
Main Dietary Patterns for Healthy Aging and Well-Being

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Methods
3. Aging and Healthy Aging
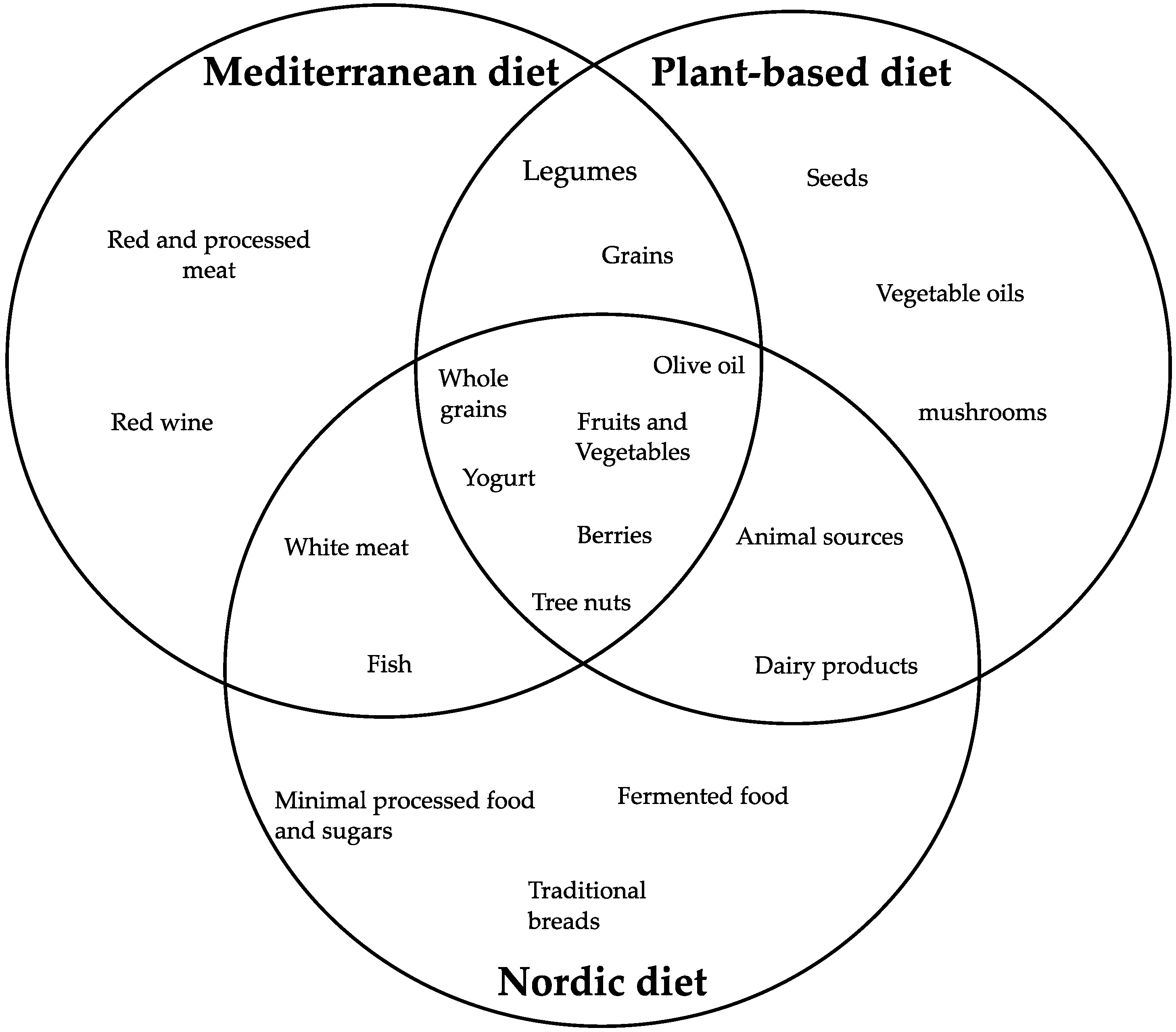
4. Main Dietary Patterns for Healthy Aging
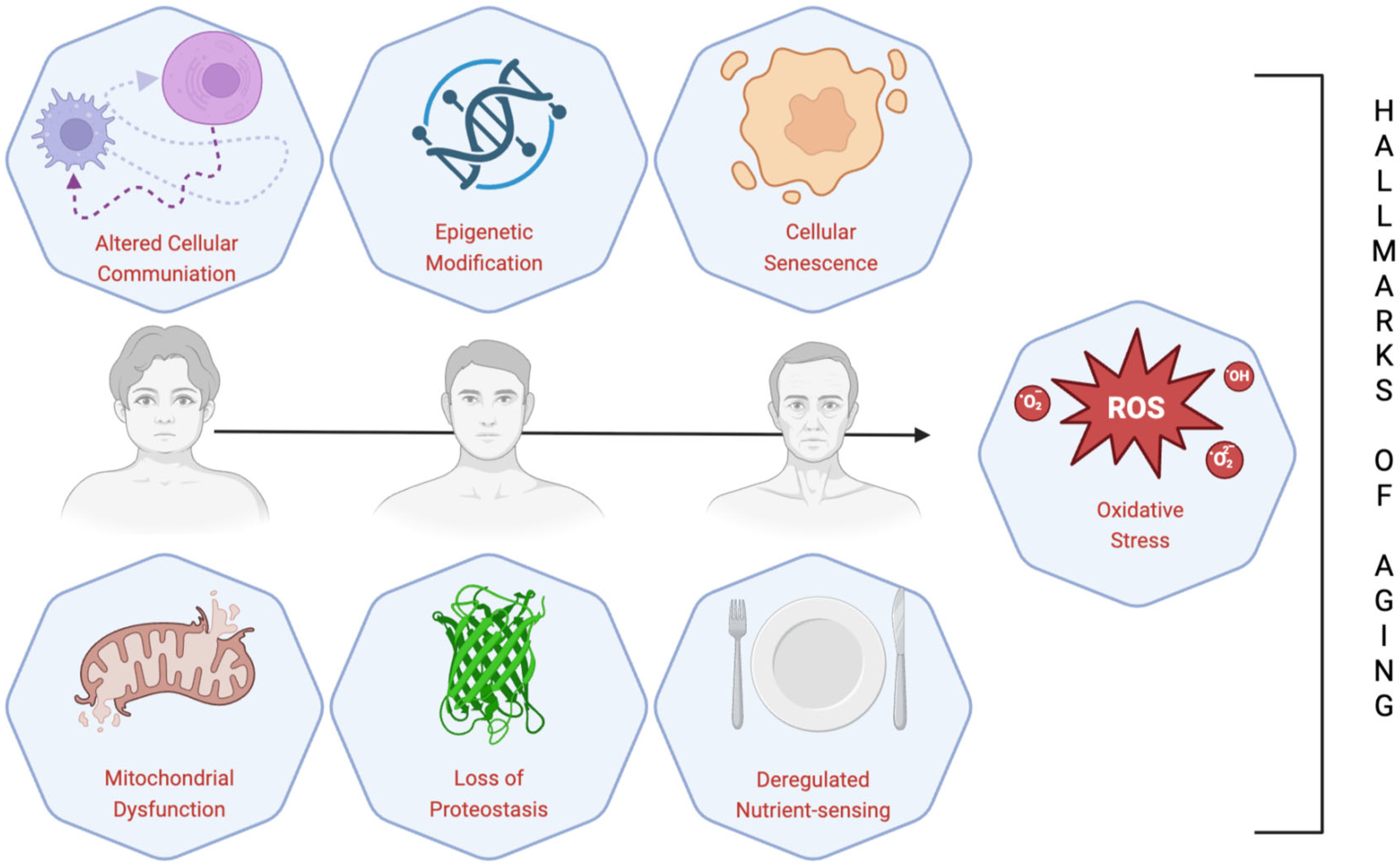
5. Aliment and Dietary Patterns Versus Hallmarks of Aging
5.1. Mediterranean Diet
5.2. Effects of Olive Oil Against Aging
5.3. Effects of Avocado Against Aging
6. Plant-Based Dietary Pattern
6.1. Effects of Fruits Against Aging
6.2. Effects of Dark Chocolate Against Aging
6.3. Effects of Yogurt Against Aging
7. Nordic Dietary Pattern
7.1. Effects of Whole Grains Against Aging
7.2. Effects of Tree Nuts Against Aging
8. Dietary Supplements from Diets
8.1. Effects of Citrus Peels Against Aging
8.2. Peanut Peel Extract Against Aging
9. Strengths and Limitations
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mitchell, E.; Walker, R. Global ageing: Successes, challenges and opportunities. Br. J. Hosp. Med. 2020, 81, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Alcazar, J.; Aagaard, P.; Haddock, B.; Kamper, R.S.; Hansen, S.K.; Prescott, E.; Alegre, L.M.; Frandsen, U.; Suetta, C. Age-and sex-specific changes in lower-limb muscle power throughout the lifespan. J. Gerontol. Ser. A 2020, 75, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, P.M.; Blackwell, T.; Cummings, S.R.; Orwoll, E.S.; Duchowny, K.A.; Kado, D.M.; Stone, K.L.; Ensrud, K.E.; Cauley, J.A.; Evans, W.J. Muscle mass assessed by the D3-creatine dilution method and incident self-reported disability and mortality in a prospective observational study of community-dwelling older men. J. Gerontol. Ser. A 2021, 76, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.S.; Gray, H.A.; Schache, A.G.; Hoermann, R.; Bicknell, J.; Joon, D.L.; Zajac, J.D.; Pandy, M.G.; Grossmann, M. Biomechanical leg muscle function during stair ambulation in men receiving androgen deprivation therapy. J. Gerontol. Ser. A 2020, 75, 1715–1722. [Google Scholar] [CrossRef]
- Ackermans, T.; Francksen, N.; Lees, C.; Papatzika, F.; Arampatzis, A.; Baltzopoulos, V.; Lisboa, P.; Hollands, M.; O’Brien, T.; Maganaris, C. Prediction of balance perturbations and falls on stairs in older people using a biomechanical profiling approach: A 12-month longitudinal study. J. Gerontol. Ser. A 2021, 76, 638–646. [Google Scholar] [CrossRef]
- Agrawal, Y.; Merfeld, D.M.; Horak, F.B.; Redfern, M.S.; Manor, B.; Westlake, K.P.; Holstein, G.R.; Smith, P.F.; Bhatt, T.; Bohnen, N.I. Aging, vestibular function, and balance: Proceedings of a national institute on aging/national institute on deafness and other communication disorders workshop. J. Gerontol. Ser. A 2020, 75, 2471–2480. [Google Scholar] [CrossRef]
- Battilana, F.; Steurer, S.; Rizzi, G.; Delgado, A.C.; Tan, K.R.; Handschin, C. Exercise-linked improvement in age-associated loss of balance is associated with increased vestibular input to motor neurons. Aging Cell 2020, 19, e13274. [Google Scholar] [CrossRef]
- Fekete, M.; Fazekas-Pongor, V.; Balazs, P.; Tarantini, S.; Nemeth, A.N.; Varga, J.T. Role of new digital technologies and telemedicine in pulmonary rehabilitation: Smart devices in the treatment of chronic respiratory diseases. Wien. Klin. Wochenschr. 2021, 133, 1201–1207. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Rudderow, A.L.; Sossong, A.M.; Lewis, K.N.; Younts, C.; Pearson, K.J.; Bernier, M.; de Cabo, R. Perinatal diet influences health and survival in a mouse model of leukemia. Geroscience 2020, 42, 1147–1155. [Google Scholar] [CrossRef]
- Ayo-Martin, O.; García-García, J.; Hernández-Fernández, F.; Gómez-Hontanilla, M.; Gómez-Fernández, I.; Andrés-Fernández, C.; Lamas, C.; Alfaro-Martínez, J.J.; Botella, F.; Segura, T. Cerebral hemodynamics in obesity: Relationship with sex, age, and adipokines in a cohort-based study. Geroscience 2021, 43, 1465–1479. [Google Scholar] [CrossRef]
- Kuzuya, M. Nutritional status related to poor health outcomes in older people: Which is better, obese or lean? Geriatr. Gerontol. Int. 2021, 21, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Allmeroth, K.; Hartman, M.D.; Purrio, M.; Mesaros, A.; Denzel, M.S. Hexosamine pathway activation improves memory but does not extend lifespan in mice. Aging Cell 2022, 21, e13711. [Google Scholar] [CrossRef]
