
Efficient Enrichment of Docosahexaenoic Acid (DHA) in Mother’s Milk and in the Brain and Retina of the Offspring by Lysophosphatidylcholine (LPC)-DHA in the Maternal Diet
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Diets, Animals, and Procedures
2.1.1. Milk Collection
2.1.2. Tissue Collection
2.2. Analysis of Fatty Acids
3. Results
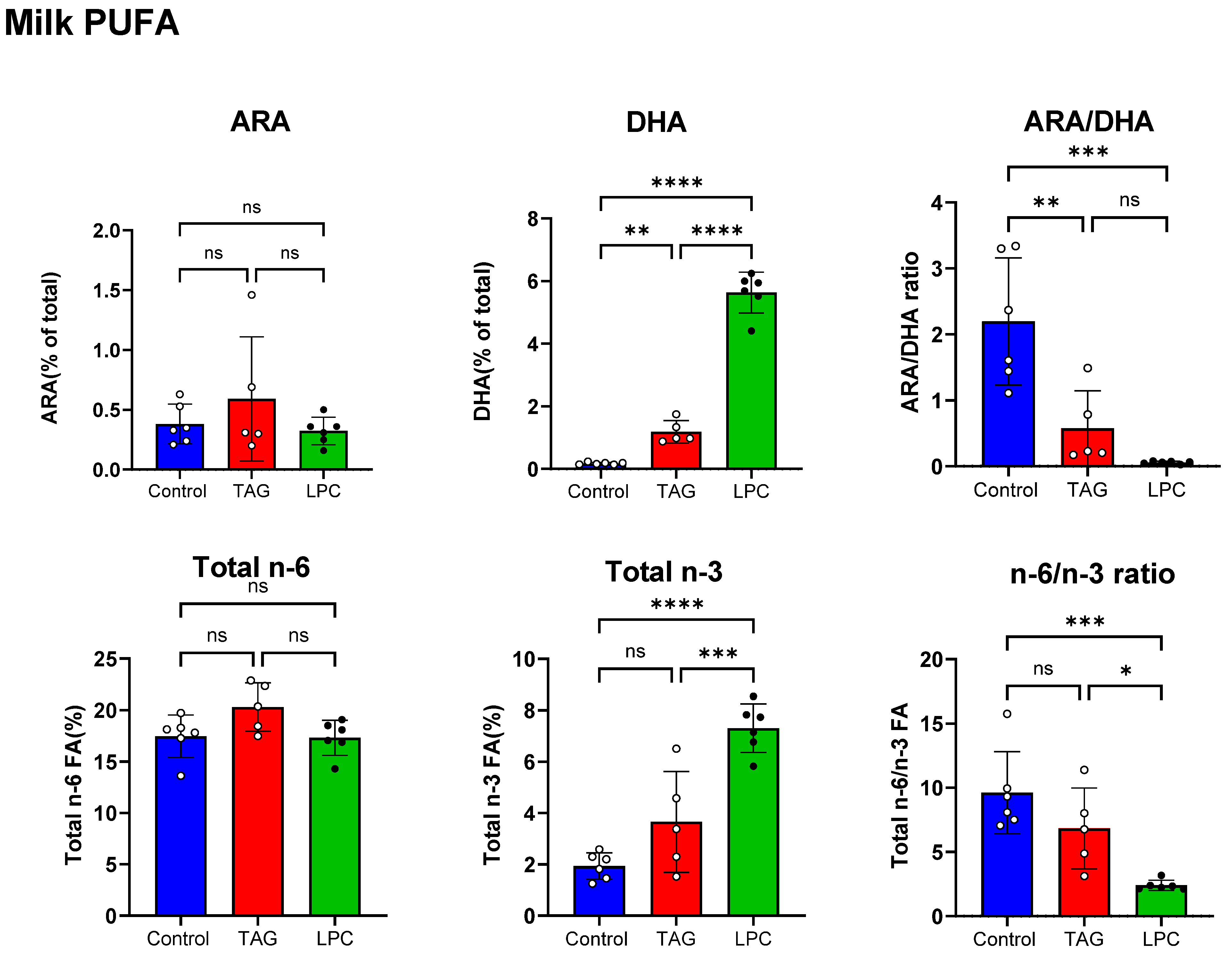
3.1. Effect of Dietary TAG-DHA and LPC-DHA on Fatty Acid Composition of Milk
3.2. Effect of TAG-DHA and LPC-DHA in Maternal Diet on Pup Brain Fatty Acid Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clandinin, M.T.; Chappell, J.E.; Leong, S.; Heim, T.; Swyer, P.R.; Chance, G.W. Intrauterine fatty acid accretion rates in human brain: Implications for fatty acid requirements. Early Hum. Dev. 1980, 4, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Mitguard, S.; Doucette, O.; Miklavcic, J. Human milk polyunsaturated fatty acids are related to neurodevelopmental, anthropometric, and allergic outcomes in early life: A systematic review. J. Dev. Orig. Health Dis. 2023, 14, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T. Animal studies of the functional consequences of suboptimal polyunsaturated fatty acid status during pregnancy, lactation and early post-natal life. Matern. Child Nutr. 2011, 7, 59–79. [Google Scholar] [CrossRef]
- Oguro, A.; Fujiyama, T.; Ishihara, Y.; Kataoka, C.; Yamamoto, M.; Eto, K.; Komohara, Y.; Imaoka, S.; Sakuragi, T.; Tsuji, M.; et al. Maternal DHA intake in mice increased DHA metabolites in the pup brain and ameliorated MeHg-induced behavioral disorder. J. Lipid Res. 2023, 64, 100458. [Google Scholar] [CrossRef]
- Dinel, A.L.; Rey, C.; Bonhomme, C.; Le Ruyet, P.; Joffre, C.; Layé, S. Dairy fat blend improves brain DHA and neuroplasticity and regulates corticosterone in mice. Prostaglandins Leukot. Essent. Fat. Acids 2016, 109, 29–38. [Google Scholar] [CrossRef]
- Lassek, W.D.; Gaulin, S.J.C. Maternal milk DHA content predicts cognitive performance in a sample of 28 nations. Matern. Child Nutr. 2015, 11, 773–779. [Google Scholar] [CrossRef]
- Hadley, K.B.; Ryan, A.S.; Forsyth, S.; Gautier, S.; Salem, N., Jr. The Essentiality of Arachidonic Acid in Infant Development. Nutrients 2016, 8, 216. [Google Scholar] [CrossRef]
- Brenna, J.T.; Varamini, B.; Jensen, R.G.; Diersen-Schade, D.A.; Boettcher, J.A.; Arterburn, L.M. Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am. J. Clin. Nutr. 2007, 85, 1457–1464. [Google Scholar] [CrossRef]
- Yuhas, R.; Pramuk, K.; Lien, E.L. Human milk fatty acid composition from nine countries varies most in DHA. Lipids 2006, 41, 851–858. [Google Scholar] [CrossRef]
- Delgado-Noguera, M.F.; Calvache, J.A.; Bonfill Cosp, X.; Kotanidou, E.P.; Galli-Tsinopoulou, A. Supplementation with long chain polyunsaturated fatty acids (LCPUFA) to breastfeeding mothers for improving child growth and development. Cochrane Database Syst. Rev. 2015, 2015, Cd007901. [Google Scholar] [CrossRef]
