
Oral–Gut Microbiota Crosstalk and Epigenetic Targets in Metabolic and Neuropsychiatric Diseases

 , and
, and
Abstract
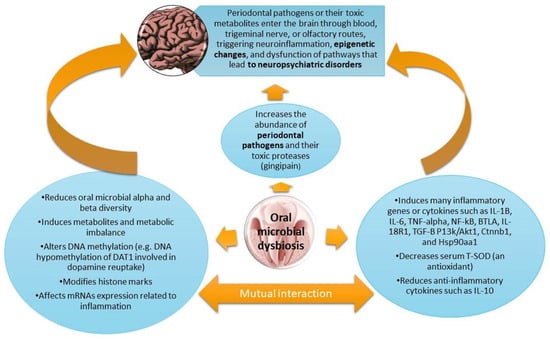
1. Introduction
2. Oral Microbiota and the Pathogenesis of Metabolic Diseases
3. Oral Microbiome and the Pathogenesis of NPDs
4. OM-Induced Inflammation Drives Metabolic and Epigenetic Alterations Underlying NPD Pathogenesis
5. Therapeutic Remedies Based on Modulation of Oral Microbiome
5.1. Oral Hygiene to Prevent or Improve OM-Induced Metabolic Dysfunctions and NPDs
5.2. Nutritional Interventions and Pre-, Pro-, and Postbiotics to Improve Oral Health and OM-Induced Epigenetic Diseases, Metabolic Diseases, and NPDs
6. Critical Points, Limitations, and Future Perspectives
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bransfield, R.C.; Mao, C.; Greenberg, R. Microbes and mental illness: Past, present, and future. In Proceedings of the Healthcare, Kyoto, Japan, 12–14 May 2023; p. 83. [Google Scholar]
- Nohesara, S.; Mostafavi Abdolmaleky, H.; Pirani, A.; Thiagalingam, S. Therapeutic Horizons: Gut Microbiome, Neuroinflammation, and Epigenetics in Neuropsychiatric Disorders. Cells 2025, 14, 1027. [Google Scholar] [CrossRef]
- Shirinian, M.; Chen, C.; Uchida, S.; Jadavji, N.M. The role of epigenetics in neuropsychiatric disorders. Front. Mol. Neurosci. 2022, 15, 985023. [Google Scholar] [CrossRef]
- Barroso, I.; McCarthy, M.I. The genetic basis of metabolic disease. Cell 2019, 177, 146–161. [Google Scholar] [CrossRef]
- Ilieva, M.S. Non-coding RNAs in neurological and neuropsychiatric disorders: Unraveling the hidden players in disease pathogenesis. Cells 2024, 13, 1063. [Google Scholar] [CrossRef]
- Zhu, H.; Guan, A.; Liu, J.; Peng, L.; Zhang, Z.; Wang, S. Noteworthy perspectives on microglia in neuropsychiatric disorders. J. Neuroinflamm. 2023, 20, 223. [Google Scholar] [CrossRef]
- Ngo, J.; Osto, C.; Villalobos, F.; Shirihai, O.S. Mitochondrial heterogeneity in metabolic diseases. Biology 2021, 10, 927. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Martín-Rodríguez, A.; Redondo-Flórez, L.; López-Mora, C.; Yáñez-Sepúlveda, R.; Tornero-Aguilera, J.F. New insights and potential therapeutic interventions in metabolic diseases. Int. J. Mol. Sci. 2023, 24, 10672. [Google Scholar] [CrossRef]
- Morella, I.M.; Brambilla, R.; Morè, L. Emerging roles of brain metabolism in cognitive impairment and neuropsychiatric disorders. Neurosci. Biobehav. Rev. 2022, 142, 104892. [Google Scholar] [CrossRef]
- López-Gambero, A.J.; Rosell-Valle, C.; Medina-Vera, D.; Navarro, J.A.; Vargas, A.; Rivera, P.; Sanjuan, C.; Rodríguez de Fonseca, F.; Suárez, J. A negative energy balance is associated with metabolic dysfunctions in the hypothalamus of a humanized preclinical model of Alzheimer’s disease, the 5XFAD mouse. Int. J. Mol. Sci. 2021, 22, 5365. [Google Scholar] [CrossRef]
- Lossi, L.; Castagna, C.; Merighi, A. An overview of the epigenetic modifications in the brain under normal and pathological conditions. Int. J. Mol. Sci. 2024, 25, 3881. [Google Scholar] [CrossRef] [PubMed]
- Nohesara, S.; Mostafavi Abdolmaleky, H.; Pirani, A.; Thiagalingam, S. Epigenetics and Gut Microbiota in the Pathogenesis and Treatment of Bipolar Disorder (BD). Cells 2025, 14, 1104. [Google Scholar] [CrossRef]
- Wu, Y.-L.; Lin, Z.-J.; Li, C.-C.; Lin, X.; Shan, S.-K.; Guo, B.; Zheng, M.-H.; Li, F.; Yuan, L.-Q.; Li, Z.-h. Epigenetic regulation in metabolic diseases: Mechanisms and advances in clinical study. Signal Transduct. Target. Ther. 2023, 8, 98. [Google Scholar] [CrossRef]
- Abdolmaleky, H.M.; Martin, M.; Zhou, J.-R.; Thiagalingam, S. Epigenetic alterations of brain non-neuronal cells in major mental diseases. Genes 2023, 14, 896. [Google Scholar] [CrossRef]
- Pizzorusso, T.; Tognini, P. Interplay between metabolism, nutrition and epigenetics in shaping brain DNA methylation, neural function and behavior. Genes 2020, 11, 742. [Google Scholar] [CrossRef]
- Huang, K.; Li, S.; Yang, M.; Teng, Z.; Xu, B.; Wang, B.; Chen, J.; Zhao, L.; Wu, H. The epigenetic mechanism of metabolic risk in bipolar disorder. Obes. Rev. 2024, 25, e13816. [Google Scholar] [CrossRef]
- Cui, J.; Zhai, Z.; Wang, S.; Song, X.; Qiu, T.; Yu, L.; Zhai, Q.; Zhang, H. The role and impact of abnormal vitamin levels in autism spectrum disorders. Food Funct. 2024, 15, 1099–1115. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi Abdolmaleky, H.; Zhou, J.-R. Gut microbiota dysbiosis, oxidative stress, inflammation, and epigenetic alterations in metabolic diseases. Antioxidants 2024, 13, 985. [Google Scholar] [CrossRef] [PubMed]
- Rivera, E.J.; Goldin, A.; Fulmer, N.; Tavares, R.; Wands, J.R.; de la Monte, S.M. Insulin and insulin-like growth factor expression and function deteriorate with progression of Alzheimer’s disease: Link to brain reductions in acetylcholine. J. Alzheimer’s Dis. 2005, 8, 247–268. [Google Scholar] [CrossRef] [PubMed]
- Nohesara, S.; Abdolmaleky, H.M.; Thiagalingam, S. Epigenetic aberrations in major psychiatric diseases related to diet and gut microbiome alterations. Genes 2023, 14, 1506. [Google Scholar] [CrossRef]
- Murru, A.; Anmella, G.; Giménez, A.; Vieta, E. Bipolar disorders, obesity, and metabolic disturbances: Mechanisms and implications. In Neurobiology of Bipolar Disorder; Elsevier: Amsterdam, The Netherlands, 2021; pp. 257–274. [Google Scholar]
- Lionaki, E.; Ploumi, C.; Tavernarakis, N. One-carbon metabolism: Pulling the strings behind aging and neurodegeneration. Cells 2022, 11, 214. [Google Scholar] [CrossRef]
- Yu, Y.; Martins, L.M. Mitochondrial One-Carbon Metabolism and Alzheimer’s Disease. Int. J. Mol. Sci. 2024, 25, 6302. [Google Scholar] [CrossRef]
- Frajerman, A.; Urban, M.; Rivollier, F.; Plaze, M.; Chaumette, B.; Krebs, M.-O.; Scoriels, L. Abnormalities in one-carbon metabolism in young patients with psychosis. Front. Psychiatry 2023, 14, 1128890. [Google Scholar] [CrossRef]
