
Vitamin B3 Ameliorates Sleep Duration and Quality in Clinical and Pre-Clinical Studies


Abstract
1. Introduction
2. Materials and Methods
2.1. Inclusion and Exclusion Criteria
2.2. Data Extraction
3. Results
3.1. Literature Search and Studies Characteristics
3.2. Safety of Nicotinamide Riboside Supplementation
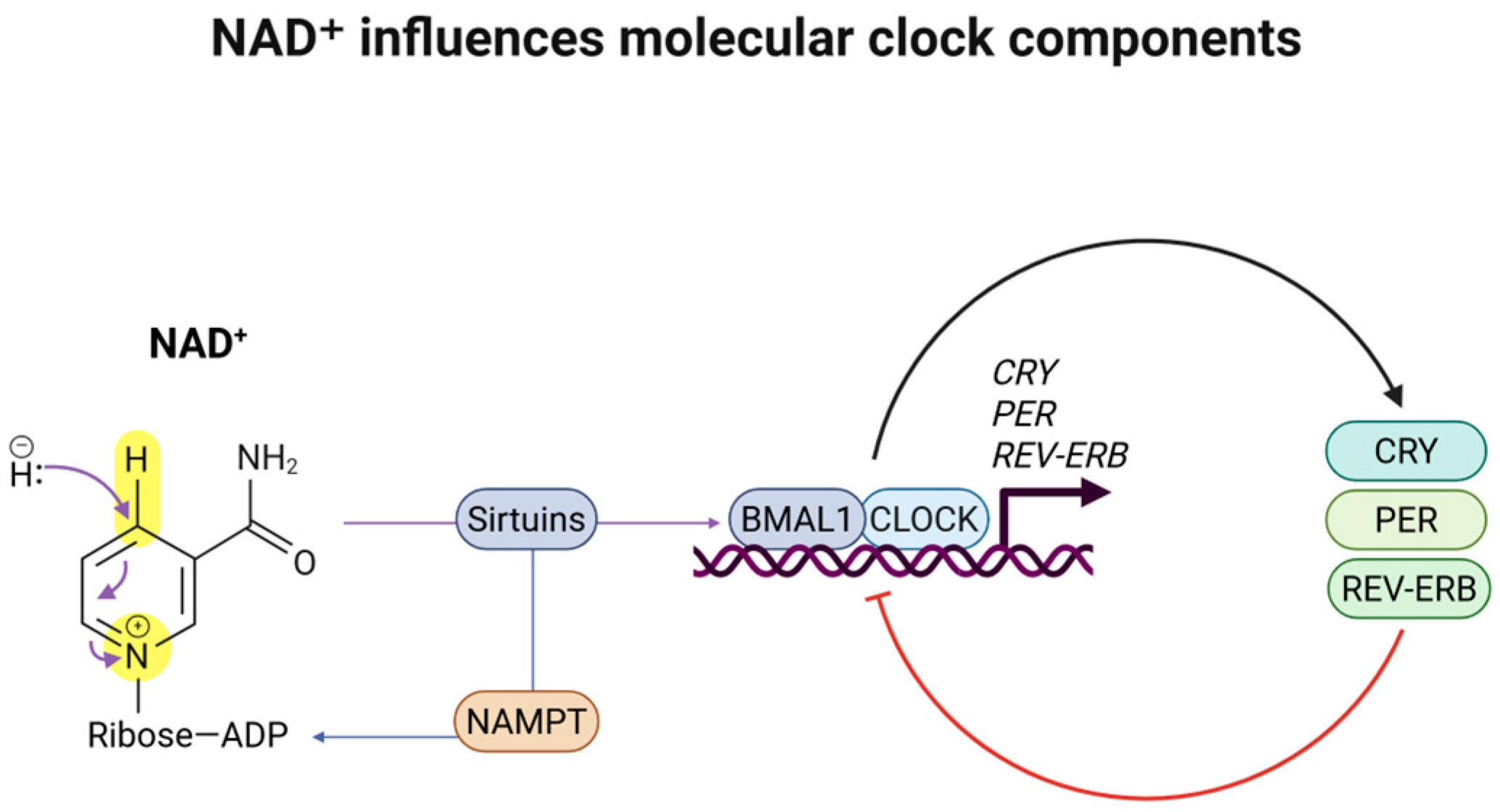
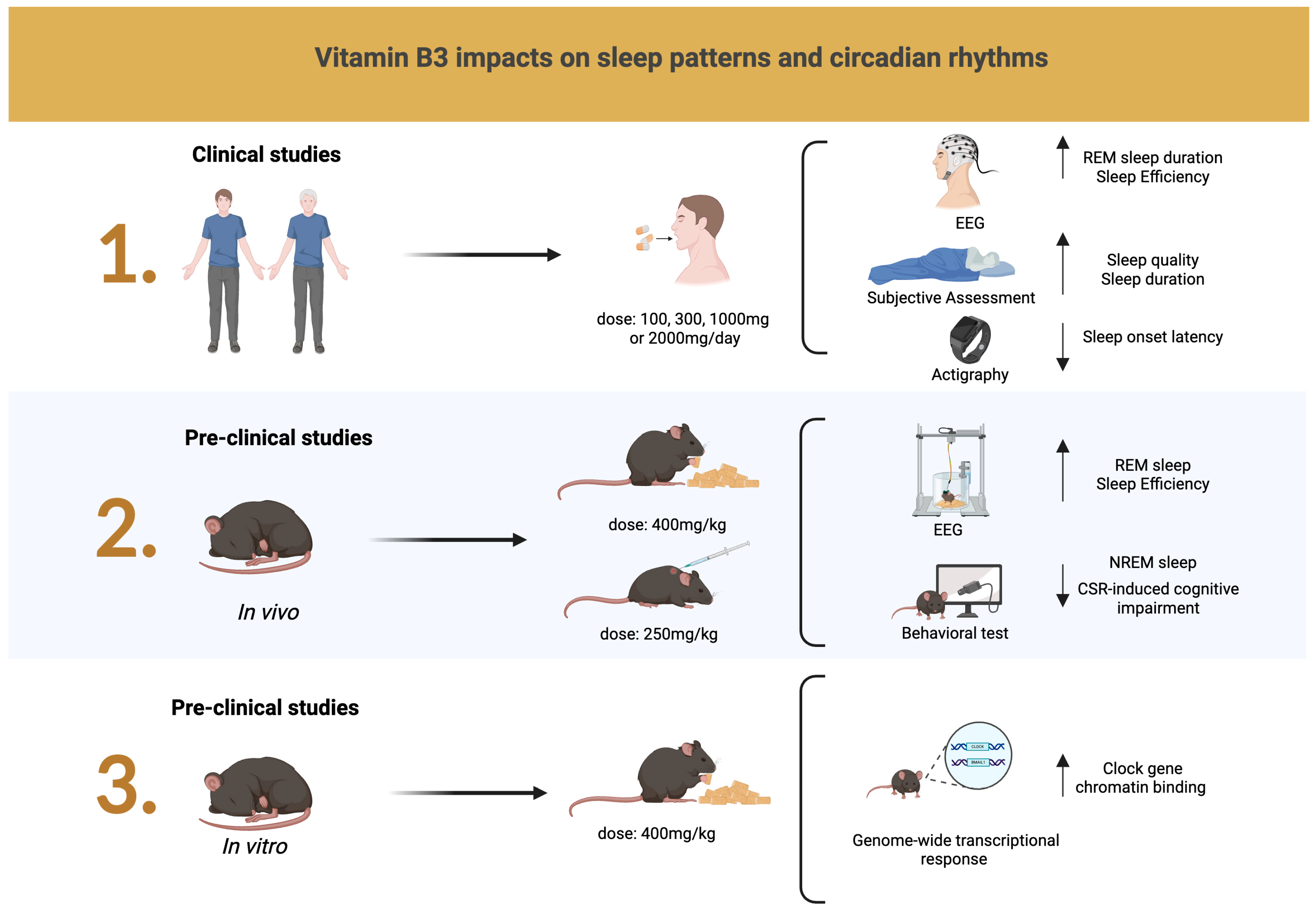
3.3. Impacts of NR Supplementation on Circadian Rhythms and Genetic Mechanisms
3.4. NR Supplementation Improves Sleep in Animal Models
3.5. NR Ameliorates Sleep Patterns in Humans
4. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMAIL1 | Brain and muscle ARNT-like protein 1 |
| EEG | Electroencephalogram |
| NAD+ | Nicotinamide adenine dinucleotide |
| NMN | Nicotinamide mononucleotide |
| NR | Nicotinamide riboside |
| NREM | Non-rapid eye movement |
| REM | Rapid eye movement |
References
- Alegre, G.F.S.; Pastore, G.M. NAD+ Precursors Nicotinamide Mononucleotide (NMN) and Nicotinamide Riboside (NR): Potential Dietary Contribution to Health. Curr. Nutr. Rep. 2023, 12, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Marinescu, A.G.; Chen, J.; Holmes, H.E.; Guarente, L.; Mendes, O.; Morris, M.; Dellinger, R.W. Safety Assessment of High-Purity, Synthetic Nicotinamide Riboside (NR-E) in a 90-Day Repeated Dose Oral Toxicity Study, With a 28-Day Recovery Arm. Int. J. Toxicol. 2020, 39, 307–320. [Google Scholar] [CrossRef]
- Altay, O.; Yang, H.; Yildirim, S.; Bayram, C.; Bolat, I.; Oner, S.; Tozlu, O.O.; Arslan, M.E.; Hacimuftuoglu, A.; Shoaie, S.; et al. Combined Metabolic Activators with Different NAD+ Precursors Improve Metabolic Functions in the Animal Models of Neurodegenerative Diseases. Biomedicines 2024, 12, 927. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Pan, Y.; Vempati, P.; Zhao, W.; Knable, L.; Ho, L.; Wang, J.; Sastre, M.; Ono, K.; Sauve, A.A.; et al. Nicotinamide riboside restores cognition through an upregulation of proliferator-activated receptor-γ coactivator 1α regulated β-secretase 1 degradation and mitochondrial gene expression in Alzheimer’s mouse models. Neurobiol. Aging 2013, 34, 1581–1588. [Google Scholar] [CrossRef]
