Potential Therapeutic Targets for Androgenetic Alopecia (AGA) in Obese Individuals as Revealed by a Gut Microbiome Analysis: A Mendelian Randomization Study

 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Methods
2.1. Data
2.2. Statistics
- (1)
- Relevance assumption: Single-nucleotide polymorphisms (SNPs) must exhibit a strong association with the exposure. This association is quantified via the F statistic, where an F > 10 indicates strong relevance (formula: F = β2/Se2)
- (2)
- Independence assumption: The SNPs should not be associated with any confounding factors that may influence both the exposure and the outcome, nor should they be directly associated with the outcome.
- (3)
2.3. Selection of Genetic Instrumental Variables (IVs)
- (1)
- To acquire more extensive data, SNPs from the GWAS data on the exposure were selected based on their strong association with the exposure; specifically, SNPs with p-values < 0.00001.
- (2)
- Based on the independence assumption of MR, each SNP must be independent of the others. To ensure that there was no linkage disequilibrium (LD) among the SNPs, we applied an LD threshold of R2 = 0.001 and a distance criterion of ≤10,000 kb.
- (3)
- To ensure that the same SNPs have identical alleles in both the exposure and the outcome, ambiguous alleles were excluded.
- (4)
- The exclusion–restriction assumption requires that SNPs associated with the exposure should not be directly related to the outcome. Consequently, the online tool National Institutes of Health LDlink (https://ldlink.nih.gov/?tab=ldtrait#ldtrait-tab (accessed on 1 November 2024)) was utilized to identify traits directly associated with the SNPs. SNPs linked to the outcome were excluded, and SNPs lacking alternative loci were removed [14].
- (5)
- The sensitivity analysis removed SNPs with potential pleiotropy.
2.4. Statistical Analysis
3. Results