- Abdolmaleky, H.M.; Zhou, J.-R.; Thiagalingam, S. Cataloging recent advances in epigenetic alterations in major mental disorders and autism. Epigenomics 2021, 13, 1231–1245. [Google Scholar] [CrossRef]
- Nohesara, S.; Mostafavi Abdolmaleky, H.; Thiagalingam, S. Substance-induced psychiatric disorders, epigenetic and microbiome alterations, and potential for therapeutic interventions. Brain Sci. 2024, 14, 769. [Google Scholar] [CrossRef]
- Tao, K.; Yuan, Y.; Xie, Q.; Dong, Z. Relationship between human oral microbiome dysbiosis and neuropsychiatric diseases: An updated overview. Behav. Brain Res. 2024, 471, 115111. [Google Scholar] [CrossRef]
- Kerstens, R.; Ng, Y.Z.; Pettersson, S.; Jayaraman, A. Balancing the oral–gut–brain axis with diet. Nutrients 2024, 16, 3206. [Google Scholar] [CrossRef]
- Inchingolo, A.D.; Malcangi, G.; Semjonova, A.; Inchingolo, A.M.; Patano, A.; Coloccia, G.; Ceci, S.; Marinelli, G.; Di Pede, C.; Ciocia, A.M. Oralbiotica/oralbiotics: The impact of oral microbiota on dental health and demineralization: A systematic review of the literature. Children 2022, 9, 1014. [Google Scholar] [CrossRef]
- Sun, X.; Li, M.; Xia, L.; Fang, Z.; Yu, S.; Gao, J.; Feng, Q.; Yang, P. Alteration of salivary microbiome in periodontitis with or without type-2 diabetes mellitus and metformin treatment. Sci. Rep. 2020, 10, 15363. [Google Scholar] [CrossRef]
- D’Angelo, E.; Fiori, F.; Ferraro, G.A.; Tessitore, A.; Nazzaro, L.; Serpico, R.; Contaldo, M. Autism Spectrum Disorder, Oral Implications, and Oral Microbiota. Children 2025, 12, 368. [Google Scholar] [CrossRef]
- Kisely, S. Periodontal health and psychiatric disorders. Curr. Oral Health Rep. 2023, 10, 111–116. [Google Scholar] [CrossRef]
- D’ambrosio, F.; Caggiano, M.; Schiavo, L.; Savarese, G.; Carpinelli, L.; Amato, A.; Iandolo, A. Chronic stress and depression in periodontitis and peri-implantitis: A narrative review on neurobiological, neurobehavioral and immune–microbiome interplays and clinical management implications. Dent. J. 2022, 10, 49. [Google Scholar] [CrossRef]
- Gupta, U.; Dey, P. The oral microbial odyssey influencing chronic metabolic disease. Arch. Physiol. Biochem. 2024, 130, 831–847. [Google Scholar] [CrossRef]
- Sun, H.-L.; Feng, Y.; Zhang, Q.; Li, J.-X.; Wang, Y.-Y.; Su, Z.; Cheung, T.; Jackson, T.; Sha, S.; Xiang, Y.-T. The microbiome–gut–brain axis and dementia: A bibliometric analysis. Int. J. Environ. Res. Public Health 2022, 19, 16549. [Google Scholar] [CrossRef]
- Alex, A.M.; Levendosky, A.A.; Bogat, G.A.; Muzik, M.; Nuttall, A.K.; Knickmeyer, R.C.; Lonstein, J.S. Stress and mental health symptoms in early pregnancy are associated with the oral microbiome. BMJ Ment Health 2024, 27, e301100. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Piras, E.; Tamburini, S.; Bu, K.; Wallach, D.S.; Remsen, B.; Cantor, A.; Kong, J.; Goetz, D.; Hoffman, K.W. Gut and oral microbiome modulate molecular and clinical markers of schizophrenia-related symptoms: A transdiagnostic, multilevel pilot study. Psychiatry Res. 2023, 326, 115279. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Meng, H.; Gao, X.; Xu, L.; Feng, X. Effect of non-surgical periodontal treatment on short chain fatty acid levels in gingival crevicular fluid of patients with generalized aggressive periodontitis. J. Periodontal Res. 2014, 49, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Leonov, G.E.; Varaeva, Y.R.; Livantsova, E.N.; Starodubova, A.V. The complicated relationship of short-chain fatty acids and oral microbiome: A narrative review. Biomedicines 2023, 11, 2749. [Google Scholar] [CrossRef]
- Gomes-Filho, I.S.; Oliveira, M.T.; Cruz, S.S.d.; Cerqueira, E.d.M.M.; Trindade, S.C.; Vieira, G.O.; Couto Souza, P.H.; Adan, L.F.F.; Hintz, A.M.; Passos-Soares, J.d.S. Periodontitis is a factor associated with dyslipidemia. Oral Dis. 2022, 28, 813–823. [Google Scholar] [CrossRef]
- Bitencourt, F.V.; Nascimento, G.G.; Costa, S.A.; Orrico, S.R.P.; Ribeiro, C.C.C.; Leite, F.R.M. The role of dyslipidemia in periodontitis. Nutrients 2023, 15, 300. [Google Scholar] [CrossRef]
- Ghitea, T.C. Correlation of periodontal bacteria with chronic inflammation present in patients with metabolic syndrome. Biomedicines 2021, 9, 1709. [Google Scholar] [CrossRef]
- Iwashita, M.; Hayashi, M.; Nishimura, Y.; Yamashita, A. The link between periodontal inflammation and obesity. Curr. Oral Health Rep. 2021, 8, 76–83. [Google Scholar] [CrossRef]
- Li, Z.; Zheng, Z.; Pathak, J.L.; Li, H.; Wu, G.; Xu, S.; Wang, T.; Cheng, H.; Piao, Z.; Jaspers, R.T. Leptin-deficient ob/ob mice exhibit periodontitis phenotype and altered oral microbiome. J. Periodontal Res. 2023, 58, 392–402. [Google Scholar] [CrossRef]
- Sato, K.; Yamazaki, K.; Kato, T.; Nakanishi, Y.; Tsuzuno, T.; Yokoji-Takeuchi, M.; Yamada-Hara, M.; Miura, N.; Okuda, S.; Ohno, H. Obesity-related gut microbiota aggravates alveolar bone destruction in experimental periodontitis through elevation of uric acid. MBio 2021, 12, e0077121. [Google Scholar] [CrossRef]
- Jia, R.; Zhang, Y.; Wang, Z.; Hu, B.; Wang, Z.; Qiao, H. Association between lipid metabolism and periodontitis in obese patients: A cross-sectional study. BMC Endocr. Disord. 2023, 23, 119. [Google Scholar] [CrossRef]
- Wang, M.; Li, L.; Qian, J.; Wang, N.; Bao, J.; Lu, J.; Chen, F.; Li, Y.; Zhang, Y.; Yan, F. Periodontitis salivary microbiota exacerbates nonalcoholic fatty liver disease in high-fat diet-induced obese mice. IScience 2023, 26, 106346. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Minty, M.; Canceill, T.; Loubières, P.; Azalbert, V.; Tercé, F.; Champion, C.; Burcelin, R.; Barthet, P.; Laurencin-Dalicieux, S. Obesity drives an oral microbiota signature of female patients with periodontitis: A pilot study. Diagnostics 2021, 11, 745. [Google Scholar] [CrossRef] [PubMed]
- Lê, S.; Laurencin-Dalicieux, S.; Minty, M.; Assoulant-Anduze, J.; Vinel, A.; Yanat, N.; Loubieres, P.; Azalbert, V.; Diemer, S.; Burcelin, R. Obesity is associated with the severity of periodontal inflammation due to a specific signature of subgingival microbiota. Int. J. Mol. Sci. 2023, 24, 15123. [Google Scholar] [CrossRef]