- Roth, T. Slow wave sleep: Does it matter? J. Clin. Sleep Med. 2009, 5, S4–S5. [Google Scholar] [CrossRef]
- Peever, J.; Fuller, P.M. The Biology of REM Sleep. Curr. Biol. 2017, 27, R1237–R1248. [Google Scholar] [CrossRef]
- Levine, D.C.; Ramsey, K.M.; Bass, J. Circadian NAD(P)(H) cycles in cell metabolism. Semin. Cell Dev. Biol. 2022, 126, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A. The two-process model of sleep regulation: Beginnings and outlook. J. Sleep Res. 2022, 31, e13598. [Google Scholar] [CrossRef]
- Quante, M.; Mariani, S.; Weng, J.; Marinac, C.R.; Kaplan, E.R.; Rueschman, M.; Mitchell, J.A.; James, P.; Hipp, J.A.; Cespedes Feliciano, E.M.; et al. Zeitgebers and their association with rest-activity patterns. Chronobiol. Int. 2019, 36, 203–213. [Google Scholar] [CrossRef]
- Eckel-Mahan, K.; Sassone-Corsi, P. Phenotyping Circadian Rhythms in Mice. Curr. Protoc. Mouse Biol. 2015, 5, 271–281. [Google Scholar] [CrossRef]
- Cuenoud, B.; Huang, Z.; Hartweg, M.; Widmaier, M.; Lim, S.; Wenz, D.; Xin, L. Effect of circadian rhythm on NAD and other metabolites in human brain. Front. Physiol. 2023, 14, 1285776. [Google Scholar] [CrossRef] [PubMed]
- Peek, C.B.; Affinati, A.H.; Ramsey, K.M.; Kuo, H.Y.; Yu, W.; Sena, L.A.; Ilkayeva, O.; Marcheva, B.; Kobayashi, Y.; Omura, C.; et al. Circadian clock NAD+ cycle drives mitochondrial oxidative metabolism in mice. Science 2013, 342, 1243417. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Ramsey, K.M.; Marcheva, B.; Bass, J. Circadian rhythms, sleep, and metabolism. J. Clin. Investig. 2011, 121, 2133–2141. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, Y.; Sahar, S.; Astarita, G.; Kaluzova, M.; Sassone-Corsi, P. Circadian Control of the NAD+ Salvage Pathway by CLOCK-SIRT1. Science 2009, 324, 654–657. [Google Scholar] [CrossRef]
