

Ligilactobacillus salivarius CECT5713 Increases Term Pregnancies in Women with Infertility of Unknown Origin: A Randomized, Triple-Blind, Placebo-Controlled Trial

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Sample Size Estimation
2.3. Study Population
2.4. Randomization
2.5. Study Intervention
2.6. Primary and Secondary Outcomes of the Study
2.7. Culture-Dependent Analysis of Vaginal Swabs and Semen Samples
2.8. DNA Extraction from Vaginal Swabs and Semen Samples
2.9. Quantification of L. salivarius DNA in Vaginal Samples
2.10. Metataxonomic Profiling
2.11. Immunological Analyses in Vaginal and Semen Samples
2.12. Statistical Analysis
3. Results
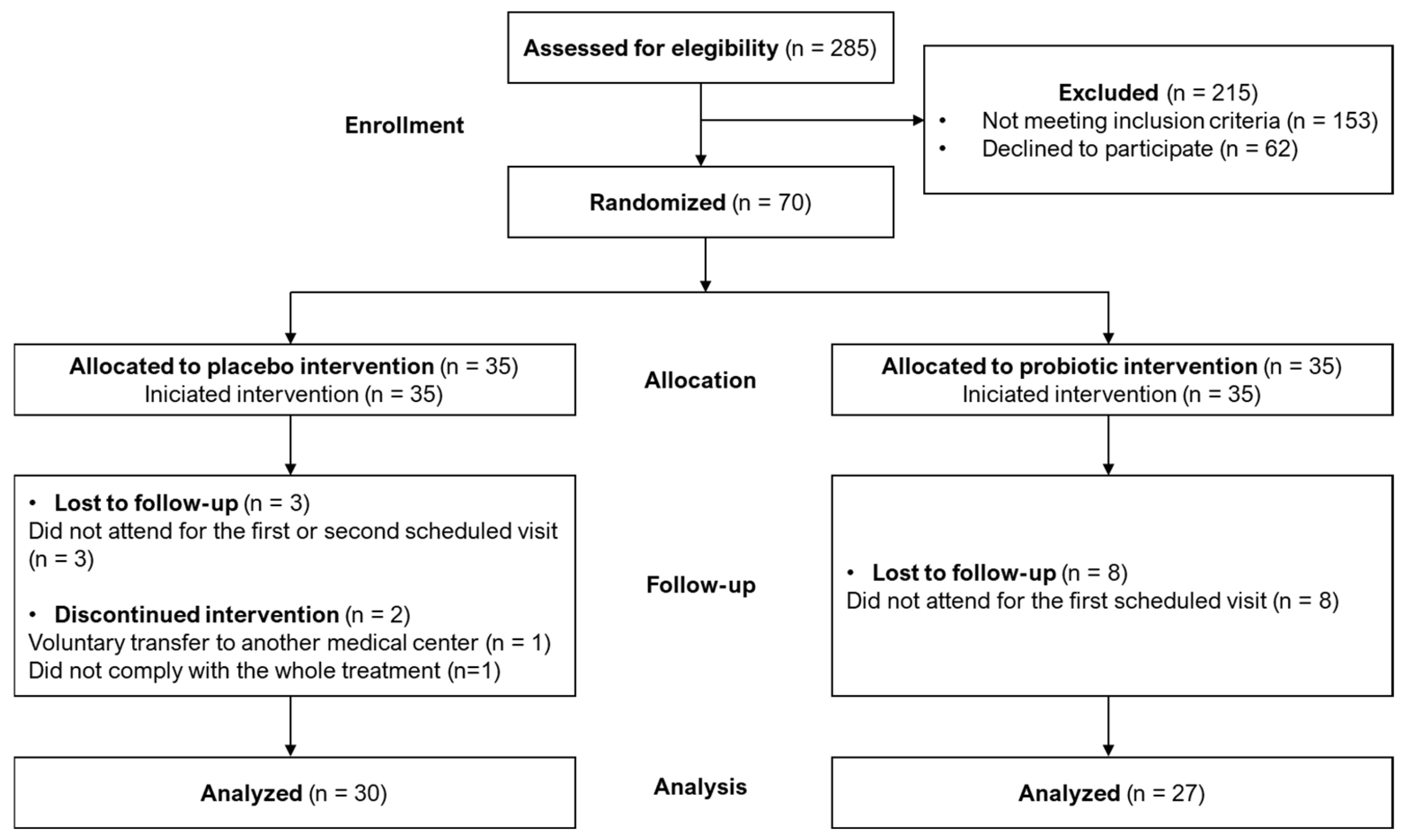
3.1. Study Randomization
3.2. Characteristics of the Participants
3.3. Pregnancy Success and Other Clinical Outcomes
3.4. Other Secondary Outcomes
3.4.1. Microbial Characterization of Vaginal Exudates Using Culture-Dependent Methods
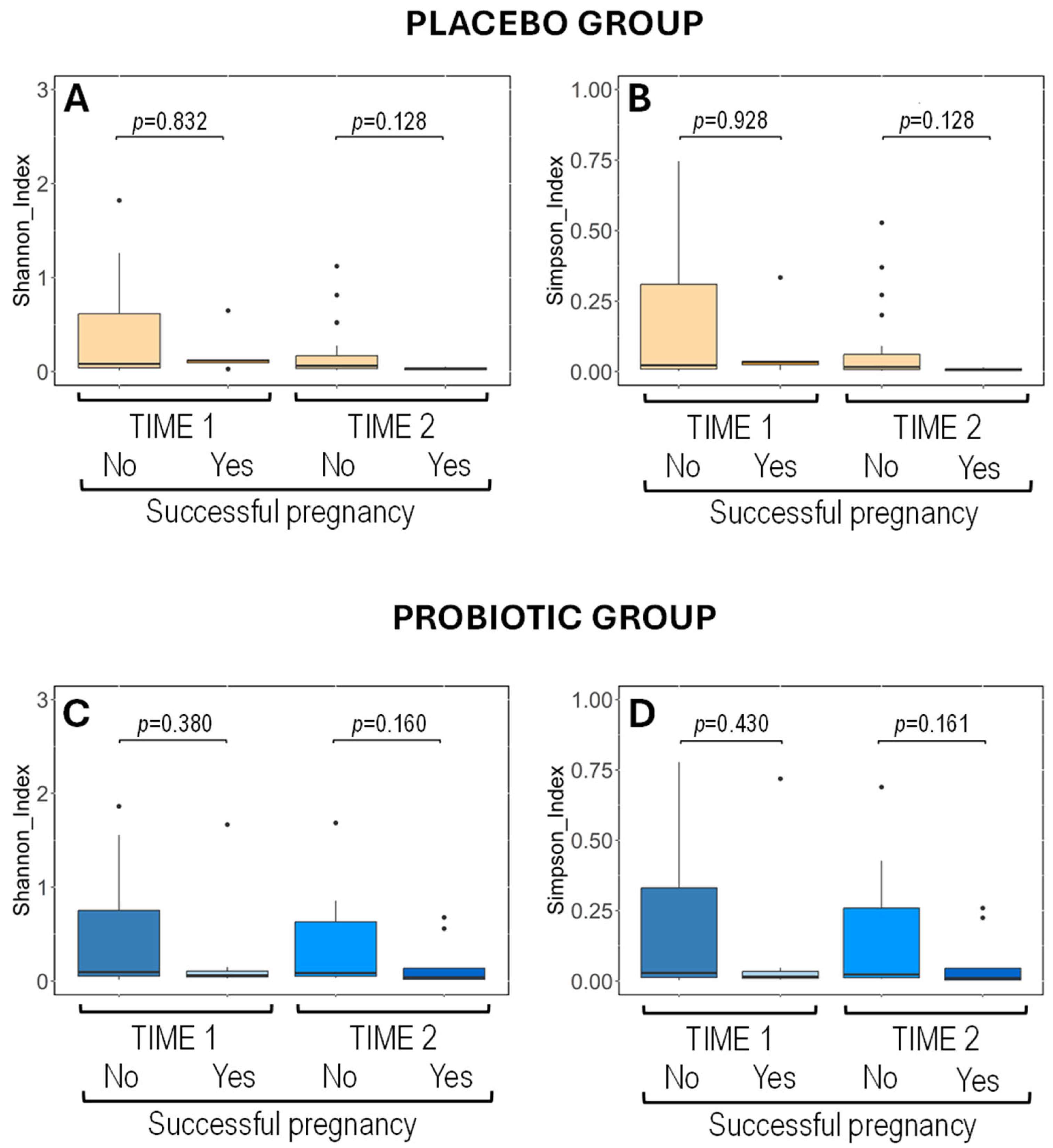
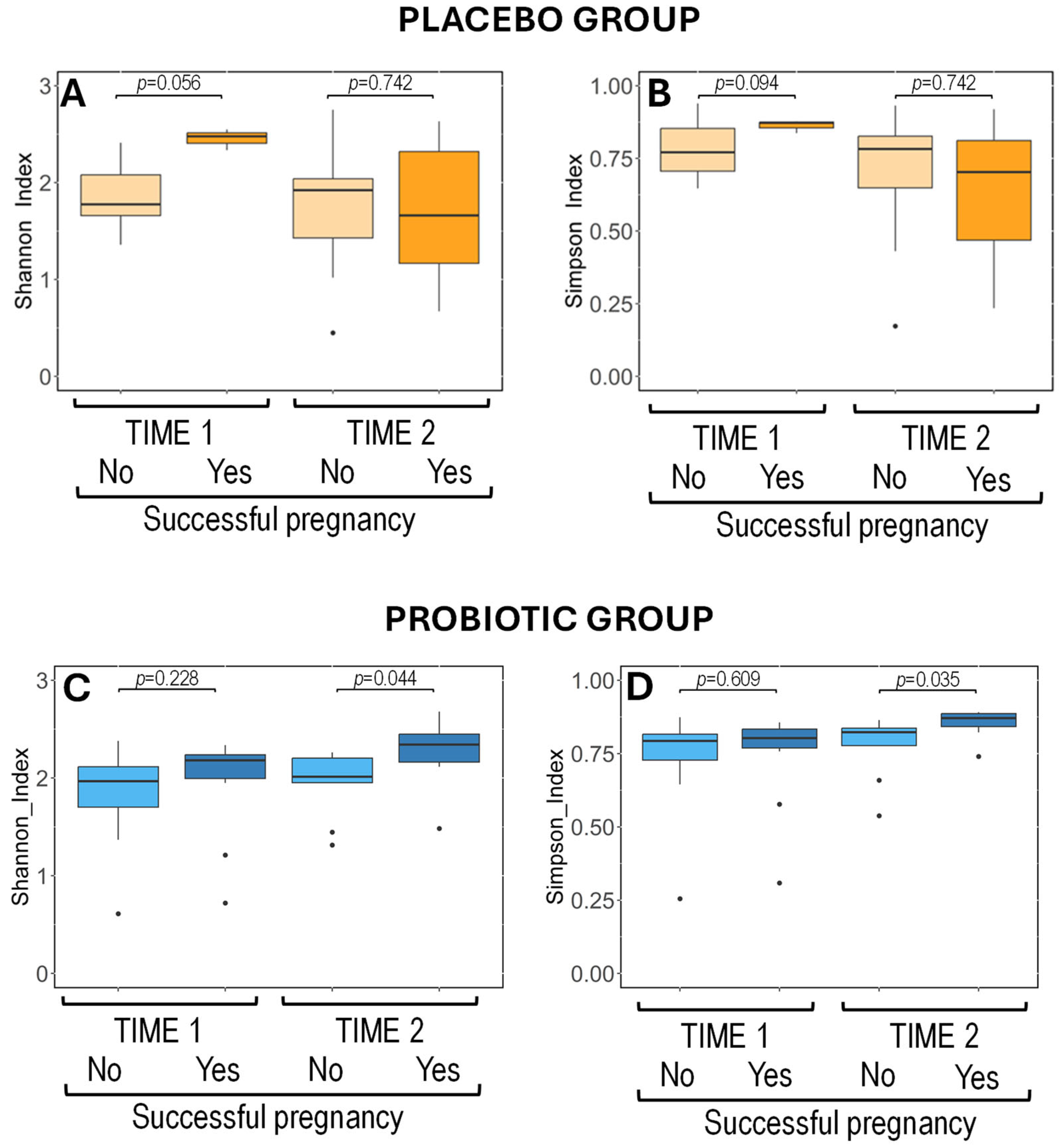
3.4.2. Metataxonomic Analysis of Vaginal Exudates
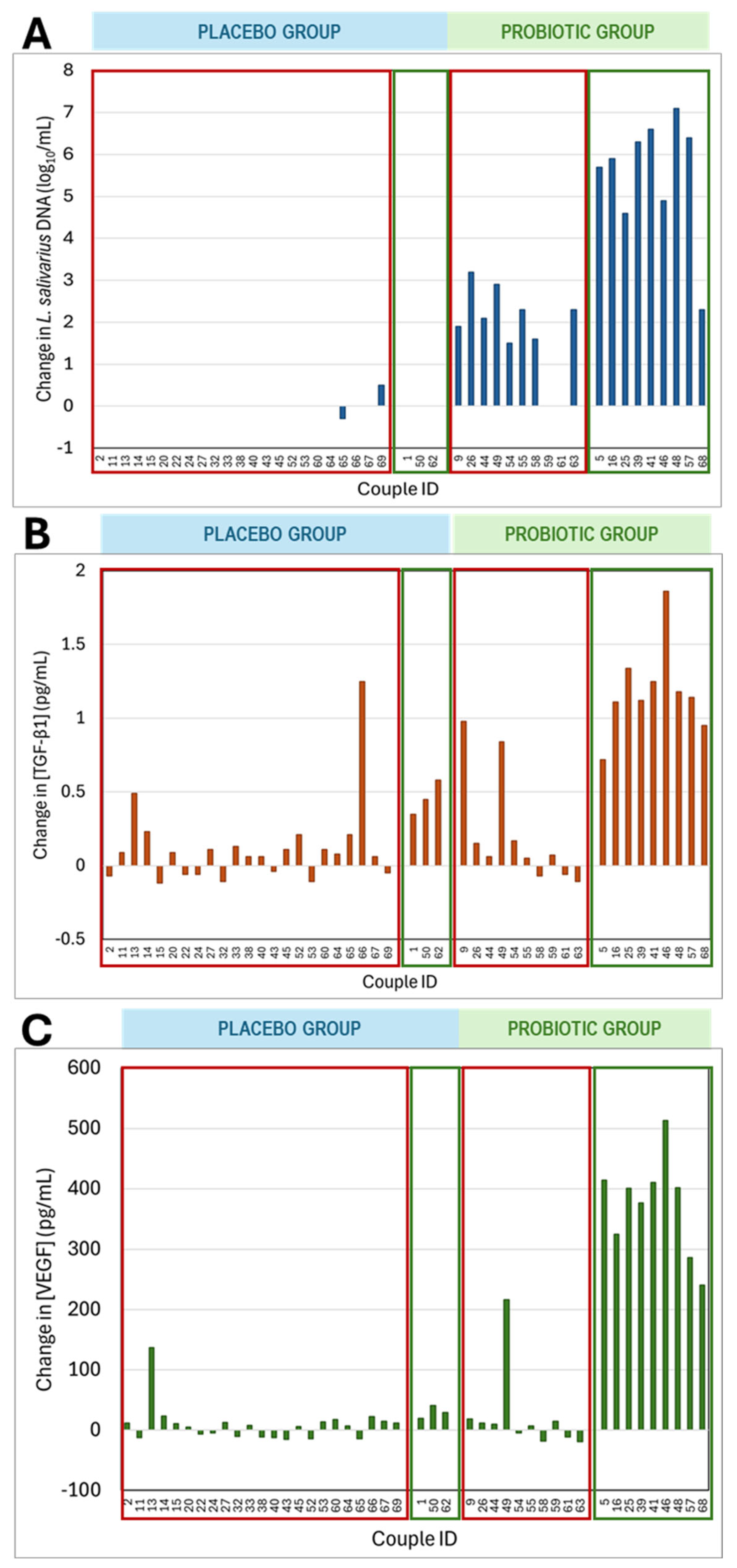
3.4.3. Detection of L. salivarius in Vaginal Exudates by RT qPCR Analysis
3.4.4. Immunological Characteristics of Vaginal Samples
3.4.5. Microbial Characterization of Semen Samples Using Culture-Dependent Methods
3.4.6. Metataxonomic Analysis of Semen Samples
3.4.7. Immunological Characteristics of Semen Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IVF | In vitro fertilization |
| TGFβ1 | Transforming growth factor β1 |
| VEGF | Vascular endothelial growth factor |
| IL | Interleukin |
| GM-CSF | Granulocyte macrophage colony-stimulating factor |
| IFNγ | Interferon γ |
| TNFα | Tumor necrosis factor α |
References
- Inhorn, M.C.; Patrizio, P. Infertility around the globe: New thinking on gender, reproductive technologies and global movements in the 21st century. Hum. Reprod. Update 2015, 21, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Boivin, J.; Bunting, L.; Collins, J.A.; Nygren, K.G. International estimates of infertility prevalence and treatment-seeking: Potential need and demand for infertility medical care. Hum. Reprod. 2007, 22, 1506–1512. [Google Scholar] [CrossRef] [PubMed]
- Carson, S.A.; Kallen, A.N. Diagnosis and management of infertility: A review. JAMA 2021, 326, 65–76. [Google Scholar] [CrossRef] [PubMed]
- WHO. ICD-11. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/infertility (accessed on 22 May 2025).
- Sirota, I.; Zarek, S.M.; Segars, J.H. Potential influence of the microbiome on infertility and assisted reproductive technology. Semin. Reprod. Med. 2014, 32, 35–42. [Google Scholar] [CrossRef]
- Campisciano, G.; Florian, F.; D’Eustacchio, A.; Stanković, D.; Ricci, G.; De Seta, F.; Comar, M. Subclinical alteration of the cervical-vaginal microbiome in women with idiopathic infertility. J. Cell. Physiol. 2017, 232, 1681–1688. [Google Scholar] [CrossRef]
- Hong, X.; Ma, J.; Yin, J.; Fang, S.; Geng, J.; Zhao, H.; Zhu, M.; Ye, M.; Zhu, X.; Xuan, Y.; et al. The association between vaginal microbiota and female infertility: A systematic review and meta-analysis. Arch. Gynecol. Obstet. 2020, 302, 569–578. [Google Scholar] [CrossRef]
- Reid, G.; Brigidi, P.; Burton, J.P.; Contractor, N.; Duncan, S.; Fargier, E.; Hill, C.; Lebeer, S.; Martín, R.; McBain, A.J.; et al. Microbes central to human reproduction. Am. J. Reprod. Immunol. 2015, 73, 1–11. [Google Scholar] [CrossRef]
- Moreno, I.; Simón, C. Deciphering the effect of reproductive tract microbiota on human reproduction. Reprod. Med. Biol. 2019, 18, 40–50. [Google Scholar] [CrossRef]
