
Role of Plasma-Derived Exosomal MicroRNAs in Mediating Type 2 Diabetes Remission

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Anthropometric and Biochemical Assessments
2.3. Blood Collection, Exosome Purification, and Exosome Characterization
2.4. Small RNA Isolation and Sequencing
2.5. Statistical Analysis
3. Results
3.1. Characteristics of the Participants
3.2. Characterization of Plasma-Derived Exosomes and Small RNA Quality Assessment
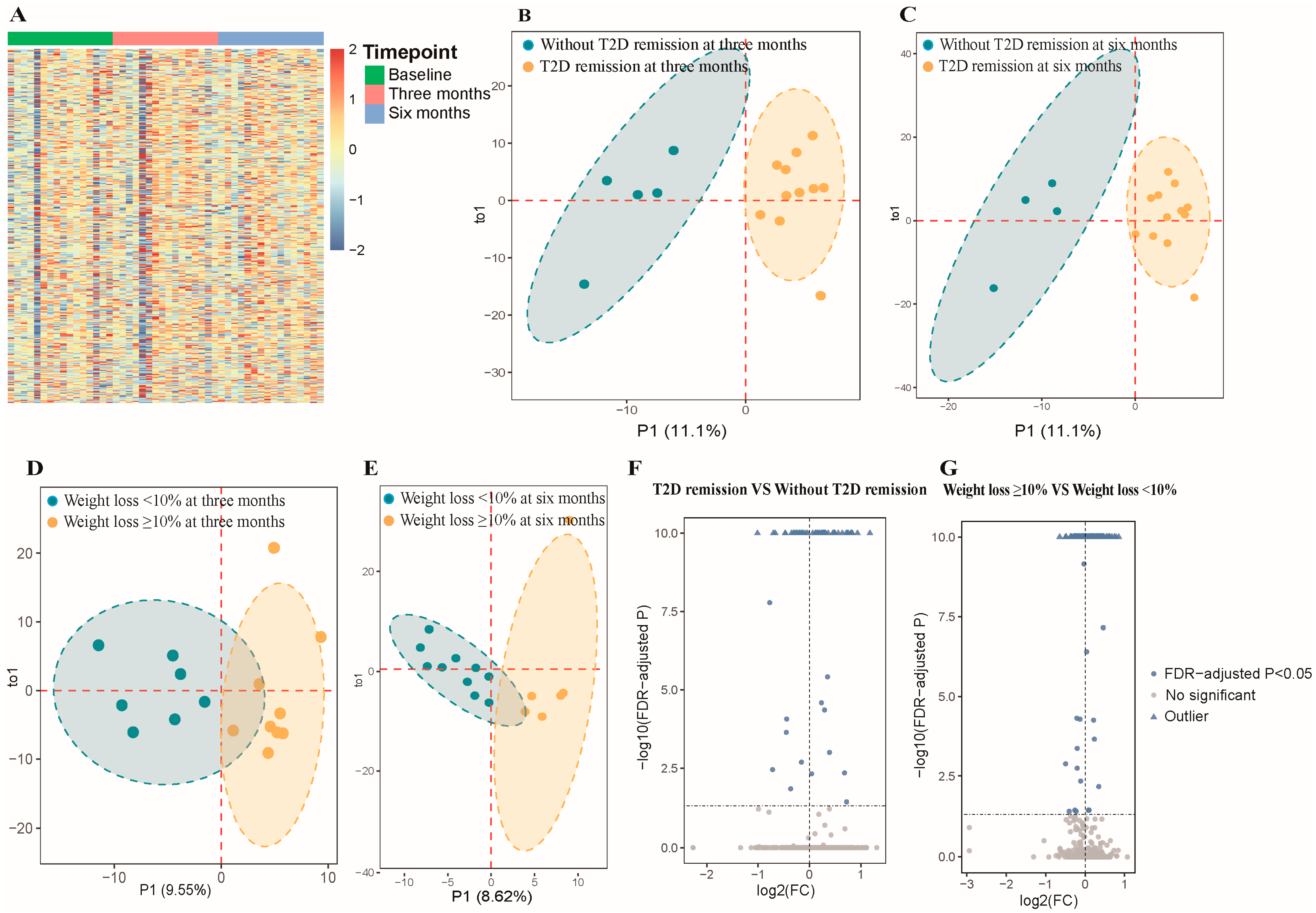
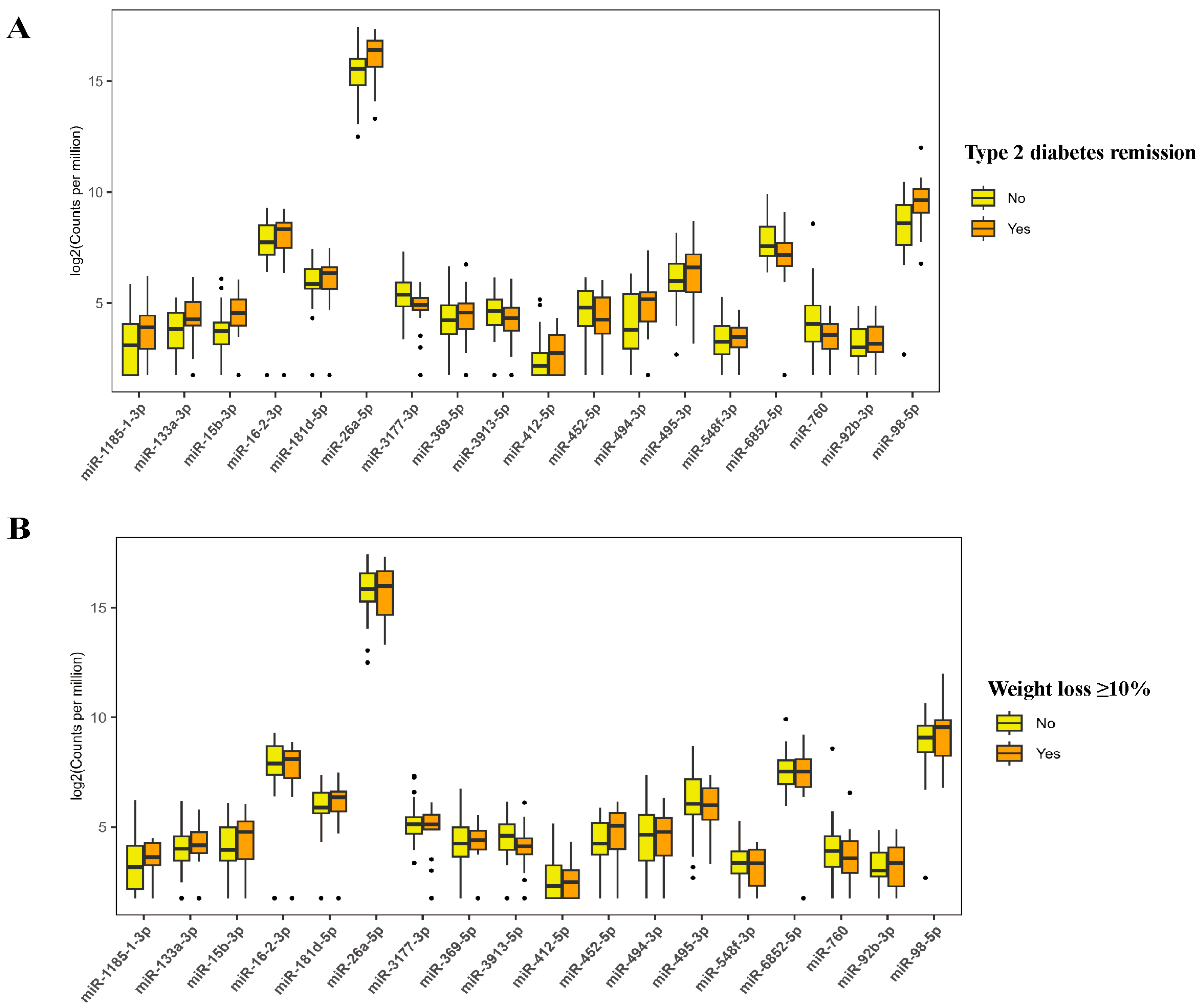
3.3. Differentially Expressed Plasma-Derived Exosomal miRNAs
