
Whey Protein Isolate-Encapsulated Astaxanthin Nanoemulsion More Effectively Mitigates Skeletal Muscle Atrophy in Dexamethasone-Induced Mice

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Emulsion
2.3. Particle Size and Zeta Potential Determination
2.4. Observation of Microscopic Morphology
2.5. Measurement of Encapsulation Efficiency of Emulsion
2.6. In Vitro Digestion Experiment
2.7. GC-IMS Analysis
2.8. Effect of the W-A on Skeletal Muscle Atrophy in Mice
2.8.1. Animal Experiment
2.8.2. Histopathological Evaluation
2.8.3. Western Blotting Experiment
2.8.4. Biochemical Assays
2.8.5. RNA Extraction and RT-qPCR
2.9. Statistical Analysis
3. Results
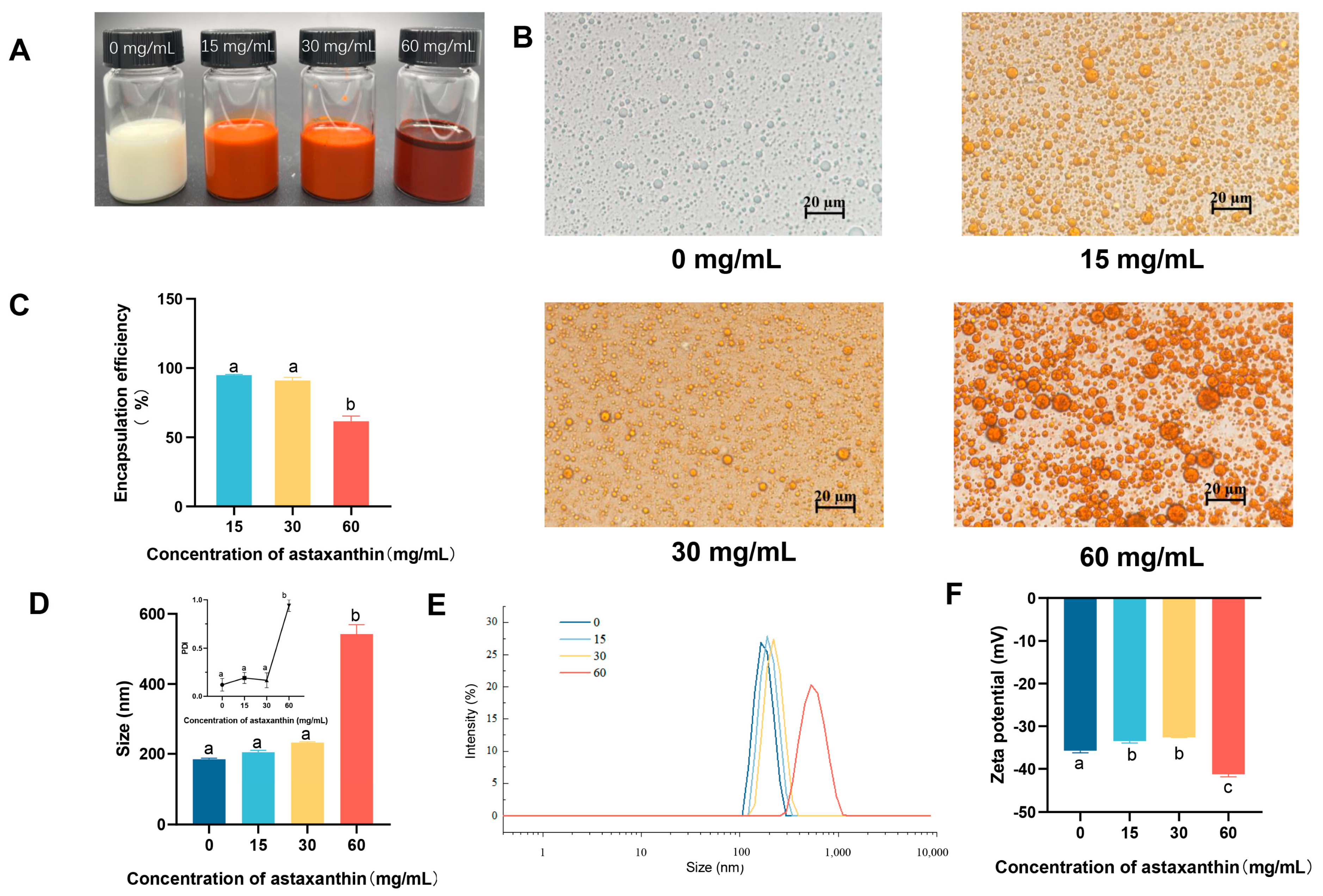
3.1. The Effect of Astaxanthin Concentration on Emulsions
3.2. The Effect of Oil/Water Ratio on Emulsions
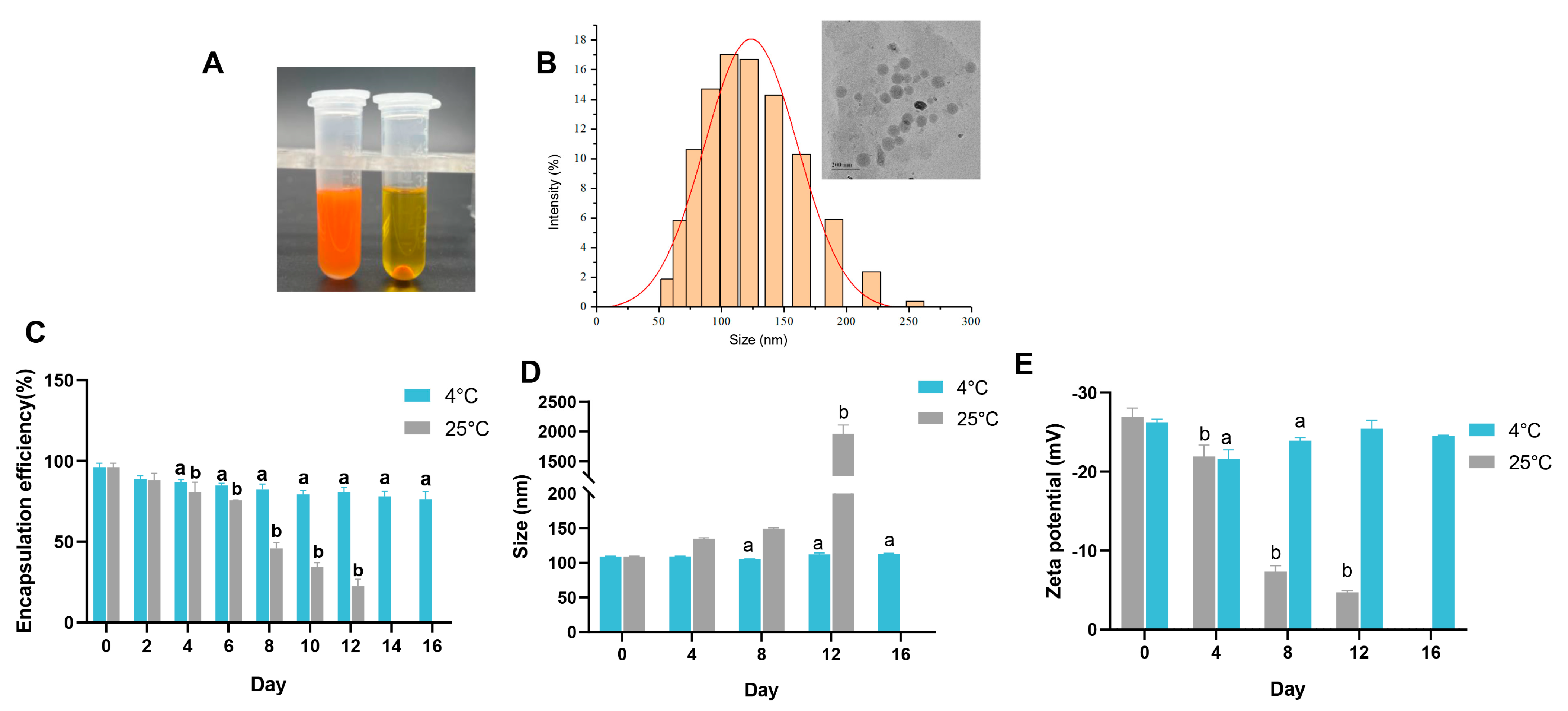
3.3. Properties of the Emulsion
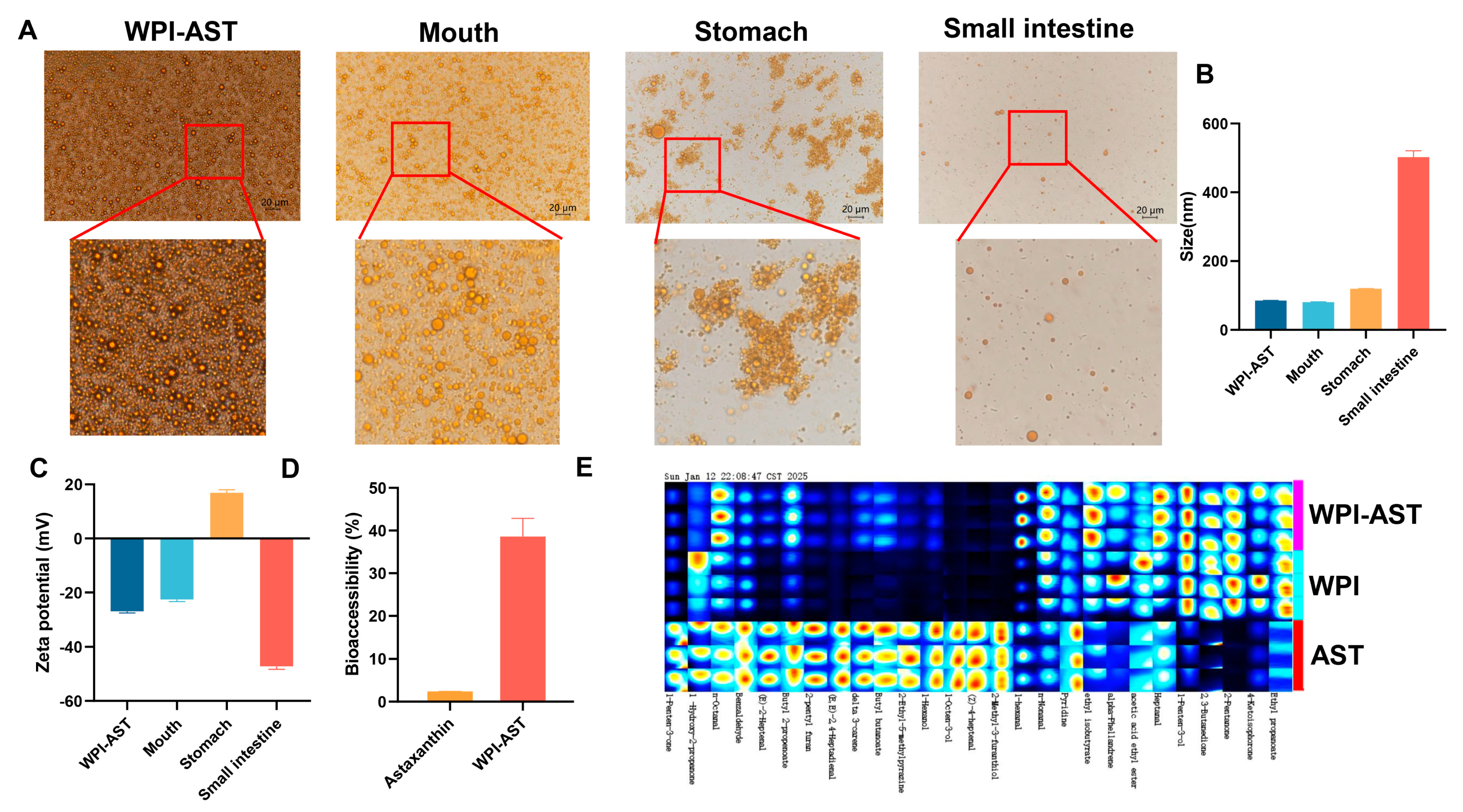
3.4. Storage Stability, In Vitro Digestive Simulation, and Undesirable Flavor Masking
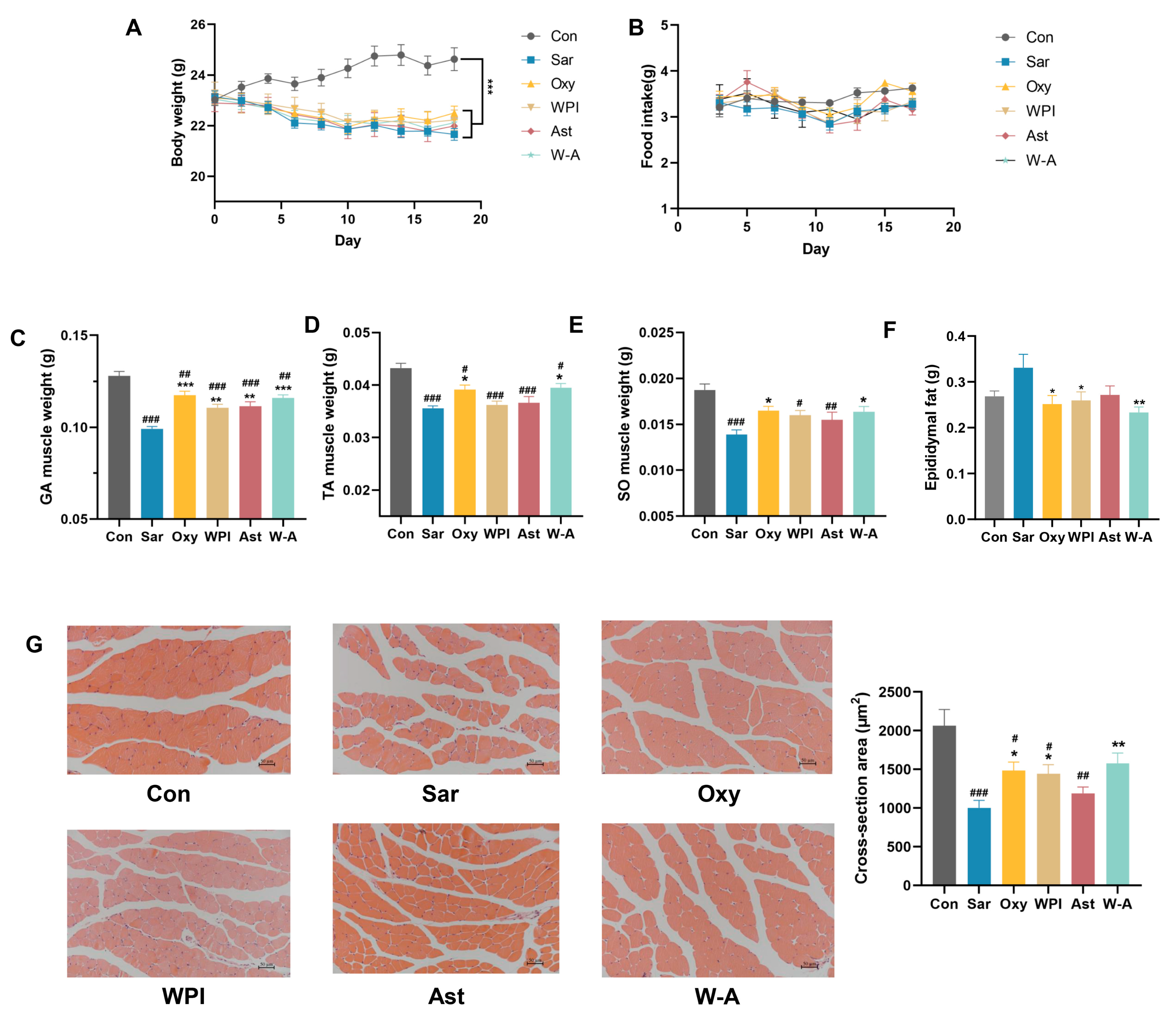
3.5. Alleviating Effect of the W-A on Dexamethasone-Induced Skeletal Muscle Atrophy in Mice
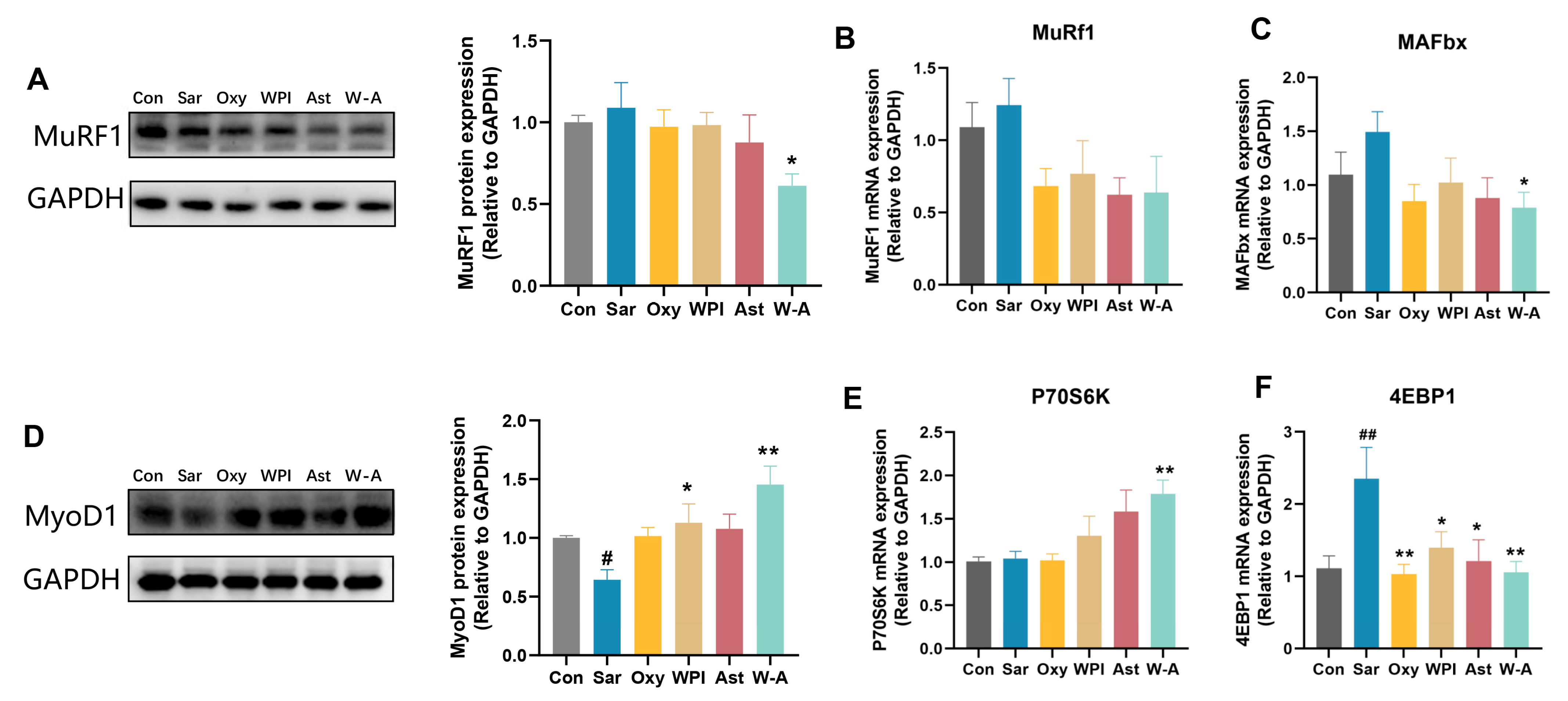
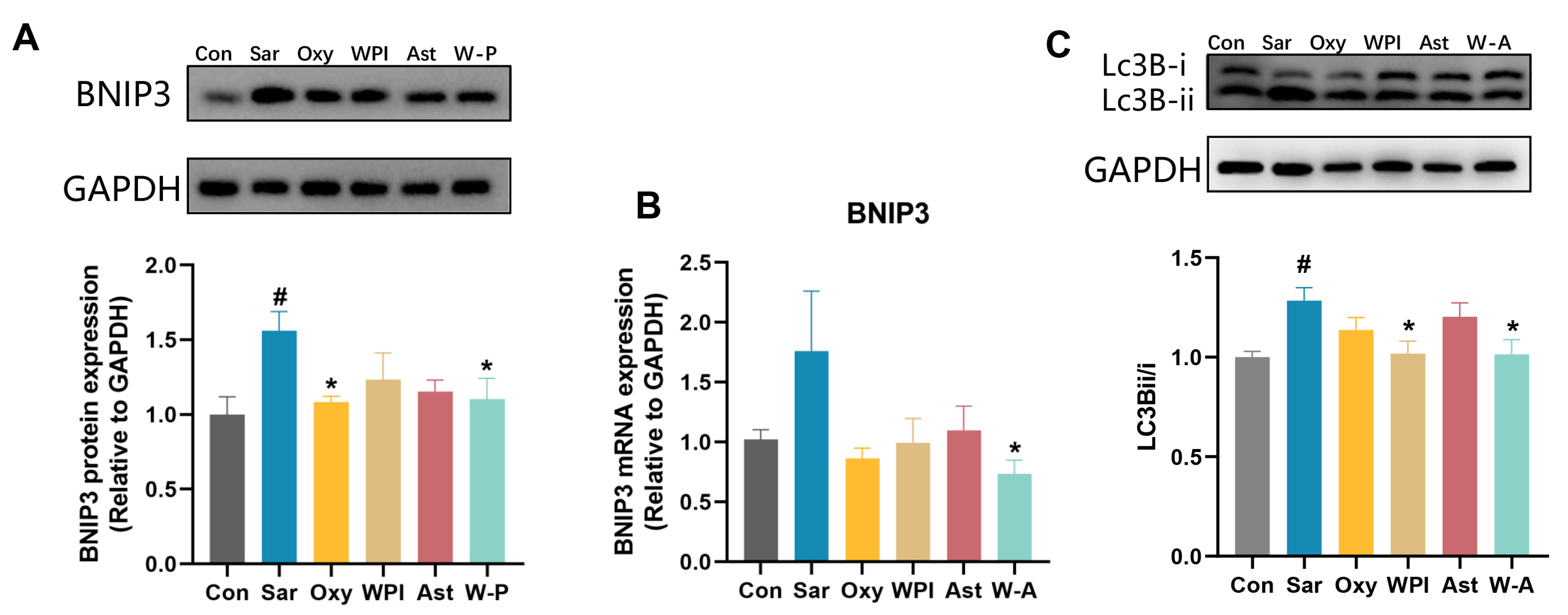
3.6. Mechanism of the W-A on Improving Skeletal Muscle Atrophy
3.7. Effects of the W-A on Glycometabolism in Skeletal Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, J.A.B.; Murach, K.A.; Dyar, K.A.; Zierath, J.R. Exercise metabolism and adaptation in skeletal muscle. Nat. Rev. Mol. Cell Biol. 2023, 24, 607–632. [Google Scholar] [CrossRef] [PubMed]
- Thyfault, J.P.; Bergouignan, A. Exercise and metabolic health: Beyond skeletal muscle. Diabetologia 2020, 63, 1464–1474. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Zhang, Y.; Qu, T.Q.; Sang, X.Q.; Li, Y.X.; Ren, F.Z.; Wen, P.C.; Sun, Y.N. Nobiletin Improves D-Galactose-Induced Aging Mice Skeletal Muscle Atrophy by Regulating Protein Homeostasis. Nutrients 2023, 15, 1801. [Google Scholar] [CrossRef] [PubMed]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of muscle atrophy and hypertrophy: Implications in health and disease. Nat. Commun. 2021, 12, 330. [Google Scholar] [CrossRef]
- Putra, C.; Konow, N.; Gage, M.; York, C.G.; Mangano, K.M. Protein Source and Muscle Health in Older Adults: A Literature Review. Nutrients 2021, 13, 743. [Google Scholar] [CrossRef]