- Bellet, M.M.; Orozco-Solis, R.; Sahar, S.; Eckel-Mahan, K.; Sassone-Corsi, P. The time of metabolism: NAD+, SIRT1, and the circadian clock. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 31–38. [Google Scholar] [CrossRef]
- Levine, D.C.; Hong, H.; Weidemann, B.J.; Ramsey, K.M.; Affinati, A.H.; Schmidt, M.S.; Cedernaes, J.; Omura, C.; Braun, R.; Lee, C.; et al. NAD(+) Controls Circadian Reprogramming through PER2 Nuclear Translocation to Counter Aging. Mol. Cell 2020, 78, 835–849.e7. [Google Scholar] [CrossRef]
- Imai, S. “Clocks” in the NAD World: NAD as a metabolic oscillator for the regulation of metabolism and aging. Biochim. Biophys. Acta 2010, 1804, 1584–1590. [Google Scholar] [CrossRef]
- Rajman, L.; Chwalek, K.; Sinclair, D.A. Therapeutic Potential of NAD-Boosting Molecules: The In Vivo Evidence. Cell Metab. 2018, 27, 529–547. [Google Scholar] [CrossRef]
- Irie, J.; Inagaki, E.; Fujita, M.; Nakaya, H.; Mitsuishi, M.; Yamaguchi, S.; Yamashita, K.; Shigaki, S.; Ono, T.; Yukioka, H.; et al. Effect of oral administration of nicotinamide mononucleotide on clinical parameters and nicotinamide metabolite levels in healthy Japanese men. Endocr. J. 2020, 67, 153–160. [Google Scholar] [CrossRef]
- Schibler, U. Senescence of Timing Reverted: NAD(+) Rejuvenates the Circadian Clock. Mol. Cell 2020, 78, 805–807. [Google Scholar] [CrossRef]
- Bushana, P.N.; Schmidt, M.A.; Rempe, M.J.; Sorg, B.A.; Wisor, J.P. Chronic dietary supplementation with nicotinamide riboside reduces sleep need in the laboratory mouse. Sleep Adv. 2023, 4, zpad044. [Google Scholar] [CrossRef]