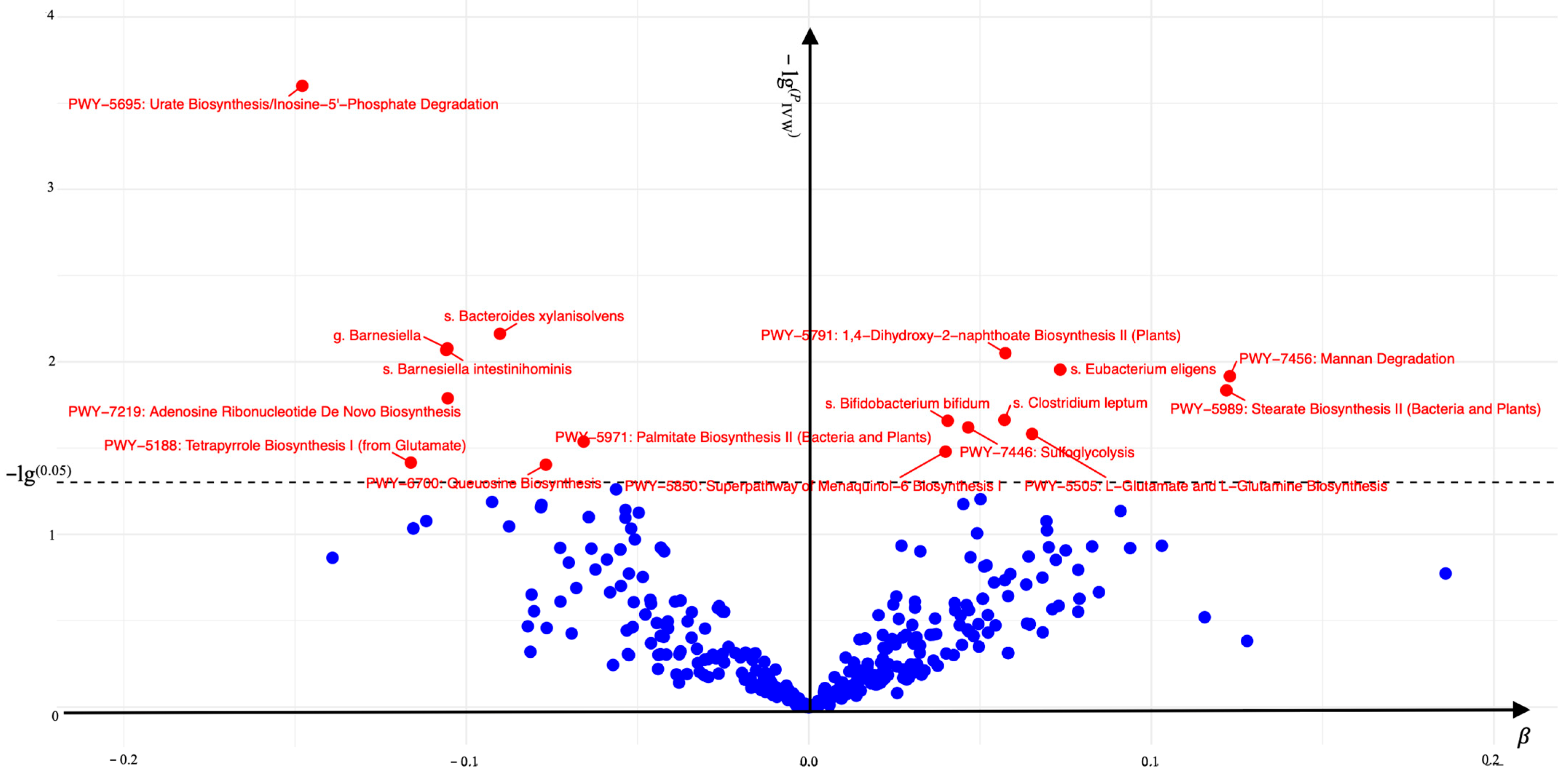
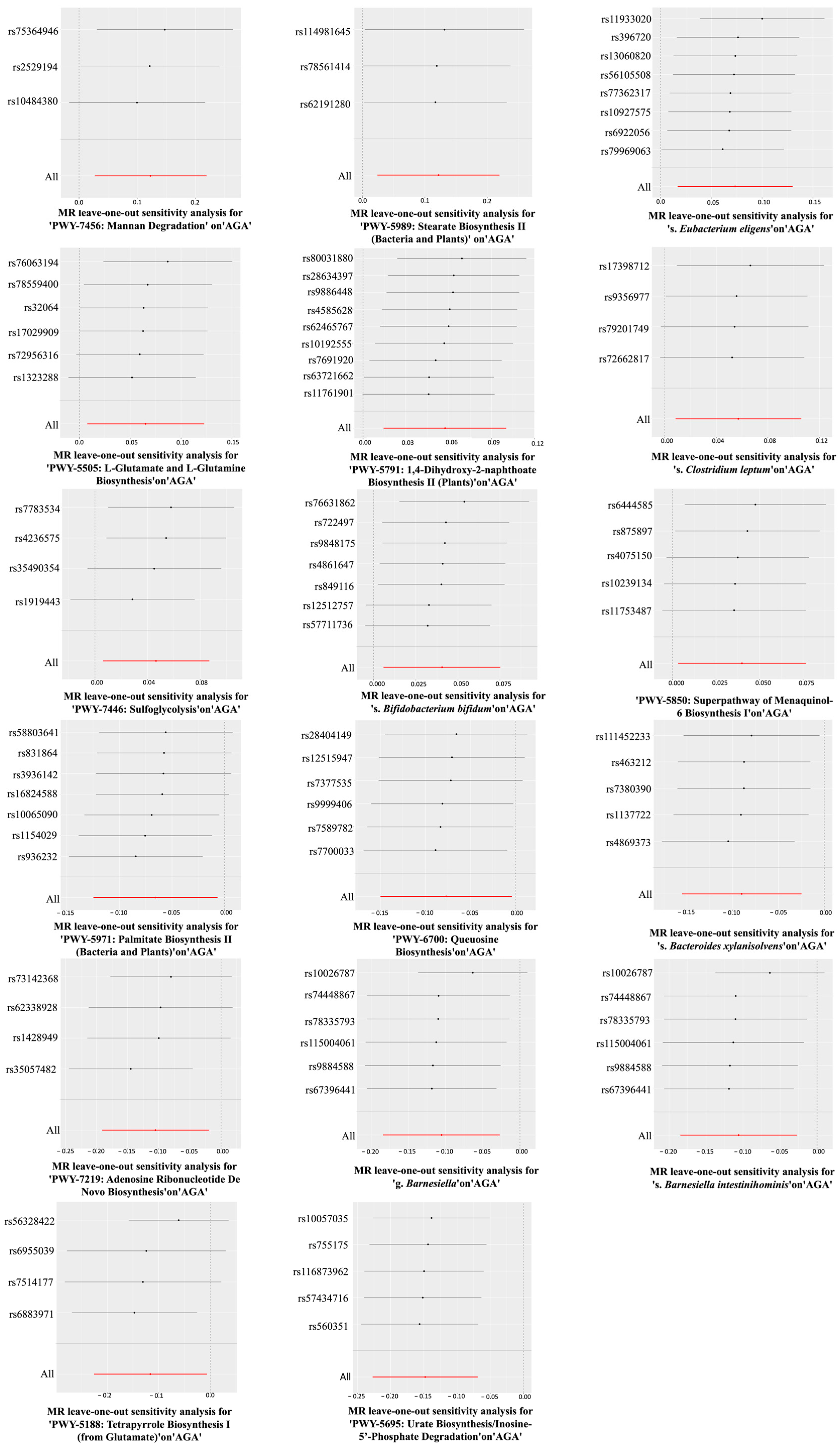
3.1. The Association Between the Gut Microbiome (Exposure 1) and AGA
3.2. The Association Between the Gut Microbiomes (Exposure 3) and Obesity
3.3. The Association Between Obesity (Exposure 2) and AGA
4. Discussion
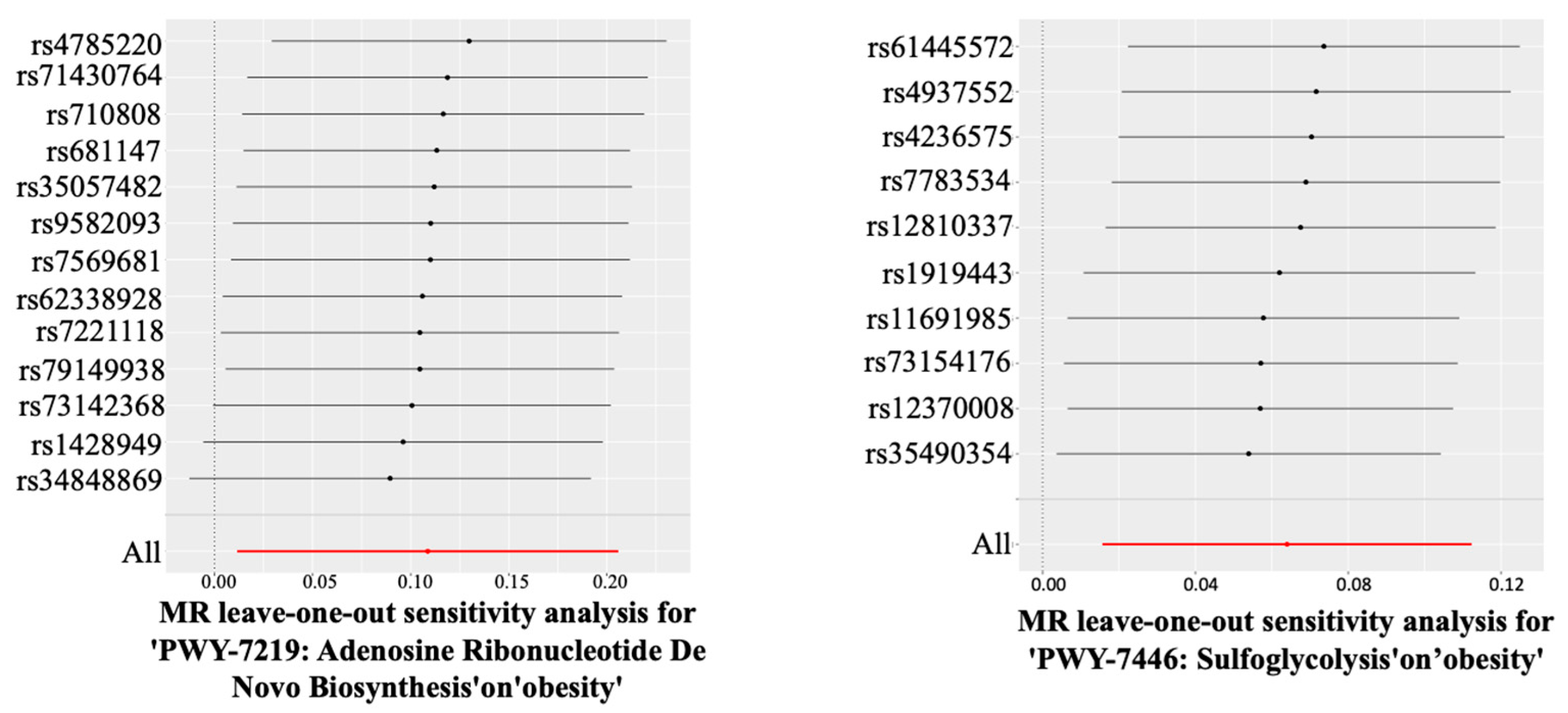
4.1. The Impact of the Gut Bacterial Pathway of Sulfoglycolysis on Obesity and AGA
4.2. The Impact of the Gut Bacterial Pathway of Adenosine Ribonucleotide De Novo Biosynthesis on Obesity and AGA
5. Conclusions
6. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sorbellini, E.; Pinto, D.; Marzani, B.; Rinaldi, F. Drug Treatment for Androgenetic Alopecia: First Italian Questionnaire Survey on What Dermatologists Think about Finasteride. Dermatol. Ther. 2018, 8, 259–267. [Google Scholar] [CrossRef]
- Wang, T.; Zhou, C.; Shen, Y.; Wang, X.; Ding, X.; Tian, S.; Liu, Y.; Peng, G.; Xue, S.; Zhou, J. Prevalence of androgenetic alopecia in China: A community-based study in six cities. Br. J. Dermatol. 2010, 162, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Paik, J.H.; Yoon, J.B.; Sim, W.Y.; Kim, B.S.; Kim, N.I. The prevalence and types of androgenetic alopecia in Korean men and women. Br. J. Dermatol. 2001, 145, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Gan, D.C.; Sinclair, R.D. Prevalence of male and female pattern hair loss in Maryborough. J. Investig. Dermatol. Symp. Proc. 2005, 10, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Lolli, F.; Pallotti, F.; Rossi, A.; Fortuna, M.C.; Caro, G.; Lenzi, A.; Sansone, A.; Lombardo, F. Androgenetic alopecia: A review. Endocrine 2017, 57, 9–17. [Google Scholar] [CrossRef]
- Chen, S.; Xie, X.; Zhang, G.; Zhang, Y. Comorbidities in androgenetic alopecia: A comprehensive review. Dermatol. Ther. 2022, 12, 2233–2247. [Google Scholar] [CrossRef]