- Park, C.Y.; Shin, S. Low dietary vitamin C intake is associated with low muscle strength among elderly Korean women. Nutr. Res. 2024, 127, 75–83. [Google Scholar] [CrossRef]
- Zhao, X.; Hua, L.; Jin, K.; Sun, Q.; Wang, R. Association between oxidative balance score and skeletal muscle mass and strength: NHANES from 2011 to 2018. Front. Nutr. 2024, 11, 1414161. [Google Scholar] [CrossRef]
- Alugoju, P.; Krishna Swamy, V.K.D.; Anthikapalli, N.V.A.; Tencomnao, T. Health benefits of astaxanthin against age-related diseases of multiple organs: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2023, 63, 10709–10774. [Google Scholar] [CrossRef]
- Waldman, H. Astaxanthin Supplementation as a Potential Strategy for Enhancing Mitochondrial Adaptations in the Endurance Athlete: An Invited Review. Nutrients 2024, 16, 1750. [Google Scholar] [CrossRef]
- Yu, X.; Ren, P.; Yang, R.; Yue, H.; Tang, Q.; Xue, C. Astaxanthin Ameliorates Skeletal Muscle Atrophy in Mice with Cancer Cachexia. Nutr. Cancer 2024, 76, 529–542. [Google Scholar] [CrossRef]
- Yue, H.; Huan, Y.; Ren, P.; Yu, X.; Tang, Q.; Xue, C.; Xu, J. Astaxanthin ameliorates dexamethasone-induced skeletal muscle atrophy and disorders of glucolipid metabolism. Biochem. Biophys. Res. Commun. 2025, 743, 151138. [Google Scholar] [CrossRef] [PubMed]
- Visentini, F.F.; Perez, A.A.; Santiago, L.G. Bioactive compounds: Application of albumin nanocarriers as delivery systems. Crit. Rev. Food Sci. Nutr. 2023, 63, 7238–7268. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Liu, X.-R.; Chen, Q.-B.; Li, Y.; Zhou, J.-L.; Zhou, L.-Y.; Zou, T. Hyaluronic acid and albumin based nanoparticles for drug delivery. J. Control. Release 2021, 331, 416–433. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, D.; Lin, B.; Zhu, L.; Li, C.; Li, X. Readily Available Oral Prebiotic Protein Reactive Oxygen Species Nanoscavengers for Synergistic Therapy of Inflammation and Fibrosis in Inflammatory Bowel Disease. ACS Nano 2024, 18, 13583–13598. [Google Scholar] [CrossRef] [PubMed]
- Shafaei, Z.; Ghalandari, B.; Vaseghi, A.; Divsalar, A.; Haertlé, T.; Saboury, A.A.; Sawyer, L. β-Lactoglobulin: An efficient nanocarrier for advanced delivery systems. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 1685–1692. [Google Scholar] [CrossRef]