- Rahman, B.; Al-Marzooq, F.; Saad, H.; Benzina, D.; Al Kawas, S. Dysbiosis of the subgingival microbiome and relation to periodontal disease in association with obesity and overweight. Nutrients 2023, 15, 826. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Wang, Z.; Wang, J.; Su, X.; Yang, J.; Zhang, Q.; Zhang, L. The oral microbiome profile and biomarker in Chinese type 2 diabetes mellitus patients. Endocrine 2020, 68, 564–572. [Google Scholar] [CrossRef]
- Wang, A.; Sun, Y.; Xu, M.; Qin, Q.; Zhu, W.; Xu, Y. The relationship with and effect of oral microbiota on NLRP3 inflammatory pathway in type 2 diabetes mellitus. Arch. Oral Biol. 2023, 155, 105801. [Google Scholar] [CrossRef]
- Guo, X.-j.; Dai, S.-x.; Lou, J.-d.; Ma, X.-x.; Hu, X.-j.; Tu, L.-p.; Cui, J.; Lu, H.; Jiang, T.; Xu, J.-t. Distribution characteristics of oral microbiota and its relationship with intestinal microbiota in patients with type 2 diabetes mellitus. Front. Endocrinol. 2023, 14, 1119201. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qian, F.; Cheng, X.; Wang, D.; Wang, Y.; Pan, Y.; Chen, L.; Wang, W.; Tian, Y. Dysbiosis of oral microbiota and metabolite profiles associated with type 2 diabetes mellitus. Microbiol. Spectr. 2023, 11, e03796-22. [Google Scholar] [CrossRef] [PubMed]
- Prince, Y.; Davison, G.M.; Davids, S.F.; Erasmus, R.T.; Kengne, A.P.; Graham, L.M.; Raghubeer, S.; Matsha, T.E. The relationship between the oral microbiota and metabolic syndrome. Biomedicines 2022, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Dong, T.; Yuan, K.-Y.; Wang, N.-J.; Xia, F.-Z.; Liu, D.; Wang, Z.-M.; Ma, R.; Lu, Y.-L.; Huang, Z.-W. Shifts in the bacterial community of supragingival plaque associated with metabolic-associated fatty liver disease. Front. Cell. Infect. Microbiol. 2020, 10, 581888. [Google Scholar] [CrossRef]
- Takagi, K.; Tamura, Y.; Narita, N.; Komatsu, S.; Yamazaki, S.; Matsumura, A.; Kubota, K.; Matsumiya, T.; Sawada, K.; Nakaji, S. Involvement of Megasphaera in the oral microbiome and dyslipidemia onset: Evidence from a community-based study in Japan. Folia Microbiol. 2025, 1–12, online ahead of print. [Google Scholar] [CrossRef]
- San-Cristobal, R.; Navas-Carretero, S.; Milagro, F.I.; Riezu-Boj, J.I.; Guruceaga, E.; Celis-Morales, C.; Livingstone, K.M.; Brennan, L.; Lovegrove, J.A.; Daniel, H. Gene methylation parallelisms between peripheral blood cells and oral mucosa samples in relation to overweight. J. Physiol. Biochem. 2016, 73, 465–474. [Google Scholar] [CrossRef]
- Kalea, A.; Hoteit, R.; Suvan, J.; Lovering, R.; Palmen, J.; Cooper, J.; Khodiyar, V.; Harrington, Z.; Humphries, S.; D’Aiuto, F. Upregulation of gingival tissue miR-200b in obese periodontitis subjects. J. Dent. Res. 2015, 94, 59S–69S. [Google Scholar] [CrossRef]
- Byun, J.-S.; Lee, H.Y.; Tian, J.; Moon, J.S.; Choi, J.; Lee, S.-H.; Kim, Y.-G.; Yi, H.-S. Effect of salivary exosomal miR-25-3p on periodontitis with insulin resistance. Front. Immunol. 2022, 12, 775046. [Google Scholar]
- Liu, L.; Xiao, Z.; Ding, W.; Wen, C.; Ge, C.; Xu, K.; Cao, S. Relationship between microRNA expression and inflammatory factors in patients with both type 2 diabetes mellitus and periodontal disease. Am. J. Transl. Res. 2022, 14, 6627. [Google Scholar]
- Radović, N.; Nikolić Jakoba, N.; Petrović, N.; Milosavljević, A.; Brković, B.; Roganović, J. Micro RNA-146a and micro RNA-155 as novel crevicular fluid biomarkers for periodontitis in non-diabetic and type 2 diabetic patients. J. Clin. Periodontol. 2018, 45, 663–671. [Google Scholar] [CrossRef]
- Al-Rawi, N.H.; Al-Marzooq, F.; Al-Nuaimi, A.S.; Hachim, M.Y.; Hamoudi, R. Salivary microRNA 155, 146a/b and 203: A pilot study for potentially non-invasive diagnostic biomarkers of periodontitis and diabetes mellitus. PLoS ONE 2020, 15, e0237004. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-Y.; Wang, P.-Y.; Lin, C.-Y.; Hung, S.-C. Association of the oral microbiome with cognitive function among older adults: Nhanes 2011–2012. J. Nutr. Health Aging 2024, 28, 100264. [Google Scholar] [CrossRef]
- Qiao, Y.; Gong, W.; Li, B.; Xu, R.; Wang, M.; Shen, L.; Shi, H.; Li, Y. Oral microbiota changes contribute to autism spectrum disorder in mice. J. Dent. Res. 2022, 101, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.W.-y.; Hau, C.C.-f.; Tong, W.-m.; Watt, R.M.; Yiu, C.K.Y.; Shum, K.K.-m. Alterations of oral microbiota in young children with autism: Unraveling potential biomarkers for early detection. J. Dent. 2025, 152, 105486. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Xu, M.; Xiao, K.; Yu, K. Association between oral microbiome and depression: A population-based study. J. Affect. Disord. 2025, 379, 441–447. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, C.; Shi, C.; Zhai, T.; Zhu, J.; Wei, D.; Shen, J.; Liu, Z.; Jia, K.; Zhao, L. Mechanisms of oral microflora in Parkinson’s disease. Behav. Brain Res. 2024, 474, 115200. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased intestinal permeability correlates with sigmoid mucosa alpha-synuclein staining and endotoxin exposure markers in early Parkinson’s disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef]
- Qian, Y.; Yang, X.; Xu, S.; Wu, C.; Song, Y.; Qin, N.; Chen, S.-D.; Xiao, Q. Alteration of the fecal microbiota in Chinese patients with Parkinson’s disease. Brain Behav. Immun. 2018, 70, 194–202. [Google Scholar]
- Panzarella, V.; Mauceri, R.; Baschi, R.; Maniscalco, L.; Campisi, G.; Monastero, R. Oral health status in subjects with amnestic mild cognitive impairment and Alzheimer’s disease: Data from the Zabut Aging Project. J. Alzheimer’s Dis. 2022, 87, 173–183. [Google Scholar]
- Wan, J.; Fan, H. Oral microbiome and alzheimer’s disease. Microorganisms 2023, 11, 2550. [Google Scholar] [CrossRef]
- Dominy, S.S.; Lynch, C.; Ermini, F.; Benedyk, M.; Marczyk, A.; Konradi, A.; Nguyen, M.; Haditsch, U.; Raha, D.; Griffin, C. Porphyromonas gingivalis in Alzheimer’s disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci. Adv. 2019, 5, eaau3333. [Google Scholar] [CrossRef] [PubMed]