- Vitale, S.G.; Ferrari, F.; Ciebiera, M.; Zgliczyńska, M.; Rapisarda, A.M.C.; Vecchio, G.M.; Pino, A.; Angelico, G.; Knafel, A.; Riemma, G.; et al. The role of genital tract microbiome in fertility: A systematic review. Int. J. Mol. Sci. 2021, 23, 180. [Google Scholar] [CrossRef]
- Reid, G.; Younes, J.A.; Van der Mei, H.C.; Gloor, G.B.; Knight, R.; Busscher, H.J. Microbiota restoration: Natural and supplemented recovery of human microbial communities. Nat. Rev. Microbiol. 2011, 9, 27–38. [Google Scholar] [CrossRef]
- Tsonis, O.; Gkrozou, F.; Paschopoulos, M. Microbiome affecting reproductive outcome in ARTs. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 102036. [Google Scholar] [CrossRef] [PubMed]
- Vidhate, P.; Wakchoure, P.; Borole, S.; Khan, A.A. Lactobacillus as probiotics: Opportunities and challenges for potential benefits in female reproductive health. Am. J. Transl. Res. 2024, 16, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Koot, Y.E.; Teklenburg, G.; Salker, M.S.; Brosens, J.J.; Macklon, N.S. Molecular aspects of implantation failure. Biochim. Biophys. Acta 2012, 1822, 1943–1950. [Google Scholar] [CrossRef]
- Ticconi, C.; Pietropolli, A.; Di Simone, N.; Piccione, E.; Fazleabas, A. Endometrial immune dysfunction in recurrent pregnancy loss. Int. J. Mol. Sci. 2019, 20, 5332. [Google Scholar] [CrossRef]
- Blancafort, C.; Llácer, J. Can probiotics enhance fertility outcome? Capacity of probiotics as a single intervention to improve the feminine genital tract microbiota in non-symptomatic reproductive-aged women. Front. Endocrinol. 2023, 13, 1081830. [Google Scholar] [CrossRef]
- Fernández, L.; Castro, I.; Arroyo, R.; Alba, C.; Beltrán, D.; Rodríguez, J.M. Application of Ligilactobacillus salivarius CECT5713 to achieve term pregnancies in women with repetitive abortion or infertility of unknown origin by microbiological and immunological modulation of the vaginal ecosystem. Nutrients 2021, 13, 162. [Google Scholar] [CrossRef]
- Martín, R.; Jiménez, E.; Olivares, M.; Marín, M.; Fernández, L.; Xaus, J.; Rodríguez, J. Lactobacillus salivarius CECT 5713, a potential probiotic strain isolated from infant feces and breast milk of a mother–child pair. Int. J. Food Microbiol. 2006, 112, 35–43. [Google Scholar] [CrossRef]
- Arroyo, R.; Martín, V.; Maldonado, A.; Jiménez, E.; Fernández, L.; Rodríguez, J.M. Treatment of infectious mastitis during lactation: Antibiotics versus oral administration of Lactobacilli isolated from breast milk. Clin. Infect. Dis. 2010, 50, 1551–1558. [Google Scholar] [CrossRef]
- Maldonado, J.; Lara-Villoslada, F.; Sierra, S.; Sempere, L.; Gómez, M.; Rodríguez, J.M.; Boza, J.; Xaus, J.; Olivares, M. Safety and tolerance of the human milk probiotic strain Lactobacillus salivarius CECT5713 in 6-month-old children. Nutrition 2010, 26, 1082–1087. [Google Scholar] [CrossRef]
- Farrington, C.P.; Manning, G. Test statistics and sample size formulae for comparative binomial trials with null hypothesis of non-zero risk difference or non-unity relative risk. Stat. Med. 1990, 9, 1447–1454. [Google Scholar] [CrossRef]
- Lackey, K.A.; Williams, J.E.; Meehan, C.L.; Zachek, J.A.; Benda, E.D.; Price, W.J.; Foster, J.A.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; et al. What’s normal? Microbiomes in human milk and infant feces are related to each other but vary geographically: The INSPIRE Study. Front. Nutr. 2019, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Harrow, S.A.; Ravindran, V.; Butler, R.C.; Marshall, J.W.; Tannock, G.W. Real-time quantitative PCR measurement of ileal Lactobacillus salivarius populations from broiler chickens to determine the influence of farming practices. Appl. Environ. Microbiol. 2007, 73, 7123–7127. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Zakrzewski, M.; Proietti, C.; Ellis, J.J.; Hasan, S.; Brion, M.J.; Berger, B.; Krause, L. Calypso: A user-friendly web-server for mining and visualizing microbiome-environment interactions. Bioinformatics 2017, 33, 782–783. [Google Scholar] [CrossRef]
