
Enhanced Antioxidant and Protective Effects of Fermented Solanum melongena L. Peel Extracts Against Ultraviolet B-Induced Skin Damage

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Fermented Solanum melongena L. (Eggplant) Peel Extracts
2.2. Cell Culture
2.3. Measurement of Cytotoxicity
2.4. Wound-Healing Analysis
2.5. Assessment of Intracellular ROS
2.6. Membrane Potential of Mitochondria
2.7. Western Blot Analysis
2.8. In Vivo Antioxidant Effect of BEPs
2.9. Histological Analysis
2.10. Statistical Analysis
3. Result
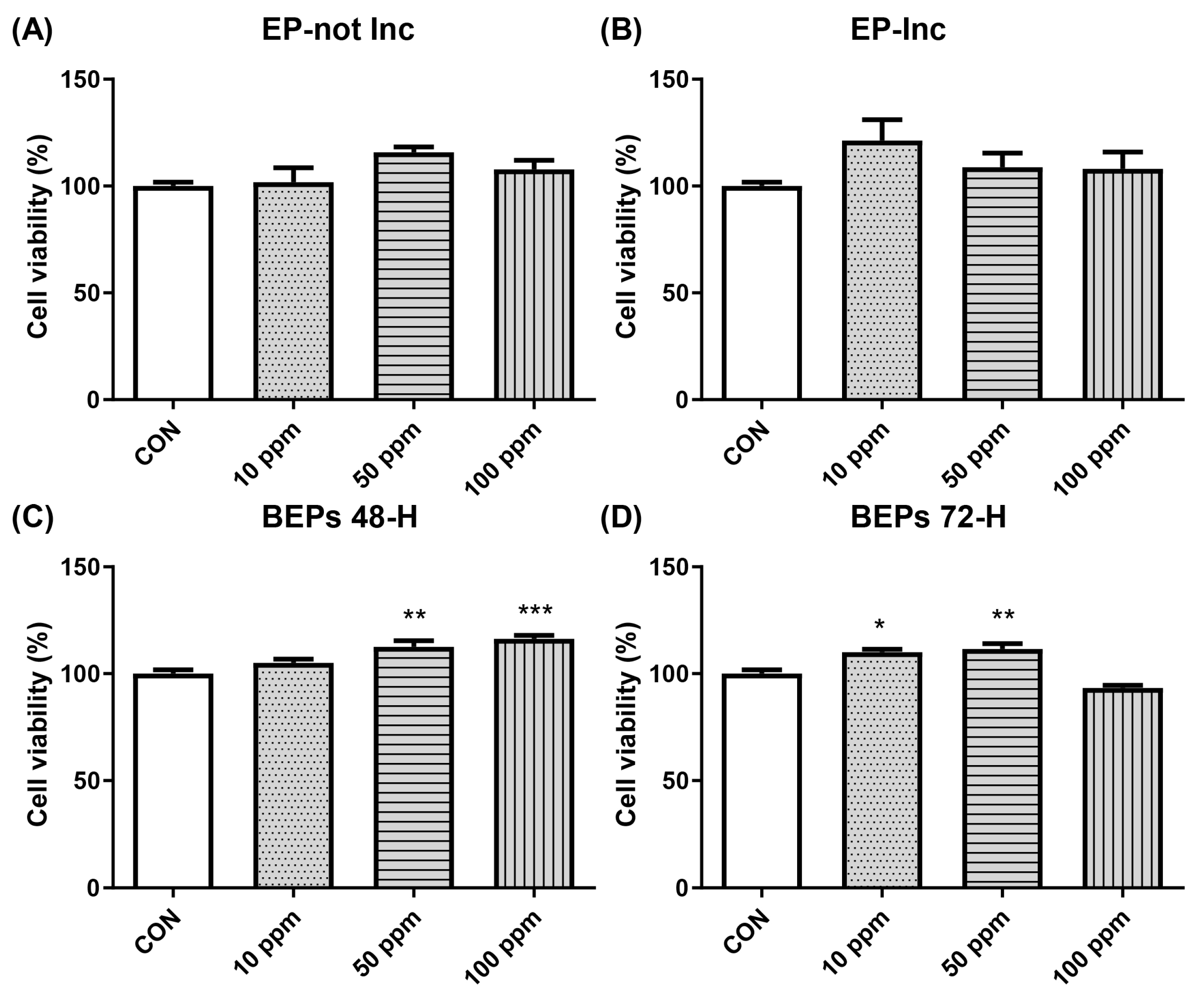
3.1. Effect of EPs and BEPs on the Viability of HDFs
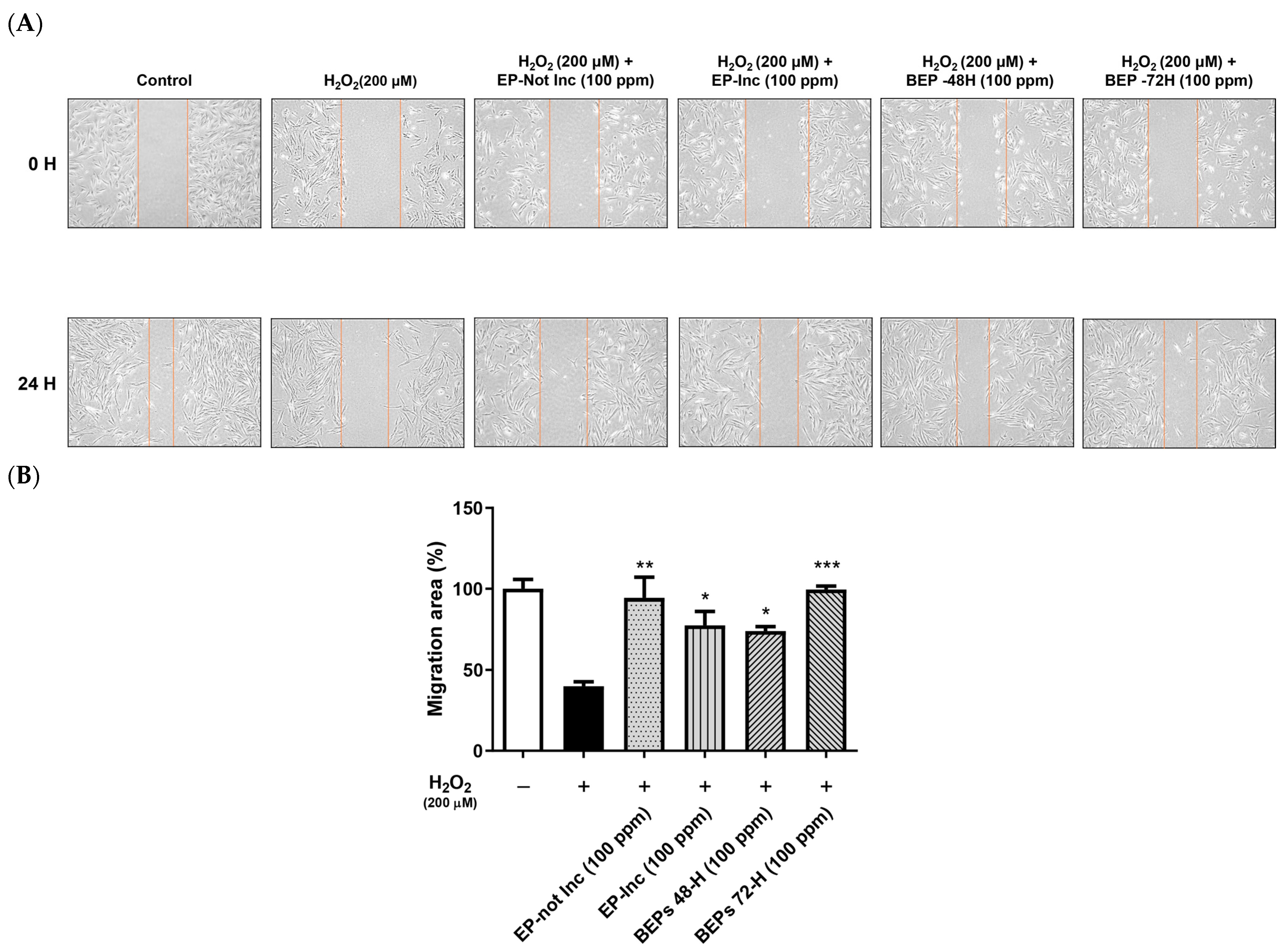
3.2. Wound-Healing Effects of BEPs in H2O2-Damaged HDFs
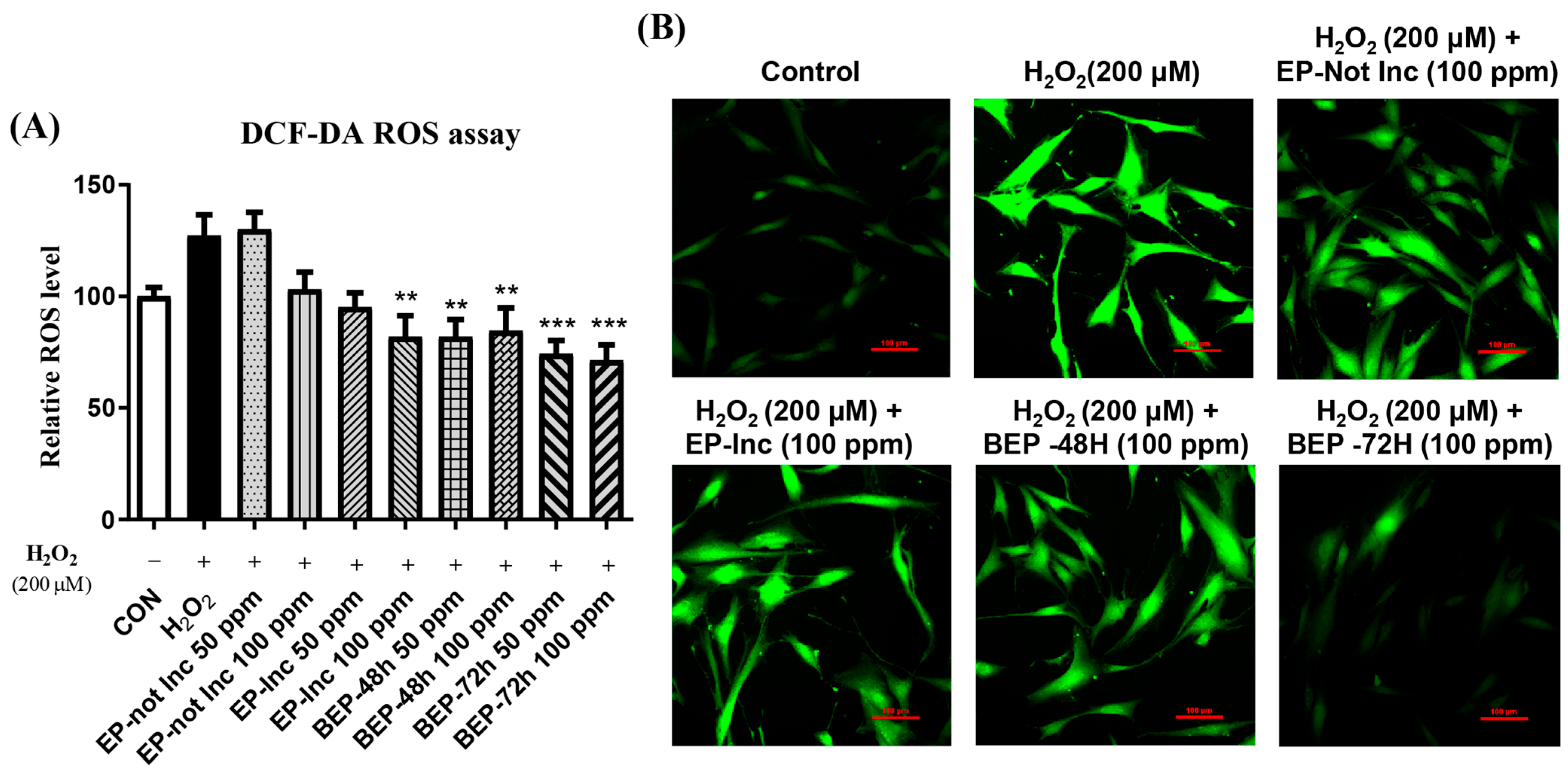
3.3. Inhibitory Effect of BEPs on ROS Generation in H2O2-Damaged HDFs
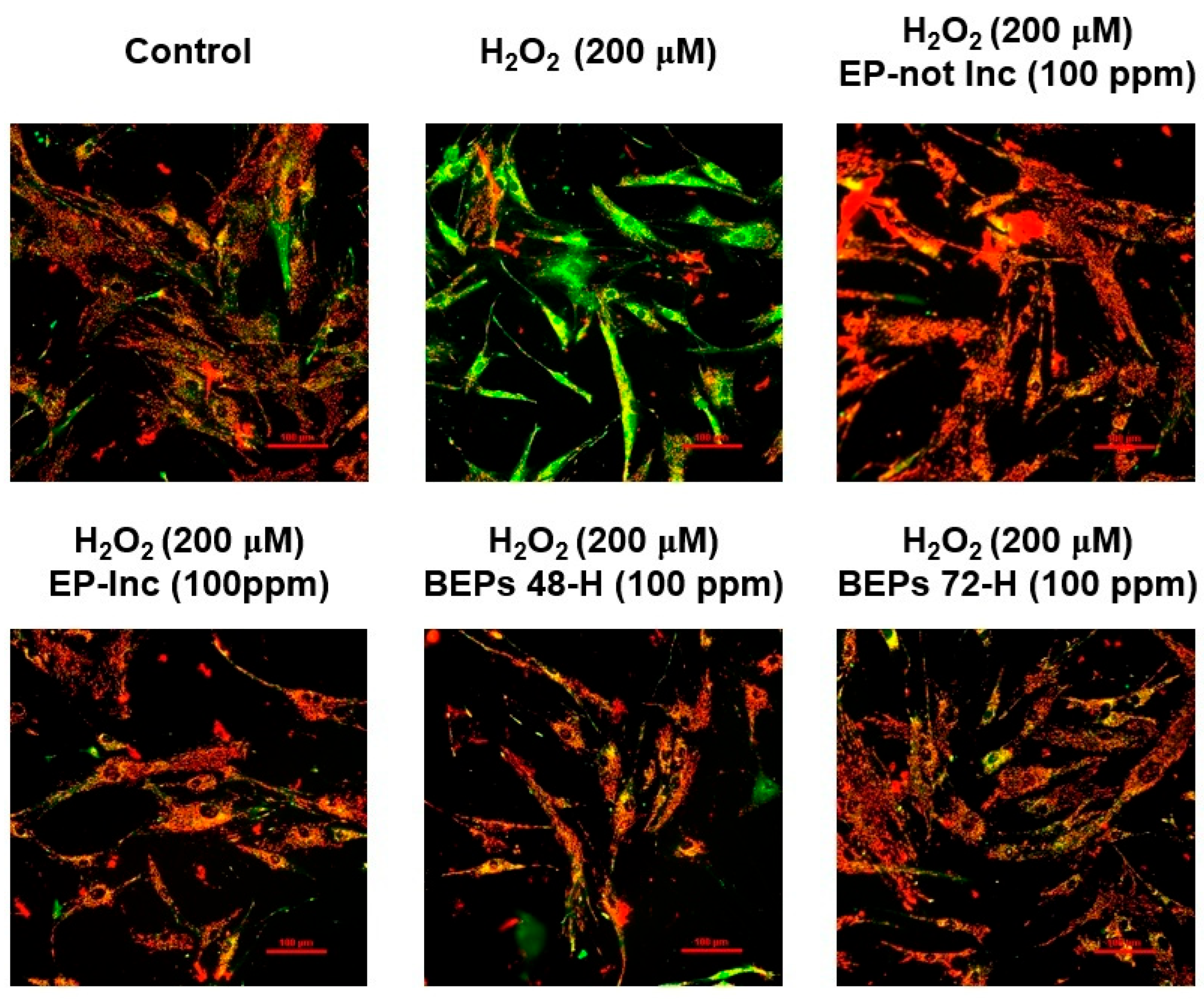
3.4. Effects of BEPs on the Membrane Potential of Mitochondria in H2O2-Damaged HDFs
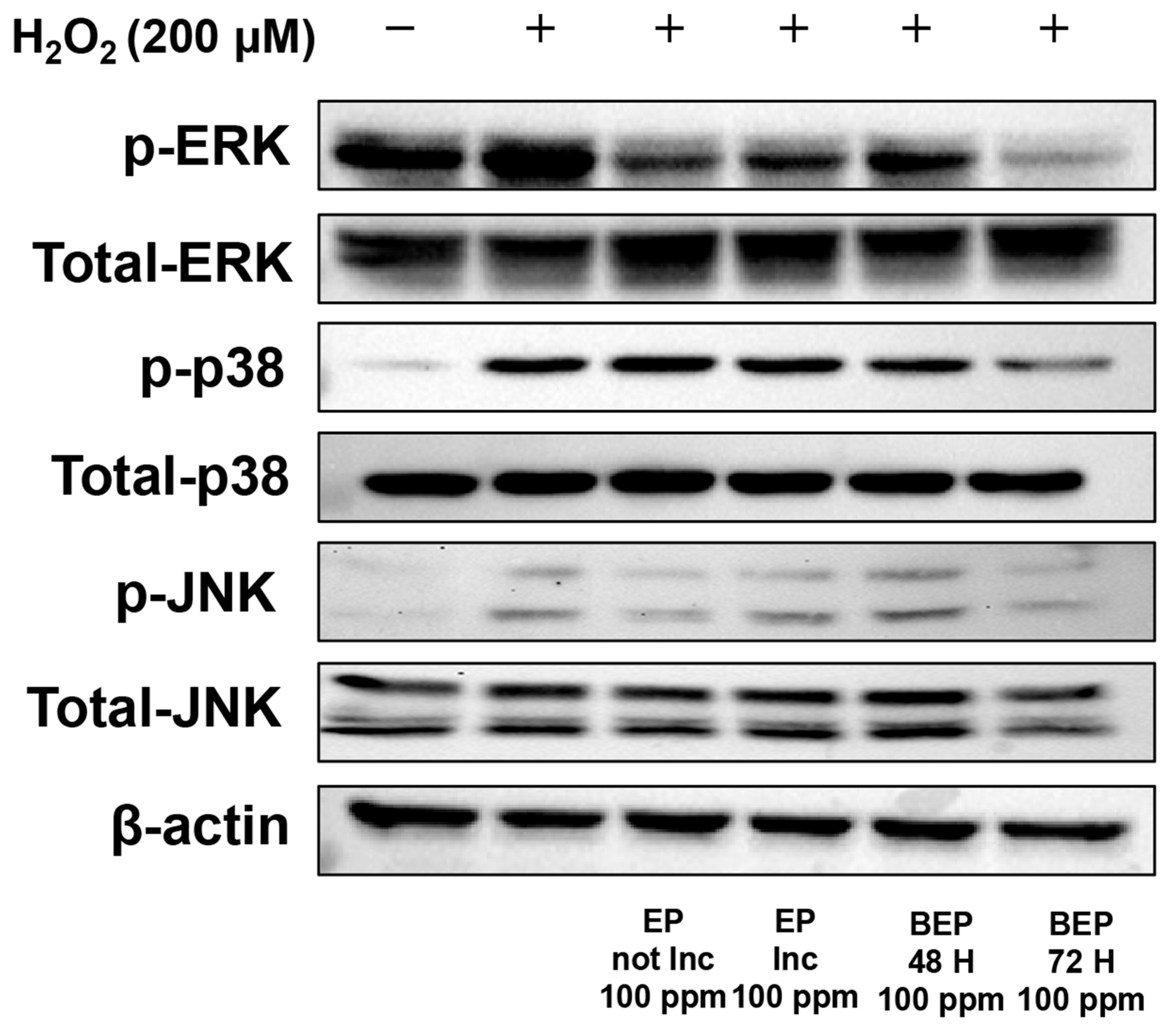
3.5. Effect of BEPs on the Phosphorylation Levels of MAPKs in H2O2-Damaged HDFs
3.6. Effect of BEPs on UVB-Induced Skin Damage
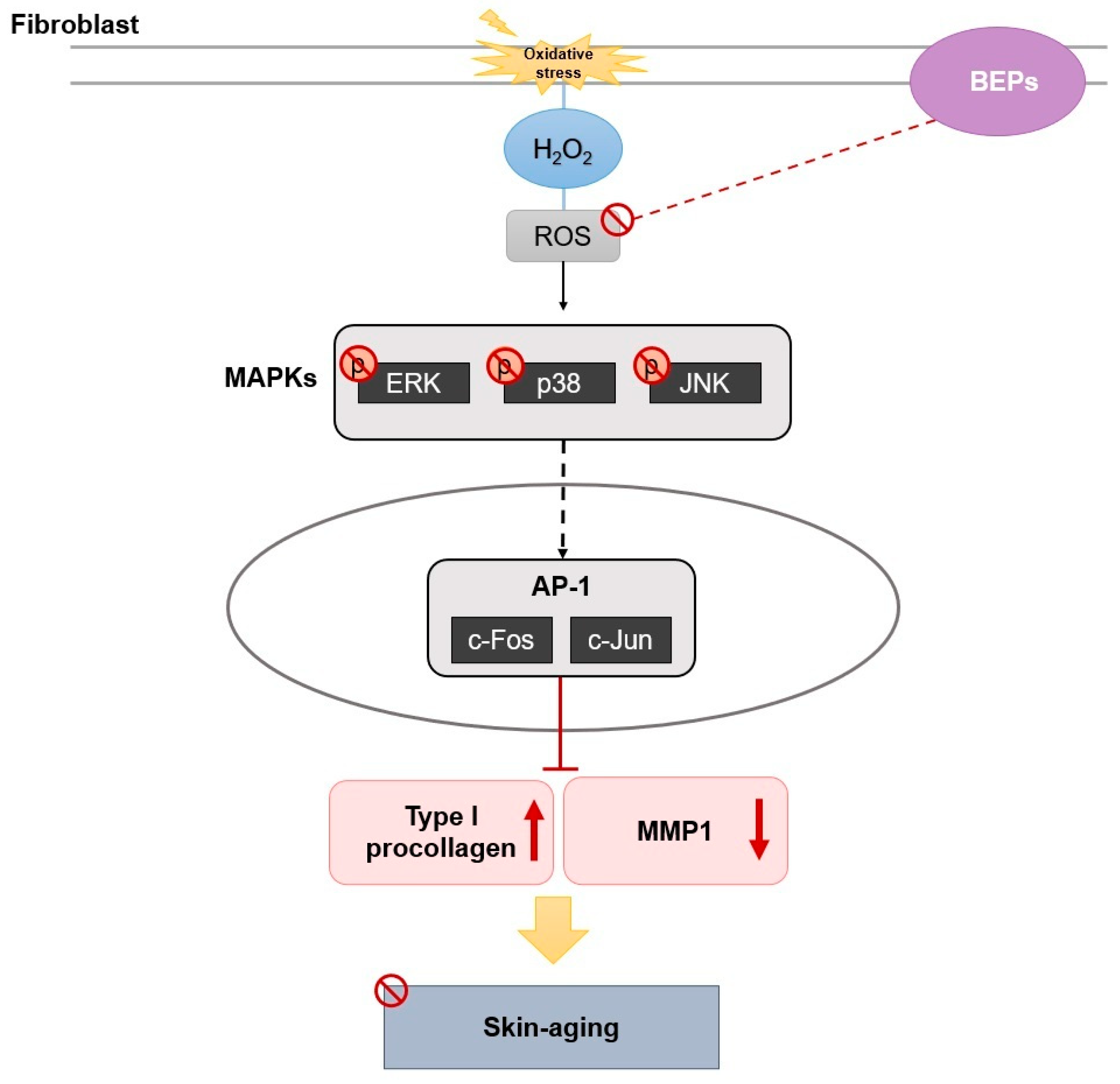
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BEPs | Bio-converted eggplant peels |
| COL1 | Collagen I |
| Eps | Eggplant peels |
| HDFs | Human dermal fibroblasts |
| MMPs | Matrix metallopeptidase |
| MAPK | Mitogen-activated protein kinase |
| UVB | Ultraviolet B |
| ROS | Reactive oxygen species |
| H2O2 | Hydrogen peroxide |
| HDFs | Human dermal fibroblasts |
| DCFDA | 2′, 7′ -dichlorodihydrofluorescein diacetate |
| ERK | Extracellular signal-regulated kinase |
| JNK | c-Jun N-terminal kinase |
| FBS | Fetal bovine serum |
| DMEM | High-glucose Dulbecco’s modified Eagle medium |
| IHC | Immunohistochemistry |
| H&E | Hematoxylin and eosin |
| MT | Masson’s trichrome |
| CPDs | Cyclobutane pyrimidine dimers |
| 6-4PPs | 6-4 photoproducts |
References