- Larki, M.; Enayati, M.H.; Rostamabadi, H. Basil seed gum promotes the electrospinnability of WPI for co-encapsulation of ZnO nanoparticles and curcumin. Carbohydr. Polym. 2022, 296, 119966. [Google Scholar] [CrossRef]
- Liao, C.D.; Huang, S.W.; Chen, H.C.; Huang, M.H.; Liou, T.H.; Lin, C.L. Comparative Efficacy of Different Protein Supplements on Muscle Mass, Strength, and Physical Indices of Sarcopenia among Community-Dwelling, Hospitalized or Institutionalized Older Adults Undergoing Resistance Training: A Network Meta-Analysis of Randomized Controlled Trials. Nutrients 2024, 16, 941. [Google Scholar] [CrossRef]
- Shen, X.; Zhao, C.; Lu, J.; Guo, M. Physicochemical Properties of Whey-Protein-Stabilized Astaxanthin Nanodispersion and Its Transport via a Caco-2 Monolayer. J. Agric. Food Chem. 2018, 66, 1472–1478. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Y.; Feng, J.; Zhang, J.; Wang, X.; Li, X.; Ji, W.; Wang, Y.; Nan, B.; Liu, J.; et al. Regulation of whey protein emulsion gel’s structure with pullulan to enhance astaxanthin bioaccessibility. Carbohydr. Polym. 2025, 351, 123113. [Google Scholar] [CrossRef]
- Chen, Y.; Su, W.; Tie, S.; Cui, W.; Yu, X.; Zhang, L.; Hua, Z.; Tan, M. Orally deliverable sequence-targeted astaxanthin nanoparticles for colitis alleviation. Biomaterials 2023, 293, 121976. [Google Scholar] [CrossRef]
- Gao, Y.; Yuan, S.; Zhang, L.; Yang, L.; Liu, F.; Li, R.W.; Li, C.; Xue, C.; Xu, J.; Tang, Q. Absorbability of Astaxanthin Was Much Lower in Obese Mice Than in Normal Mice. J. Agric. Food Chem. 2020, 68, 11161–11169. [Google Scholar] [CrossRef]
- Shen, X.; Fang, T.; Zheng, J.; Guo, M. Physicochemical Properties and Cellular Uptake of Astaxanthin-Loaded Emulsions. Molecules 2019, 24, 727. [Google Scholar] [CrossRef]
- Santos, D.S.; Morais, J.A.V.; Vanderlei, Í.A.C.; Santos, A.S.; Azevedo, R.B.; Muehlmann, L.A.; Júnior, O.R.P.; Mortari, M.R.; da Silva, J.R.; da Silva, S.W.; et al. Oral delivery of fish oil in oil-in-water nanoemulsion: Development, colloidal stability and modulatory effect on in vivo inflammatory induction in mice. Biomed. Pharmacother. 2021, 133, 110980. [Google Scholar] [CrossRef]
- Xu, G.; Wang, C.; Yao, P. Stable emulsion produced from casein and soy polysaccharide compacted complex for protection and oral delivery of curcumin. Food Hydrocoll. 2017, 71, 108–117. [Google Scholar] [CrossRef]
