

Structural Characterization and Efficacy in Alleviating Lung Inflammation of Sialylated Glycopeptides from Edible Bird’s Nest

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Experimental Animals
2.3. Preparation of Sialylated Glycopeptides (SCP) from EBN
2.4. Structural Characterization of SCP
2.4.1. Neutral Sugar and SA Content Quantification
2.4.2. Amino Acid Composition Analysis
2.4.3. Molecular Weight Distribution Determination
2.4.4. FTIR Spectral Analysis
2.5. Animal Grouping and Biological Sample Collection
2.6. Lung Inflammatory Markers
2.7. Lung Histopathology
2.8. Lung Metabolomics
2.9. Statistical Analysis
3. Results
3.1. Chemical Composition Analysis of SCP
3.2. Primary Structure Analysis of SCP
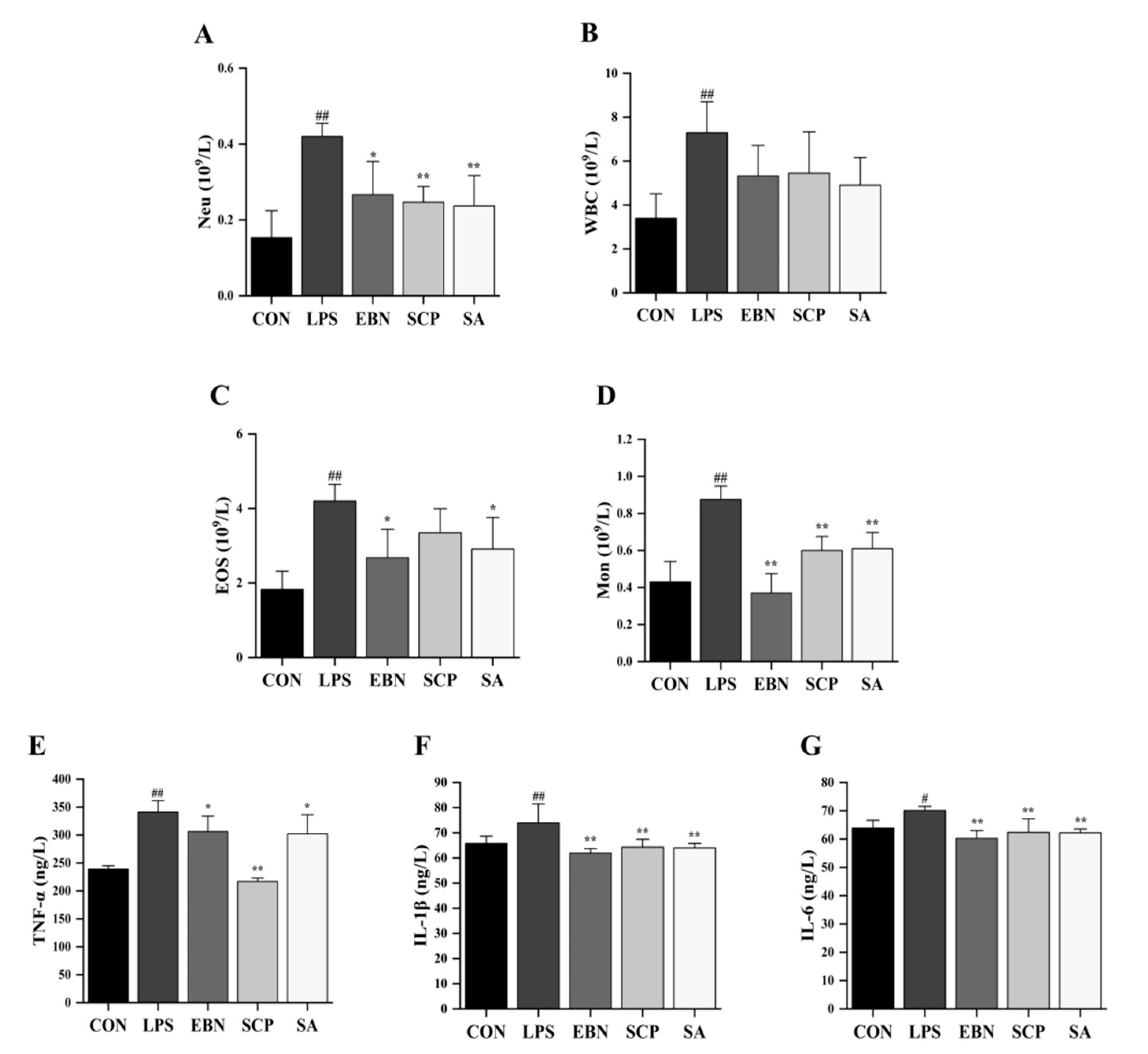
3.3. SCP Alleviated the Changes of LPS-Induced Lung Inflammation Levels
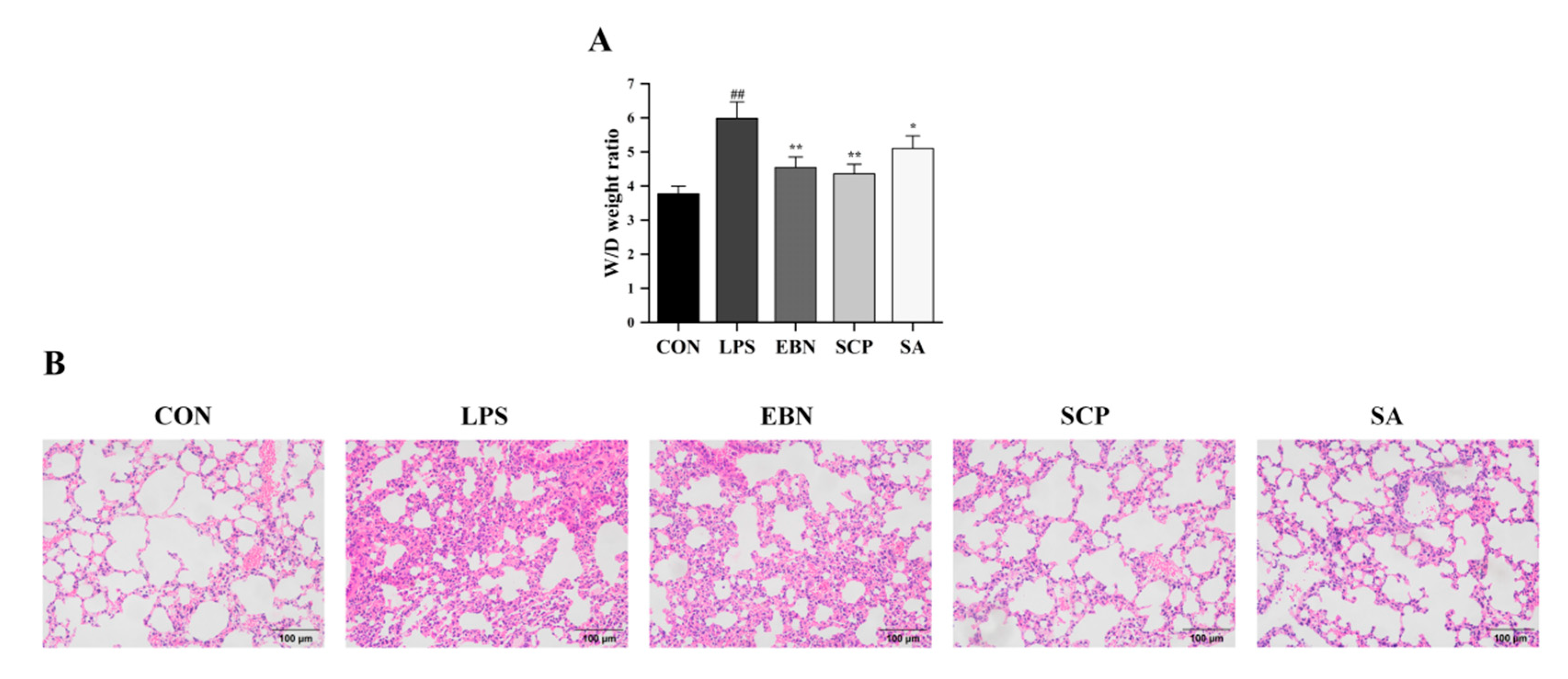
3.4. SCP Alleviated the Changes of LPS-Induced Lung Tissue Pathological Structures
3.5. SCP Alleviated the Change of LPS-Induced Lung Tissue Metabolic Levels
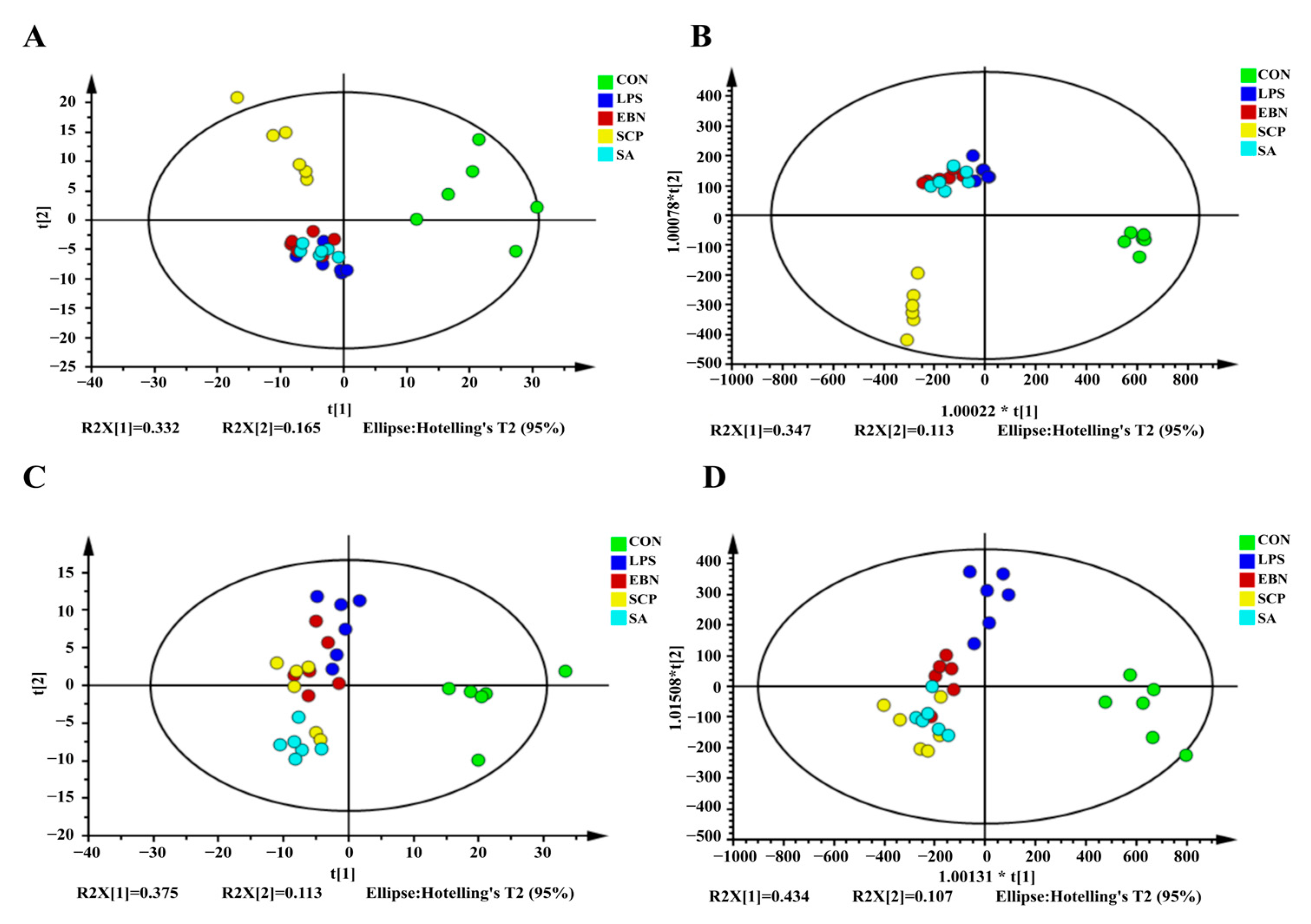
3.5.1. Analysis of the Metabolic Profiles
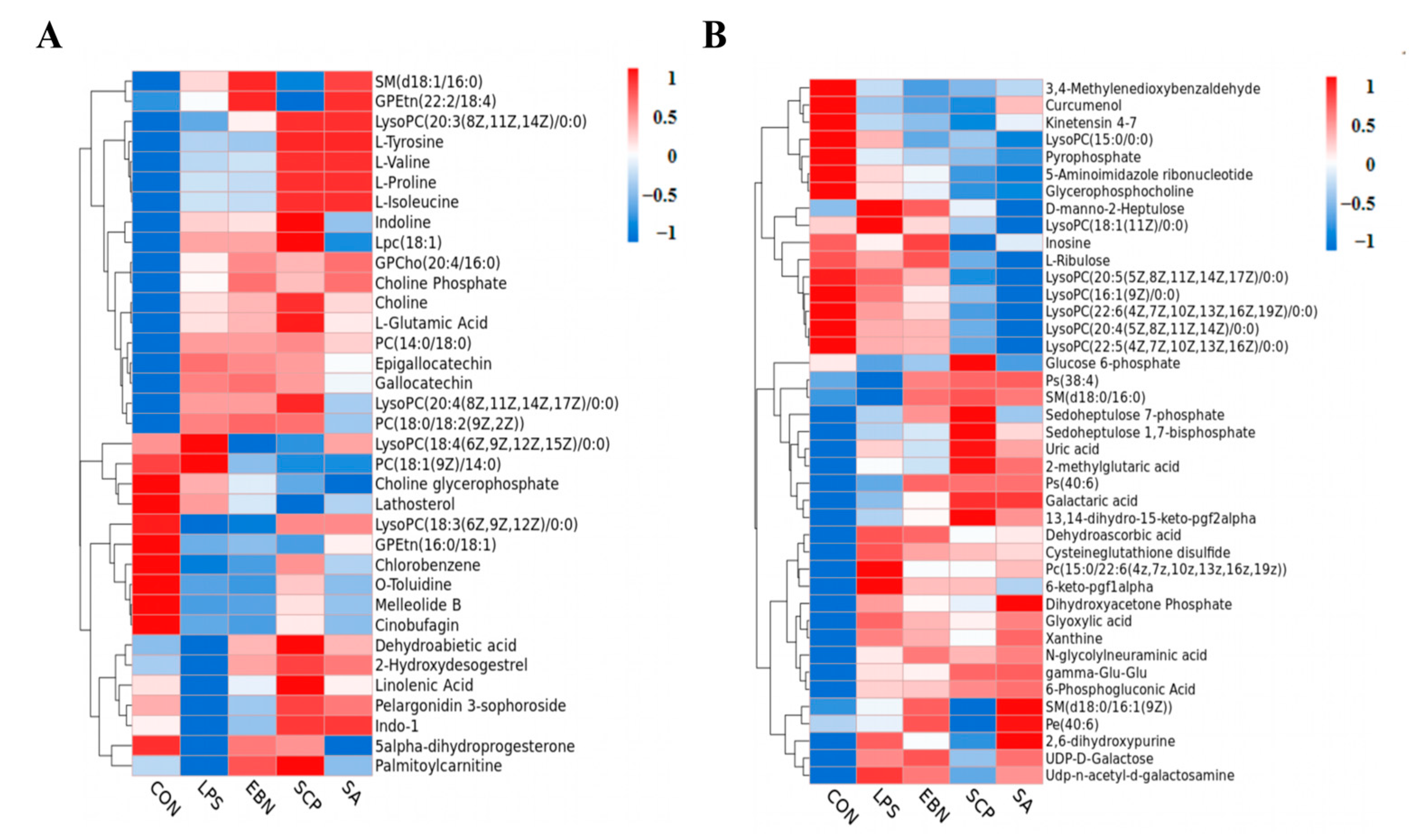
3.5.2. Screening of Differential Metabolites
3.5.3. Metabolic Pathway Analysis of Differential Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, M.; Qing, Y.; Fan, Q.; Li, S.; Yan, T.; Amakye, W.K.; Xu, Y.; Liu, X.; Ren, J. Identification of the Wound Healing Activity Peptidome of Edible Bird’s Nest Protein Hydrolysate and the In Silico Evaluation of Its Transport and Absorption Potential in Skin. J. Agric. Food Chem. 2024, 72, 12529–12540. [Google Scholar] [CrossRef]
- Dai, Y.; Cao, J.; Wang, Y.; Chen, Y.; Jiang, L. A comprehensive review of edible bird’s nest. Food Res. Int. 2021, 140, 109875. [Google Scholar] [CrossRef]