- Jasani, B.; Simmer, K.; Patole, S.K.; Rao, S.C. Long chain polyunsaturated fatty acid supplementation in infants born at term. Cochrane Database Syst. Rev. 2017, 3, CD000376. [Google Scholar] [CrossRef] [PubMed]
- Ulven, S.M.; Holven, K.B. Comparison of bioavailability of krill oil versus fish oil and health effect. Vasc. Health Risk Manag. 2015, 11, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Sugasini, D.; Yalagala, P.C.R.; Goggin, A.; Tai, L.M.; Subbaiah, P.V. Enrichment of brain docosahexaenoic acid (DHA) is highly dependent upon the molecular carrier of dietary DHA: Lysophosphatidylcholine is more efficient than either phosphatidylcholine or triacylglycerol. J. Nutr. Biochem. 2019, 74, 108231. [Google Scholar] [CrossRef] [PubMed]
- Yalagala, P.C.R.; Sugasini, D.; Zaldua, S.B.; Tai, L.M.; Subbaiah, P.V. Lipase treatment of dietary krill oil, but not fish oil, enables enrichment of brain eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). Mol. Nutrtion Food Res. 2020, 64, e2000059. [Google Scholar] [CrossRef]
- Sugasini, D.; Thomas, R.; Yalagala, P.C.R.; Tai, L.M.; Subbaiah, P.V. Dietary docosahexaenoic acid (DHA) as lysophosphatidylcholine, but not as free acid, enriches brain DHA and improves memory in adult mice. Sci. Rep. 2017, 7, 11263. [Google Scholar] [CrossRef]
- Wong, B.H.; Silver, D.L. Mfsd2a: A Physiologically Important Lysolipid Transporter in the Brain and Eye. Adv. Exp. Med. Biol. 2020, 1276, 223–234. [Google Scholar] [CrossRef]
- Sugasini, D.; Park, J.C.; McAnany, J.J.; Kim, T.H.; Ma, G.; Yao, X.; Antharavally, B.; Oroskar, A.; Oroskar, A.A.; Layden, B.T.; et al. Improvement of retinal function in Alzheimer disease-associated retinopathy by dietary lysophosphatidylcholine-EPA/DHA. Sci. Rep. 2023, 13, 9179. [Google Scholar] [CrossRef]
- Sugasini, D.; Yalagala, P.C.R.; Subbaiah, P.V. Efficient Enrichment of Retinal DHA with Dietary Lysophosphatidylcholine-DHA: Potential Application for Retinopathies. Nutrients 2020, 12, 3114. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Oosting, A.; Verkade, H.J.; Kegler, D.; van de Heijning, B.J.; van der Beek, E.M. Rapid and selective manipulation of milk fatty acid composition in mice through the maternal diet during lactation. J. Nutr. Sci. 2015, 4, e19. [Google Scholar] [CrossRef]
- Kao, B.T.; DePeters, E.J.; Van Eenennaam, A.L. Mice raised on milk transgenically enriched with n-3 PUFA have increased brain docosahexaenoic acid. Lipids 2006, 41, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, A.; Nieto, S.; Sanhueza, J.; Nuñez, M.J.; Ferrer, C. Tissue accretion and milk content of docosahexaenoic acid in female rats after supplementation with different docosahexaenoic acid sources. Ann. Nutr. Metab. 2005, 49, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Fidler, N.; Sauerwald, T.; Pohl, A.; Demmelmair, H.; Koletzko, B. Docosahexaenoic acid transfer into human milk after dietary supplementation: A randomized clinical trial. J. Lipid Res. 2000, 41, 1376–1383. [Google Scholar] [CrossRef]