- Boonlao, N.; Shrestha, S.; Sadiq, M.B.; Anal, A.K. Influence of whey protein-xanthan gum stabilized emulsion on stability and in vitro digestibility of encapsulated astaxanthin. J. Food Eng. 2020, 272, 109859. [Google Scholar] [CrossRef]
- Liu, X.; McClements, D.J.; Cao, Y.; Xiao, H. Chemical and Physical Stability of Astaxanthin-Enriched Emulsion-Based Delivery Systems. Food Biophys. 2016, 11, 302–310. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Zhao, W.; Wei, Z.; Xue, C. Foam-templated oleogels constructed by whey protein isolate and xanthan gum: Multiple-effect delivery vehicle for Antarctic krill oil. Int. J. Biol. Macromol. 2024, 256, 128391. [Google Scholar] [CrossRef] [PubMed]
- Zaafar, D.; Khalil, H.M.A.; Elkhouly, G.E.; Sedeky, A.S.; Ahmed, Y.H.; Khalil, M.G.; Abo-Zeid, Y. Preparation and characterization of Sorafenib nano-emulsion: Impact on pharmacokinetics and toxicity; an in vitro and in vivo study. Drug Deliv. Transl. Res. 2024, 14, 3089–3111. [Google Scholar] [CrossRef]
- Yarnpakdee, S.; Benjakul, S.; Nalinanon, S.; Kristinsson, H.G. Lipid oxidation and fishy odour development in protein hydrolysate from Nile tilapia (Oreochromis niloticus) muscle as affected by freshness and antioxidants. Food Chem. 2012, 132, 1781–1788. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, D.; Huang, Q.; Liu, Z.; Luo, X.; Xiong, S.; Yin, T. Adsorption kinetics and thermodynamics of yeast β-glucan for off-odor compounds in silver carp mince. Food Chem. 2020, 319, 126232. [Google Scholar] [CrossRef]
- Zhao, M.; Ma, H.; Hou, Y.; Li, J.; Zou, T.; Zhang, D.; Wen, R.; Li, H.; Song, H. Characterization of Key Odor-Active Off-Flavor Compounds in Aged Pasteurized Yogurt by Sensory-Directed Flavor Analysis. J. Agric. Food Chem. 2022, 70, 14439–14447. [Google Scholar] [CrossRef]
- Cecchi, L.; Migliorini, M.; Giambanelli, E.; Rossetti, A.; Cane, A.; Mulinacci, N. New Volatile Molecular Markers of Rancidity in Virgin Olive Oils under Nonaccelerated Oxidative Storage Conditions. J. Agric. Food Chem. 2019, 67, 13150–13163. [Google Scholar] [CrossRef]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef]
- Rovito, D.; Rerra, A.I.; Ueberschlag-Pitiot, V.; Joshi, S.; Karasu, N.; Dacleu-Siewe, V.; Rayana, K.B.; Ghaibour, K.; Parisotto, M.; Ferry, A.; et al. Myod1 and GR coordinate myofiber-specific transcriptional enhancers. Nucleic Acids Res. 2021, 49, 4472–4492. [Google Scholar] [CrossRef]