- Anic, K.; Schmidt, M.W.; Furtado, L.; Weidenbach, L.; Battista, M.J.; Schmidt, M.; Schwab, R.; Brenner, W.; Ruckes, C.; Lotz, J. Intermittent fasting—Short-and long-term quality of life, fatigue, and safety in healthy volunteers: A prospective, clinical trial. Nutrients 2022, 14, 4216. [Google Scholar] [CrossRef] [PubMed]
- Duque, G.; Al Saedi, A.; Rivas, D.; Miard, S.; Ferland, G.; Picard, F.; Gaudreau, P. Differential effects of long-term caloric restriction and dietary protein source on bone and marrow fat of the aging rat. J. Gerontol. Ser. A 2020, 75, 2031–2036. [Google Scholar] [CrossRef]
- Lauretti, E.; Nenov, M.; Dincer, O.; Iuliano, L.; Praticò, D. Extra virgin olive oil improves synaptic activity, short-term plasticity, memory, and neuropathology in a tauopathy model. Aging Cell 2020, 19, e13076. [Google Scholar] [CrossRef]
- Ros, M.; Carrascosa, J.M. Current nutritional and pharmacological anti-aging interventions. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1866, 165612. [Google Scholar] [CrossRef]
- Rudnicka, E.; Napierała, P.; Podfigurna, A.; Męczekalski, B.; Smolarczyk, R.; Grymowicz, M. The World Health Organization (WHO) approach to healthy ageing. Maturitas 2020, 139, 6–11. [Google Scholar] [CrossRef]
- Zhou, M.; Zhao, G.; Zeng, Y.; Zhu, J.; Cheng, F.; Liang, W. Aging and cardiovascular disease: Current status and challenges. Rev. Cardiovasc. Med. 2022, 23, 135. [Google Scholar] [CrossRef] [PubMed]
- Juttukonda, M.R.; Donahue, M.J. Neuroimaging of vascular reserve in patients with cerebrovascular diseases. Neuroimage 2019, 187, 192–208. [Google Scholar] [CrossRef]
- O’Brien, M.S.; McDougall, J.J. Age and frailty as risk factors for the development of osteoarthritis. Mech. Ageing Dev. 2019, 180, 21–28. [Google Scholar] [CrossRef]
- Wang, X.; Hu, J.; Wu, D. Risk factors for frailty in older adults. Medicine 2022, 101, e30169. [Google Scholar] [CrossRef] [PubMed]
- Berardelli, I.; Belvisi, D.; Nardella, A.; Falcone, G.; Lamis, D.A.; Fabbrini, G.; Berardelli, A.; Girardi, P.; Pompili, M. Suicide in Parkinson’s disease: A systematic review. CNS Neurol. Disord. Drug Targets (Former. Curr. Drug Targets-CNS Neurol. Disord.) 2019, 18, 466–477. [Google Scholar] [CrossRef]
- Heavener, K.S.; Bradshaw, E.M. The aging immune system in Alzheimer’s and Parkinson’s diseases. In Proceedings of Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2022; pp. 649–657. [Google Scholar]
- Chatsirisupachai, K.; Lagger, C.; de Magalhães, J.P. Age-associated differences in the cancer molecular landscape. Trends Cancer 2022, 8, 962–971. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef]
- World Health, A. Decade of Healthy Ageing: The Global Strategy and Action Plan on Ageing and Health 2016–2020: Towards a World in Which Everyone can Live a Long and Healthy Life: Report by the Director-General; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Jivraj, S.; Goodman, A.; Pongiglione, B.; Ploubidis, G.B. Living longer but not necessarily healthier: The joint progress of health and mortality in the working-age population of England. Popul. Stud. 2020, 74, 399–414. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Wen, J.; Phau, I.; Ying, T.; Aston, J.; Wang, W. The role of tourism in healthy aging: An interdisciplinary literature review and conceptual model. J. Hosp. Tour. Manag. 2023, 56, 356–366. [Google Scholar] [CrossRef]
- Hu, F.B. Diet strategies for promoting healthy aging and longevity: An epidemiological perspective. J. Intern. Med. 2024, 295, 508–531. [Google Scholar] [CrossRef]
- Banerjee, A.; Mukherjee, S.; Maji, B.K. Worldwide flavor enhancer monosodium glutamate combined with high lipid diet provokes metabolic alterations and systemic anomalies: An overview. Toxicol. Rep. 2021, 8, 938–961. [Google Scholar] [CrossRef]
- Mehar, P.; Bera, R.; Swarnim, S.; Mishra, D. Composition of Common Junk Food Items and Their Contribution to the Dietary Requirement of Children and Adolescents. Indian Pediatr. 2023, 60, 221–223. [Google Scholar] [CrossRef]
- Singh, S.A.; Dhanasekaran, D.; Ganamurali, N.; LP, S.S. Junk food-induced obesity- a growing threat to youngsters during the pandemic. Obes. Med. 2021, 26, 100364. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.A.; El Moshy, S.; Rady, D.; Radwan, I.A.; Abbass, M.M.; Al Jawaldeh, A. The effect of unhealthy dietary habits on the incidence of dental caries and overweight/obesity among Egyptian school children (A cross-sectional study). Front. Public Health 2022, 10, 953545. [Google Scholar] [CrossRef] [PubMed]