- Moher, D.; Hopewell, S.; Schulz, K.F.; Montori, V.; Gøtzsche, P.C.; Devereaux, P.J.; Elbourne, D.; Egger, M.; Altman, D.G. CONSORT 2010 explanation and elaboration: Updated guidelines for reporting parallel group randomised trials. BMJ 2010, 340, c869. [Google Scholar] [CrossRef] [PubMed]
- Hyman, R.W.; Herndon, C.N.; Jiang, H.; Palm, C.; Fukushima, M.; Bernstein, D.; Vo, K.C.; Zelenko, Z.; Davis, R.W.; Giudice, L.C. The dynamics of the vaginal microbiome during infertility therapy with in vitro fertilization-embryo transfer. J. Assist. Reprod. Genet. 2012, 29, 105–115. [Google Scholar] [CrossRef]
- Haahr, T.; Jensen, J.S.; Thomsen, L.; Duus, L.; Rygaard, K.; Humaidan, P. Abnormal vaginal microbiota may be associated with poor reproductive outcomes: A prospective study in IVF patients. Hum. Reprod. 2016, 31, 795–803. [Google Scholar] [CrossRef]
- Koedooder, R.; Singer, M.; Schoenmakers, S.; Savelkoul, P.H.M.; Morré, S.A.; de Jonge, J.D.; Poort, L.; Cuypers, W.J.S.S.; Beckers, N.G.M.; Broekmans, F.J.M.; et al. The vaginal microbiome as a predictor for outcome of in vitro fertilization with or without intracytoplasmic sperm injection: A prospective study. Hum. Reprod. 2019, 34, 1042–1054. [Google Scholar] [CrossRef]
- Baud, A.; Hillion, K.H.; Plainvert, C.; Tessier, V.; Tazi, A.; Mandelbrot, L.; Poyart, C.; Kennedy, S.P. Microbial diversity in the vaginal microbiota and its link to pregnancy outcomes. Sci. Rep. 2023, 13, 9061. [Google Scholar] [CrossRef]
- Hertz, F.B.; Holm, J.B.; Pallejá, A.; Björnsdóttir, M.K.; Mikkelsen, L.S.; Brandsborg, E.; Frimodt-Møller, N. Vaginal microbiome following orally administered probiotic. APMIS 2022, 130, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Lyra, A.; Ala-Jaakkola, R.; Yeung, N.; Datta, N.; Evans, K.; Hibberd, A.; Lehtinen, M.J.; Forssten, S.D.; Ibarra, A.; Pesonen, T.; et al. A healthy vaginal microbiota remains stable during oral probiotic supplementation: A randomised controlled trial. Microorganisms 2023, 11, 499. [Google Scholar] [CrossRef] [PubMed]
- Oerlemans, E.; Ahannach, S.; Wittouck, S.; Dehay, E.; De Boeck, I.; Ballet, N.; Rodriguez, B.; Tuyaerts, I.; Lebeer, S. Impacts of menstruation, community type, and an oral yeast probiotic on the vaginal microbiome. mSphere 2022, 7, e0023922. [Google Scholar] [CrossRef]
- Yang, S.; Reid, G.; Challis, J.R.G.; Gloor, G.B.; Asztalos, E.; Money, D.; Seney, S.; Bocking, A.D. Effect of oral probiotic Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14 on the vaginal microbiota, cytokines and chemokines in pregnant women. Nutrients 2020, 12, 368. [Google Scholar] [CrossRef]
- Vujic, G.; Jajac Knez, A.; Despot Stefanovic, V.; Kuzmic Vrbanovic, V. Efficacy of orally applied probiotic capsules for bacterial vaginosis and other vaginal infections: A double-blind, randomized, placebo-controlled study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 168, 75–79. [Google Scholar] [CrossRef]
- Macklaim, J.M.; Clemente, J.C.; Knight, R.; Gloor, G.B.; Reid, G. Changes in vaginal microbiota following antimicrobial and probiotic therapy. Microb. Ecol. Health Dis. 2015, 26, 27799. [Google Scholar] [CrossRef]
- Wieërs, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; Philippart de Foy, J.M.; Dequenne, I.; de Timary, P.; Cani, P.D. How probiotics affect the microbiota. Front. Cell. Infect. Microbiol. 2020, 9, 454. [Google Scholar] [CrossRef]