- Yang, C.-C.; Hsieh, F.-N.; Lin, L.-Y.; Hsu, C.-K.; Sheu, H.-M.; Chen, W. Higher body mass index is associated with greater severity of alopecia in men with male-pattern androgenetic alopecia in Taiwan: A cross-sectional study. J. Am. Acad. Dermatol. 2014, 70, 297–302.e291. [Google Scholar] [CrossRef]
- Sanke, S.; Chander, R.; Jain, A.; Garg, T.; Yadav, P. A comparison of the hormonal profile of early androgenetic alopecia in men with the phenotypic equivalent of polycystic ovarian syndrome in women. JAMA Dermatol. 2016, 152, 986–991. [Google Scholar] [CrossRef]
- Shen, J.; Obin, M.S.; Zhao, L. The gut microbiota, obesity and insulin resistance. Mol. Asp. Med. 2013, 34, 39–58. [Google Scholar] [CrossRef]
- Zhao, L. The gut microbiota and obesity: From correlation to causality. Nat. Rev. Microbiol. 2013, 11, 639–647. [Google Scholar] [CrossRef]
- Mayer, E.A. Gut feelings: The emerging biology of gut–brain communication. Nat. Rev. Neurosci. 2011, 12, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The central nervous system and the gut microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, E.; Glymour, M.M.; Holmes, M.V.; Kang, H.; Morrison, J.; Munafò, M.R.; Palmer, T.; Schooling, C.M.; Wallace, C.; Zhao, Q.; et al. Mendelian randomization. Nat. Rev. Methods Primers 2022, 2, 6. [Google Scholar] [CrossRef]
- Sanderson, E.; Davey Smith, G.; Windmeijer, F.; Bowden, J. An examination of multivariable Mendelian randomization in the single-sample and two-sample summary data settings. Int. J. Epidemiol. 2019, 48, 713–727. [Google Scholar] [CrossRef]
- Morinaga, H.; Mohri, Y.; Grachtchouk, M.; Asakawa, K.; Matsumura, H.; Oshima, M.; Takayama, N.; Kato, T.; Nishimori, Y.; Sorimachi, Y. Obesity accelerates hair thinning by stem cell-centric converging mechanisms. Nature 2021, 595, 266–271. [Google Scholar] [CrossRef]