- Efe, N.; Dawson, P. A review: Sugar-based confectionery and the importance of ingredients. Eur. J. Agric. Food Sci. 2022, 4, 1–8. [Google Scholar] [CrossRef]
- Dominguez, L.J.; Veronese, N.; Baiamonte, E.; Guarrera, M.; Parisi, A.; Ruffolo, C.; Tagliaferri, F.; Barbagallo, M. Healthy Aging and Dietary Patterns. Nutrients 2022, 14, 889. [Google Scholar] [CrossRef] [PubMed]
- Gush, L.; Shah, S.; Gilani, F. Macronutrients and micronutrients. In A Prescription for Healthy Living; Elsevier: Amsterdam, The Netherlands, 2021; pp. 255–273. [Google Scholar]
- Hoti, G.; Matencio, A.; Rubin Pedrazzo, A.; Cecone, C.; Appleton, S.L.; Khazaei Monfared, Y.; Caldera, F.; Trotta, F. Nutraceutical Concepts and Dextrin-Based Delivery Systems. Int. J. Mol. Sci. 2022, 23, 4102. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Beltrán-Velasco, A.I.; Redondo-Flórez, L.; Martín-Rodríguez, A.; Tornero-Aguilera, J.F. Global Impacts of Western Diet and Its Effects on Metabolism and Health: A Narrative Review. Nutrients 2023, 15, 2749. [Google Scholar] [CrossRef]
- Cena, H.; Calder, P.C. Defining a healthy diet: Evidence for the role of contemporary dietary patterns in health and disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef]
- Muscogiuri, G.; Verde, L.; Sulu, C.; Katsiki, N.; Hassapidou, M.; Frias-Toral, E.; Cucalón, G.; Pazderska, A.; Yumuk, V.D.; Colao, A. Mediterranean diet and obesity-related disorders: What is the evidence? Curr. Obes. Rep. 2022, 11, 287–304. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Willett, W.C. The Mediterranean diet and health: A comprehensive overview. J. Intern. Med. 2021, 290, 549–566. [Google Scholar] [CrossRef]
- Lemonakis, N.; Mougios, V.; Halabalaki, M.; Dagla, I.; Tsarbopoulos, A.; Skaltsounis, A.-L.; Gikas, E. Effect of Supplementation with Olive Leaf Extract Enriched with Oleuropein on the Metabolome and Redox Status of Athletes’ Blood and Urine—A Metabolomic Approach. Metabolites 2022, 12, 195. [Google Scholar] [CrossRef]
- Perrone, P.; D’Angelo, S. Extra virgin olive oil as a functional food for athletes: Recovery, health, and performance. J. Phys. Educ. Sport 2025, 25, 370–381. [Google Scholar]
- d’Angelo, S.; Cusano, P. Who Practices Sports can Be Vegetarian? Universidad de Alicante: Alicante, Spain, 2020. [Google Scholar]
- Notariale, R.; Perrone, P.; Mele, L.; Lettieri, G.; Piscopo, M.; Manna, C. Olive oil phenols prevent mercury-induced phosphatidylserine exposure and morphological changes in human erythrocytes regardless of their different scavenging activity. Int. J. Mol. Sci. 2022, 23, 5693. [Google Scholar] [CrossRef] [PubMed]
- Tabanez, M.; Santos, I.R.; Ikebara, J.M.; Camargo, M.L.; Dos Santos, B.A.; Freire, B.M.; Batista, B.L.; Takada, S.H.; Squitti, R.; Kihara, A.H. The Impact of Hydroxytyrosol on the Metallomic-Profile in an Animal Model of Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 14950. [Google Scholar] [CrossRef]
- Perrone, P.; Notariale, R.; Lettieri, G.; Mele, L.; La Pietra, V.; Piscopo, M.; Manna, C. Protective effects of olive oil antioxidant phenols on mercury-induced phosphatidylserine externalization in erythrocyte membrane: Insights into scramblase and flippase activity. Free. Radic. Biol. Med. 2025, 227, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Siddique, A.B.; Kilgore, P.C.; Tajmim, A.; Singh, S.S.; Meyer, S.A.; Jois, S.D.; Cvek, U.; Trutschl, M.; Sayed, K.A.E. (−)-Oleocanthal as a dual c-MET-COX2 inhibitor for the control of lung cancer. Nutrients 2020, 12, 1749. [Google Scholar] [CrossRef] [PubMed]
- Ahamad, J.; Toufeeq, I.; Khan, M.A.; Ameen, M.S.M.; Anwer, E.T.; Uthirapathy, S.; Mir, S.R.; Ahmad, J. Oleuropein: A natural antioxidant molecule in the treatment of metabolic syndrome. Phytother. Res. 2019, 33, 3112–3128. [Google Scholar] [CrossRef] [PubMed]
- Rishmawi, S.; Haddad, F.; Dokmak, G.; Karaman, R. A comprehensive review on the anti-cancer effects of oleuropein. Life 2022, 12, 1140. [Google Scholar] [CrossRef]
- Wilson, D.M., III; Cookson, M.R.; Van Den Bosch, L.; Zetterberg, H.; Holtzman, D.M.; Dewachter, I. Hallmarks of neurodegenerative diseases. Cell 2023, 186, 693–714. [Google Scholar] [CrossRef]