- Merenstein, D.; Pot, B.; Leyer, G.; Ouwehand, A.C.; Preidis, G.A.; Elkins, C.A.; Hill, C.; Lewis, Z.T.; Shane, A.L.; Zmora, N.; et al. Emerging issues in probiotic safety: 2023 perspectives. Gut Microbes 2023, 15, 2185034. [Google Scholar] [CrossRef]
- Grazul, H.; Kanda, L.L.; Gondek, D. Impact of probiotic supplements on microbiome diversity following antibiotic treatment of mice. Gut Microbes 2016, 7, 101–114. [Google Scholar] [CrossRef]
- Kabbani, T.A.; Pallav, K.; Dowd, S.E.; Villafuerte-Galvez, J.; Vanga, R.R.; Castillo, N.E.; Hansen, J.; Dennis, M.; Leffler, D.A.; Kelly, C.P. Prospective randomized controlled study on the effects of Saccharomyces boulardii CNCM I-745 and amoxicillin-clavulanate or the combination on the gut microbiota of healthy volunteers. Gut Microbes 2017, 8, 17–32. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.Z.; Federici, S.; et al. Post-antibiotic gut mucosal microbiome reconstitution is impaired by probiotics and improved by autologous FMT. Cell 2018, 174, 1406–1423.e16. [Google Scholar] [CrossRef] [PubMed]
- Castagnoli, R.; Pala, F.; Bosticardo, M.; Licari, A.; Delmonte, O.M.; Villa, A.; Marseglia, G.L.; Notarangelo, L.D. Gut microbiota–host interactions in inborn errors of immunity. Int. J. Mol. Sci. 2021, 22, 1416. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Castro, I.; Arroyo, R.; Alba, C.; Beltrán, D.; Rodríguez, J.M. Immunomodulation of the vaginal ecosystem by Ligilactobacillus salivarius CECT 30632 improves pregnancy rates among women with infertility of unknown origin or habitual abortions. Nutrients 2023, 15, 362. [Google Scholar] [CrossRef]
- Guo, X.; Yi, H.; Li, T.C.; Wang, Y.; Wang, H.; Chen, X. Role of vascular Endothelial Growth Factor (VEGF) in Human Embryo Implantation: Clinical Implications. Biomolecules 2021, 11, 253. [Google Scholar] [CrossRef]
- Wen, B.; Liao, H.; Lin, W.; Li, Z.; Ma, X.; Xu, Q.; Yu, F. The role of TGF-β during pregnancy and pregnancy complications. Int. J. Mol. Sci. 2023, 24, 16882. [Google Scholar] [CrossRef]
- Qian, D.; Lin, H.-Y.; Wang, H.-M.; Zhang, X.; Liu, D.-L.; Li, Q.-L.; Zhu, C. Involvement of ERK1/2 pathway in TGF-beta1-induced VEGF secretion in normal human cytotrophoblast cells. Mol. Reprod. Dev. 2004, 68, 198–204. [Google Scholar] [CrossRef]
- Wahl, S.M.; Wen, J.; Moutsopoulos, N. TGF-beta: A mobile purveyor of immune privilege. Immunol. Rev. 2006, 213, 213–227. [Google Scholar] [CrossRef]
- Qian, J.; Zhang, N.; Lin, J.; Wang, C.; Pan, X.; Chen, L.; Li, D.; Wang, L. Distinct pattern of Th17/Treg cells in pregnant women with a history of unexplained recurrent spontaneous abortion. Biosci. Trends 2018, 12, 157–167. [Google Scholar] [CrossRef]
- Garmendia, J.V.; De Sanctis, C.V.; Hajdúch, M.; De Sanctis, J.B. Microbiota and Recurrent Pregnancy Loss (RPL); More than a Simple Connection. Microorganisms 2024, 12, 1641. [Google Scholar] [CrossRef]
- Syriou, V.; Papanikolaou, D.; Kozyraki, A.; Goulis, D.G. Cytokines and male infertility. Eur. Cytokine Netw. 2018, 29, 73–82. [Google Scholar] [CrossRef]
- Renu, K.; Subramaniam, M.D.; Chakraborty, R.; Myakala, H.; Iyer, M.; Bharathi, G.; Siva, K.; Vellingiri, B.; Valsala Gopalakrishnan, A. The role of Interleukin-4 in COVID-19 associated male infertility—A hypothesis. J. Reprod. Immunol. 2020, 142, 103213. [Google Scholar] [CrossRef] [PubMed]
- Naz, R.K.; Kaplan, P. Increased levels of interleukin-6 in seminal plasma of infertile men. J. Androl. 1994, 15, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Camejo, M.I.; Segnini, A.; Proverbio, F. Interleukin-6 (IL-6) in seminal plasma of infertile men, and lipid peroxidation of their sperm. Arch. Androl. 2001, 47, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Koçak, I.; Yenisey, C.; Dündar, M.; Okyay, P.; Serter, M. Relationship between seminal plasma interleukin-6 and tumor necrosis factor alpha levels with semen parameters in fertile and infertile men. Urol. Res. 2002, 30, 263–267. [Google Scholar] [CrossRef]