- Elsaie, L.T.; Elshahid, A.R.; Hasan, H.M.; Soultan, F.A.Z.M.; Jafferany, M.; Elsaie, M.L. Cross sectional quality of life assessment in patients with androgenetic alopecia. Dermatol. Ther. 2020, 33, e13799. [Google Scholar] [CrossRef]
- Mui, J.W.-Y.; De Souza, D.P.; Saunders, E.C.; McConville, M.J.; Williams, S.J. Remodeling of Carbon Metabolism During Sulfoglycolysis in Escherichia coli. Appl. Environ. Microbiol. 2023, 89, e02016–e02022. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.-O. NADPH dynamics: Linking insulin resistance and β-Cells ferroptosis in diabetes mellitus. Int. J. Mol. Sci. 2023, 25, 342. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride metabolism in the liver. Compr. Physiol. 2017, 8, 1. [Google Scholar] [CrossRef]
- Porseryd, T.; Caspillo, N.R.; Volkova, K.; Elabbas, L.; Källman, T.; Dinnétz, P.; Olsson, P.-E.; Porsch-Hällström, I. Testis transcriptome alterations in zebrafish (Danio rerio) with reduced fertility due to developmental exposure to 17α-ethinyl estradiol. Gen. Comp. Endocrinol. 2018, 262, 44–58. [Google Scholar] [CrossRef]
- Wei, Y.; Tong, Y.; Zhang, Y. New mechanisms for bacterial degradation of sulfoquinovose. Biosci. Rep. 2022, 42, BSR20220314. [Google Scholar] [CrossRef] [PubMed]
- Alkazemi, D.; Rahman, A.; Habra, B. Alterations in glutathione redox homeostasis among adolescents with obesity and anemia. Sci. Rep. 2021, 11, 3034. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ahn, Y.J.; Asmis, R. Sexual dimorphism in glutathione metabolism and glutathione-dependent responses. Redox Biol. 2020, 31, 101410. [Google Scholar] [CrossRef]
- Mavralc, G.O.P.K.N.; Korund, Z.E.U.; Kaplanf, E.A.S. Exhibit Anti-Inflammatory, Antioxidant, and Anti-Androgen Effect in Rat Model of Experimental Polycystic Ovarian Syndrome. Med. Princ. Pract. 2024, 33, 385–391. [Google Scholar]
- Kamigaki, M.; Sakaue, S.; Tsujino, I.; Ohira, H.; Ikeda, D.; Itoh, N.; Ishimaru, S.; Ohtsuka, Y.; Nishimura, M. Oxidative stress provokes atherogenic changes in adipokine gene expression in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2006, 339, 624–632. [Google Scholar] [CrossRef]