- Grewal, R.; Reutzel, M.; Dilberger, B.; Hein, H.; Zotzel, J.; Marx, S.; Tretzel, J.; Sarafeddinov, A.; Fuchs, C.; Eckert, G.P. Purified oleocanthal and ligstroside protect against mitochondrial dysfunction in models of early Alzheimer’s disease and brain ageing. Exp. Neurol. 2020, 328, 113248. [Google Scholar] [CrossRef]
- Chen, X.; Ji, Y.; Liu, R.; Zhu, X.; Wang, K.; Yang, X.; Liu, B.; Gao, Z.; Huang, Y.; Shen, Y. Mitochondrial dysfunction: Roles in skeletal muscle atrophy. J. Transl. Med. 2023, 21, 503. [Google Scholar] [CrossRef]
- Dambrova, M.; Makrecka-Kuka, M.; Kuka, J.; Vilskersts, R.; Nordberg, D.; Attwood, M.M.; Smesny, S.; Sen, Z.D.; Guo, A.C.; Oler, E. Acylcarnitines: Nomenclature, biomarkers, therapeutic potential, drug targets, and clinical trials. Pharmacol. Rev. 2022, 74, 506–551. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Muñoz-Pinto, M.F.; Cano, M. Irisin enhances longevity by boosting SIRT1, AMPK, autophagy and telomerase. Expert Rev. Mol. Med. 2023, 25, e4. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Li, H.; Liao, P.; Chen, L.; Pan, Y.; Zheng, Y.; Zhang, C.; Liu, D.; Zheng, M.; Gao, J. Mitochondrial dysfunction: Mechanisms and advances in therapy. Signal Transduct. Target. Ther. 2024, 9, 124. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Benítez, M.; Calderón-Fernández, A.; Canales-Cortés, S.; Alegre-Cortés, E.; Uribe-Carretero, E.; Paredes-Barquero, M.; Gimenez-Bejarano, A.; Duque González, G.; Gómez-Suaga, P.; Ortega-Vidal, J.; et al. Biological effects of olive oil phenolic compounds on mitochondria. Mol. Cell Oncol. 2022, 9, 2044263. [Google Scholar] [CrossRef]
- Cárceles Rodríguez, B.; Durán Zuazo, V.H.; Franco Tarifa, D.; Cuadros Tavira, S.; Sacristan, P.C.; García-Tejero, I.F. Irrigation Alternatives for Avocado (Persea americana Mill.) in the Mediterranean Subtropical Region in the Context of Climate Change: A Review. Agriculture 2023, 13, 1049. [Google Scholar] [CrossRef]
- Carracedo, M.G.; Alonso, S.B.; Cabrera, R.S.B.; Jiménez-Arias, D.; Pérez Pérez, J.A. Development of Retrotransposon-Based Molecular Markers for Characterization of Persea americana (Avocado) Cultivars and Horticultural Races. Agronomy 2022, 12, 1510. [Google Scholar] [CrossRef]
- Torres-Isidro, O.; González-Montoya, M.; Vargas-Vargas, M.A.; Florian-Rodriguez, U.; García-Berumen, C.I.; Montoya-Pérez, R.; Saavedra-Molina, A.; Calderón-Cortés, E.; Rodríguez-Orozco, A.R.; Cortés-Rojo, C. Anti-Aging Potential of Avocado Oil via Its Antioxidant Effects. Pharmaceuticals 2025, 18, 246. [Google Scholar] [CrossRef]
- Maiese, K. Targeting the core of neurodegeneration: FoxO, mTOR, and SIRT1. Neural Regen. Res. 2021, 16, 448–455. [Google Scholar] [CrossRef]
- Zhong, Q.; Xiao, X.; Qiu, Y.; Xu, Z.; Chen, C.; Chong, B.; Zhao, X.; Hai, S.; Li, S.; An, Z. Protein posttranslational modifications in health and diseases: Functions, regulatory mechanisms, and therapeutic implications. MedComm 2023, 4, e261. [Google Scholar] [CrossRef]
- Igarashi, M.; Iwasa, K.; Hayakawa, T.; Tsuduki, T.; Kimura, I.; Maruyama, K.; Yoshikawa, K. Dietary oleic acid contributes to the regulation of food intake through the synthesis of intestinal oleoylethanolamide. Front. Endocrinol. 2023, 13, 1056116. [Google Scholar] [CrossRef]
- Toyokuni, S.; Yanatori, I.; Kong, Y.; Zheng, H.; Motooka, Y.; Jiang, L. Ferroptosis at the crossroads of infection, aging and cancer. Cancer Sci. 2020, 111, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Mann, J.; Reznik, E.; Santer, M.; Fongheiser, M.A.; Smith, N.; Hirschhorn, T.; Zandkarimi, F.; Soni, R.K.; Dafré, A.L.; Miranda-Vizuete, A. Ferroptosis inhibition by oleic acid mitigates iron-overload-induced injury. Cell Chem. Biol. 2024, 31, 249–264.e7. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.; Saravia, C.; Vergara, C.E.; Avila, F.; Valdés, H.; Ortiz-Viedma, J. Avocado oil: Characteristics, properties, and applications. Molecules 2019, 24, 2172. [Google Scholar] [CrossRef]
- Lowry, J.R.; Marshall, N.; Wenzel, T.J.; Murray, T.E.; Klegeris, A. The dietary fatty acids α-linolenic acid (ALA) and linoleic acid (LA) selectively inhibit microglial nitric oxide production. Mol. Cell. Neurosci. 2020, 109, 103569. [Google Scholar] [CrossRef]