- Haditsch, U.; Roth, T.; Rodriguez, L.; Hancock, S.; Cecere, T.; Nguyen, M.; Arastu-Kapur, S.; Broce, S.; Raha, D.; Lynch, C.C. Alzheimer’s disease-like neurodegeneration in Porphyromonas gingivalis infected neurons with persistent expression of active gingipains. J. Alzheimer’s Dis. 2020, 75, 1361–1376. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.-C.; Tsai, S.-J.; Hsu, J.-W.; Huang, K.-L.; Chen, T.-J.; Chen, M.-H. Risk of periodontitis in adolescents with bipolar disorder: A cohort study of 21,255 subjects. Eur. Child Adolesc. Psychiatry 2024, 33, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Fatima, Z.; Bey, A.; Azmi, S.; Gupta, N.; Khan, A. Mental depression as a risk factor for periodontal disease: A case–control study. Eur. J. Gen. Dent. 2016, 5, 86–89. [Google Scholar] [CrossRef]
- Yolken, R.; Prandovszky, E.; Severance, E.G.; Hatfield, G.; Dickerson, F. The oropharyngeal microbiome is altered in individuals with schizophrenia and mania. Schizophr. Res. 2021, 234, 51–57. [Google Scholar] [CrossRef]
- Ling, Z.; Cheng, Y.; Liu, X.; Yan, X.; Wu, L.; Shao, L.; Gao, J.; Lei, W.; Song, Q.; Zhao, L. Altered oral microbiota and immune dysfunction in Chinese elderly patients with schizophrenia: A cross-sectional study. Transl. Psychiatry 2023, 13, 383. [Google Scholar] [CrossRef]
- Lin, D.; Fu, Z.; Liu, J.; Perrone-Bizzozero, N.; Hutchison, K.E.; Bustillo, J.; Du, Y.; Pearlson, G.; Calhoun, V.D. Association between the oral microbiome and brain resting state connectivity in schizophrenia. Schizophr. Res. 2024, 270, 392–402. [Google Scholar] [CrossRef]
- Huang, H.; Yang, N.; Chen, M.-m.; Chen, X.; Chen, W.; Li, X.; Chen, Y.; Deng, Z.; Zhou, W.; Xu, S.-x. Altered oral health and microbiota in drug-free patients with schizophrenia. BMC Psychiatry 2025, 25, 274. [Google Scholar] [CrossRef]
- Guo, H.; Li, B.; Yao, H.; Liu, D.; Chen, R.; Zhou, S.; Ji, Y.; Zeng, L.; Du, M. Profiling the oral microbiomes in patients with Alzheimer’s disease. Oral Dis. 2023, 29, 1341–1355. [Google Scholar] [CrossRef]
- Na, H.S.; Jung, N.Y.; Song, Y.; Kim, S.Y.; Kim, H.J.; Lee, J.Y.; Chung, J. A distinctive subgingival microbiome in patients with periodontitis and Alzheimer’s disease compared with cognitively unimpaired periodontitis patients. J. Clin. Periodontol. 2024, 51, 43–53. [Google Scholar] [CrossRef]
- Sritana, N.; Phungpinij, A. Analysis of Oral Microbiota in Elderly Thai Patients with Alzheimer’s Disease and Mild Cognitive Impairment. Int. J. Environ. Res. Public Health 2024, 21, 1242. [Google Scholar] [CrossRef]
- Mihaila, D.; Donegan, J.; Barns, S.; LaRocca, D.; Du, Q.; Zheng, D.; Vidal, M.; Neville, C.; Uhlig, R.; Middleton, F.A. The oral microbiome of early stage Parkinson’s disease and its relationship with functional measures of motor and non-motor function. PLoS ONE 2019, 14, e0218252. [Google Scholar] [CrossRef] [PubMed]
- Manghi, P.; Filosi, M.; Zolfo, M.; Casten, L.G.; Garcia-Valiente, A.; Mattevi, S.; Heidrich, V.; Golzato, D.; Perini, S.; Thomas, A.M. Large-scale metagenomic analysis of oral microbiomes reveals markers for autism spectrum disorders. Nat. Commun. 2024, 15, 9743. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, B.; Lapsley, C.; McDowell, A.; Miliotis, G.; McLafferty, M.; O’Neill, S.M.; Coleman, S.; McGinnity, T.M.; Bjourson, A.J.; Murray, E.K. Variations in the oral microbiome are associated with depression in young adults. Sci. Rep. 2021, 11, 15009. [Google Scholar] [CrossRef] [PubMed]
- Agranyoni, O.; Rowley, T.; Johnson, S.B.; Volk, H.; Schleif, W.; Hernandez, R.G.; Klein, L.M.; Yolken, R.H.; Sabunciyan, S. Oral microbiome composition is associated with depressive symptoms during pregnancy. Brain Behav. Immun. Health 2025, 45, 100978. [Google Scholar] [CrossRef]
- Nohesara, S.; Abdolmaleky, H.M.; Dickerson, F.; Pinto-Tomas, A.A.; Jeste, D.V.; Thiagalingam, S. Associations of microbiome pathophysiology with social activity and behavior are mediated by epigenetic modulations: Avenues for designing innovative therapeutic strategies. Neurosci. Biobehav. Rev. 2025, 174, 106208. [Google Scholar] [CrossRef]
- Abdolmaleky, H.M.; Nohesara, S.; Zhou, J.-R.; Thiagalingam, S. Epigenetics in evolution and adaptation to environmental challenges: Pathways for disease prevention and treatment. Epigenomics 2025, 17, 317–333. [Google Scholar] [CrossRef]
- Hernandez Martinez, C.d.J.; Glessner, J.; Finoti, L.S.; Silva, P.F.; Messora, M.; Coletta, R.D.; Hakonarson, H.; Palioto, D.B. Methylome-wide analysis in systemic microbial-induced experimental periodontal disease in mice with different susceptibility. Front. Cell. Infect. Microbiol. 2024, 14, 1369226. [Google Scholar] [CrossRef]
- Takano, M.; Nishihara, R.; Sugano, N.; Matsumoto, K.; Yamada, Y.; Takane, M.; Fujisaki, Y.; Ito, K. The effect of systemic anti-tumor necrosis factor-alpha treatment on Porphyromonas gingivalis infection in type 2 diabetic mice. Arch. Oral Biol. 2010, 55, 379–384. [Google Scholar] [CrossRef]
- Palioto, D.B.; Finoti, L.S.; Kinane, D.F.; Benakanakere, M. Epigenetic and inflammatory events in experimental periodontitis following systemic microbial challenge. J. Clin. Periodontol. 2019, 46, 819–829. [Google Scholar] [CrossRef]
- Niwa, T.; Ushijima, T. Induction of epigenetic alterations by chronic inflammation and its significance on carcinogenesis. Adv. Genet. 2010, 71, 41–56. [Google Scholar]
- Claycombe, K.J.; Brissette, C.A.; Ghribi, O. Epigenetics of inflammation, maternal infection, and nutrition. J. Nutr. 2015, 145, 1109S–1115S. [Google Scholar] [CrossRef] [PubMed]
- Ligthart, S.; Marzi, C.; Aslibekyan, S.; Mendelson, M.M.; Conneely, K.N.; Tanaka, T.; Colicino, E.; Waite, L.L.; Joehanes, R.; Guan, W. DNA methylation signatures of chronic low-grade inflammation are associated with complex diseases. Genome Biol. 2016, 17, 255. [Google Scholar] [CrossRef] [PubMed]