- Ling, J.W.A.; Chang, L.S.; Babji, A.S.; Latip, J.; Koketsu, M.; Lim, S.J. Review of sialic acid’s biochemistry, sources, extraction and functions with special reference to edible bird’s nest. Food Chem. 2022, 367, 130755. [Google Scholar] [CrossRef]
- Quek, M.C.; Chin, N.L.; Yusof, Y.A.; Law, C.L.; Tan, S.W. Characterization of edible bird’s nest of different production, species and geographical origins using nutritional composition, physicochemical properties and antioxidant activities. Food Res. Int. 2018, 109, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Careena, S.; Sani, D.; Tan, S.N.; Lim, C.W.; Hassan, S.; Norhafizah, M.; Kirby, B.P.; Ideris, A.; Stanslas, J.; Bin Basri, H.; et al. Effect of Edible Bird’s Nest Extract on Lipopolysaccharide-Induced Impairment of Learning and Memory in Wistar Rats. Evid.-Based Complement. Altern. Med. 2018, 2018, 9318789. [Google Scholar] [CrossRef] [PubMed]
- Rashed, A.A.; Ahmad, H.; Abdul Khalid, S.K.; Rathi, D.-N.G. The Potential Use of Sialic Acid from Edible Bird’s Nest to Attenuate Mitochondria Dysfunction by In Vitro Study. Front. Pharmacol. 2021, 12, 633303. [Google Scholar] [CrossRef]
- Ling, J.W.A.; Chang, L.S.; Babji, A.S.; Lim, S.J. Recovery of value-added glycopeptides from edible bird’s nest (EBN) co-products: Enzymatic hydrolysis, physicochemical characteristics and bioactivity. J. Sci. Food Agric. 2020, 100, 4714–4722. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Ma, J.H.; Tang, S.; Yu, Z.; Liu, S. Lycium barbarum Glycopeptide alleviated retinal inflammation via NF-κB/MAPK pathways in microglial cells. Investig. Ophthalmol. Vis. Sci. 2024, 65, 205. [Google Scholar]
- He, H.; Wang, J.; Gong, P.; Xiao, Y.; Li, S.; Wang, J.; Geng, F. Structural identification and immunomodulatory effects of chicken egg white glycopeptides. LWT 2024, 200, 116195. [Google Scholar] [CrossRef]
- Zeng, H.; Jian, Y.; Xie, Y.; Fan, Q.; Chang, Q.; Zheng, B.; Zhang, Y. Edible bird’s nest inhibits the inflammation and regulates the immunological balance of lung injury mice by SO2. Food Front. 2022, 3, 773–784. [Google Scholar] [CrossRef]