- Motta, J.R.; Jung, I.; Azzolin, V.F.; Teixeira, C.F.; Braun, L.E.; De Oliveira Nerys, D.A.; Motano, M.A.E.; Duarte, M.; Maia-Ribeiro, E.A.; da Cruz, I.B.M.; et al. Avocado oil (Persea americana) protects SH-SY5Y cells against cytotoxicity triggered by cortisol by the modulation of BDNF, oxidative stress, and apoptosis molecules. J. Food Biochem. 2021, 45, e13596. [Google Scholar] [CrossRef]
- Aschbacher, K.; O’Donovan, A.; Wolkowitz, O.M.; Dhabhar, F.S.; Su, Y.; Epel, E. Good stress, bad stress and oxidative stress: Insights from anticipatory cortisol reactivity. Psychoneuroendocrinology 2013, 38, 1698–1708. [Google Scholar] [CrossRef]
- Latt, H.M.; Matsushita, H.; Morino, M.; Koga, Y.; Michiue, H.; Nishiki, T.; Tomizawa, K.; Matsui, H. Oxytocin inhibits corticosterone-induced apoptosis in primary hippocampal neurons. Neuroscience 2018, 379, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Svandova, E.; Lesot, H.; Sharpe, P.; Matalova, E. Making the head: Caspases in life and death. Front. Cell Dev. Biol. 2023, 10, 1075751. [Google Scholar] [CrossRef]
- Rahaman, M.M.; Hossain, R.; Herrera-Bravo, J.; Islam, M.T.; Atolani, O.; Adeyemi, O.S.; Owolodun, O.A.; Kambizi, L.; Daştan, S.D.; Calina, D. Natural antioxidants from some fruits, seeds, foods, natural products, and associated health benefits: An update. Food Sci. Nutr. 2023, 11, 1657–1670. [Google Scholar]
- Yeung, S.S.Y.; Kwan, M.; Woo, J. Healthy Diet for Healthy Aging. Nutrients 2021, 13, 4310. [Google Scholar] [CrossRef]
- Liu, W.; Hu, B.; Dehghan, M.; Mente, A.; Wang, C.; Yan, R.; Rangarajan, S.; Tse, L.A.; Yusuf, S.; Liu, X. Fruit, vegetable, and legume intake and the risk of all-cause, cardiovascular, and cancer mortality: A prospective study. Clin. Nutr. 2021, 40, 4316–4323. [Google Scholar] [CrossRef] [PubMed]
- Fiorito, G.; Caini, S.; Palli, D.; Bendinelli, B.; Saieva, C.; Ermini, I.; Valentini, V.; Assedi, M.; Rizzolo, P.; Ambrogetti, D. DNA methylation-based biomarkers of aging were slowed down in a two-year diet and physical activity intervention trial: The DAMA study. Aging Cell 2021, 20, e13439. [Google Scholar] [CrossRef]
- Zhou, Y.-F.; Song, X.-Y.; Wu, J.; Chen, G.-C.; Neelakantan, N.; van Dam, R.M.; Feng, L.; Yuan, J.-M.; Pan, A.; Koh, W.-P. Association between dietary patterns in midlife and healthy ageing in Chinese adults: The Singapore Chinese Health Study. J. Am. Med. Dir. Assoc. 2021, 22, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Shen, Y.; Ren, Y.; Mou, Q.; Lin, T.; Zhu, L.; Ren, T. Combined intake of blueberry juice and probiotics ameliorate mitochondrial dysfunction by activating SIRT1 in alcoholic fatty liver disease. Nutr. Metab. 2021, 18, 50. [Google Scholar] [CrossRef]
- Prasun, P.; Ginevic, I.; Oishi, K. Mitochondrial dysfunction in nonalcoholic fatty liver disease and alcohol related liver disease. Transl. Gastroenterol. Hepatol. 2021, 6, 4. [Google Scholar] [CrossRef]
- Fromenty, B.; Roden, M. Mitochondrial alterations in fatty liver diseases. J. Hepatol. 2023, 78, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Man, S.; Sun, B.; Ma, L.; Guo, L.; Huang, L.; Gao, W. Gut liver brain axis in diseases: The implications for therapeutic interventions. Signal Transduct. Target. Ther. 2023, 8, 443. [Google Scholar] [CrossRef]
- Sellami, M.; Bragazzi, N.; Prince, M.S.; Denham, J.; Elrayess, M. Regular, intense exercise training as a healthy aging lifestyle strategy: Preventing DNA damage, telomere shortening and adverse DNA methylation changes over a lifetime. Front. Genet. 2021, 12, 652497. [Google Scholar] [CrossRef]
- Chakravarti, D.; LaBella, K.A.; DePinho, R.A. Telomeres: History, health, and hallmarks of aging. Cell 2021, 184, 306–322. [Google Scholar] [CrossRef]
- D’Angelo, S. Diet and Aging: The Role of Polyphenol-Rich Diets in Slow Down the Shortening of Telomeres: A Review. Antioxidants 2023, 12, 2086. [Google Scholar] [CrossRef]
- Meccariello, R.; D’Angelo, S. Impact of polyphenolic-food on longevity: An elixir of life. An overview. Antioxidants 2021, 10, 507. [Google Scholar] [CrossRef] [PubMed]
- Pes, G.M.; Dore, M.P.; Tsofliou, F.; Poulain, M. Diet and longevity in the Blue Zones: A set-and-forget issue? Maturitas 2022, 164, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Tian, G.; Xue, H.; Zhang, X.; Zhao, Y.; Cheng, G. Higher adherence to the ‘vegetable-rich’ dietary pattern is related to longer telomere length in women. Clin. Nutr. 2018, 37, 1232–1237. [Google Scholar] [CrossRef]