- Lorente-Sorolla, C.; Garcia-Gomez, A.; Català-Moll, F.; Toledano, V.; Ciudad, L.; Avendaño-Ortiz, J.; Maroun-Eid, C.; Martín-Quirós, A.; Martínez-Gallo, M.; Ruiz-Sanmartín, A. Inflammatory cytokines and organ dysfunction associate with the aberrant DNA methylome of monocytes in sepsis. Genome Med. 2019, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Jiao, Y.; Larsson, L.; Almeida, L.; Garaicoa-Pazmino, C.; Le, J.; Squarize, C.; Inohara, N.; Giannobile, W.V.; Castilho, R. Epigenetic modifications of histones in periodontal disease. J. Dent. Res. 2016, 95, 215–222. [Google Scholar] [CrossRef]
- Liaw, A.; Liu, C.; Bartold, M.; Ivanovski, S.; Han, P. Salivary histone deacetylase in periodontal disease: A cross-sectional pilot study. J. Periodontal Res. 2023, 58, 433–443. [Google Scholar] [CrossRef]
- Liaw, A.; Liu, C.; Bartold, M.; Ivanovski, S.; Han, P. Effect of non-surgical periodontal therapy on salivary histone deacetylases expression: A prospective clinical study. J. Clin. Periodontol. 2024, 51, 926–935. [Google Scholar] [CrossRef]
- Görgülü, N.G.; Doğan, B. Effect of non-surgical periodontal treatment on salivary and serum biomarkers in stage III grade B and C periodontitis. J. Periodontol. 2022, 93, 1191–1205. [Google Scholar] [CrossRef]
- Olsen, I.; Singhrao, S.K.; Osmundsen, H. Periodontitis, pathogenesis and progression: miRNA-mediated cellular responses to Porphyromonas gingivalis. J. Oral Microbiol. 2017, 9, 1333396. [Google Scholar] [CrossRef]
- Nayar, G.; Gauna, A.; Chukkapalli, S.; Velsko, I.; Kesavalu, L.; Cha, S. Polymicrobial infection alter inflammatory microRNA in rat salivary glands during periodontal disease. Anaerobe 2016, 38, 70–75. [Google Scholar] [CrossRef]
- Sharma, R.; Frasch, M.G.; Zelgert, C.; Zimmermann, P.; Fabre, B.; Wilson, R.; Waldenberger, M.; MacDonald, J.W.; Bammler, T.K.; Lobmaier, S.M. Maternal–fetal stress and DNA methylation signatures in neonatal saliva: An epigenome-wide association study. Clin. Epigenetics 2022, 14, 87. [Google Scholar] [CrossRef]
- Ragusa, M.; Santagati, M.; Mirabella, F.; Lauretta, G.; Cirnigliaro, M.; Brex, D.; Barbagallo, C.; Domini, C.N.; Gulisano, M.; Barone, R. Potential associations among alteration of salivary miRNAs, saliva microbiome structure, and cognitive impairments in autistic children. Int. J. Mol. Sci. 2020, 21, 6203. [Google Scholar] [CrossRef] [PubMed]
- DiCarlo, G.E.; Mabry, S.J.; Cao, X.; McMillan, C.; Woynaroski, T.G.; Harrison, F.E.; Reddy, I.A.; Matthies, H.J.; Flynn, C.R.; Wallace, M.T. Autism-associated variant in the SLC6A3 gene alters the oral microbiome and metabolism in a murine model. Front. Psychiatry 2021, 12, 655451. [Google Scholar] [CrossRef] [PubMed]
- Mercante, F.; Abbaspour, A.; Pucci, M.; Sabatucci, A.; Rania, M.; Konstantinidou, F.; Gatta, V.; Stuppia, L.; Cifani, C.; Bulik, C.M. Epigenetic alterations and microbiota changes in the saliva of individuals with binge-eating spectrum disorders compared with normal weight healthy controls. Life Sci. 2025, 374, 123695. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, L.; Ren, L.; Zhu, R.; Jiang, Y.; Qiao, Y.; Li, Y. Dysbiosis and Metabolic Dysregulation of Salivary Microbiota in Schizophrenia. J. Multidiscip. Healthc. 2025, 18, 813–825. [Google Scholar] [CrossRef]
- François, M.; Pascovici, D.; Wang, Y.; Vu, T.; Liu, J.-W.; Beale, D.; Hor, M.; Hecker, J.; Faunt, J.; Maddison, J. Saliva Proteome, Metabolome and Microbiome Signatures for Detection of Alzheimer’s Disease. Metabolites 2024, 14, 714. [Google Scholar] [CrossRef]
- Lemon, K.P.; Klepac-Ceraj, V.; Schiffer, H.K.; Brodie, E.L.; Lynch, S.V.; Kolter, R. Comparative analyses of the bacterial microbiota of the human nostril and oropharynx. mBio 2010, 1, e00129-10. [Google Scholar] [CrossRef]
- Hicks, S.D.; Uhlig, R.; Afshari, P.; Williams, J.; Chroneos, M.; Tierney-Aves, C.; Wagner, K.; Middleton, F.A. Oral microbiome activity in children with autism spectrum disorder. Autism Res. 2018, 11, 1286–1299. [Google Scholar] [CrossRef]
- Nohesara, S.; Mostafavi Abdolmaleky, H.; Pirani, A.; Pettinato, G.; Thiagalingam, S. The Obesity–Epigenetics–Microbiome Axis: Strategies for Therapeutic Intervention. Nutrients 2025, 17, 1564. [Google Scholar] [CrossRef]
- Costa, C.F.; Correia-de-Sá, T.; Araujo, R.; Barbosa, F.; Burnet, P.W.; Ferreira-Gomes, J.; Sampaio-Maia, B. The oral-gut microbiota relationship in healthy humans: Identifying shared bacteria between environments and age groups. Front. Microbiol. 2024, 15, 1475159. [Google Scholar] [CrossRef]
- Bao, J.; Li, L.; Zhang, Y.; Wang, M.; Chen, F.; Ge, S.; Chen, B.; Yan, F. Periodontitis may induce gut microbiota dysbiosis via salivary microbiota. Int. J. Oral Sci. 2022, 14, 32. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Jiao, Y.; Gillilland, M.G.; Hayashi, A.; Imai, J.; Sugihara, K.; Miyoshi, M.; Brazil, J.C.; Kuffa, P. The intermucosal connection between the mouth and gut in commensal pathobiont-driven colitis. Cell 2020, 182, 447–462.e414. [Google Scholar] [CrossRef] [PubMed]
- Irie, K.; Azuma, T.; Tomofuji, T.; Yamamoto, T. Exploring the role of IL-17A in oral dysbiosis-associated periodontitis and its correlation with systemic inflammatory disease. Dent. J. 2023, 11, 194. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, P.; Xu, F.; Zheng, Y.; Zhao, H. Advances in the study of IL-17 in neurological diseases and mental disorders. Front. Neurol. 2023, 14, 1284304. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Mao, T.; Fan, J.; Shen, Y.; Xue, L.; Du, K.; Li, Y.; Wang, L.; Wang, X. IL-17A Induces Circadian Disruptions Through the Epigenetic Repression of BMAL1 in Mice With Alzheimer’s Disease. J. Cell. Mol. Med. 2025, 29, e70546. [Google Scholar] [CrossRef]
- Schamarek, I.; Anders, L.; Chakaroun, R.M.; Kovacs, P.; Rohde-Zimmermann, K. The role of the oral microbiome in obesity and metabolic disease: Potential systemic implications and effects on taste perception. Nutr. J. 2023, 22, 28. [Google Scholar] [CrossRef]