- Ma, F.; Liu, D. Sketch of the edible bird’s nest and its important bioactivities. Food Res. Int. 2012, 48, 559–567. [Google Scholar] [CrossRef]
- Cheng, X.R.; Zhang, C.X.; Zhao, X.Y.; Li, Q.S.; Song, Y.; Zhang, L.; Ma, J.H.; Chen, Y.Y.; Yuan, M.; Wang, D.L. Prophylactic effects of sialylated glycopeptides from edible bird’s nest on neuroinflammation in lipopolysaccharide-treated mice via the gut-brain axis. Food Sci. Hum. Wellness 2024. [Google Scholar] [CrossRef]
- Ahmad, S.; Zhang, X.L.; Ahmad, A. Epigenetic regulation of pulmonary inflammation. Semin. Cell Dev. Biol. 2024, 154, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Pervizaj-Oruqaj, L.; Ferrero, M.R.; Matt, U.; Herold, S. The guardians of pulmonary harmony: Alveolar macrophages orchestrating the symphony of lung inflammation and tissue homeostasis. Eur. Respir. Rev. 2024, 33, 230263. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhu, J.; Zhu, B.; Yin, H.; Wang, D.; Deng, Y.; Zhang, M.; Zhang, C. Recent advances in the structure and immunomodulatory activity of food-derived glycoprotein complex. J. Future Foods 2025, 5, 542–550. [Google Scholar] [CrossRef]
- Xie, Y.K.; Pan, X.Y.; Liang, X.R.; Zhai, K.F.; Yu, Q. Research progress on structural characterization and bioactivities of Poria cocos and Ganoderma polysaccharides. Food Med. Homol. 2025, 2, 9420040. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, W.; Wang, X.; Yu, L.; Wei, Y.; Wei, L.; Xie, X.; Li, X. Ganoderma lucidum polysaccharide inhibits LPS-induced inflammatory injury to mammary epithelial cells. J. Future Foods 2023, 3, 49–54. [Google Scholar] [CrossRef]
- Ye, J.; Guan, M.; Lu, Y.; Zhang, D.; Li, C.; Zhou, C. Arbutin attenuates LPS-induced lung injury via Sirt1/Nrf2/NF-κBp65 pathway. Pulm. Pharmacol. Ther. 2019, 54, 53–59. [Google Scholar] [CrossRef]