- Li, X.; Li, M.; Cheng, J.; Guan, S.; Hou, L.; Zu, S.; Yang, L.; Wu, H.; Li, H.; Fan, Y.; et al. Association of healthy and unhealthy plant-based diets with telomere length. Clin. Nutr. 2024, 43, 1694–1701. [Google Scholar] [CrossRef]
- Satija, A.; Bhupathiraju, S.N.; Rimm, E.B.; Spiegelman, D.; Chiuve, S.E.; Borgi, L.; Willett, W.C.; Manson, J.E.; Sun, Q.; Hu, F.B. Plant-based dietary patterns and incidence of type 2 diabetes in US men and women: Results from three prospective cohort studies. PLoS Med. 2016, 13, e1002039. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, W.; Li, S.; Tu, H.; Jia, J.; Zhao, W.; Xu, A.; Xu, W.; Tsai, M.K.; Chu, D.T.-W. Association between plant-based dietary pattern and biological aging trajectory in a large prospective cohort. BMC Med. 2023, 21, 310. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shen, J.; Xuan, J.; Zhu, A.; Ji, J.S.; Liu, X.; Cao, Y.; Zong, G.; Zeng, Y.; Wang, X. Plant-based dietary patterns in relation to mortality among older adults in China. Nat. Aging 2022, 2, 224–230. [Google Scholar] [CrossRef]
- Li, H.; Zeng, X.; Wang, Y.; Zhang, Z.; Zhu, Y.; Li, X.; Hu, A.; Zhao, Q.; Yang, W. A prospective study of healthful and unhealthful plant-based diet and risk of overall and cause-specific mortality. Eur. J. Nutr. 2022, 61, 387–398. [Google Scholar] [CrossRef]
- Galiè, S.; Canudas, S.; Muralidharan, J.; García-Gavilán, J.; Bulló, M.; Salas-Salvadó, J. Impact of nutrition on telomere health: Systematic review of observational cohort studies and randomized clinical trials. Adv. Nutr. 2020, 11, 576–601. [Google Scholar] [CrossRef]
- Gavia-García, G.; Rosado-Pérez, J.; Arista-Ugalde, T.L.; Aguiñiga-Sánchez, I.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. Telomere Length and Oxidative Stress and Its Relation with Metabolic Syndrome Components in the Aging. Biology 2021, 10, 253. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, H.; Gutin, B.; Sesso, H.D.; Dong, Y. Higher chocolate intake is associated with longer telomere length among adolescents. Pediatr. Res. 2020, 87, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Sarıtaş, S.; Duman, H.; Pekdemir, B.; Rocha, J.M.; Oz, F.; Karav, S. Functional chocolate: Exploring advances in production and health benefits. Int. J. Food Sci. Technol. 2024, 59, 5303–5325. [Google Scholar] [CrossRef]
- Six, I.; Guillaume, N.; Jacob, V.; Mentaverri, R.; Kamel, S.; Boullier, A.; Slama, M. The Endothelium and COVID-19: An Increasingly Clear Link Brief Title: Endotheliopathy in COVID-19. Int. J. Mol. Sci. 2022, 23, 6196. [Google Scholar] [CrossRef] [PubMed]
- Sfera, A.; Osorio, C.; Zapata Martin del Campo, C.M.; Pereida, S.; Maurer, S.; Maldonado, J.C.; Kozlakidis, Z. Endothelial senescence and chronic fatigue syndrome, a COVID-19 based hypothesis. Front. Cell. Neurosci. 2021, 15, 673217. [Google Scholar] [CrossRef]
- Stobiecka, M.; Król, J.; Brodziak, A. Antioxidant Activity of Milk and Dairy Products. Animals 2022, 12, 245. [Google Scholar] [CrossRef]
- Li, L.; Ma, C.-m.; Yang, Y.; Wang, B.; Liu, X.-f.; Wang, Y.; Bian, X.; Zhang, G.; Zhang, N. Exploring the potential of plant-derived metal ion binding peptides: Preparation, structure-activity relationship, and biological activities. Trends Food Sci. Technol. 2024, 152, 104650. [Google Scholar] [CrossRef]
- Zeng, J.; Li, Y.; Zou, Y.; Yang, Y.; Yang, T.; Zhou, Y. Intestinal toxicity alleviation and efficacy potentiation through therapeutic administration of Lactobacillus paracasei GY-1 in the treatment of gout flares with colchicine. Food Funct. 2024, 15, 1671–1688. [Google Scholar] [CrossRef]
- Shan, S.; Zheng, T.; Zhang, C.; Song, X.; Chen, J.; Shi, Y.; You, J.; Cheng, G.; Xiong, J. Yogurt and Streptococcus thermophilus metabolites ameliorated telomere attrition in D-galactose-induced ageing mice and t-BHP-challenged HepG2 cells. Int. J. Food Sci. Technol. 2020, 55, 2509–2516. [Google Scholar] [CrossRef]
- Chew, H.; Solomon, V.A.; Fonteh, A.N. Involvement of lipids in Alzheimer’s disease pathology and potential therapies. Front. Physiol. 2020, 11, 598. [Google Scholar] [CrossRef]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to detect senescence-associated beta-galactosidase (SA-βgal) activity, a biomarker of senescent cells in culture and in vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef]