- Lê, S.; Cecchin-Albertoni, C.; Thomas, C.; Kemoun, P.; Minty, M.; Blasco-Baque, V. The role of dysbiotic oral microbiota in cardiometabolic diseases: A narrative review. Diagnostics 2023, 13, 3184. [Google Scholar] [CrossRef]
- Al-Roub, A.; Al Madhoun, A.; Akhter, N.; Thomas, R.; Miranda, L.; Jacob, T.; Al-Ozairi, E.; Al-Mulla, F.; Sindhu, S.; Ahmad, R. IL-1β and TNFα cooperativity in regulating IL-6 expression in adipocytes depends on CREB binding and H3K14 acetylation. Cells 2021, 10, 3228. [Google Scholar] [CrossRef]
- Suárez, L.J.; Garzón, H.; Arboleda, S.; Rodríguez, A. Oral dysbiosis and autoimmunity: From local periodontal responses to an imbalanced systemic immunity. A review. Front. Immunol. 2020, 11, 591255. [Google Scholar] [CrossRef]
- Luo, H.; Wu, B.; Kamer, A.R.; Adhikari, S.; Sloan, F.; Plassman, B.L.; Tan, C.; Qi, X.; Schwartz, M.D. Oral health, diabetes, and inflammation: Effects of oral hygiene behaviour. Int. Dent. J. 2022, 72, 484–490. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, Y.; Chen, Y.; Yu, L.; Zhou, J.; Wang, N.; Liu, T.; Fu, C. Associations of oral hygiene with incident hypertension and type 2 diabetes mellitus: A population based cohort study in Southwest China. J. Clin. Hypertens. 2022, 24, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Avcu, N.; Avcu, F.; Beyan, C.; Ural, A.U.; ad Kaptan, K.; Özyurt, M.; Nevruz, O.; Yalçin, A. The relationship between gastric-oral Helicobacter pylori and oral hygiene in patients with vitamin B12–deficiency anemia. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2001, 92, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Ge, J.; Pan, Q.; Hu, N.; Hua, F. Salivary microbiome variations in type 2 diabetes mellitus patients with different stages of periodontitis. BMC Oral Health 2024, 24, 1424. [Google Scholar] [CrossRef] [PubMed]
- Ebersole, J.L.; Kirakodu, S.S.; Zhang, X.; Dawson, D., III; Miller, C.S. Salivary microbiome and biomarker characteristics of diabetics with periodontitis. Mol. Oral Microbiol. 2025, 40, 37–49. [Google Scholar] [CrossRef]
- Qin, H.; Li, G.; Xu, X.; Zhang, C.; Zhong, W.; Xu, S.; Yin, Y.; Song, J. The role of oral microbiome in periodontitis under diabetes mellitus. J. Oral Microbiol. 2022, 14, 2078031. [Google Scholar] [CrossRef]
- Li, H.; Zou, Y.; Ding, G. Dietary Factors Associated with Dental Erosion: A Meta-Analysis. PLoS ONE 2012, 7, e42626. [Google Scholar] [CrossRef]
- Jagelavičienė, E.; Vaitkevičienė, I.; Šilingaitė, D.; Šinkūnaitė, E.; Daugėlaitė, G. The relationship between vitamin D and periodontal pathology. Medicina 2018, 54, 45. [Google Scholar] [CrossRef]
- Graells, J.; Ojeda, R.M.; Muniesa, C.; Gonzalez, J.; Saavedra, J. Glossitis with linear lesions: An early sign of vitamin B12 deficiency. J. Am. Acad. Dermatol. 2009, 60, 498–500. [Google Scholar] [CrossRef]
- Ramsey, D.; Muskin, P.R. Vitamin deficiencies and mental health: How are they linked. Curr. Psychiatry 2013, 12, 37–43. [Google Scholar]
- Sultana, O.F.; Hia, R.A.; Reddy, P.H. A combinational therapy for preventing and delaying the onset of alzheimer’s disease: A focus on probiotic and vitamin co-supplementation. Antioxidants 2024, 13, 202. [Google Scholar] [CrossRef]
- Wang, S.-D.; Wang, X.; Zhao, Y.; Xue, B.-H.; Wang, X.-T.; Chen, Y.-X.; Zhang, Z.-Q.; Tian, Y.-R.; Xie, F.; Qian, L.-J. Homocysteine-induced disturbances in DNA methylation contribute to development of stress-associated cognitive decline in rats. Neurosci. Bull. 2022, 38, 887–900. [Google Scholar] [CrossRef]
- Holmes, H.E.; Valentin, R.E.; Jernerén, F.; de Jager Loots, C.A.; Refsum, H.; Smith, A.D.; Guarente, L.; Dellinger, R.W.; Sampson, D.; Initiative, A.s.D.N. Elevated homocysteine is associated with increased rates of epigenetic aging in a population with mild cognitive impairment. Aging Cell 2024, 23, e14255. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Kuo, H.K.; Lai, Y.L. The association between serum folate levels and periodontal disease in older adults: Data from the National Health and Nutrition Examination Survey 2001/02. J. Am. Geriatr. Soc. 2007, 55, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Roffman, J.L.; Lamberti, J.S.; Achtyes, E.; Macklin, E.A.; Galendez, G.C.; Raeke, L.H.; Silverstein, N.J.; Smoller, J.W.; Hill, M.; Goff, D.C. Randomized multicenter investigation of folate plus vitamin B12 supplementation in schizophrenia. JAMA Psychiatry 2013, 70, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Saxena, P.; Kumar, S.; Vaibhav, K. Role of Folic acid as adjuvant treatment in Schizophrenia: A randomized controlled trial. Eur. Psychiatry 2023, 66, S634–S635. [Google Scholar]
- An, Y.; Feng, L.; Zhang, X.; Wang, Y.; Wang, Y.; Tao, L.; Qin, Z.; Xiao, R. Dietary intakes and biomarker patterns of folate, vitamin B6, and vitamin B12 can be associated with cognitive impairment by hypermethylation of redox-related genes NUDT15 and TXNRD1. Clin. Epigenetics 2019, 11, 139. [Google Scholar] [CrossRef]
- Buzatu, R.; Luca, M.M.; Bumbu, B.A. Does Vitamin C Supplementation Provide a Protective Effect in Periodontal Health? A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2024, 25, 8598. [Google Scholar] [CrossRef]
- Guo, H.; Ding, J.; Liu, Q.; Li, Y.; Liang, J.; Zhang, Y. Vitamin C and metabolic syndrome: A meta-analysis of observational studies. Front. Nutr. 2021, 8, 728880. [Google Scholar] [CrossRef]
- Plevin, D.; Galletly, C. The neuropsychiatric effects of vitamin C deficiency: A systematic review. BMC Psychiatry 2020, 20, 315. [Google Scholar] [CrossRef]
- Blaschke, K.; Ebata, K.T.; Karimi, M.M.; Zepeda-Martínez, J.A.; Goyal, P.; Mahapatra, S.; Tam, A.; Laird, D.J.; Hirst, M.; Rao, A. Vitamin C induces Tet-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature 2013, 500, 222–226. [Google Scholar] [CrossRef]