- Csekes, E.; Rackova, L. Skin Aging, Cellular Senescence and Natural Polyphenols. Int. J. Mol. Sci. 2021, 22, 12641. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Lee, J.; Kim, J.; Jung, T.; Cho, B.; Choi, S.; Lee, D.; Kim, J.; Kim, J.; Lee, O. Ulmus Macrocarpa Hance Extracts Attenuated H(2)O(2) and UVB-Induced Skin Photo-Aging by Activating Antioxidant Enzymes and Inhibiting MAPK Pathways. Int. J. Mol. Sci. 2017, 18, 1200. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Kwon, S.; Choi, J.; Na, J.; Huh, C.; Choi, H.; Park, K. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [PubMed]
- Philippeos, C.; Telerman, S.B.; Oules, B.; Pisco, A.O.; Shaw, T.J.; Elgueta, R.; Lombardi, G.; Driskell, R.R.; Soldin, M.; Lynch, M.D.; et al. Spatial and Single-Cell Transcriptional Profiling Identifies Functionally Distinct Human Dermal Fibroblast Subpopulations. J. Investig. Dermatol. 2018, 138, 811–825. [Google Scholar] [CrossRef]
- Harper, R.A.; Grove, G. Human Skin Fibroblasts Derived from Papillary and Reticular Dermis: Differences in Growth Potential in Vitro. Science 1979, 204, 526–527. [Google Scholar] [CrossRef]
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.H.; Lee, Y.H.; Rho, N.; Park, K.Y. Skin Aging from Mechanisms to Interventions: Focusing on Dermal Aging. Front. Physiol. 2023, 14, 1195272. [Google Scholar] [CrossRef]
- Wlaschek, M.; Tantcheva-Poor, I.; Naderi, L.; Ma, W.; Schneider, L.A.; Razi-Wolf, Z.; Schuller, J.; Scharffetter-Kochanek, K. Solar UV Irradiation and Dermal Photoaging. J. Photochem. Photobiol. B. 2001, 63, 41–51. [Google Scholar] [CrossRef]
- Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef]
- Rysava, A.; Vostalova, J.; Rajnochova Svobodova, A. Effect of Ultraviolet Radiation on the Nrf2 Signaling Pathway in Skin Cells. Int. J. Radiat. Biol. 2021, 97, 1383–1403. [Google Scholar] [CrossRef]
- Brenneisen, P.; Briviba, K.; Wlaschek, M.; Wenk, J.; Scharffetter-Kochanek, K. Hydrogen Peroxide (H2O2) Increases the Steady-State mRNA Levels of Collagenase/MMP-1 in Human Dermal Fibroblasts. Free Radic. Biol. Med. 1997, 22, 515–524. [Google Scholar]
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunaga, M.; Tsuru, K.; Horikawa, T. UV-Induced Skin Damage. Toxicology 2003, 189, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Mohania, D.; Chandel, S.; Kumar, P.; Verma, V.; Digvijay, K.; Tripathi, D.; Choudhury, K.; Mitten, S.K.; Shah, D. Ultraviolet Radiations: Skin Defense-Damage Mechanism. Adv. Exp. Med. Biol. 2017, 996, 71–87. [Google Scholar] [PubMed]