- Remie, C.M.E.; Roumans, K.H.M.; Moonen, M.P.B.; Connell, N.J.; Havekes, B.; Mevenkamp, J.; Lindeboom, L.; de Wit, V.H.W.; van de Weijer, T.; Aarts, S.; et al. Nicotinamide riboside supplementation alters body composition and skeletal muscle acetylcarnitine concentrations in healthy obese humans. Am. J. Clin. Nutr. 2020, 112, 413–426. [Google Scholar] [CrossRef]
- Gindri, I.M.; Ferrari, G.; Pinto, L.P.S.; Bicca, J.; Dos Santos, I.K.; Dallacosta, D.; Roesler, C.R.M. Evaluation of safety and effectiveness of NAD in different clinical conditions: A systematic review. Am. J. Physiol. Endocrinol. Metab. 2024, 326, E417–E427. [Google Scholar] [CrossRef] [PubMed]
- Conze, D.; Brenner, C.; Kruger, C.L. Safety and Metabolism of Long-term Administration of NIAGEN (Nicotinamide Riboside Chloride) in a Randomized, Double-Blind, Placebo-controlled Clinical Trial of Healthy Overweight Adults. Sci. Rep. 2019, 9, 9772. [Google Scholar] [CrossRef] [PubMed]
- Beaton, J.M.; Pegram, G.V.; Smythies, J.R.; Bradley, R.J. The effects of nicotinamide on mouse sleep. Experientia 1974, 30, 926–927. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xiao, J.; Lin, Z.; Xu, X.; Chen, J. NAD+ supplement relieved chronic sleep restriction (CSR)-induced microglial proinflammation in vivo and in vitro. J. Neuroimmunol. 2024, 397, 578469. [Google Scholar] [CrossRef]
- Trammell, S.A.; Schmidt, M.S.; Weidemann, B.J.; Redpath, P.; Jaksch, F.; Dellinger, R.W.; Li, Z.; Abel, E.D.; Migaud, M.E.; Brenner, C. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat. Commun. 2016, 7, 12948. [Google Scholar] [CrossRef]
- Martens, C.R.; Denman, B.A.; Mazzo, M.R.; Armstrong, M.L.; Reisdorph, N.; McQueen, M.B.; Chonchol, M.; Seals, D.R. Chronic nicotinamide riboside supplementation is well-tolerated and elevates NAD(+) in healthy middle-aged and older adults. Nat. Commun. 2018, 9, 1286. [Google Scholar] [CrossRef]
- Kim, M.; Seol, J.; Sato, T.; Fukamizu, Y.; Sakurai, T.; Okura, T. Effect of 12-Week Intake of Nicotinamide Mononucleotide on Sleep Quality, Fatigue, and Physical Performance in Older Japanese Adults: A Randomized, Double-Blind Placebo-Controlled Study. Nutrients 2022, 14, 755. [Google Scholar] [CrossRef]