- Horvath, T.L.; Andrews, Z.B.; Diano, S. Fuel utilization by hypothalamic neurons: Roles for ROS. Trends Endocrinol. Metab. 2009, 20, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhou, Y.; Zhu, L.; Wang, Z.; Ma, L.; He, Y.; Fu, P. Comparative metagenomic analysis of the vaginal microbiome in healthy women. Synth. Syst. Biotechnol. 2021, 6, 77–84. [Google Scholar] [CrossRef]
- Feng, T.; Zhao, X.; Gu, P.; Yang, W.; Wang, C.; Guo, Q.; Long, Q.; Liu, Q.; Cheng, Y.; Li, J. Adipocyte-derived lactate is a signalling metabolite that potentiates adipose macrophage inflammation via targeting PHD2. Nat. Commun. 2022, 13, 5208. [Google Scholar] [CrossRef]
- Brooks, G.A.; Arevalo, J.A.; Osmond, A.D.; Leija, R.G.; Curl, C.C.; Tovar, A.P. Lactate in contemporary biology: A phoenix risen. J. Physiol. 2022, 600, 1229–1251. [Google Scholar] [CrossRef]
- Liu, C.; Wu, J.; Zhu, J.; Kuei, C.; Yu, J.; Shelton, J.; Sutton, S.W.; Li, X.; Yun, S.J.; Mirzadegan, T. Lactate inhibits lipolysis in fat cells through activation of an orphan G-protein-coupled receptor, GPR81. J. Biol. Chem. 2009, 284, 2811–2822. [Google Scholar] [CrossRef]
- Ahmed, K.; Tunaru, S.; Tang, C.; Müller, M.; Gille, A.; Sassmann, A.; Hanson, J.; Offermanns, S. An autocrine lactate loop mediates insulin-dependent inhibition of lipolysis through GPR81. Cell Metab. 2010, 11, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Bai, M.; Wang, S.; Chen, L.; Li, Z.; Li, C.; Cao, P.; Chen, Y. Lactate is a key mediator that links obesity to insulin resistance via modulating cytokine production from adipose tissue. Diabetes 2022, 71, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.N.; Fan, T.W. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef] [PubMed]