- Eggert-Kruse, W.; Kiefer, I.; Beck, C.; Demirakca, T.; Strowitzki, T. Role for tumor necrosis factor alpha (TNF-alpha) and interleukin 1-beta (IL-1beta) determination in seminal plasma during infertility investigation. Fertil. Steril. 2007, 87, 810–823. [Google Scholar] [CrossRef]
- Attia, H.; Finocchi, F.; Orciani, M.; Mehdi, M.; Zidi Jrah, I.; Lazzarini, R.; Balercia, G.; Mattioli Belmonte, M. Pro-inflammatory cytokines and microRNAs in male infertility. Mol. Biol. Rep. 2021, 48, 5935–5942. [Google Scholar] [CrossRef]
- Vilanova, L.T.; Rauch, M.C.; Mansilla, A.; Zambrano, A.; Brito, M.; Werner, E.; Alfaro, V.; Cox, J.F.; Concha, I.I. Expression of granulocyte-macrophage colony stimulating factor (GM-CSF) in male germ cells: GM-CSF enhances sperm motility. Theriogenology 2003, 60, 1083–1095. [Google Scholar] [CrossRef]
- Tanhaye Kalate Sabz, F.; Amjadi, F.S.; Zandieh, Z.; Hosseini, E.; Aflatoonian, R.; Tabatabaei, M.; Mohammadian, M.; Ashrafi, M. GM-CSF (granulocyte-macrophage colony-stimulating factor) treatment improves sperm parameters in men with oligoasthenoteratospermia via PI3K/AKT pathway. Andrologia 2022, 54, 1618–1630. [Google Scholar] [CrossRef]
- Hosseini, E.; Afradiasbagharani, P.; Mohammadian, M.; Amjadi, F.; Tabatabaei, M.; Tanhaye Kalate Sabz, F.; Zandieh, Z. Granulocyte-Macrophage Colony-Stimulating Factor Cytokine Addition After the Freeze-Thawing Process Improves Human Sperm Motility and Vitality in Asthenoteratozoospermia Patients. Biopreserv. Biobank. 2024, 22, 38–45. [Google Scholar] [CrossRef]
- Tanhaye Kalate Sabz, F.; Hosseini, E.; Amjadi, F.S.; Mohammadian, M.; Zandieh, Z.; Mohammadian, F.; Kafaeinezhad, R.; Ashrafi, M. In vitro effect of granulocyte-macrophage colony-stimulating factor (GM-CSF) on the expression of genes related to sperm motility and energy metabolism and intracytoplasmic sperm injection outcomes in obstructive azoospermic patients. Mol. Biol. Rep. 2024, 51, 727. [Google Scholar] [CrossRef]
- Saini, A.; McPherson, N.O.; Nottle, M. Addition of GM-CSF during in vitro oocyte maturation improves embryo development and implantation and birth rate in mice. Reprod. Fertil. 2024, 5, e240020. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, D.J.; Macpherson, A.M.; Tremellen, K.P.; Mottershead, D.G.; Gilchrist, R.B.; Robertson, S.A. TGF-β mediates proinflammatory seminal fluid signaling in human cervical epithelial cells. J. Immunol. 2012, 189, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A.; Prins, J.R.; Sharkey, D.J.; Moldenhauer, L.M. Seminal fluid and the generation of regulatory T cells for embryo implantation. Am. J. Reprod. Immunol. 2013, 69, 315–330. [Google Scholar] [CrossRef]
- Chu, T.; Kawinski, E. Plasmin, substilisin-like endoproteases, tissue plasminogen activator, and urokinase plasminogen activator are involved in activation of latent TGF-β1 in human seminal plasma. Biochem. Biophys. Res. Commun. 1998, 253, 128–134. [Google Scholar] [CrossRef]
- Emami, N.; Diamandis, E.P. Potential role of multiple members of the kallikrein-related peptidase family of serine proteases in activating latent TGFβ1 in semen. Biol. Chem. 2010, 391, 85–95. [Google Scholar] [CrossRef]
- Jepsen, I.E.; Saxtorph, M.H.; Englund, A.L.M.; Petersen, K.B.; Wissing, M.L.M.; Hviid, T.V.F.; Macklon, N. Probiotic treatment with specific lactobacilli does not improve an unfavorable vaginal microbiota prior to fertility treatment-A randomized, double-blinded, placebo-controlled trial. Front. Endocrinol. 2022, 13, 1057022. [Google Scholar] [CrossRef]
