effects of dietary malate esters derivatives from Bletilla striata against SiO(2) nanoparticles through activation of Nrf2 pathway. Chin. Herb. Med. 2023, 15, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.S.; Wu, W.Z.; Liu, S.B.; Ling-Hu, T.; Zhao, Y.H.; Gao, Y.; Qin, X.M. Stable Isotope-Resolved Metabolomics Studies on Corticosteroid-Induced PC12 Cells: A Strategy for Evaluating Glucose Catabolism in an in Vitro Model of Depression. J. Proteome Res. 2022, 21, 788–797. [Google Scholar] [CrossRef]
- Wang, P.; Yi, T.; Mao, S.; Li, M. Neuroprotective mechanism of human umbilical cord mesenchymal stem cell-derived extracellular vesicles improving the phenotype polarization of microglia via the PI3K/AKT/Nrf2 pathway in vascular dementia. Synapse 2023, 77, e22268. [Google Scholar] [CrossRef]
- Xia, C.X.; Gao, A.X.; Dong, T.T.; Tsim, K.W. Flavonoids from Seabuckthorn (Hippophae rhamnoides L.) mimic neurotrophic functions in inducing neurite outgrowth in cultured neurons: Signaling via PI3K/Akt and ERK pathways. Phytomedicine 2023, 115, 154832. [Google Scholar] [CrossRef]
- Sun, J.Y.; Liu, Y.T.; Jiang, S.N.; Guo, P.M.; Wu, X.Y.; Yu, J. Essential oil from the roots of Paeonia lactiflora pall. has protective effect against corticosterone-induced depression in mice via modulation of PI3K/Akt signaling pathway. Front. Pharmacol. 2022, 13, 999712. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, Y.; Yang, Z.; Liu, M.; Zhang, C.; Zhao, Y.; Song, C. ω-3 DPA Protected Neurons from Neuroinflammation by Balancing Microglia M1/M2 Polarizations through Inhibiting NF-κB/MAPK p38 Signaling and Activating Neuron-BDNF-PI3K/AKT Pathways. Mar. Drugs 2021, 19, 587. [Google Scholar] [CrossRef]
- Zuo, C.; Cao, H.; Song, Y.; Gu, Z.; Huang, Y.; Yang, Y.; Miao, J.; Zhu, L.; Chen, J.; Jiang, Y.; et al. Nrf2: An all-rounder in depression. Redox Biol. 2022, 58, 102522. [Google Scholar] [CrossRef]
- Li, H.; Xue, X.; Li, L.; Li, Y.; Wang, Y.; Huang, T.; Wang, Y.; Meng, H.; Pan, B.; Niu, Q. Aluminum-Induced Synaptic Plasticity Impairment via PI3K-Akt-mTOR Signaling Pathway. Neurotox. Res. 2020, 37, 996–1008. [Google Scholar] [CrossRef]
- Cigliano, L.; Spagnuolo, M.S.; Boscaino, F.; Ferrandino, I.; Monaco, A.; Capriello, T.; Cocca, E.; Iannotta, L.; Treppiccione, L.; Luongo, D.; et al. Dietary Supplementation with Fish Oil or Conjugated Linoleic Acid Relieves Depression Markers in Mice by Modulation of the Nrf2 Pathway. Mol. Nutr. Food Res. 2019, 63, e1900243. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, L.; Zhu, T.; Xu, S.; He, J.; Mao, N.; Liu, Z.; Wang, D. Neuroprotective effects of Lycium barbarum polysaccharide on light-induced oxidative stress and mitochondrial damage via the Nrf2/HO-1 pathway in mouse hippocampal neurons. Int. J. Biol. Macromol. 2023, 251, 126315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.C.; Yao, W.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Wu, J.; Ushida, Y.; Suganuma, H.; et al. Prophylactic effects of sulforaphane on depression-like behavior and dendritic changes in mice after inflammation. J. Nutr. Biochem. 2017, 39, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Ma, C.; Zhu, C.; Wang, Y.; Zou, X.; Li, H.; Li, J.; He, Y.; Wu, C. Prediction models for depression risk among older adults: Systematic review and critical appraisal. Ageing Res. Rev. 2023, 83, 101803. [Google Scholar] [CrossRef]
- Akhmouch, A.A.; Hriouech, S.; Chefchaou, H.; Tanghort, M.; Mzabi, A.; Chami, N.; Remmal, A. The Combination of Amoxicillin and 1,8-Cineole Improves the Bioavailability and the Therapeutic Effect of Amoxicillin in a Rabbit Model. Antibiotics 2022, 11, 1294. [Google Scholar] [CrossRef]
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: A review. J. Sci. Food Agric. 2018, 98, 833–848. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Wang, D.; Shi, Y.; Wang, Y.; Wu, Y.; Wu, X.; Shah, B.A.; Ye, G. 1,8-Cineole Alleviates Hippocampal Oxidative Stress in CUMS Mice via the PI3K/Akt/Nrf2 Pathway. Nutrients 2025, 17, 1027. https://doi.org/10.3390/nu17061027
Wu W, Wang D, Shi Y, Wang Y, Wu Y, Wu X, Shah BA, Ye G. 1,8-Cineole Alleviates Hippocampal Oxidative Stress in CUMS Mice via the PI3K/Akt/Nrf2 Pathway. Nutrients. 2025; 17(6):1027. https://doi.org/10.3390/nu17061027
Chicago/Turabian StyleWu, Wenze, Dequan Wang, Yuzhu Shi, Yichen Wang, Yongzi Wu, Xinyan Wu, Basit Ali Shah, and Gang Ye. 2025. "1,8-Cineole Alleviates Hippocampal Oxidative Stress in CUMS Mice via the PI3K/Akt/Nrf2 Pathway" Nutrients 17, no. 6: 1027. https://doi.org/10.3390/nu17061027
APA StyleWu, W., Wang, D., Shi, Y., Wang, Y., Wu, Y., Wu, X., Shah, B. A., & Ye, G. (2025). 1,8-Cineole Alleviates Hippocampal Oxidative Stress in CUMS Mice via the PI3K/Akt/Nrf2 Pathway. Nutrients, 17(6), 1027. https://doi.org/10.3390/nu17061027