- Liang, F.; Zhou, Y.; Zhang, Z.; Zhang, Z.; Shen, J. Association of vitamin D in individuals with periodontitis: An updated systematic review and meta-analysis. BMC Oral Health 2023, 23, 387. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Kim, J. Serum vitamin D status and metabolic syndrome: A systematic review and dose-response meta-analysis. Nutr. Res. Pract. 2021, 15, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Chai, B.; Gao, F.; Wu, R.; Dong, T.; Gu, C.; Lin, Q.; Zhang, Y. Vitamin D deficiency as a risk factor for dementia and Alzheimer’s disease: An updated meta-analysis. BMC Neurol. 2019, 19, 284. [Google Scholar] [CrossRef] [PubMed]
- Wilczynski, K.M.; Checinska, K.; Kulczyk, K.; Janas-Kozik, M. Vitamin D deficiency and depressive symptoms: Meta-analysis of studies. Psychiatr. Pol. 2022, 56, 1327–1344. [Google Scholar]
- Zhu, J.-l.; Luo, W.-w.; Cheng, X.; Li, Y.; Zhang, Q.-z.; Peng, W.-x. Vitamin D deficiency and schizophrenia in adults: A systematic review and meta-analysis of observational studies. Psychiatry Res. 2020, 288, 112959. [Google Scholar] [CrossRef]
- Jiang, X.; Xia, L.; Tang, T.; Fan, X.; Wang, R.; Wang, M.; Yang, W.; Yan, J.; Qi, K.; Li, P. Decreased vitamin D bio-availability with altered DNA methylation of its metabolism genes in association with the metabolic disorders among the school-aged children with degree I, II, and III obesity. J. Nutr. Biochem. 2024, 129, 109627. [Google Scholar] [CrossRef]
- Xue, J.; Schoenrock, S.A.; Valdar, W.; Tarantino, L.M.; Ideraabdullah, F.Y. Maternal vitamin D depletion alters DNA methylation at imprinted loci in multiple generations. Clin. Epigenetics 2016, 8, 107. [Google Scholar] [CrossRef]
- Fischbach, M.A. Microbiome: Focus on causation and mechanism. Cell 2018, 174, 785–790. [Google Scholar] [CrossRef]
- Walter, J.; Armet, A.M.; Finlay, B.B.; Shanahan, F. Establishing or exaggerating causality for the gut microbiome: Lessons from human microbiota-associated rodents. Cell 2020, 180, 221–232. [Google Scholar] [CrossRef]
- Homayouni Rad, A.; Pourjafar, H.; Mirzakhani, E. A comprehensive review of the application of probiotics and postbiotics in oral health. Front. Cell. Infect. Microbiol. 2023, 13, 1120995. [Google Scholar] [CrossRef]
- Luo, S.-C.; Wei, S.-M.; Luo, X.-T.; Yang, Q.-Q.; Wong, K.-H.; Cheung, P.C.; Zhang, B.-B. How probiotics, prebiotics, synbiotics, and postbiotics prevent dental caries: An oral microbiota perspective. npj Biofilms Microbiomes 2024, 10, 14. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Metabolic Disorder | Type of Study/Number of Participants | Microbiota Testing Method/Relevant Pathway or Oral Disease | Key Findings and Affected Oral Bacteria | Ref. |
|---|---|---|---|---|
| Obesity | Human/9 healthy normal weight and 10 classed as obese | 16S rRNA gene sequening/periodontal inflammation | Higher abundance of the Capnocytophaga genus (2.47% vs. 0.27%) in subjects classed as obese vs. normal weight | [48] |
| Obesity | Human/34 healthy normal weight and 11 classed as obese | 16S rRNA gene sequening/periodontal inflammation | Reduction in OM diversity and an elevation in periodontal inflammation; greater abundance of Staphylococcaceae (1.13% vs. 0.48%) in periodontal microbiota in subjects classed as obese vs. normal weight | [49] |
| Obesity | Human/25 classed as obese or overweight vs. 25 of normal weight | 16S rRNA gene sequencing/moderate–severe periodontitis | Increased abundance of periopathogens, including Aggregatibacter and actinomycetemcomitans in subjects with obesity | [50] |
| Type 2 diabetes (T2DM) | Human/280 patients with T2DM and 162 healthy controls | 16S rRNA gene sequencing/increased risk of periodontitis | Increased Neisseria (14.12%), Streptococcus, Haemophilus, and Pseudomonas genera, decreased Acinetobacteria, and elevated Firmicutes/Bacteroidetes ratio (7.6% vs. 2.74%) in T2DM vs. controls | [51] |
| T2DM | Human/15 patients with T2DM vs. controls | 16S rRNA gene sequencing/inflammation | Reduced saliva Fusobacteriota and Campilobacterota and increased Proteobacteria abundance in T2DM, triggering the NLRP3 inflammasome pathway | [52] |
| T2DM | Human/183 patients with type 2 diabetes and 74 controls | 16S rRNA gene sequencing/inflammatory mediators of oral and intestinal flora | Higher oral Streptococcus, Actinobacteria, Rothia, Cetobacterium, and intestinal Bifidobacterium, Streptococcus, and Blautia correlate with T2DM; upregulated glycine betaine degradation pathway in oral and intestinal flora | [53] |
| T2DM | Human/10 patients with type 2 diabetes and 10 controls | Metagenomic sequencing/OM dysbiosis in T2DM and increased risk of oral diseases | Increased periodontal pathogens like P. gingivalis and Prevotella melaninogenica and harmful salivary metabolites such as cadaverine and L-(+)-leucine | [54] |
| Metabolic syndrome (MetS) | Human/128 subgingival plaque samples from participants with and without MetS | Metagenomic and 16S rRNA gene sequencing/inflammation | Increased abundance of Actinomyces dentalis, Actinomyces naeslundii, Actinomyces viscosus, Corynebacterium matruchotii, Leptotrichia buccalis, and Streptococcus sanguinis in MetS | [55] |
| Metabolic-associated fatty liver disease (MAFLD) | Human/24 patients with MAFLD and 22 healthy controls | 16S rRNA gene sequencing/chronic low-grade inflammation | Increased Actinomyces and Prevotella 2 spp. in supragingival plaques in MAFLD; insulin resistance correlates with the abundance of Granulicatella, Veillonella, Streptococcus, and Scardovia spp., and obesity with Streptococcus, Olsenella, Scardovia, and Selenomonas spp. | [56] |
| Dyslipidemia | Human/763 tongue coating samples | 16S rRNA gene sequencing/lipid metabolism | Increased relative abundance of Megasphaera in dyslipidemia; dyslipidemia is linked to the abundance of Veillonella, Atopobium, Stomatobaculum, Tanneralla, and Megasphaera | [57] |
| Neuropsychiatric Disorders | Study Type/Samples/Microbiota Testing Method | Oral Microbiota Changes in Patients | Effects on the Disease State | Ref. |
|---|---|---|---|---|
| Schizophrenia (SCZ) and mania | Human/throat swab/16S rRNA gene sequencing | Decreased Neisseria subflava, Weeksellaceae, and Prevotella and increased Streptococci | Correlation between Neisseria subflava and cognitive performance | [77] |