3.4. Pathway Enrichment Analysis
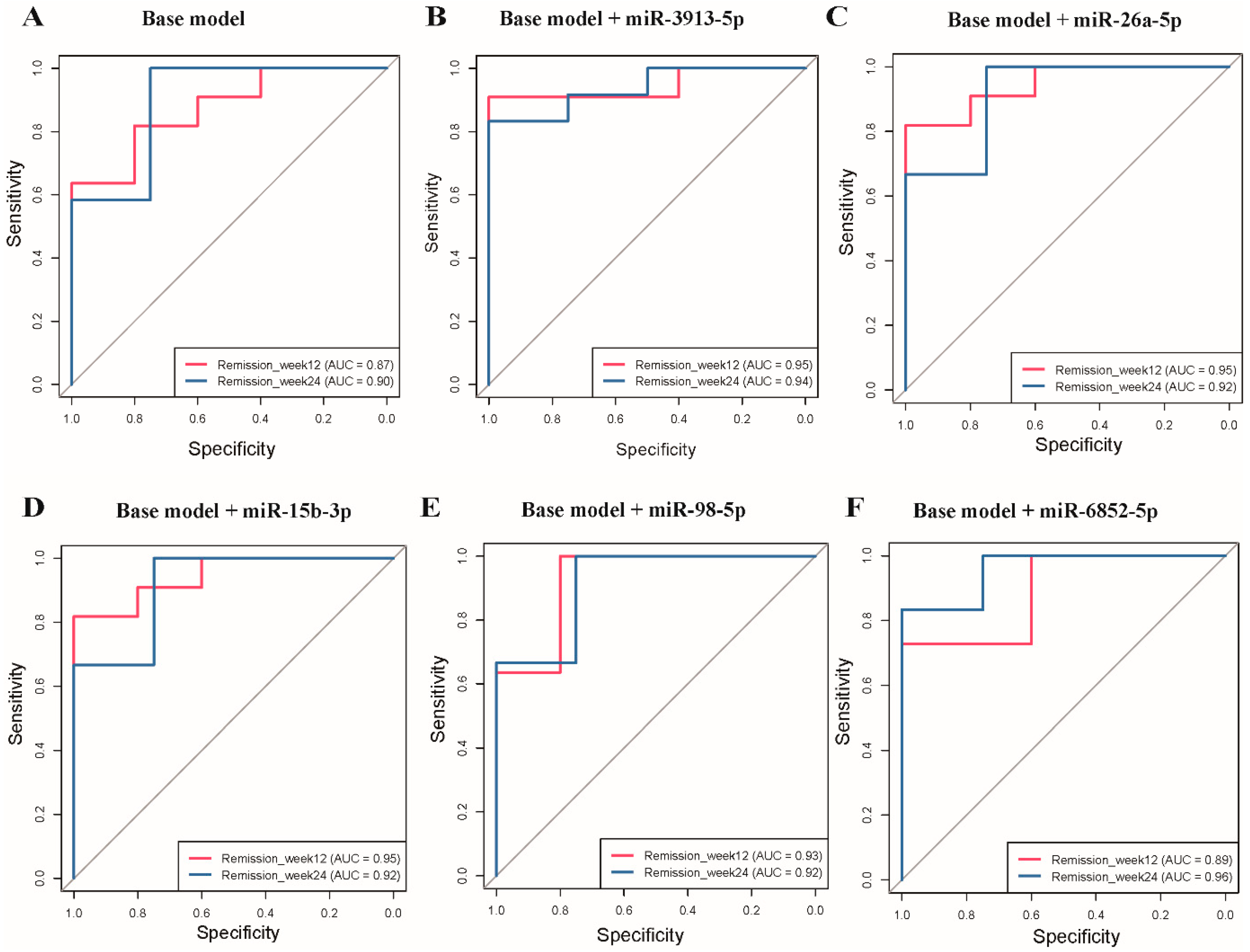
3.5. Receiver Operating Characteristic Curve Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diabetes. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 18 September 2024).