- Kim, J.; Kim, S.Y.; Noh, E.; Song, H.; Lee, G.; Kwon, K.; Lee, Y. Reversine Inhibits MMP-1 and MMP-3 Expressions by Suppressing of ROS/MAPK/AP-1 Activation in UV-Stimulated Human Keratinocytes and Dermal Fibroblasts. Exp. Dermatol. 2018, 27, 298–301. [Google Scholar] [CrossRef]
- Jang, H.; Kim, G.; Kim, J.; Kang, S.Y.; Youn, H.; Park, J.; Ro, S.Y.; Chung, E.; Park, K.; Kim, J. Fisetin Inhibits UVA-Induced Expression of MMP-1 and MMP-3 through the NOX/ROS/MAPK Pathway in Human Dermal Fibroblasts and Human Epidermal Keratinocytes. Int. J. Mol. Sci. 2023, 24, 17358. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, J.; Shin, D.W. The Molecular Mechanism of Polyphenols with Anti-Aging Activity in Aged Human Dermal Fibroblasts. Molecules 2022, 27, 4351. [Google Scholar] [CrossRef]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in Skin Health, Aging, and Disease. Cell Death Dis. 2020, 11, 444. [Google Scholar] [CrossRef]
- Kim, J.; Go, M.Y.; Jeon, C.Y.; Shin, J.U.; Kim, M.; Lim, H.W.; Shin, D.W. Pinitol Improves Diabetic Foot Ulcers in Streptozotocin-Induced Diabetes Rats through Upregulation of Nrf2/HO-1 Signaling. Antioxidants 2024, 14, 15. [Google Scholar] [CrossRef]
- Estrella-Osuna, D.E.; Tapia-Hernandez, J.A.; Ruiz-Cruz, S.; Marquez-Rios, E.; Ornelas-Paz, J.d.J.; Del-Toro-Sanchez, C.L.; Ocano-Higuera, V.M.; Rodriguez-Felix, F.; Estrada-Alvarado, M.I.; Cira-Chavez, L.A. Nanoencapsulation of Eggplant (Solanum melongena L.) Peel Extract in Electrospun Gelatin Nanofiber: Preparation, Characterization, and in Vitro Release. Nanomaterials 2022, 12, 2303. [Google Scholar] [CrossRef]
- Dranca, F.; Oroian, M. Optimization of Ultrasound-Assisted Extraction of Total Monomeric Anthocyanin (TMA) and Total Phenolic Content (TPC) from Eggplant (Solanum melongena L.) Peel. Ultrason. Sonochem. 2016, 31, 637–646. [Google Scholar] [CrossRef]