- Imai, S.-i.; Guarente, L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 2014, 24, 464–471. [Google Scholar] [CrossRef]
- Beaton, J.M.; Smythies, J.R.; Bradley, R.J. The behavioral effects of L-methionine and related compounds in rats and mice. Biol. Psychiatry 1975, 10, 45–52. [Google Scholar] [PubMed]
- Scherer, B.; Krämer, W. Influence of niacinamide administration on brain 5-HT and a possible mode of action. Life Sci. 1972, 11, 189–195. [Google Scholar] [CrossRef]
- Soon, C.S.; Thota, R.; Owen, L.; Tian, L.; Martin, F.P.; Mantantzis, K.; Cherta-Murillo, A.; Campos, V.C.; Chkroun, C.; Lavalle, L.; et al. Mulberry leaf extract combined with tryptophan improves sleep and post wake mood in adults with sleep complaints—A randomized cross-over study. Eur. J. Nutr. 2025, 64, 124. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.R.; Pegram, G.V.; Hyde, P.R.; Beaton, J.M.; Smythies, J.R. The effects of nicotinamide upon sleep in humans. Biol. Psychiatry 1977, 12, 139–143. [Google Scholar] [PubMed]
- Morifuji, M.; Higashi, S.; Ebihara, S.; Nagata, M. Ingestion of β-nicotinamide mononucleotide increased blood NAD levels, maintained walking speed, and improved sleep quality in older adults in a double-blind randomized, placebo-controlled study. Geroscience 2024, 46, 4671–4688. [Google Scholar] [CrossRef]
- Nguyen, H.D. Higher intakes of nutrients and regular drinking are associated with habitual sleep duration in pre- and postmenopausal women with comorbidities. Sleep Health 2023, 9, 688–697. [Google Scholar] [CrossRef]
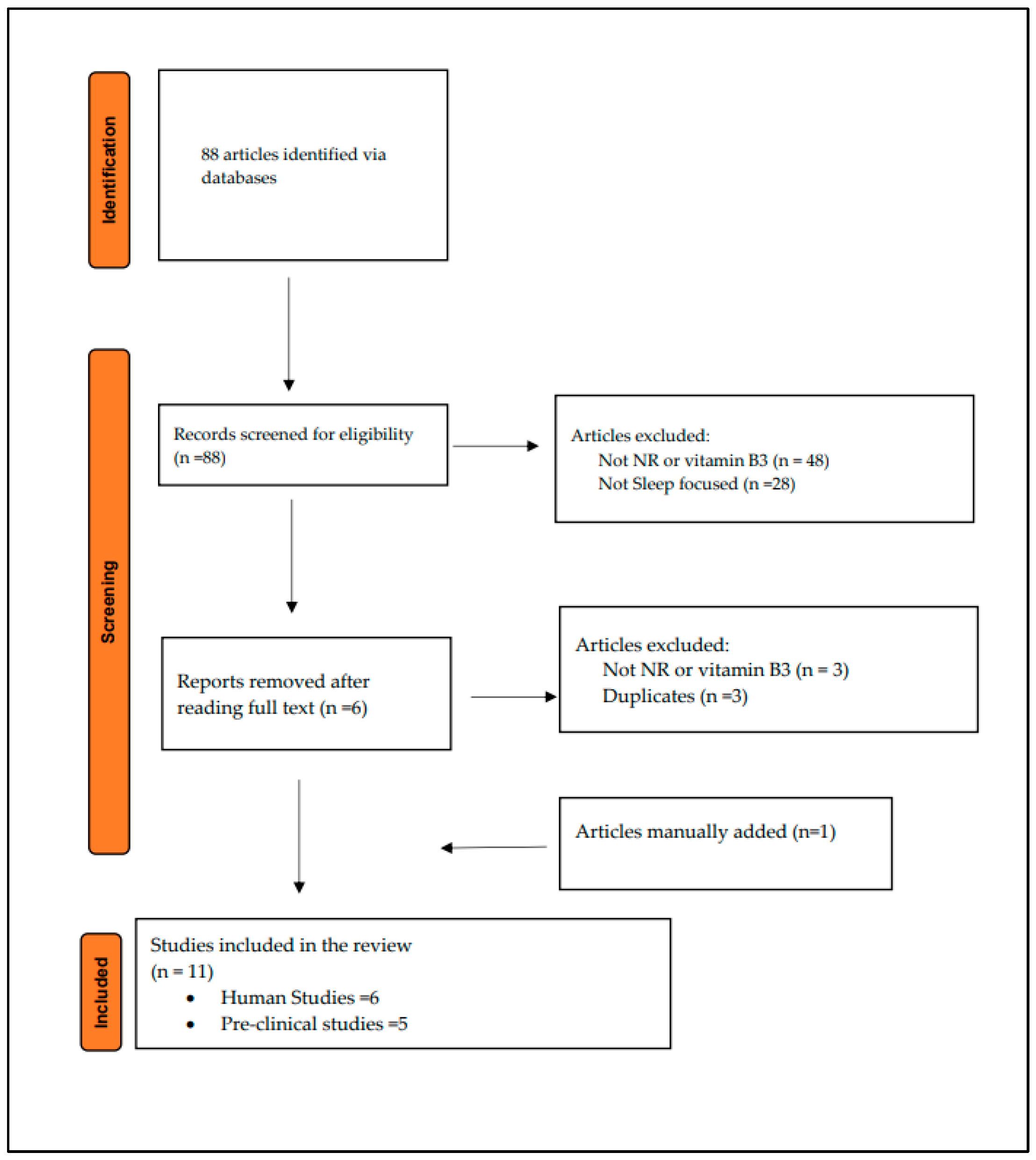

{kind=link}
{kind=link}
{kind=link}
| MeSH Terms | Search Results |
|---|---|
| ((nicotinamide riboside) OR (vitamin B3)) AND (sleep) | 87 |