- Perry, R.A., Jr.; Brown, L.A.; Lee, D.E.; Brown, J.L.; Baum, J.I.; Greene, N.P.; Washington, T.A. Differential effects of leucine supplementation in young and aged mice at the onset of skeletal muscle regeneration. Mech. Ageing Dev. 2016, 157, 7–16. [Google Scholar] [CrossRef]
- Ulla, A.; Uchida, T.; Miki, Y.; Sugiura, K.; Higashitani, A.; Kobayashi, T.; Ohno, A.; Nakao, R.; Hirasaka, K.; Sakakibara, I.; et al. Morin attenuates dexamethasone-mediated oxidative stress and atrophy in mouse C2C12 skeletal myotubes. Arch. Biochem. Biophys. 2021, 704, 108873. [Google Scholar] [CrossRef]
- Troncoso, R.; Paredes, F.; Parra, V.; Gatica, D.; Vásquez-Trincado, C.; Quiroga, C.; Bravo-Sagua, R.; López-Crisosto, C.; Rodriguez, A.E.; Oyarzún, A.P.; et al. Dexamethasone-induced autophagy mediates muscle atrophy through mitochondrial clearance. Cell Cycle 2014, 13, 2281–2295. [Google Scholar] [CrossRef]
- Gao, A.; Jiang, J.; Xie, F.; Chen, L. Bnip3 in mitophagy: Novel insights and potential therapeutic target for diseases of secondary mitochondrial dysfunction. Clin. Chim. Acta 2020, 506, 72–83. [Google Scholar] [CrossRef]
- Malkawi, A.K.; Alzoubi, K.H.; Jacob, M.; Matic, G.; Ali, A.; Al Faraj, A.; Almuhanna, F.; Dasouki, M.; Abdel Rahman, A.M. Metabolomics Based Profiling of Dexamethasone Side Effects in Rats. Front. Pharmacol. 2018, 9, 46. [Google Scholar] [CrossRef]
- Mangum, J.E.; Hardee, J.P.; Fix, D.K.; Puppa, M.J.; Elkes, J.; Altomare, D.; Bykhovskaya, Y.; Campagna, D.R.; Schmidt, P.J.; Sendamarai, A.K.; et al. Pseudouridine synthase 1 deficient mice, a model for Mitochondrial Myopathy with Sideroblastic Anemia, exhibit muscle morphology and physiology alterations. Sci. Rep. 2016, 6, 26202. [Google Scholar] [CrossRef]
- Ermini, M.; Szelényi, I.; Moser, P.; Verzár, F. The Ageing of Skeletal (Striated) Muscle by Changes of Recovery Metabolism. Gerontologia 2009, 17, 300–311. [Google Scholar] [CrossRef]
- Wende, A.R.; Huss, J.M.; Schaeffer, P.J.; Giguère, V.; Kelly, D.P. PGC-1alpha coactivates PDK4 gene expression via the orphan nuclear receptor ERRalpha: A mechanism for transcriptional control of muscle glucose metabolism. Mol. Cell. Biol. 2005, 25, 10684–10694. [Google Scholar] [CrossRef]