- Gao, Z.; Liu, X.; Wang, W.; Yang, Q.; Dong, Y.; Xu, N.; Zhang, C.; Song, X.; Ren, Z.; Zhao, F.; et al. Characteristic anti-inflammatory and antioxidative effects of enzymatic- and acidic- hydrolysed mycelium polysaccharides by Oudemansiella radicata on LPS-induced lung injury. Carbohydr. Polym. 2019, 204, 142–151. [Google Scholar] [CrossRef]
- Wang, S.; Tan, S.; Beng, H.; Liu, F.; Huang, J.; Kuai, Y.; Zhang, R.; Tan, W. Protective effect of isosteviol sodium against LPS-induced multiple organ injury by regulating of glycerophospholipid metabolism and reducing macrophage-driven inflammation. Pharmacol. Res. 2021, 172, 105781. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, K.; Zhang, J.; Xu, G.; Guo, Z.; Lu, X.; Liang, C.; Gu, X.; Huang, L.; Liu, S.; et al. Cordyceps militaris solid medium extract alleviates lipopolysaccharide-induced acute lung injury via regulating gut microbiota and metabolism. Front. Immunol. 2025, 15, 1528222. [Google Scholar] [CrossRef] [PubMed]
- Hui Yan, T.; Mun, S.L.; Lee, J.L.; Lim, S.J.; Daud, N.A.; Babji, A.S.; Sarbini, S.R. Bioactive sialylated-mucin (SiaMuc) glycopeptide produced from enzymatic hydrolysis of edible swiftlet’s nest (ESN): Degree of hydrolysis, nutritional bioavailability, and physicochemical characteristics. Int. J. Food Prop. 2022, 25, 252–277. [Google Scholar] [CrossRef]
- Doerner, K.C.; White, B.A. Detection of glycoproteins separated by nondenaturing polyacrylamide gel electrophoresis using the periodic acid-Schiff stain. Anal. Biochem. 1990, 187, 147–150. [Google Scholar] [CrossRef]
- Martins, M.S.; Nascimento, M.H.; Barbosa, L.L.; Campos, L.C.G.; Singh, M.N.; Martin, F.L.; Romão, W.; Filgueiras, P.R.; Barauna, V.G. Detection and quantification using ATR-FTIR spectroscopy of whey protein concentrate adulteration with wheat flour. LWT 2022, 172, 114161. [Google Scholar] [CrossRef]