- Docimo, T.; Francese, G.; Ruggiero, A.; Batelli, G.; De Palma, M.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Mennella, G.; Tucci, M. Phenylpropanoids Accumulation in Eggplant Fruit: Characterization of Biosynthetic Genes and Regulation by a MYB Transcription Factor. Front. Plant Sci. 2016, 6, 1233. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Shen, L.; Yu, J.; Wan, H.; Guo, A.; Chen, J.; Long, Y.; Zhao, J.; Pei, G. Rapamycin and Other Longevity-Promoting Compounds Enhance the Generation of Mouse Induced Pluripotent Stem Cells. Aging Cell 2011, 10, 908–911. [Google Scholar] [CrossRef] [PubMed]
- Dranca, F.; Oroian, M. Total Monomeric Anthocyanin, Total Phenolic Content and Antioxidant Activity of Extracts from Eggplant (Solanum melongena L.) Peel using Ultrasonic Treatments. J. Food Process Eng. 2015, 40, e12312. [Google Scholar] [CrossRef]
- Han, K.J.; Lee, J.; Lee, N.; Paik, H. Antioxidant and Anti-Inflammatory Effect of Probiotic Lactobacillus Plantarum KU15149 Derived from Korean Homemade Diced-Radish Kimchi. J. Microbiol. Biotechnol. 2020, 30, 591–598. [Google Scholar] [CrossRef]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus Plantarum and its Probiotic and Food Potentialities. Probiotics Antimicrob. Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Hong, S.M.; Kang, M.C.; Jin, M.; Lee, T.H.; Lim, B.O.; Kim, S.Y. Fermented Blueberry and Black Rice Containing Lactobacillus Plantarum MG4221: A Novel Functional Food for Particulate Matter (PM(2.5))/Dinitrochlorobenzene (DNCB)-Induced Atopic Dermatitis. Food Funct. 2021, 12, 3611–3623. [Google Scholar] [CrossRef]
- Rwubuzizi, R.; Kim, H.; Holzapfel, W.H.; Todorov, S.D. Beneficial, Safety, and Antioxidant Properties of Lactic Acid Bacteria: A Next Step in their Evaluation as Potential Probiotics. Heliyon 2023, 9, e15610. [Google Scholar] [CrossRef]
- DUz, M.; DoGan, Y.N.; DoGan, I. Antioxidant Activitiy of Lactobacillus Plantarum, Lactobacillus Sake and Lactobacillus Curvatus Strains Isolated from Fermented Turkish Sucuk. An. Acad. Bras. Cienc. 2020, 92, e20200105. [Google Scholar] [CrossRef]