- Anastassakis, K. Androgenetic Alopecia From A to Z: Vol. 2 Drugs, Herbs, Nutrition and Supplements; Springer Nature: Berlin, Germany, 2022. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward MR Analysis | Reserve MR Analysis | ||||||
|---|---|---|---|---|---|---|---|
| Exposure | Outcome | SNPs | OR (95% CI) | PIVW | PHeterogeneity | Ppleiotropy | PIVW |
| PWY-7456: Mannan Degradation | Obesity | 7 | 0.921 (0.818–1.038) | 0.176 | 0.580 | 0.897 | 0.852 |
| PWY-5989: Stearate Biosynthesis II (Bacteria and Plants) | Obesity | 8 | 0.975 (0.872–1.091) | 0.661 | 0.509 | 0.110 | 0.482 |
| s. Eubacterium eligens | Obesity | 13 | 0.961 (0.854–1.082) | 0.512 | 0.038 | 0.319 | 0.873 |
| PWY-5505: L-Glutamate and L-Glutamine Biosynthesis | Obesity | 16 | 1.008 (0.931–1.093) | 0.839 | 0.127 | 0.186 | 0.437 |
| PWY-5791: 1,4-Dihydroxy-2-naphthoate Biosynthesis II (Plants) | Obesity | 16 | 0.999 (0.930–1.074) | 0.988 | 0.102 | 0.245 | 0.133 |
| s. Clostridium leptum | Obesity | 7 | 1.037 (0.960–1.120) | 0.356 | 0.805 | 0.464 | 0.274 |
| PWY-7446: Sulfoglycolysis | Obesity | 10 | 1.066 (1.016–1.119) | 0.009 | 0.669 | 0.715 | 0.464 |
| s. Bifidobacterium bifidum | Obesity | 16 | 1.030 (0.985–1.076) | 0.191 | 0.744 | 0.692 | 0.641 |
| PWY-5850: Superpathway of Menaquinol-6 Biosynthesis I | Obesity | 11 | 1.001 (0.947–1.058) | 0.971 | 0.192 | 0.577 | 0.073 |
| PWY-5971: Palmitate Biosynthesis II (Bacteria and Plants) | Obesity | 14 | 0.975 (0.872–1.091) | 0.661 | 0.072 | 0.065 | 0.923 |
| PWY-6700: Queuosine Biosynthesis | Obesity | 16 | 1.019 (0.933–1.113) | 0.678 | 0.555 | 0.489 | 0.433 |
| s. Bacteroides xylanisolvens | Obesity | 13 | 0.951 (0.870–1.039) | 0.268 | 0.221 | 0.728 | 0.615 |
| PWY-7219: Adenosine Ribonucleotide De Novo Biosynthesis | Obesity | 13 | 1.115 (1.012–1.229) | 0.028 | 0.935 | 0.831 | 0.432 |
| g. Barnesiella | Obesity | 14 | 1.078 (0.990–1.173) | 0.082 | 0.405 | 0.122 | 0.297 |