| ((nicotinamide riboside) AND (vitamin B3)) AND (sleep) | 1 |
| Author | Year | Study Population | NR Supplementation Regimen | Treatment Type and Duration | Sleep Assessments | Sleep Findings |
|---|---|---|---|---|---|---|
| Pre-Clinical Studies | ||||||
| Beaton, et al. [25] | 1974 | Mouse | 250 mg/kg nicotinamide riboside | Daily injections | Sleep–wake cycle |  REM (mice) REM (mice) |
| Beaton, et al. [19] | 1975 | Swiss mice and rats |
| Daily injections 21 days | Sleep–wake cycle |  REM (mice) REM (mice) |
| Bushana et al. [12] | 2023 | C57BL/6J mice on a 12:12 LD cycle n = 33, 12 females, 11 males; aged 10–12 weeks |
| 6–10 weeks | Implanted with an EEG and electromyographic (EMG) to monitor a 6 h sleep deprivation protocol and 30 min recovery sessions |  17% NREMS 17% NREMS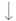 Cumulative manifestation of sleep need during quiet wake Cumulative manifestation of sleep need during quiet wake |
| Chen et al. [14] | 2024 | C57BL/6 mice (6–8 weeks, 22–26 g) |
| 14 days | Chronic sleep restriction (CSR) with a forced locomotion protocol |  CSR-induced cognitive decline CSR-induced cognitive decline |
| Levine et al. [8] | 2020 | 4-month-old mice housed in a 12:12 LD cycle | 400 mg/kg/day of NR | Four months | Clock gene PER2 and BMAL1 expression | Restored BMAL1 levels Enhanced PER2 |
| Human Studies | ||||||
| Irie et al. [9] | 2020 | 10 healthy Japanese males. | 100, 250, and 500 mg of nicotinamide mononucleotide (NMN) for a 5 h pharmacokinetics assessment | Single oral administration | Pittsburgh Sleep Quality Index (PSQI) | No significant findings |
| Kim et al. [17] | 2022 | 108 Japanese older adults (mean age 72.2) | 250 mg of vitamin B3 or placebo for 12 weeks | Daily oral administration | Pittsburgh Sleep Quality Index (PSQI) |  Sleep duration Sleep duration  Sleep quality (PSQI global score) Sleep quality (PSQI global score) |