- Barnes, R.P.; Fouquerel, E.; Opresko, P.L. The impact of oxidative DNA damage and stress on telomere homeostasis. Mech. Ageing Dev. 2019, 177, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Hor, Y.-Y.; Ooi, C.-H.; Khoo, B.-Y.; Choi, S.-B.; Seeni, A.; Shamsuddin, S.; Oon, C.-E.; Ong, K.-L.; Jeong, W.-S.; Liong, M.-T. Lactobacillus strains alleviated aging symptoms and aging-induced metabolic disorders in aged rats. J. Med. Food 2019, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Krznarić, Ž.; Karas, I.; Ljubas Kelečić, D.; Vranešić Bender, D. The Mediterranean and Nordic diet: A review of differences and similarities of two sustainable, health-promoting dietary patterns. Front. Nutr. 2021, 8, 683678. [Google Scholar] [CrossRef]
- Yang, M.; Chen, L.; Wang, J.; Msigwa, G.; Osman, A.I.; Fawzy, S.; Rooney, D.W.; Yap, P.-S. Circular economy strategies for combating climate change and other environmental issues. Environ. Chem. Lett. 2023, 21, 55–80. [Google Scholar] [CrossRef]
- Stanek, A.; Grygiel-Górniak, B.; Brożyna-Tkaczyk, K.; Myśliński, W.; Cholewka, A.; Zolghadri, S. The Influence of Dietary Interventions on Arterial Stiffness in Overweight and Obese Subjects. Nutrients 2023, 15, 1440. [Google Scholar] [CrossRef]
- Song, S.; Lee, Y.-M.; Lee, Y.Y.; Yeum, K.-J. Oat (Avena sativa) Extract against Oxidative Stress-Induced Apoptosis in Human Keratinocytes. Molecules 2021, 26, 5564. [Google Scholar] [CrossRef]
- Abuetabh, Y.; Wu, H.H.; Chai, C.; Al Yousef, H.; Persad, S.; Sergi, C.M.; Leng, R. DNA damage response revisited: The p53 family and its regulators provide endless cancer therapy opportunities. Exp. Mol. Med. 2022, 54, 1658–1669. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Goerzen, D.; Jena, P.V.; Zeng, E.; Pasquali, M.; Meidl, R.A.; Heller, D.A. Human and environmental safety of carbon nanotubes across their life cycle. Nat. Rev. Mater. 2024, 9, 63–81. [Google Scholar] [CrossRef]
- Neophytou, C.M.; Trougakos, I.P.; Erin, N.; Papageorgis, P. Apoptosis Deregulation and the Development of Cancer Multi-Drug Resistance. Cancers 2021, 13, 4363. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 inflammasome activation and cell death. Cell. Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef]
- Agarwal, A.; Rizwana; Tripathi, A.D.; Kumar, T.; Sharma, K.P.; Patel, S.K.S. Nutritional and functional new perspectives and potential health benefits of quinoa and chia seeds. Antioxidants 2023, 12, 1413. [Google Scholar] [CrossRef] [PubMed]
- Noratto, G.D.; Murphy, K.; Chew, B.P. Quinoa intake reduces plasma and liver cholesterol, lessens obesity-associated inflammation, and helps to prevent hepatic steatosis in obese db/db mouse. Food Chem. 2019, 287, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhou, J.; Cao, Y.; Zhang, J.; Zou, L. Anti-aging effects of polysaccharides from quinoa (Chenopodium quinoa Willd.) in improving memory and cognitive function. J. Funct. Foods 2022, 94, 105097. [Google Scholar] [CrossRef]
- Makhijani, P.; Basso, P.J.; Chan, Y.T.; Chen, N.; Baechle, J.; Khan, S.; Furman, D.; Tsai, S.; Winer, D.A. Regulation of the immune system by the insulin receptor in health and disease. Front. Endocrinol. 2023, 14, 1128622. [Google Scholar] [CrossRef]
- Orea-Soufi, A.; Paik, J.; Bragança, J.; Donlon, T.A.; Willcox, B.J.; Link, W. FOXO transcription factors as therapeutic targets in human diseases. Trends Pharmacol. Sci. 2022, 43, 1070–1084. [Google Scholar] [CrossRef]
- D’Amico, R.; Cordaro, M.; Fusco, R.; Peritore, A.F.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Mandalari, G.; Caccamo, D.; Cuzzocrea, S.; et al. Consumption of Cashew (Anacardium occidentale L.) Nuts Counteracts Oxidative Stress and Tissue Inflammation in Mild Hyperhomocysteinemia in Rats. Nutrients 2022, 14, 1474. [Google Scholar] [CrossRef]
- Mata, A.; Cadenas, S. The Antioxidant Transcription Factor Nrf2 in Cardiac Ischemia–Reperfusion Injury. Int. J. Mol. Sci. 2021, 22, 11939. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Gao, H.; Zhang, Y.; Lan, T.; Wang, J.; Zhao, H.; Zhang, H.; Xue, M.; Liang, H. Folic acid protects against ethanol-induced hepatic mitophagy imbalance by ROS scavenging and attenuating the elevated Hcy levels. J. Agric. Food Chem. 2023, 71, 14276–14288. [Google Scholar] [CrossRef]