- Wang, L.; Li, X.; Wang, Z.; Bancks, M.P.; Carnethon, M.R.; Greenland, P.; Zhong, V.W. Trends in Prevalence of Diabetes and Control of Risk Factors in Diabetes Among US Adults, 1999–2018. JAMA 2021, 326, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sattar, N.; Welsh, P.; Leslie, W.S.; Thom, G.; McCombie, L.; Brosnahan, N.; Lean, M.E. Dietary weight-management for type 2 diabetes remissions in south asians: The south asian diabetes remission randomised trial for proof-of-concept and feasibility (STANDby). Lancet Reg. Health-Southeast Asia 2023, 9, 100111. [Google Scholar] [CrossRef] [PubMed]
- Lean, M.E.; Leslie, W.S.; Barnes, A.C.; Brosnahan, N.; Thom, G.; McCombie, L.; Taylor, R. Primary care-led weight management for remission of type 2 diabetes (DiRECT): An open-label, cluster-randomised trial. Lancet 2018, 391, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Feng, N.; Wang, S.; Liu, Y.; Wang, J.; Tan, Y.; Zhong, V.W. Low-calorie diets and remission of type 2 diabetes in Chinese: Phenotypic changes and individual variability. Nutr. J. 2025, 24, 42. [Google Scholar] [CrossRef]
- He, X.; Kuang, G.; Wu, Y.; Ou, C. Emerging roles of exosomal miRNAs in diabetes mellitus. Clin. Transl. Med. 2021, 11, e468. [Google Scholar] [CrossRef]
- Fluitt, M.B.; Mohit, N.; Gambhir, K.K.; Nunlee-Bland, G. To the future: The role of exosome-derived microRNAs as markers, mediators, and therapies for endothelial dysfunction in type 2 diabetes mellitus. J. Diabetes Res. 2022, 2022, 5126968. [Google Scholar] [CrossRef]
- Jafari, N.; Llevenes, P.; Denis, G.V. Exosomes as novel biomarkers in metabolic disease and obesity-related cancers. Nat. Rev. Endocrinol. 2022, 18, 327–328. [Google Scholar] [CrossRef]
- Hernández-Gómez, K.G.; Avila-Nava, A.; González-Salazar, L.E.; Noriega, L.G.; Serralde-Zúñiga, A.E.; Guizar-Heredia, R.; Guevara-Cruz, M. Modulation of MicroRNAs and Exosomal MicroRNAs after Dietary Interventions for Obesity and Insulin Resistance: A Narrative Review. Metabolites 2023, 13, 1190. [Google Scholar] [CrossRef]
- Palmer, J.D.; Soule, B.P.; Simone, B.A.; Zaorsky, N.G.; Jin, L.; Simone, N.L. MicroRNA expression altered by diet: Can food be medicinal? Ageing Res. Rev. 2014, 17, 16–24. [Google Scholar] [CrossRef]
- Taheri, S. Type 2 diabetes remission: A new mission in diabetes care. Diabetes Care 2023, 47, 47–49. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.; Shen, Y.; Shi, S.; Tang, T. miREvo: An integrative microRNA evolutionary analysis platform for next-generation sequencing experiments. BMC Bioinf. 2012, 13, 140. [Google Scholar] [CrossRef] [PubMed]
- miRTargetLink 2.0—Interactive miRNA Target Gene and Target Pathway Networks|Nucleic Acids Research|Oxford Academic. Available online: https://academic.oup.com/nar/article/49/W1/W409/6261786?login=false (accessed on 19 September 2024).