- Thanaboonyawat, I.; Pothisan, S.; Petyim, S.; Laokirkkiat, P. Pregnancy outcomes after vaginal probiotic supplementation before frozen embryo transfer: A randomized controlled study. Sci. Rep. 2023, 13, 11892. [Google Scholar] [CrossRef]
- Naghi Jafarabadi, M.; Hadavi, F.; Ahmadi, M.; Masoumi, M.; Zabihzadeh, S. Intravaginal probiotics before embryo transfer do not improve pregnancy rates in recurrent implantation failure cases: An RCT. Int. J. Reprod. Biomed. 2024, 22, 363–374. [Google Scholar] [CrossRef]
- Di Pierro, F.; Sinatra, F.; Cester, M.; Da Ros, L.; Pistolato, M.; Da Parè, V.; Fabbro, L.; Maccari, D.; Dotto, S.; Sossai, S.; et al. Effect of L. crispatus M247 administration on pregnancy outcomes in women undergoing IVF: A controlled, retrospective, observational, and open-label study. Microorganisms 2023, 11, 2796. [Google Scholar] [CrossRef]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen–gut microbiome axis: Physiological and clinical implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef]
- Kwon, M.S.; Lee, H.K. Host and microbiome interplay shapes the vaginal microenvironment. Front. Immunol. 2022, 13, 919728. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zheng, L.W.; Ma, S.; Zhao, D.H.; Xu, Y. The gut microbiota: Emerging biomarkers and potential treatments for infertility-related diseases. Front. Cell. Infect. Microbiol. 2024, 14, 1450310. [Google Scholar] [CrossRef] [PubMed]
- Plottel, C.S.; Blaser, M.J. Microbiome and malignancy. Cell Host Microbe 2011, 10, 324–335. [Google Scholar] [CrossRef]
- Salliss, M.E.; Farland, L.V.; Mahnert, N.D.; Herbst-Kralovetz, M.M. The role of gut and genital microbiota and the estrobolome in endometriosis, infertility and chronic pelvic pain. Hum. Reprod. Update 2021, 28, 92–131. [Google Scholar] [CrossRef]
- Aragón, A.; Jurado, R.; Jara, J.; Rodríguez, J.M.; Orgaz, B. Investigating the metabolism of estrogens in Ligilactobacillus salivarius strains isolated from human milk and vaginal microbiota. Nutrients 2024, 16, 861. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic Characteristics | Placebo Group (N = 30) Median (Q1, Q3) | Probiotic Group (N = 27) Median (Q1, Q3) | p-Value 1 |
|---|---|---|---|
| Women | |||
| Age at recruitment (years) | 36.52 (35.17, 37.88) | 37.02 (34.91, 38.29) | 0.533 |
| Weight (kg) | 61.5 (56.0, 71.0) | 60.0 (56.0, 67.0) | 0.455 |
| Height (cm) | 162.5 (158.0, 168.0) | 163.0 (158.0, 167.0) | 0.867 |
| Male partners | |||
| Age at recruitment (years) | 36.04 (34.53, 37.96) | 36.91 (33.40, 39.67) | 0.835 |
| Weight (kg) | 80.0 (74.0, 90.0) | 77.0 (75.0, 85.0) | 0.437 |
| Height (cm) | 178.5 (175.0, 183.0) | 178.0 (173.0, 181.0) | 0.423 |
| Placebo Group (N = 30) n/N (%) | Probiotic Group (N = 27) n/N (%) | p-Value 1 | |
|---|---|---|---|
| Spontaneous pregnancies before ovarian stimulation | 3/30 (10.0) | 3/27 (11.1) | 1.000 |
| Ovarian puncture | |||
| No | 5/27 (19.2) | 2/24 (8.3) | 0.423 |
| Yes | 22/27 (80.8) | 22/24 (91.7) | |
| Embryo transfer | |||
| No | 8/22 (33.3) | 5/22 (22.7) | 0.322 * |
| Yes | 14/22 (66.7) | 17/22 (77.3) | |
| Frozen embryo | 10/14 (71.4) | 13/17 (76.5) | 1.000 |
| Fresh embryo | 4/14 (28.6) | 4/17 (23.5) | |
| Pregnancy after embryo transfer | |||
| No | 10/14 (71.4) | 6/17 (35.3) | 0.045 * |
| Yes | 4/14 (28.6) | 11/17 (64.7) | |
| Frozen embryo | 3/4 (75.0) | 9/11 (81.8) | 1.000 |
| Fresh embryo | 1/4 (25.0) | 2/11 (18.2) | |
| Total pregnancies 2 | 8/30 (26.7) | 14/27 (51.8) | 0.051 * |
| Abortions | 2/30 (6.7) | 1/27 (3.7) | 0.540 |
| Successful pregnancies | 6/30 (20.0) | 13/27 (48.1) | 0.024 * |