- Saw, C.L.L.; Yang, A.Y.; Guo, Y.; Kong, A.-N.T. Astaxanthin and omega-3 fatty acids individually and in combination protect against oxidative stress via the Nrf2–ARE pathway. Food Chem. Toxicol. 2013, 62, 869–875. [Google Scholar] [CrossRef]
- Kanwugu, O.N.; Glukhareva, T.V. Activation of Nrf2 pathway as a protective mechanism against oxidative stress-induced diseases: Potential of astaxanthin. Arch. Biochem. Biophys. 2023, 741, 109601. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Hu, Z.; Shen, Q.; Liang, H.; Jiang, X. Resveratrol attenuated homocysteine-induced apoptosis of rat ventricular myocytes. Int. J. Cardiol. 2014, 177, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, N.; Condello, S.; Currò, M.; Parisi, G.; Ientile, R.; Caccamo, D. NF-kappaB activation is associated with homocysteine-induced injury in Neuro2a cells. BMC Neurosci. 2008, 9, 62. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Elewa, Y.H.A.; Zahran, M.H.; Alexiou, A.; Papadakis, M.; Batiha, G.E.-S. Parkinson’s disease risk and hyperhomocysteinemia: The possible link. Cell. Mol. Neurobiol. 2023, 43, 2743–2759. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Free radical theory of aging: Dietary implications. Am. J. Clin. Nutr. 1972, 25, 839–843. [Google Scholar] [CrossRef]
- Remigante, A.; Spinelli, S.; Straface, E.; Gambardella, L.; Russo, M.; Cafeo, G.; Caruso, D.; Falliti, G.; Dugo, P.; Dossena, S.; et al. Mechanisms underlying the anti-aging activity of bergamot (Citrus bergamia) extract in human red blood cells. Front. Physiol. 2023, 14, 1225552. [Google Scholar] [CrossRef]
- Fernandes, A.C.; Santana, Á.L.; Vieira, N.C.; Gandra, R.L.; Rubia, C.; Castro-Gamboa, I.; Macedo, J.A.; Macedo, G.A. In vitro effects of peanut skin polyphenolic extract on oxidative stress, adipogenesis, and lipid accumulation. J. Food Process. Preserv. 2021, 45, e15815. [Google Scholar] [CrossRef]
- Fernandes, A.C.F.; Martins, I.M.; Moreira, D.K.T.; Macedo, G.A. Use of agro-industrial residues as potent antioxidant, antiglycation agents, and α-amylase and pancreatic lipase inhibitory activity. J. Food Process. Preserv. 2020, 44, e14397. [Google Scholar] [CrossRef]
- Ho, G.T.T.; Wangensteen, H.; Barsett, H. Elderberry and elderflower extracts, phenolic compounds, and metabolites and their effect on complement, RAW 264.7 macrophages and dendritic cells. Int. J. Mol. Sci. 2017, 18, 584. [Google Scholar] [CrossRef]
- Liu, D.; He, X.-Q.; Wu, D.-T.; Li, H.-B.; Feng, Y.-B.; Zou, L.; Gan, R.-Y. Elderberry (Sambucus nigra L.): Bioactive compounds, health functions, and applications. J. Agric. Food Chem. 2022, 70, 4202–4220. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Junk Food Type | Components | Health Impacts |
|---|---|---|
| Sweetened soda, soft drinks | High-fructose corn syrup | Weight gain and diabetes, hypertension, atherosclerosis, coronary heart disease. |
| French fries, ice-cream | Trans fats | Increase in inflammatory markers, cancer, and diabetes. |
| Burgers and sandwiches | Per/poly fluoroalkyl substances (PFAS) | Breast cancer, infertility, weakened immune system. |
| Processed cheese, chicken nuggets | Phosphate additives | Kidney disease, bone problems. |
| Processed red meats | Sodium nitrite | Stomach cancer, renal inflammation. |
| Candy | Sugar, sweeteners, corn syrup | Weight gain and cardiovascular health. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinci, G.; Davì, F.; Pellegrino, T.; Fusco, R.; Cordaro, M.; Di Paola, R. Main Dietary Patterns for Healthy Aging and Well-Being. Nutrients 2025, 17, 2009. https://doi.org/10.3390/nu17122009
Vinci G, Davì F, Pellegrino T, Fusco R, Cordaro M, Di Paola R. Main Dietary Patterns for Healthy Aging and Well-Being. Nutrients. 2025; 17(12):2009. https://doi.org/10.3390/nu17122009
Chicago/Turabian StyleVinci, Graziano, Federica Davì, Teresina Pellegrino, Roberta Fusco, Marika Cordaro, and Rosanna Di Paola. 2025. "Main Dietary Patterns for Healthy Aging and Well-Being" Nutrients 17, no. 12: 2009. https://doi.org/10.3390/nu17122009
APA StyleVinci, G., Davì, F., Pellegrino, T., Fusco, R., Cordaro, M., & Di Paola, R. (2025). Main Dietary Patterns for Healthy Aging and Well-Being. Nutrients, 17(12), 2009. https://doi.org/10.3390/nu17122009