- Kuo, T.; Liu, P.H.; Chen, T.C.; Lee, R.A.; New, J.; Zhang, D.; Lei, C.; Chau, A.; Tang, Y.; Cheung, E.; et al. Transcriptional regulation of FoxO3 gene by glucocorticoids in murine myotubes. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E572–E585. [Google Scholar] [CrossRef]
- Lundell, L.S.; Massart, J.; Altıntaş, A.; Krook, A.; Zierath, J.R. Regulation of glucose uptake and inflammation markers by FOXO1 and FOXO3 in skeletal muscle. Mol. Metab. 2019, 20, 79–88. [Google Scholar] [CrossRef]
- Abbas, S.; Hayat, K.; Karangwa, E.; Bashari, M.; Zhang, X. An Overview of Ultrasound-Assisted Food-Grade Nanoemulsions. Food Eng. Rev. 2013, 5, 139–157. [Google Scholar] [CrossRef]
- Boonlao, N.; Ruktanonchai, U.R.; Anal, A.K. Enhancing bioaccessibility and bioavailability of carotenoids using emulsion-based delivery systems. Colloids Surf. B Biointerfaces 2022, 209, 112211. [Google Scholar] [CrossRef]
- Gao, Y.; Yuan, S.; Chen, Y.; Liu, F.; Wei, Z.; Cao, W.; Li, R.W.; Xu, J.; Xue, C.; Tang, Q. The improvement effect of astaxanthin-loaded emulsions on obesity is better than that of astaxanthin in the oil phase. Food Funct. 2022, 13, 3720–3731. [Google Scholar] [CrossRef]
- Ward, M.S.; Zhao, D.Y.; Bernstein, P.S. Macular and serum carotenoid concentrations in patients with malabsorption syndromes. J. Ocul. Biol. Dis. Inform. 2008, 1, 12–18. [Google Scholar] [CrossRef]
- Martínez-Álvarez, Ó.; Calvo, M.M.; Gómez-Estaca, J. Recent Advances in Astaxanthin Micro/Nanoencapsulation to Improve Its Stability and Functionality as a Food Ingredient. Mar. Drugs 2020, 18, 406. [Google Scholar] [CrossRef]
- Qi, H.; Gao, Y.; Zhang, Z.; Zhang, X.; Tian, D.; Jiang, Y.; Zhang, L.; Zeng, N.; Yang, R. HouShiHeiSan attenuates sarcopenia in middle cerebral artery occlusion (MCAO) rats. J. Ethnopharmacol. 2025, 337, 118917. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Yan, Y.; Deng, Z.; Zhou, L.; She, M.; Yang, Y.; Zhang, M.; Wang, D. Saikosaponin A and D attenuate skeletal muscle atrophy in chronic kidney disease by reducing oxidative stress through activation of PI3K/AKT/Nrf2 pathway. Phytomed. Int. J. Phytother. Phytopharm. 2023, 114, 154766. [Google Scholar] [CrossRef] [PubMed]