| Time 1 | p-Value 1 | Time 2 | p-Value | |||
|---|---|---|---|---|---|---|
| Placebo group | Probiotic group | Placebo group | Probiotic group | |||
| Prevalence, n/N (%) 2 | 2/29 (6.9) | 2/27 (7.4) | 1.000 | 2/26 (7.7) | 17/19 (89.5) # | <0.001 * |
| Concentration, median (Q1, Q3) | 2.15 (1.60, 2.70) | 2.10 (1.80, 2.40) | 0.699 | 2.25 (2.10, 2.40) | 4.60 (2.30, 5.90) | 0.352 |
| Not pregnancy | Pregnancy | Not pregnancy | Pregnancy | |||
| Prevalence, n/N (%) | 2/35 (5.7) | 2/21 (9.5) | 1.000 | 9/30 (30.0) # | 10/15 (66.7) # | 0.019 * |
| Concentration, median (Q1, Q3) | 2.15 (1.60, 2.70) | 2.10 (1.80, 2.40) | 0.699 | 2.10 (1.90, 2.30) | 5.80 (4.70, 6.40) # | <0.001 |
| Not pregnancy + miscarriage | Successful pregnancy | Not pregnancy + miscarriage | Successful pregnancy | |||
| Prevalence, n/N (%) | 2/38 (5.3) | 2/18 (11.1) | 0.589 | 10/35 (30.0) # | 9/12 (66.7) # | 0.007 |
| Concentration, median (Q1, Q3) | 2.15 (1.60, 2.70) | 2.10 (1.80, 2.40) | 0.699 | 2.20 (1.90, 2.40) | 5.90 (4.90, 6.40) # | <0.001 |
| Time 1 | p-Value 1 | Time 2 | p-Value | |||
|---|---|---|---|---|---|---|
| Placebo group (n = 29) | Probiotic group (n = 27) | Placebo (n = 26) | Probiotic (n = 19) | |||
| TGFβ1 2 | 1.29 (1.11, 1.44) | 1.42 (1.25, 1.72) | 0.014 | 1.25 (1.17, 1.62) | 2.33 (1.27, 2.97) # | 0.002 |
| VEGF | 119.0 (98.0, 154.0) | 133.0 (98.0, 150.0) | 0.566 | 110.0 (97.0, 140.0) | 349.0 (101.0, 529.0) | 0.019 |
| Not pregnancy (n = 35) | Pregnancy (n = 21) | Not pregnancy (n = 30) | Pregnancy (n = 15) | |||
| TGFβ1 | 1.25 (1.06, 1.37) | 1.67 (1.49, 1.83) | <0.001 | 1.23 (1.17, 1.42) | 2.69 (2.27, 3.01) # | <0.001 |
| VEGF | 102.0 (85.0, 122.0) | 162.0 (145.0, 217.0) | <0.001 | 107.5 (92.0, 123.0) | 431.0 (303.0, 564.0) # | <0.001 |
| Not pregnancy + miscarriage (n = 38) | Successful pregnancy (n = 18) | Not pregnancy + miscarriage (n = 33) | Successful pregnancy (n = 12) | |||
| TGFβ1 | 1.27 (1.07, 1.38) | 1.72 (1.57, 1.86) | <0.001 | 1.27 (1.19, 1.45) | 2.86 (2.38, 3.04) # | <0.001 |
| VEGF | 104.0 (87.0, 130.0) | 160.0 (145.0, 231.0) | <0.001 | 109.0 (94.0, 129.0) | 485.5 (347.0, 582.5) # | <0.001 |
| Compound 2 | Time 1 | p-Value 1 | Time 2 | p-Value | ||
|---|---|---|---|---|---|---|
| Placebo group (n = 29) | Probiotic group (n = 27) | Placebo group (n = 25) | Probiotic group (n = 19) | |||
| IL-2 | 3.56 (3.41, 3.82) | 3.59 (3.31, 3.87) | 0.646 | 3.67 (3.45, 3.99) | 3.47 (3.23, 3.71) | 0.090 |
| IL-4 | 0.58 (0.53, 0.63) | 0.65 (0.60, 0.69) | <0.001 | 0.61 (0.57, 0.66) | 0.53 (0.43, 0.61) # | 0.002 |
| IL-6 | 3.27 (2.74, 3.71) | 3.62 (3.23, 3.82) | 0.103 | 3.41 (3.12, 3.76) | 1.88 (1.43, 2.72) # | <0.001 |
| IL-8 | 197.5 (141.2, 298.5) | 247.7 (201.9, 325.5) | 0.156 | 201.8 (164.6, 277.8) | 200.1 (177.4, 311.6) | 0.434 |
| IL-10 | 2.45 (2.12, 2.90) | 2.34 (1.93, 3.06) | 0.731 | 2.45 (2.14, 2.92) | 2.48 (2.03, 2.84) | 0.653 |
| GM-CSF | 1.29 (1.17, 1.42) | 1.31 (1.17, 1.45) | 0.889 | 1.26 (1.14, 1.49) | 1.34 (1.23, 1.47) | 0.192 |
| IFNγ | 50.94 (40.11, 64.60) | 52.03 (44.94, 62.08) | 0.664 | 48.81 (40.23, 69.66) | 27.17 (21.29, 33.07) # | <0.001 |
| TNFα | 6.64 (4.94, 7.03) | 7.14 (6.50, 7.87) | 0.022 | 6.17 (5.18, 7.23) | 4.04 (3.87, 5.02) # | <0.001 |
| TGFβ1 | 348.2 (245.3, 398.6) | 257.3 (145.8, 361.3) | 0.091 | 345.5 (234.7, 388.1) | 491.8 (431.8, 573.0) # | <0.001 |
| Not pregnancy (n = 35) | Pregnancy (n = 21) | Not pregnancy (n = 30) | Pregnancy (n = 14) | |||
| IL-2 | 3.59 (3.29, 4.02) | 3.53 (3.41, 3.78) | 0.987 | 3.65 (3.33, 3.96) | 3.62 (3.44, 3.71) | 0.830 |
| IL-4 | 0.62 (0.58, 0.67) | 0.59 (0.56, 0.67) | 0.351 | 0.61 (0.57, 0.65) | 0.45 (0.43, 0.48) # | <0.001 |