- Ke, R.Q.; Wang, Y.; Hong, S.H.; Xiao, L.X. Anti-diabetic effect of quercetin in type 2 diabetes mellitus by regulating the microRNA-92b-3p/EGR1 axis. J. Physiol. Pharmacol. 2023, 74, 149–158. [Google Scholar] [CrossRef]
- Yang, F.; Zhao, S.; Zhang, X.; Ding, S.; Xu, Y. RNF6 Targeted by miR-26a-5p Protects Pancreatic β-Cell Function Against Type 2 Diabetes. Diabetes Metab. Syndr. Obes. 2022, 15, 93–102. [Google Scholar] [CrossRef]
- Oliveira, M.S.; Rheinheimer, J.; Moehlecke, M.; Rodrigues, M.; Assmann, T.S.; Leitao, C.B.; de Souza, B.M. UCP2, IL18, and miR-133a-3p are dysregulated in subcutaneous adipose tissue of patients with obesity. Mol. Cell. Endocrinol. 2020, 509, 110805. [Google Scholar] [CrossRef]
- Du, G.; Cheng, X.; Zhang, Z.; Han, L.; Wu, K.; Li, Y.; Lin, X. TGF-Beta Induced Key Genes of Osteogenic and Adipogenic Differentiation in Human Mesenchymal Stem Cells and MiRNA–mRNA Regulatory Networks. Front. Genet. 2021, 12, 759596. [Google Scholar] [CrossRef]
- Mu, N.; Zhang, T.; Zhu, Y.; Lu, B.; Zheng, Q.; Duan, J. The mechanism by which miR-494-3p regulates PGC1-α-mediated inhibition of mitophagy in cardiomyocytes and alleviation of myocardial ischemia—Reperfusion injury. BMC Cardiovasc. Disord. 2023, 23, 204. [Google Scholar] [CrossRef]
- Rovira-Llopis, S.; Bañuls, C.; Diaz-Morales, N.; Hernandez-Mijares, A.; Rocha, M.; Victor, V.M. Mitochondrial dynamics in type 2 diabetes: Pathophysiological implications. Redox Biol. 2017, 11, 637–645. [Google Scholar] [CrossRef]
- Kumar, A.; Supowit, S.; Potts, J.D.; DiPette, D.J. Alpha-calcitonin gene-related peptide prevents pressure-overload induced heart failure: Role of apoptosis and oxidative stress. Physiol. Rep. 2019, 7, e14269. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.H. Effect of miR-412-5p-loaded exosomes in H9c2 cardiomyocytes via the MAPK pathway. Iran. J. Basic. Med. Sci. 2024, 27, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Corsello, T.; Kudlicki, A.S.; Garofalo, R.P.; Casola, A. Cigarette smoke condensate exposure changes RNA content of extracellular vesicles released from small airway epithelial cells. Cells 2019, 8, 1652. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [PubMed]
- Benchoula, K.; Arya, A.; Parhar, I.S.; Hwa, W.E. FoxO1 signaling as a therapeutic target for type 2 diabetes and obesity. Eur. J. Pharmacol. 2021, 891, 173758. [Google Scholar] [CrossRef]
- Jiang, Y.; Yan, F.; Feng, Z.; Lazarovici, P.; Zheng, W. Signaling Network of Forkhead Family of Transcription Factors (FOXO) in Dietary Restriction. Cells 2020, 9, 100. [Google Scholar] [CrossRef]
- Costa, I.; Lima, M.; Medeiros, A.; Bezerra, L.; Santos, P.; Serquiz, A.; Morais, A.H. An insulin receptor-binding multifunctional protein from tamarindus indica L. Presents a hypoglycemic effect in a diet-induced type 2 diabetes—Preclinical study. Foods 2022, 11, 2207. [Google Scholar] [CrossRef]