| Morifuji et al. [23] | 2024 | 60 Japanese older adults (mean age 69.0) | 250 mg of β-nicotinamide mononucleotide or placebo for 12 weeks | Daily oral administration | Pittsburgh Sleep Quality Index (PSQI |  Sleep quality (PSQI global score) Sleep quality (PSQI global score) |
| Nguyen et al. [26] | 2023 | 2084 pre- and postmenopausal Korean women 18–80 years old (KNHASES Study, 2016–2018) | 12 nutrients (vitamin B1, B3, vitamin C, PUFA, n-6 fatty acid, iron, potassium, phosphorus, calcium, fiber, carbohydrate) | Daily oral administration | Habitual sleep duration per night: <5 h = very short 5–6 = short 7–8 h = normal ≥9 = long | |
| Robinson, et al. [22] | 1977 | 6 adults with normal sleep (3 males and 2 females) and 2 females with severe insomnia | 1–3 g of NR twice daily for 23 days | Daily oral administration | In-lab sleep recording |  Sleep efficiency Sleep efficiency REM sleep duration REM sleep duration |
| Soon et al. [24] | 2025 | 43 adults |
| Morning vs. evening 14-day oral administration + 28-day washout period | Actigraphy Karolinska Sleepiness Scale | Evening supplements: 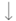 Sleep onset Sleep onset Sleep quality Sleep quality |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weiss, C. Vitamin B3 Ameliorates Sleep Duration and Quality in Clinical and Pre-Clinical Studies. Nutrients 2025, 17, 1982. https://doi.org/10.3390/nu17121982
Weiss C. Vitamin B3 Ameliorates Sleep Duration and Quality in Clinical and Pre-Clinical Studies. Nutrients. 2025; 17(12):1982. https://doi.org/10.3390/nu17121982
Chicago/Turabian StyleWeiss, Carleara. 2025. "Vitamin B3 Ameliorates Sleep Duration and Quality in Clinical and Pre-Clinical Studies" Nutrients 17, no. 12: 1982. https://doi.org/10.3390/nu17121982
APA StyleWeiss, C. (2025). Vitamin B3 Ameliorates Sleep Duration and Quality in Clinical and Pre-Clinical Studies. Nutrients, 17(12), 1982. https://doi.org/10.3390/nu17121982