- Chen, X.; Yang, X.; Liu, T.; Guan, M.; Feng, X.; Dong, W.; Chu, X.; Liu, J.; Tian, X.; Ci, X.; et al. Kaempferol regulates MAPKs and NF-κB signaling pathways to attenuate LPS-induced acute lung injury in mice. Int. Immunopharmacol. 2012, 14, 209–216. [Google Scholar] [CrossRef]
- Cheng, X.R.; Guan, L.J.; Muskat, M.N.; Cao, C.C.; Guan, B. Effects of Ejiao peptide–iron chelates on intestinal inflammation and gut microbiota in iron deficiency anemic mice. Food Funct. 2021, 12, 10887–10902. [Google Scholar] [CrossRef]
- Hong, T.; Yin, J.Y.; Nie, S.P.; Xie, M.Y. Applications of infrared spectroscopy in polysaccharide structural analysis: Progress, challenge and perspective. Food Chem: X 2021, 12, 100168. [Google Scholar] [CrossRef] [PubMed]
- Mistry, B. A Handbook of Spectroscopic Data CHEMISTRY (UV, JR, PMR, JJCNMR and Mass Spectroscopy); Oxford Book Company Jaipur: Jaipur, India, 2009; pp. 39–42. [Google Scholar]
- Mohamad Ibrahim, R.; Mohamad Nasir, N.N.; Abu Bakar, M.Z.; Mahmud, R.; Ab Razak, N.A. The Authentication and Grading of Edible Bird’s Nest by Metabolite, Nutritional, and Mineral Profiling. Foods 2021, 10, 1574. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Zhao, J.; Gu, S.; Chen, S.; Jia, W.; Huang, S.; Wu, J. Potentilla anserina L. polysaccharide ameliorates LPS-induced acute lung injury and relevant intestinal mucosal barrier impairment. Int. J. Biol. Macromol. 2025, 305, 140667. [Google Scholar] [CrossRef]
- Zhang, S.; Lai, X.; Liu, X.; Li, Y.; Li, B.; Huang, X.; Zhang, Q.; Chen, W.; Lin, L.; Yang, G. Competitive Enzyme-Linked Immunoassay for Sialoglycoprotein of Edible Bird’s Nest in Food and Cosmetics. J. Agric. Food Chem. 2012, 60, 3580–3585. [Google Scholar] [CrossRef]
- Sriwilaijaroen, N.; Hanamatsu, H.; Yokota, I.; Nishikaze, T.; Ijichi, T.; Takahashi, T.; Sakoda, Y.; Furukawa, J.-I.; Suzuki, Y. Edible bird’s nest: N- and O-glycan analysis and synergistic anti-avian influenza virus activity with neuraminidase inhibitors. Antivir. Res. 2024, 232, 106040. [Google Scholar] [CrossRef] [PubMed]