- AlKalbani, N.S.; Turner, M.S.; Ayyash, M.M. Isolation, Identification, and Potential Probiotic Characterization of Isolated Lactic Acid Bacteria and in Vitro Investigation of the Cytotoxicity, Antioxidant, and Antidiabetic Activities in Fermented Sausage. Microb. Cell Factories 2019, 18, 188. [Google Scholar] [CrossRef]
- Antognoni, F.; Mandrioli, R.; Potente, G.; Taneyo Saa, D.L.; Gianotti, A. Changes in Carotenoids, Phenolic Acids and Antioxidant Capacity in Bread Wheat Doughs Fermented with Different Lactic Acid Bacteria Strains. Food Chem. 2019, 292, 211–216. [Google Scholar] [CrossRef]
- Tsai, W.; Chou, C.; Chiang, Y.; Lin, C.; Lee, C. Regulatory Effects of Lactobacillus Plantarum-GMNL6 on Human Skin Health by Improving Skin Microbiome. Int. J. Med. Sci. 2021, 18, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus Plantarum with Functional Properties: An Approach to Increase Safety and Shelf-Life of Fermented Foods. BioMed Res. Int. 2018, 2018, 9361614. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.; Tsai, W.; Jheng, Y.; Su, S.; Wang, S.; Lin, C.; Chen, Y.; Chang, W. The Beneficial Effects of Lactobacillus Reuteri ADR-1 Or ADR-3 Consumption on Type 2 Diabetes Mellitus: A Randomized, Double-Blinded, Placebo-Controlled Trial. Sci. Rep. 2018, 8, 16791. [Google Scholar] [CrossRef]
- Chang, C.; Lin, T.; Tsai, Y.; Wu, T.; Lai, W.; Lu, C.; Lai, H. Next Generation Probiotics in Disease Amelioration. J. Food Drug Anal. 2019, 27, 615–622. [Google Scholar] [CrossRef]
- Le, B.; Yang, S.H. Efficacy of Lactobacillus Plantarum in Prevention of Inflammatory Bowel Disease. Toxicol. Rep. 2018, 5, 314–317. [Google Scholar] [CrossRef]
- Sharma, R.; Diwan, B.; Singh, B.P.; Kulshrestha, S. Probiotic Fermentation of Polyphenols: Potential Sources of Novel Functional Foods. Food Prod. Process. Nutr. 2022, 4, 21. [Google Scholar] [CrossRef]
- Li, Z.; Teng, J.; Lyu, Y.; Hu, X.; Zhao, Y.; Wang, M. Enhanced Antioxidant Activity for Apple Juice Fermented with Lactobacillus Plantarum ATCC14917. Molecules 2018, 24, 51. [Google Scholar] [CrossRef]
- Hu, Y.; Rao, S.; Wang, Z.; Cao, J.; Tan, Y.; Luo, J.; Li, H.; Zhang, W.; Chen, C.; Xie, H. Exosomes from Human Umbilical Cord Blood Accelerate Cutaneous Wound Healing through miR-21-3p-Mediated Promotion of Angiogenesis and Fibroblast Function. Theranostics 2018, 8, 169–184. [Google Scholar] [CrossRef]