- Castillero, E.; Martín, A.I.; Nieto-Bona, M.P.; Fernández-Galaz, C.; López-Menduiña, M.; Villanúa, M.; López-Calderón, A. Fenofibrate administration to arthritic rats increases adiponectin and leptin and prevents oxidative muscle wasting. Endocr. Connect. 2012, 1, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nabuco, H.C.G.; Tomeleri, C.M.; Fernandes, R.R.; Junior, P.S.; Cavalcante, E.F.; Cunha, P.M.; Antunes, M.; Nunes, J.P.; Venturini, D.; Barbosa, D.S.; et al. Effect of whey protein supplementation combined with resistance training on body composition, muscular strength, functional capacity, and plasma-metabolism biomarkers in older women with sarcopenic obesity: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. ESPEN 2019, 32, 88–95. [Google Scholar] [CrossRef]
- Cheah, K.J.; Cheah, L.J. Benefits and side effects of protein supplementation and exercise in sarcopenic obesity: A scoping review. Nutr. J. 2023, 22, 52. [Google Scholar] [CrossRef]
- Liao, Y.; Zhou, X.; Xu, S.; Meng, Z.; Li, D.; Yang, X.; Liu, L.; Yang, W. Non-coding RNAs-associated ceRNA networks involved in the amelioration of skeletal muscle aging after whey protein supplementation. J. Nutr. Biochem. 2022, 104, 108968. [Google Scholar] [CrossRef]
- Katsanos, C.S.; Kobayashi, H.; Sheffield-Moore, M.; Aarsland, A.; Wolfe, R.R. A high proportion of leucine is required for optimal stimulation of the rate of muscle protein synthesis by essential amino acids in the elderly. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E381–E387. [Google Scholar] [CrossRef]
- Luiking, Y.C.; Deutz, N.E.; Memelink, R.G.; Verlaan, S.; Wolfe, R.R. Postprandial muscle protein synthesis is higher after a high whey protein, leucine-enriched supplement than after a dairy-like product in healthy older people: A randomized controlled trial. Nutr. J. 2014, 13, 9. [Google Scholar] [CrossRef]
- Kawamura, A.; Aoi, W.; Abe, R.; Kobayashi, Y.; Wada, S.; Kuwahata, M.; Higashi, A. Combined intake of astaxanthin, β-carotene, and resveratrol elevates protein synthesis during muscle hypertrophy in mice. Nutrition 2020, 69, 110561. [Google Scholar] [CrossRef]
- Liang, H.; Ward, W.F. PGC-1alpha: A key regulator of energy metabolism. Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef]