| SCZ | Human/swabs from the middle site of the tongue/Illumina MiSeq sequencing | Decreased Prevotella and Veillonella; increased Streptococcus and Fusobacterium | OM composition affects peripheral inflammatory cytokines | [78] |
| SCZ | Human/saliva/16S rRNA gene sequencing | Increased in some Gram-positive (Actinomyces, Rothia, Atopobium, Streptococcus) and Gram-negative bacteria (Prevotella, Leptotrichia Porphyromonas, Lautropia, and Capnocytophaga) | OM dysbiosis is associated with brain functional connectivity changes | [79] |
| SCZ | Human/saliva/16S rRNA gene sequencing | Decreased Actinobacteriota in patients | A poor oral environment is associated with altered OM in SCZ | [80] |
| Alzheimer’s disease (AD) | Human/saliva and periodontal samples/16S rRNA gene sequencing | Streptococcus oralis and Porphyromonas gingivalis are the predominant salivary and periodontal bacteria, respectively | Association between altered OM and cognition in AD | [81] |
| AD vs. cognitively unimpaired patients with periodontitis | Human/buccal, supragingival, and subgingival plaque samples/16S rRNA gene sequencing | Increased Atopobium rimae, Dialister pneumosintes, Olsenella sp. HMT 807, Saccharibacteria (TM7) sp. HMT 348, and several species of Prevotella in AD | Direct association between periodontitis caused by OM dysbiosis and greater cognitive decline | [82] |
| AD | Human/saliva/16S rRNA gene sequencing | Increased Fusobacteriota and Peptostreptococcaceae and decreased Veillonella vs. the MCI and controls | Association between abnormal immune responses and inflammatory processes and OM changes in AD | [83] |
| Parkinson’s disease (PD) | Human/saliva/shotgun metatranscriptomic profiling | Decrease in a bacteriophage (Streptococcus phage PhiSpn 200), and increase in three yeast species (Candida albicans, Candida dubliniensis, and Saccharomyces cerevisiae) in PD | Significant alterations in several indices of motor, cognitive, and sensory function | [84] |
| Autism spectrum disorders (ASDs) | Human/saliva/metagenomic approach | Increased Actinomyces hongkongensis, Actinomyces johnsonii, Cutibacterium acnes, the Eikenella species NML 130454, and Rothia dentocariosa | Association between the OM composition and cognitive impairments in ASDs | [85] |
| Depression | Human/saliva/16S rRNA gene sequencing | Increased Neisseria spp. and Prevotella nigrescens | OM dysbiosis is associated with depression | [86] |
| Depression | Human/saliva/16S rRNA gene sequencing | Decreased Neisseria genus | Negative correlation between Neisseria genus and pro-inflammatory cytokines | [87] |
| Experiment | Epigenetic Changes | Immunoassays/Outcome | Ref. |
|---|---|---|---|
| Orally administered P. gingivalis in mice to induce periodontitis | Methylation of PI3K/Akt1, Ctnnb1, and Hsp90aa1 | ELISA/increases inflammation (TNF signaling), which is reversed by TNF-α inhibition | [90,91] |
| P. gingivalis gavage in mice, which induces periodontitis | Increases DNMT3b expression, likely due to “repercussion” | Immunofluorescence staining and Procarta Multiplex Cytokine Kit/increases inflammatory markers BTLA and IL-18R1 | [92] |
| Monocytes exposed to LPS or monocytes of patients with sepsis | DNA methylation changes related to inflammatory genes | The cytometric bead array and ELISA/alterations in IL-10 and IL-6 levels | [96] |
| Oral dysbiosis in vivo and oral epithelial cells exposure to LPS in vitro | Induces histone modifications | Immunohistochemistry, immunofluorescence, and immunoblotting/activates the transcriptional coactivators p300/CBP, and promotes NF-κB accumulation | [97] |
| Clinical gingivitis | Downregulates HDAC4, HDAC8, and HDAC10 in saliva in gingivitis, and HDAC4, HDAC6, HDAC8, and HDAC9 in periodontitis | ELISA plate reader/3–6 months non-surgical periodontal therapy improved HDACs changes; decreased salivary MMP-8 and MAF (inflammatory) and increased serum T-SOD (antioxidant) levels | [98,99,100] |
| P. gingivalis infection in THP-1 macrophages or P. gingivalis LPS | Upregulates miR-132 via TLR2/4 and NF-κB signaling and miR-146a | RT-PCR and ELISA/reduces IRAK1/TRAF6 expression and increases IL-1β, IL-6, and TNF-α secretion | [101] |
| In vivo polymicrobial periodontal infection in ApoE−/− mice | Induces miR-146a in periodontal tissues and the spleen | RT-PCR/reduces IRAK1/TRAF6 expression and increases IL-1β, IL-6, and TNF-α secretion [101] | [102] |
| Tannerella abundance in saliva in ASD | Increases salivary miR-141-3p expression | Increases cognitive dysfunction | [104] |
| Increased salivary Bacilli and reduced Lachnospirales in eating disorder | Upregulation of exosomal let-7b-5p, mir-15b-5p, mir-429, and mir-221-3p, and hypomethylation of DAT1 | RT-PCR/increases dopamine transporter (DAT1) gene expression affecting dopamine signaling | [106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mostafavi, S.; Nohesara, S.; Pirani, A.; Mostafavi Abdolmaleky, H.; Thiagalingam, S. Oral–Gut Microbiota Crosstalk and Epigenetic Targets in Metabolic and Neuropsychiatric Diseases. Nutrients 2025, 17, 3367. https://doi.org/10.3390/nu17213367
Mostafavi S, Nohesara S, Pirani A, Mostafavi Abdolmaleky H, Thiagalingam S. Oral–Gut Microbiota Crosstalk and Epigenetic Targets in Metabolic and Neuropsychiatric Diseases. Nutrients. 2025; 17(21):3367. https://doi.org/10.3390/nu17213367
Chicago/Turabian StyleMostafavi, Sahar, Shabnam Nohesara, Ahmad Pirani, Hamid Mostafavi Abdolmaleky, and Sam Thiagalingam. 2025. "Oral–Gut Microbiota Crosstalk and Epigenetic Targets in Metabolic and Neuropsychiatric Diseases" Nutrients 17, no. 21: 3367. https://doi.org/10.3390/nu17213367
APA StyleMostafavi, S., Nohesara, S., Pirani, A., Mostafavi Abdolmaleky, H., & Thiagalingam, S. (2025). Oral–Gut Microbiota Crosstalk and Epigenetic Targets in Metabolic and Neuropsychiatric Diseases. Nutrients, 17(21), 3367. https://doi.org/10.3390/nu17213367