- Bourgoin-Voillard, S.; Goron, A.; Seve, M.; Moinard, C. Regulation of the proteome by amino acids. Proteomics 2016, 16, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Shim, E.K.S.; Chandra, G.F.; Lee, S.-Y. Thermal analysis methods for the rapid identification and authentication of swiftlet (Aerodramus fuciphagus) edible bird’s nest—A mucin glycoprotein. Food Res. Int. 2017, 95, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shi, Y. Investigation the interaction between quinoa glycopeptides and polyphenol: Spectroscopic analyses and antioxidant activities. J. Food Meas. Charact. 2023, 17, 5594–5605. [Google Scholar] [CrossRef]
- Sun, N.; Cui, P.; Lin, S.; Yu, C.; Tang, Y.; Wei, Y.; Xiong, Y.; Wu, H. Characterization of sea cucumber (Stichopus japonicus) ovum hydrolysates: Calcium chelation, solubility and absorption into intestinal epithelial cells. J. Sci. Food Agric. 2017, 97, 4604–4611. [Google Scholar] [CrossRef]
- Medzhitov, R. The spectrum of inflammatory responses. Science 2021, 374, 1070–1075. [Google Scholar] [CrossRef]
- Ning, E.-J.; Sun, C.-W.; Wang, X.-F.; Chen, L.; Li, F.-F.; Zhang, L.-X.; Wang, L.-P.; Ma, Y.-N.; Zhu, J.; Li, X.; et al. Artemisia argyi polysaccharide alleviates intestinal inflammation and intestinal flora dysbiosis in lipopolysaccharide-treated mice. Food Med. Homol. 2024, 1, 9420008. [Google Scholar] [CrossRef]
- Bezerra, F.S.; Lanzetti, M.; Nesi, R.T.; Nagato, A.C.; Silva, C.P.E.; Kennedy-Feitosa, E.; Melo, A.C.; Cattani-Cavalieri, I.; Porto, L.C.; Valenca, S.S. Oxidative Stress and Inflammation in Acute and Chronic Lung Injuries. Antioxidants 2023, 12, 548. [Google Scholar] [CrossRef]