| s. Barnesiella intestinihominis | Obesity | 13 | 1.076 (0.984–1.176) | 0.110 | 0.373 | 0.177 | 0.295 |
| PWY-5188: Tetrapyrrole Biosynthesis I (from Glutamate) | Obesity | 11 | 1.030 (0.934–1.137) | 0.553 | 0.693 | 0.955 | 0.811 |
| PWY-5695: Urate Biosynthesis/Inosine-5′-Phosphate Degradation | Obesity | 11 | 0.999 (0.889–1.122) | 0.984 | 0.282 | 0.356 | 0.778 |
| Forward MR Analysis | Reserve MR Analysis | ||||||
|---|---|---|---|---|---|---|---|
| Exposure | Outcome | SNPs | OR (95% CI) | PIVW | PHeterogeneity | Ppleiotropy | PIVW |
| Obesity | AGA | 20 | 1.050 (0.994~1.110) | 0.08 | 0.181 | 0.534 | 0.124 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Liao, X.; Tang, S.; Wang, Q.; Lin, H.; Yu, X.; Xiao, Y.; Tao, X.; Zhong, T. Potential Therapeutic Targets for Androgenetic Alopecia (AGA) in Obese Individuals as Revealed by a Gut Microbiome Analysis: A Mendelian Randomization Study. Nutrients 2025, 17, 1892. https://doi.org/10.3390/nu17111892
Li Y, Liao X, Tang S, Wang Q, Lin H, Yu X, Xiao Y, Tao X, Zhong T. Potential Therapeutic Targets for Androgenetic Alopecia (AGA) in Obese Individuals as Revealed by a Gut Microbiome Analysis: A Mendelian Randomization Study. Nutrients. 2025; 17(11):1892. https://doi.org/10.3390/nu17111892
Chicago/Turabian StyleLi, Yongwei, Xi Liao, Siwen Tang, Qian Wang, Heng Lin, Xi Yu, Ying Xiao, Xiaoyu Tao, and Tian Zhong. 2025. "Potential Therapeutic Targets for Androgenetic Alopecia (AGA) in Obese Individuals as Revealed by a Gut Microbiome Analysis: A Mendelian Randomization Study" Nutrients 17, no. 11: 1892. https://doi.org/10.3390/nu17111892
APA StyleLi, Y., Liao, X., Tang, S., Wang, Q., Lin, H., Yu, X., Xiao, Y., Tao, X., & Zhong, T. (2025). Potential Therapeutic Targets for Androgenetic Alopecia (AGA) in Obese Individuals as Revealed by a Gut Microbiome Analysis: A Mendelian Randomization Study. Nutrients, 17(11), 1892. https://doi.org/10.3390/nu17111892