- Kanazashi, M.; Tanaka, M.; Nakanishi, R.; Maeshige, N.; Fujino, H. Effects of astaxanthin supplementation and electrical stimulation on muscle atrophy and decreased oxidative capacity in soleus muscle during hindlimb unloading in rats. J. Physiol. Sci. 2019, 69, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guan, K.; Wang, R.; Zhu, A.; Ma, Y. Synergistic effects of MFG-E8 and whey protein on mitigating d-galactose-induced sarcopenia through PI3K/AKT/PGC-1α and MAPK/ERK signaling pathways. J. Dairy Sci. 2024, 107, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Ji, A.; Chen, W.; Zhang, T.; Shi, R.; Wang, X.; Wang, Y.; Xu, H.; Li, D. Whey protein and soy protein prevent obesity by upregulating uncoupling protein 1 to activate brown adipose tissue and promote white adipose tissue browning in high-fat diet-fed mice. Food Funct. 2022, 13, 12836–12851. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, H. Astaxanthin Modulation of Signaling Pathways That Regulate Autophagy. Mar. Drugs 2019, 17, 546. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| MuRF1 | ACCACAGAGGGTAAAGAAGAACA | GCAGAGAGAAGACACACTTCCC |
| MAFbx | ACAAAGGAAGTACGAAGGAGCG | GGCAGTCGAGAAGTCCAGTC |
| P70S6K | TGTCAGCCCAGTCAAATTCTCTCC | ACATCCATCTGCTCTATCCCACTTG |
| 4EBP1 | ACCCAGTCCTGCTCCTCACTC | CTCGGTATAGACAGAGGCACAAGG |
| BNIP3 | TTCTCACTGTGACAGCCCAC | TCTTCCTCAGACAGAGTGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huan, Y.; Yue, H.; Song, Y.; Zhang, W.; Wei, B.; Tang, Q. Whey Protein Isolate-Encapsulated Astaxanthin Nanoemulsion More Effectively Mitigates Skeletal Muscle Atrophy in Dexamethasone-Induced Mice. Nutrients 2025, 17, 750. https://doi.org/10.3390/nu17050750
Huan Y, Yue H, Song Y, Zhang W, Wei B, Tang Q. Whey Protein Isolate-Encapsulated Astaxanthin Nanoemulsion More Effectively Mitigates Skeletal Muscle Atrophy in Dexamethasone-Induced Mice. Nutrients. 2025; 17(5):750. https://doi.org/10.3390/nu17050750
Chicago/Turabian StyleHuan, Yuchen, Han Yue, Yanli Song, Wenmei Zhang, Biqian Wei, and Qingjuan Tang. 2025. "Whey Protein Isolate-Encapsulated Astaxanthin Nanoemulsion More Effectively Mitigates Skeletal Muscle Atrophy in Dexamethasone-Induced Mice" Nutrients 17, no. 5: 750. https://doi.org/10.3390/nu17050750
APA StyleHuan, Y., Yue, H., Song, Y., Zhang, W., Wei, B., & Tang, Q. (2025). Whey Protein Isolate-Encapsulated Astaxanthin Nanoemulsion More Effectively Mitigates Skeletal Muscle Atrophy in Dexamethasone-Induced Mice. Nutrients, 17(5), 750. https://doi.org/10.3390/nu17050750