- Zhu, B.; Liang, L.; Hui, L.; Lu, Y. Exploring the Role of Dermal Sheath Cells in Wound Healing and Fibrosis. Wound Repair Regen. 2024, 32, 735–745. [Google Scholar] [CrossRef]
- Ma, W.; Wlaschek, M.; Tantcheva-Poor, I.; Schneider, L.A.; Naderi, L.; Razi-Wolf, Z.; Schuller, J.; Scharffetter-Kochanek, K. Chronological Ageing and Photoageing of the Fibroblasts and the Dermal Connective Tissue. Clin. Exp. Dermatol. 2001, 26, 592–599. [Google Scholar] [CrossRef]
- Scharffetter-Kochanek, K.; Brenneisen, P.; Wenk, J.; Herrmann, G.; Ma, W.; Kuhr, L.; Meewes, C.; Wlaschek, M. Photoaging of the Skin from Phenotype to Mechanisms. Exp. Gerontol. 2000, 35, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Role of Metabolic H2O2 Generation: Redox Signaling and Oxidative Stress. J. Biol. Chem. 2014, 289, 8735–8741. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Boyman, L.; Karbowski, M.; Lederer, W.J. Regulation of Mitochondrial ATP Production: Ca(2+) Signaling and Quality Control. Trends Mol. Med. 2020, 26, 21–39. [Google Scholar] [CrossRef]
- Shadel, G.S.; Horvath, T.L. Mitochondrial ROS Signaling in Organismal Homeostasis. Cell 2015, 163, 560–569. [Google Scholar] [CrossRef]
- Natarelli, N.; Gahoonia, N.; Aflatooni, S.; Bhatia, S.; Sivamani, R.K. Dermatologic Manifestations of Mitochondrial Dysfunction: A Review of the Literature. Int. J. Mol. Sci. 2024, 25, 3303. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial Reactive Oxygen Species (ROS) and ROS-Induced ROS Release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial Membrane Potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef]
- Li, L.; Ngo, H.T.T.; Hwang, E.; Wei, X.; Liu, Y.; Liu, J.; Yi, T. Conditioned Medium from Human Adipose-Derived Mesenchymal Stem Cell Culture Prevents UVB-Induced Skin Aging in Human Keratinocytes and Dermal Fibroblasts. Int. J. Mol. Sci. 2019, 21, 49. [Google Scholar] [CrossRef]