- Su, T.; Hou, J.; Liu, T.; Dai, P.; Qin, L.; Ding, L.; Guo, X. MiR-34a-5p and miR-452-5p: The novel regulators of pancreatic endocrine dysfunction in diabetic zucker rats? Int. J. Med. Sci. 2021, 18, 3171. [Google Scholar] [CrossRef]
- Jang, C.; Oh, S.F.; Wada, S.; Rowe, G.C.; Liu, L.; Chan, M.C.; Arany, Z. A branched-chain amino acid metabolite drives vascular fatty acid transport and causes insulin resistance. Nat. Med. 2016, 22, 421–426. [Google Scholar] [CrossRef]
- Peng, Z.; Aggarwal, R.; Zeng, N.; He, L.; Stiles, E.X.; Debebe, A.; Stiles, B.L. KT1 regulates endoplasmic reticulum stress and mediates the adaptive response of pancreatic β cells. Mol. Cell. Biol. 2020, 40, e00031-20. [Google Scholar] [CrossRef]
- Swolin-Eide, D.; Forsander, G.; Pundziute Lyckå, A.; Novak, D.; Grillari, J.; Diendorfer, A.B.; Magnusson, P. Circulating microRNAs in young individuals with long-duration type 1 diabetes in comparison with healthy controls. Sci. Rep. 2023, 13, 11634. [Google Scholar] [CrossRef]
- Arora, S.; Singh, P.; Tabassum, G.; Dohare, R.; Syed, M.A. miR-495–3p regulates sphingolipid metabolic reprogramming to induce Sphk1/ceramide mediated mitophagy and apoptosis in NSCLC. Free Radic. Biol. Med. 2022, 189, 71–84. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Zhang, Z.; Zhang, X.; Zhu, Y.; Zhang, C.; Bi, Y. Extracellular vesicles mediate the communication of adipose tissue with brain and promote cognitive impairment associated with insulin resistance. Cell Metab. 2022, 34, 1264–1279.e8. [Google Scholar] [CrossRef]
- Zhang, T.; Li, L.; Shang, Q.; Lv, C.; Wang, C.; Su, B. Circulating miR-126 is a potential biomarker to predict the onset of type 2 diabetes mellitus in susceptible individuals. Biochem. Biophys. Res. Commun. 2015, 463, 60–63. [Google Scholar] [CrossRef]
- Jiménez-Lucena, R.; Camargo, A.; Alcalá-Diaz, J.F.; Romero-Baldonado, C.; Luque, R.M.; van Ommen, B.; López-Miranda, J.A. plasma circulating miRNAs profile predicts type 2 diabetes mellitus and prediabetes: From the CORDIOPREV study. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef]
- Wojciechowska, G.; Szczerbinski, L.; Kretowski, M.; Niemira, M.; Hady, H.R.; Kretowski, A. Exploring microRNAs as predictive biomarkers for type 2 diabetes mellitus remission after sleeve gastrectomy: A pilot study. Obesity 2022, 30, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chen, X.; He, C.; Chen, Z.; Chen, Q.; Chen, J.; Bo, H. Sanhuang xiexin decoction synergizes insulin/PI3K-akt/FoxO signaling pathway to inhibit hepatic glucose production and alleviate T2DM. J. Ethnopharmacol. 2023, 306, 116162. [Google Scholar] [CrossRef]
- Prattichizzo, F.; Matacchione, G.; Giuliani, A.; Sabbatinelli, J.; Olivieri, F.; de Candia, P.; Ceriello, A. Extracellular vesicle-shuttled miRNAs: A critical appraisal of their potential as nano-diagnostics and nano-therapeutics in type 2 diabetes mellitus and its cardiovascular complications. Theranostics 2021, 11, 1031–1045. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Baseline | Six Months | ||||