| IL-6 | 3.59 (3.23, 3.87) | 3.33 (1.99, 3.69) | 0.043 | 3.29 (3.02, 3.71) | 1.53 (1.41, 1.95) # | <0.001 |
| IL-8 | 237.1 (178.4, 325.5) | 207.1 (134.8, 265.7) | 0.126 | 228.9 (180.2, 301.4) | 183.7 (142.6, 200.1) | 0.031 |
| IL-10 | 2.54 (2.01, 3.07) | 2.32 (1.91, 2.74) | 0.150 | 2.59 (2.10, 2.92) | 2.48 (2.06, 2.65) | 0.512 |
| GM-CSF | 1.31 (1.17, 1.46) | 1.28 (1.15, 1.38) | 0.630 | 1.27 (1.14, 1.42) | 1.36 (1.23, 1.59) | 0.107 |
| IFNγ | 58.10 (41.40, 69.69) | 47.48 (44.21, 54.29) | 0.148 | 48.67 (35.12, 67.02) | 23.47 (21.19, 29.12) # | <0.001 |
| TNFα | 6.91 (5.73, 7.93) | 6.71 (4.97, 7.26) | 0.356 | 6.03 (5.02, 7.23) | 4.02 (3.87, 4.82) # | <0.001 |
| TGFβ1 | 347.3 (164.2, 402.1) | 257.3 (240.1, 333.9) | 0.498 | 374.4 (245.7, 451.8) | 489.9 (415.2, 579.3) # | 0.004 |
| Not pregnancy + miscarriage (n = 38) | Successful pregnancy (n = 18) | Not pregnancy + miscarriage (n = 33) | Successful pregnancy (n = 11) | |||
| IL-2 | 3.58 (3.31, 3.87) | 3.56 (3.38, 3.78) | 0.958 | 3.65 (3.33, 3.94) | 3.62 (3.44, 3.71) | 0.968 |
| IL-4 | 0.62 (0.58, 0.67) | 0.59 (0.49, 0.67) | 0.312 | 0.61 (0.57, 0.64) | 0.44 (0.41, 0.46) # | <0.001 |
| IL-6 | 3.47 (3.21, 3.87) | 3.42 (1.99, 3.71) | 0.188 | 3.24 (2.98, 3.58) | 1.46 (1.27, 1.64) # | <0.001 |
| IL-8 | 232.6 (178.3, 310.2) | 223.4 (134.8, 278.4) | 0.366 | 221.8 (178.4, 297.1) | 199.0 (134.5, 261.5) | 0.175 |
| IL-10 | 2.46 (2.01, 3.06) | 2.26 (1.91, 2.79) | 0.258 | 2.45 (2.10, 2.92) | 2.48 (2.06, 2.69) | 0.892 |
| GM-CSF | 1.32 (1.18, 1.48) | 1.28 (1.09, 1.35) | 0.247 | 1.27 (1.15, 1.45) | 1.34 (1.23, 1.59) | 0.350 |
| IFNγ | 55.67 (40.11, 69.69) | 48.27 (45.42, 54.29) | 0.277 | 48.52 (35.17, 67.02) | 22.38 (20.29, 29.12) # | <0.001 |
| TNFα | 6.91 (5.67, 7.93) | 6.80 (5.29, 7.26) | 0.516 | 5.92 (4.98, 7.14) | 3.94 (3.52, 4.04) # | <0.001 |
| TGFβ1 | 347.7 (174.5, 402.1) | 274.2 (229.2, 333.2) | 0.273 | 382.4 (299.7, 444.3) | 503.1 (486.5, 594.3) # | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huerga López, C.; Sánchez Martín, M.J.; Herráez Moreta, A.; Calvo Urrutia, M.; Cristóbal García, I.; Díaz Morillo, C.; Blanco-Rojo, R.; Sáez, M.E.; Olivares, M.; Arroyo, R.; et al. Ligilactobacillus salivarius CECT5713 Increases Term Pregnancies in Women with Infertility of Unknown Origin: A Randomized, Triple-Blind, Placebo-Controlled Trial. Nutrients 2025, 17, 1860. https://doi.org/10.3390/nu17111860
Huerga López C, Sánchez Martín MJ, Herráez Moreta A, Calvo Urrutia M, Cristóbal García I, Díaz Morillo C, Blanco-Rojo R, Sáez ME, Olivares M, Arroyo R, et al. Ligilactobacillus salivarius CECT5713 Increases Term Pregnancies in Women with Infertility of Unknown Origin: A Randomized, Triple-Blind, Placebo-Controlled Trial. Nutrients. 2025; 17(11):1860. https://doi.org/10.3390/nu17111860
Chicago/Turabian StyleHuerga López, Cristina, María J. Sánchez Martín, Aránzazu Herráez Moreta, Marta Calvo Urrutia, Ignacio Cristóbal García, Cristina Díaz Morillo, Ruth Blanco-Rojo, María E. Sáez, Mónica Olivares, Rebeca Arroyo, and et al. 2025. "Ligilactobacillus salivarius CECT5713 Increases Term Pregnancies in Women with Infertility of Unknown Origin: A Randomized, Triple-Blind, Placebo-Controlled Trial" Nutrients 17, no. 11: 1860. https://doi.org/10.3390/nu17111860
APA StyleHuerga López, C., Sánchez Martín, M. J., Herráez Moreta, A., Calvo Urrutia, M., Cristóbal García, I., Díaz Morillo, C., Blanco-Rojo, R., Sáez, M. E., Olivares, M., Arroyo, R., Herranz, C., Alba, C., Rodríguez, J. M., & Fernández, L. (2025). Ligilactobacillus salivarius CECT5713 Increases Term Pregnancies in Women with Infertility of Unknown Origin: A Randomized, Triple-Blind, Placebo-Controlled Trial. Nutrients, 17(11), 1860. https://doi.org/10.3390/nu17111860