- Kim, M.; Park, Y.G.; Lee, H.; Lim, S.J.; Nho, C.W. Youngiasides A and C Isolated from Youngia Denticulatum Inhibit UVB-Induced MMP Expression and Promote Type I Procollagen Production Via Repression of MAPK/AP-1/NF-kappaB and Activation of AMPK/Nrf2 in HaCaT Cells and Human Dermal Fibroblasts. J. Agric. Food Chem. 2015, 63, 5428–5438. [Google Scholar] [CrossRef]
- Bonifant, H.; Holloway, S. A Review of the Effects of Ageing on Skin Integrity and Wound Healing. Br. J. Community Nurs. 2019, 24, S28–S33. [Google Scholar] [CrossRef] [PubMed]
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, Prevention and Therapy. Br. J. Dermatol. 2007, 157, 874–887. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.E.B.; Gibbs, N.K.; Griffiths, C.E.M.; Sherratt, M.J. Damage to Skin Extracellular Matrix Induced by UV Exposure. Antioxid. Redox Signal. 2014, 21, 1063–1077. [Google Scholar] [CrossRef]
- Rittie, L.; Fisher, G.J. UV-Light-Induced Signal Cascades and Skin Aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [CrossRef]
- Fisher, G.J.; Voorhees, J.J. Molecular Mechanisms of Photoaging and its Prevention by Retinoic Acid: Ultraviolet Irradiation Induces MAP Kinase Signal Transduction Cascades that Induce Ap-1-Regulated Matrix Metalloproteinases that Degrade Human Skin in Vivo. J. Investig. Dermatol. Symp. Proc. 1998, 3, 61–68. [Google Scholar] [PubMed]
- Curran, S.; Murray, G.I. Matrix Metalloproteinases in Tumour Invasion and Metastasis. J. Pathol. 1999, 189, 300–308. [Google Scholar] [CrossRef]
- Kahari, V.M.; Saarialho-Kere, U. Matrix Metalloproteinases in Skin. Exp. Dermatol. 1997, 6, 199–213. [Google Scholar] [CrossRef]
- Lee, J.; Kim, E.; Kim, J.H.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Kim, J.; Kim, S.H.; Park, C.; Seo, D.B.; et al. Antimelanogenesis and Skin-Protective Activities of Panax Ginseng Calyx Ethanol Extract. J. Ginseng Res. 2018, 42, 389–399. [Google Scholar] [CrossRef]
- Bratic, A.; Larsson, N. The Role of Mitochondria in Aging. J. Clin. Investig. 2013, 123, 951–957. [Google Scholar] [CrossRef]
- Trifunovic, A.; Larsson, N. Mitochondrial Dysfunction as a Cause of Ageing. J. Intern. Med. 2008, 263, 167–178. [Google Scholar] [CrossRef]