|---|---|---|---|---|---|---|
| Overall | Overall | Weight Loss ≥ 10% | Weight Loss < 10% | Type 2 Diabetes Remission | Without Type 2 Diabetes Remission | |
| N | 16 | 16 | 6 | 10 | 12 | 4 |
| Age, mean (SD), year | 37.3 (8.0) | 37.3 (8.0) | 40.7 (4.6) | 35.3 (9.1) | 37.0 (7.6) | 38.3 (10.5) |
| Men, n (%) | 12 (75.0) | 12 (75.0) | 4 (66.6) | 8 (80.0) | 11 (91.7) | 1 (25.0) |
| Duration of T2D, mean (SD), year | 2.5 (2.1) | 2.5 (2.1) | 1.8 (1.7) | 2.9 (2.3) | 2.2 (2.4) | 3.5 (0.8) |
| Family history of T2D, n (%) | 8 (50.0) | 8 (50.0) | 5 (83.3) | 3 (30.0) | 6 (50.0) | 2 (50.0) |
| Body weight, mean (SD), kg | 92.4 (26.7) | 82.4 (24.1) | 70.3 (11.3) | 89.6 (27.2) | 84.8 (26.6) | 75.0 (14.5) |
| Body mass index, mean (SD), kg/m2 | 31.2 (6.9) | 27.8 (6.7) | 24.1 (2.5) | 30.0 (7.6) | 28.1 (7.5) | 26.9 (4.4) |
| Body fat mass, mean (SD), kg | 32.3 (16.9) | 23.4 (14.7) | 15.8 (5.9) | 28.5 (16.9) | 23.8 (16.3) | 22.0 (6.3) |
| Body fat percentage, % | 33.6 (9.8) | 26.9 (9.6) | 22.9 (8.8) | 29.5 (9.6) | 25.9 (9.9) | 30.7 (8.5) |
| Body muscle mass, mean (SD), kg | 55.9 (13.3) | 55.1 (12.6) | 51.2 (12.2) | 57.7 (12.8) | 57.2 (11.9) | 46.6 (14.0) |
| Body muscle percentage, % | 61.9 (9.3) | 68.4 (9.7) | 72.6 (9.9) | 65.6 (8.9) | 69.5 (10.1) | 63.9 (7.4) |
| Trunk fat mass, mean (SD), kg | 18.0 (9.1) | 12.9 (8.4) | 8.4 (3.9) | 15.9 (9.4) | 13.3 (9.3) | 11.5 (3.5) |
| Fasting blood glucose, mean (SD), mmol/L | 7.2 (1.7) | 6.2 (1.9) | 5.3 (0.7) | 6.8 (2.2) | 5.5 (0.7) | 8.6 (2.6) |
| Hemoglobin A1c, mean (SD), % | 8.0 (1.8) | 6.2 (1.0) | 5.8 (0.7) | 6.5 (1.1) | 5.8 (0.4) | 7.6 (1.0) |
| HOMA2-IR | 2.5 (1.0) | 2.0 (0.8) | 1.2 (0.3) | 2.4 (0.5) | 2.0 (0.8) | 1.8 (0.9) |
| HOMA2-%β | 100.7 (51.2) | 108.7 (49.8) | 101.9 (36.7) | 112.7 (57.8) | 127.7 (41.3) | 51.6 (18.9) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Shi, S.; Jiang, X.; Yang, G.; Wu, D.; Li, K.; Zhong, V.W.; Du, X. Role of Plasma-Derived Exosomal MicroRNAs in Mediating Type 2 Diabetes Remission. Nutrients 2025, 17, 2450. https://doi.org/10.3390/nu17152450
Wang S, Shi S, Jiang X, Yang G, Wu D, Li K, Zhong VW, Du X. Role of Plasma-Derived Exosomal MicroRNAs in Mediating Type 2 Diabetes Remission. Nutrients. 2025; 17(15):2450. https://doi.org/10.3390/nu17152450
Chicago/Turabian StyleWang, Sujing, Shuxiao Shi, Xuanwei Jiang, Guangrui Yang, Deshan Wu, Kexin Li, Victor W. Zhong, and Xihao Du. 2025. "Role of Plasma-Derived Exosomal MicroRNAs in Mediating Type 2 Diabetes Remission" Nutrients 17, no. 15: 2450. https://doi.org/10.3390/nu17152450
APA StyleWang, S., Shi, S., Jiang, X., Yang, G., Wu, D., Li, K., Zhong, V. W., & Du, X. (2025). Role of Plasma-Derived Exosomal MicroRNAs in Mediating Type 2 Diabetes Remission. Nutrients, 17(15), 2450. https://doi.org/10.3390/nu17152450