- Qian, N.; Zhang, C.-X.; Fang, G.-D.; Qiu, S.; Song, Y.; Yuan, M.; Wang, D.-L.; Cheng, X.-R. Interventional Effects of Edible Bird’s Nest and Free Sialic Acids on LPS-Induced Brain Inflammation in Mice. Nutrients 2025, 17, 531. [Google Scholar] [CrossRef]
- Pang, H.; Hu, Z. Metabolomics in drug research and development: The recent advances in technologies and applications. Acta Pharm. Sin. B 2023, 13, 3238–3251. [Google Scholar] [CrossRef]
- Sarkar, C.; Lipinski, M.M. Glycerophospholipid dysregulation after traumatic brain injury. Neurochem. Int. 2024, 175, 105701. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, S.; Bong How, S.; Gummer, J.; Trengove, R.; Moodley, Y. Metabolomics in chronic lung diseases. Respirology 2020, 25, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Guo, C.; Huang, S.; Sun, B. Inhaled nitric oxide alleviates hyperoxia suppressed phosphatidylcholine synthesis in endotoxin-induced injury in mature rat lungs. Respir. Res. 2006, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, Y.; Liu, L.; Wang, Q.; Zeng, J.; Chen, C. PM2.5 exposure perturbs lung microbiome and its metabolic profile in mice. Sci. Total Environ. 2020, 721, 137432. [Google Scholar] [CrossRef]
- Hassoun, P.M.; Yu, F.-S.; Cote, C.G.; Zulueta, J.J.; Sawhney, R.; Skinner, K.A.; Skinner, H.B.; Parks, D.A.; Lanzillo, J.J. Upregulation of Xanthine Oxidase by Lipopolysaccharide, Interleukin-1, and Hypoxia. Am. J. Respir. Crit. Care Med. 1998, 158, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Qi, R.; Li, T.; Zhang, X.; Shi, Y.; Xu, M.; Zhu, T. Kidney Organoids as a Novel Platform to Evaluate Lipopolysaccharide-Induced Oxidative Stress and Apoptosis in Acute Kidney Injury. Front. Med. 2021, 8, 766073. [Google Scholar] [CrossRef]