- Kaunda, J.S.; Zhang, Y. The Genus Solanum: An Ethnopharmacological, Phytochemical and Biological Properties Review. Nat. Prod. Bioprospect 2019, 9, 77–137. [Google Scholar] [CrossRef] [PubMed]
- Elizalde-Romero, C.A.; Montoya-Inzunza, L.A.; Contreras-Angulo, L.A.; Heredia, J.B.; Gutierrez-Grijalva, E.P. Solanum Fruits: Phytochemicals, Bioaccessibility and Bioavailability, and their Relationship with their Health-Promoting Effects. Front. Nutr. 2021, 8, 790582. [Google Scholar] [CrossRef] [PubMed]
- Sembring, H.S.B.; Chin, K.B. Antioxidant Activities of Eggplant (Solanum melongena) Powder with Different Drying Methods and Addition Levels to Pork Sausages. Food Sci. Anim. Resour. 2021, 41, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Hanson, P.M.; Yang, R.; Tsou, S.C.S.; Ledesma, D.; Engle, L.; Lee, T. Diversity in Eggplant (Solanum melongena) for Superoxide Scavenging Activity, Total Phenolics, and Ascorbic Acid. J. Food Compos. Anal. 2006, 19, 594–600. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.H.; Kim, J.; Jo, Y.C.; Jo, Y.H.; Jeong, Y.H.; Jeong, S.A.; Lim, B.O.; Shin, D.W. Enhanced Antioxidant and Protective Effects of Fermented Solanum melongena L. Peel Extracts Against Ultraviolet B-Induced Skin Damage. Nutrients 2025, 17, 847. https://doi.org/10.3390/nu17050847
Lee JH, Kim J, Jo YC, Jo YH, Jeong YH, Jeong SA, Lim BO, Shin DW. Enhanced Antioxidant and Protective Effects of Fermented Solanum melongena L. Peel Extracts Against Ultraviolet B-Induced Skin Damage. Nutrients. 2025; 17(5):847. https://doi.org/10.3390/nu17050847
Chicago/Turabian StyleLee, Joo Hwa, Jinsick Kim, Yu Chang Jo, Yun Hoo Jo, Yeong Hwan Jeong, Soo Ah Jeong, Beong Ou Lim, and Dong Wook Shin. 2025. "Enhanced Antioxidant and Protective Effects of Fermented Solanum melongena L. Peel Extracts Against Ultraviolet B-Induced Skin Damage" Nutrients 17, no. 5: 847. https://doi.org/10.3390/nu17050847
APA StyleLee, J. H., Kim, J., Jo, Y. C., Jo, Y. H., Jeong, Y. H., Jeong, S. A., Lim, B. O., & Shin, D. W. (2025). Enhanced Antioxidant and Protective Effects of Fermented Solanum melongena L. Peel Extracts Against Ultraviolet B-Induced Skin Damage. Nutrients, 17(5), 847. https://doi.org/10.3390/nu17050847