- Macura, I.J.; Djuricic, I.; Major, T.; Milanovic, D.; Sobajic, S.; Kanazir, S.; Ivkovic, S. The supplementation of a high dose of fish oil during pregnancy and lactation led to an elevation in Mfsd2a expression without any changes in docosahexaenoic acid levels in the retina of healthy 2-month-old mouse offspring. Front. Nutr. 2023, 10, 1330414. [Google Scholar] [CrossRef]
- Sherry, C.L.; Oliver, J.S.; Marriage, B.J. Docosahexaenoic acid supplementation in lactating women increases breast milk and plasma docosahexaenoic acid concentrations and alters infant omega 6:3 fatty acid ratio. Prostaglandins Leukot. Essent. Fat. Acids 2015, 95, 63–69. [Google Scholar] [CrossRef]
- Gould, J.F.; Bednarz, J.M.; Sullivan, T.R.; McPhee, A.J.; Gibson, R.A.; Makrides, M. Subgroup analyses of a randomized trial of DHA supplementation for infants born preterm with assessments of cognitive development up to 7-years of age: What happens in infants born <29 weeks’ gestation? Prostaglandins Leukot. Essent. Fat. Acids 2023, 198–199, 102593. [Google Scholar] [CrossRef]
- Makrides, M.; Gibson, R.A.; McPhee, A.J.; Collins, C.T.; Davis, P.G.; Doyle, L.W.; Simmer, K.; Colditz, P.B.; Morris, S.; Smithers, L.G.; et al. Neurodevelopmental outcomes of preterm infants fed high-dose docosahexaenoic acid: A randomized controlled trial. JAMA 2009, 301, 175–182. [Google Scholar] [CrossRef]
- Luxwolda, M.F.; Kuipers, R.S.; Koops, J.H.; Muller, S.; de Graaf, D.; Dijck-Brouwer, D.A.; Muskiet, F.A. Interrelationships between maternal DHA in erythrocytes, milk and adipose tissue. Is 1 wt% DHA the optimal human milk content? Data from four Tanzanian tribes differing in lifetime stable intakes of fish. Br. J. Nutr. 2014, 111, 854–866. [Google Scholar] [CrossRef]
- Ureta-Velasco, N.; Montealegre-Pomar, A.; Keller, K.; Escuder-Vieco, D.; Fontecha, J.; Calvo, M.V.; Megino-Tello, J.; Serrano, J.C.E.; García-Lara, N.R.; Pallás-Alonso, C.R. Associations of Dietary Intake and Nutrient Status with Micronutrient and Lipid Composition in Breast Milk of Donor Women. Nutrients 2023, 15, 3486. [Google Scholar] [CrossRef]
- Harris, W.S.; Connor, W.E.; Lindsey, S. Will dietary omega-3 fatty acids change the composition of human milk? Am. J. Clin. Nutr. 1984, 40, 780–785. [Google Scholar] [CrossRef]