- Yuan, Q.; Xie, F.; Huang, W.; Hu, M.; Yan, Q.; Chen, Z.; Zheng, Y.; Liu, L. The review of alpha-linolenic acid: Sources, metabolism, and pharmacology. Phytother. Res. 2022, 36, 164–188. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, B.; Zhang, X.; Chen, X.; Zhu, J.; Zou, Y.; Li, J. Alpha-linolenic acid protects against lipopolysaccharide-induced acute lung injury through anti-inflammatory and anti-oxidative pathways. Microb. Pathog. 2020, 142, 104077. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids | Percentage Composition (%) | Amino Acids | Percentage Composition (%) |
|---|---|---|---|
| Asp | 9.97 ± 0.04 | Glu | 8.25 ± 0.04 |
| Cys | 0.90 ± 0.02 | Val * | 8.03 ± 0.03 |
| Ser | 7.05 ± 0.02 | Met * | 1.07 ± 0.01 |
| His * | 3.56 ± 0.06 | Phe * | 6.89 ± 0.02 |
| Gly | 4.40 ± 0.00 | Ile * | 3.40 ± 0.01 |
| Thr * | 6.32 ± 0.02 | Leu * | 7.62 ± 0.01 |
| Arg | 7.14 ± 0.01 | Lys * | 4.13 ± 0.02 |
| Ala | 3.44 ± 0.01 | Pro | 10.39 ± 0.06 |
| Tyr | 6.23 ± 0.03 | Trp * | 1.22 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Zhang, C.; Fang, G.; Qiu, S.; Yuan, M.; Qian, N.; Wang, D.; Cheng, X. Structural Characterization and Efficacy in Alleviating Lung Inflammation of Sialylated Glycopeptides from Edible Bird’s Nest. Nutrients 2025, 17, 1745. https://doi.org/10.3390/nu17101745
Li Q, Zhang C, Fang G, Qiu S, Yuan M, Qian N, Wang D, Cheng X. Structural Characterization and Efficacy in Alleviating Lung Inflammation of Sialylated Glycopeptides from Edible Bird’s Nest. Nutrients. 2025; 17(10):1745. https://doi.org/10.3390/nu17101745
Chicago/Turabian StyleLi, Qiushi, Chenxi Zhang, Guandong Fang, Shuang Qiu, Man Yuan, Nan Qian, Dongliang Wang, and Xiangrong Cheng. 2025. "Structural Characterization and Efficacy in Alleviating Lung Inflammation of Sialylated Glycopeptides from Edible Bird’s Nest" Nutrients 17, no. 10: 1745. https://doi.org/10.3390/nu17101745
APA StyleLi, Q., Zhang, C., Fang, G., Qiu, S., Yuan, M., Qian, N., Wang, D., & Cheng, X. (2025). Structural Characterization and Efficacy in Alleviating Lung Inflammation of Sialylated Glycopeptides from Edible Bird’s Nest. Nutrients, 17(10), 1745. https://doi.org/10.3390/nu17101745