- Liscano, Y.; Sanchez-Palacio, N. A Critical Look at Omega-3 Supplementation: A Thematic Review. Healthcare 2023, 11, 3065. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, S.I. Brain arachidonic and docosahexaenoic acid cascades are selectively altered by drugs, diet and disease. Prostaglandins Leukot. Essent. Fat Acids 2008, 79, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.; Zhao, A.; Yang, B.; Zhang, Y.; Linderborg, K.M. Tissue-Specific Content of Polyunsaturated Fatty Acids in (n-3) Deficiency State of Rats. Foods 2022, 11, 208. [Google Scholar] [CrossRef]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, 1467s–1476s. [Google Scholar] [CrossRef]
- Beligere, N.; Perumalswamy, V.; Tandon, M.; Mittal, A.; Floora, J.; Vijayakumar, B.; Miller, M.T. Retinopathy of prematurity and neurodevelopmental disabilities in premature infants. Semin. Fetal Neonatal Med. 2015, 20, 346–353. [Google Scholar] [CrossRef]
- Connor, K.M.; SanGiovanni, J.P.; Lofqvist, C.; Aderman, C.M.; Chen, J.; Higuchi, A.; Hong, S.; Pravda, E.A.; Majchrzak, S.; Carper, D.; et al. Increased dietary intake of -ë-3-polyunsaturated fatty acids reduces pathological retinal angiogenesis. Nat. Med. 2007, 13, 868. [Google Scholar] [CrossRef]
- Carnielli, V.P.; Verlato, G.; Pederzini, F.; Luijendijk, I.; Boerlage, A.; Pedrotti, D.; Sauer, P.J. Intestinal absorption of long-chain polyunsaturated fatty acids in preterm infants fed breast milk or formula. Am. J. Clin. Nutr. 1998, 67, 97–103. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yalagala, P.C.R.; Sugasini, D.; Chantapim, S.; Caal, K.; Sun, H.; Nicastro, S.; Sargis, R.M.; Gregg, B.; Subbaiah, P.V. Efficient Enrichment of Docosahexaenoic Acid (DHA) in Mother’s Milk and in the Brain and Retina of the Offspring by Lysophosphatidylcholine (LPC)-DHA in the Maternal Diet. Nutrients 2025, 17, 1864. https://doi.org/10.3390/nu17111864
Yalagala PCR, Sugasini D, Chantapim S, Caal K, Sun H, Nicastro S, Sargis RM, Gregg B, Subbaiah PV. Efficient Enrichment of Docosahexaenoic Acid (DHA) in Mother’s Milk and in the Brain and Retina of the Offspring by Lysophosphatidylcholine (LPC)-DHA in the Maternal Diet. Nutrients. 2025; 17(11):1864. https://doi.org/10.3390/nu17111864
Chicago/Turabian StyleYalagala, Poorna C. R., Dhavamani Sugasini, Sutape Chantapim, Karyna Caal, Haijing Sun, Sofia Nicastro, Robert M. Sargis, Brigid Gregg, and Papasani V. Subbaiah. 2025. "Efficient Enrichment of Docosahexaenoic Acid (DHA) in Mother’s Milk and in the Brain and Retina of the Offspring by Lysophosphatidylcholine (LPC)-DHA in the Maternal Diet" Nutrients 17, no. 11: 1864. https://doi.org/10.3390/nu17111864
APA StyleYalagala, P. C. R., Sugasini, D., Chantapim, S., Caal, K., Sun, H., Nicastro, S., Sargis, R. M., Gregg, B., & Subbaiah, P. V. (2025). Efficient Enrichment of Docosahexaenoic Acid (DHA) in Mother’s Milk and in the Brain and Retina of the Offspring by Lysophosphatidylcholine (LPC)-DHA in the Maternal Diet. Nutrients, 17(11), 1864. https://doi.org/10.3390/nu17111864