
3,3′-Diindolylmethane Ameliorates Metabolism Dysfunction-Associated Fatty Liver Disease via AhR/p38 MAPK Signaling

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Drugs and Dosages
2.2. Animals
2.3. Biochemical Analysis
2.4. Histopathological Analysis
2.5. Cell Culture and Cell Treatment
2.6. CCK-8 Assay
2.7. Oil Red O Staining
2.8. Flow Cytometry
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
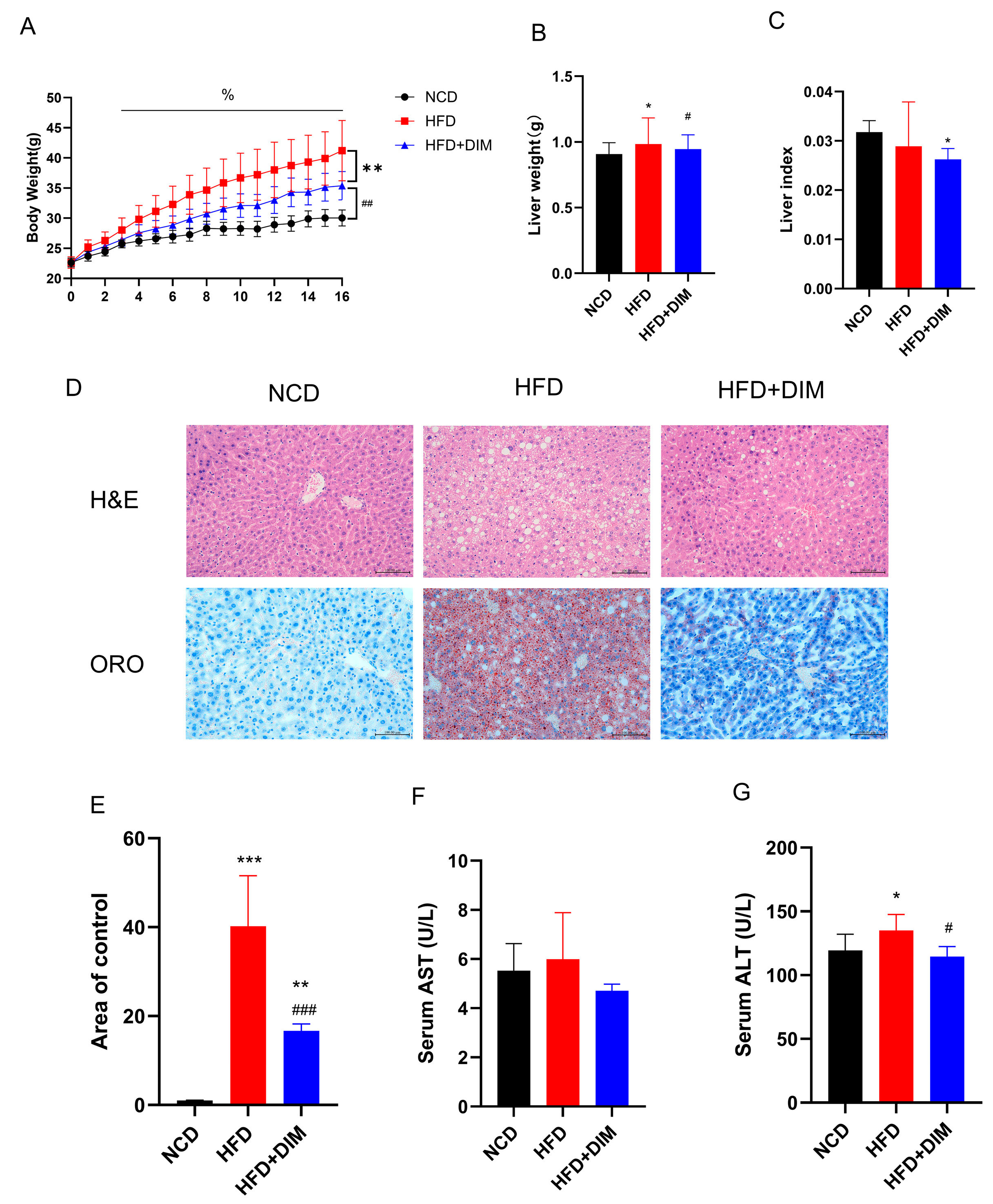
3.1. DIM Ameliorated Obesity and Liver Damage Induced by HFD
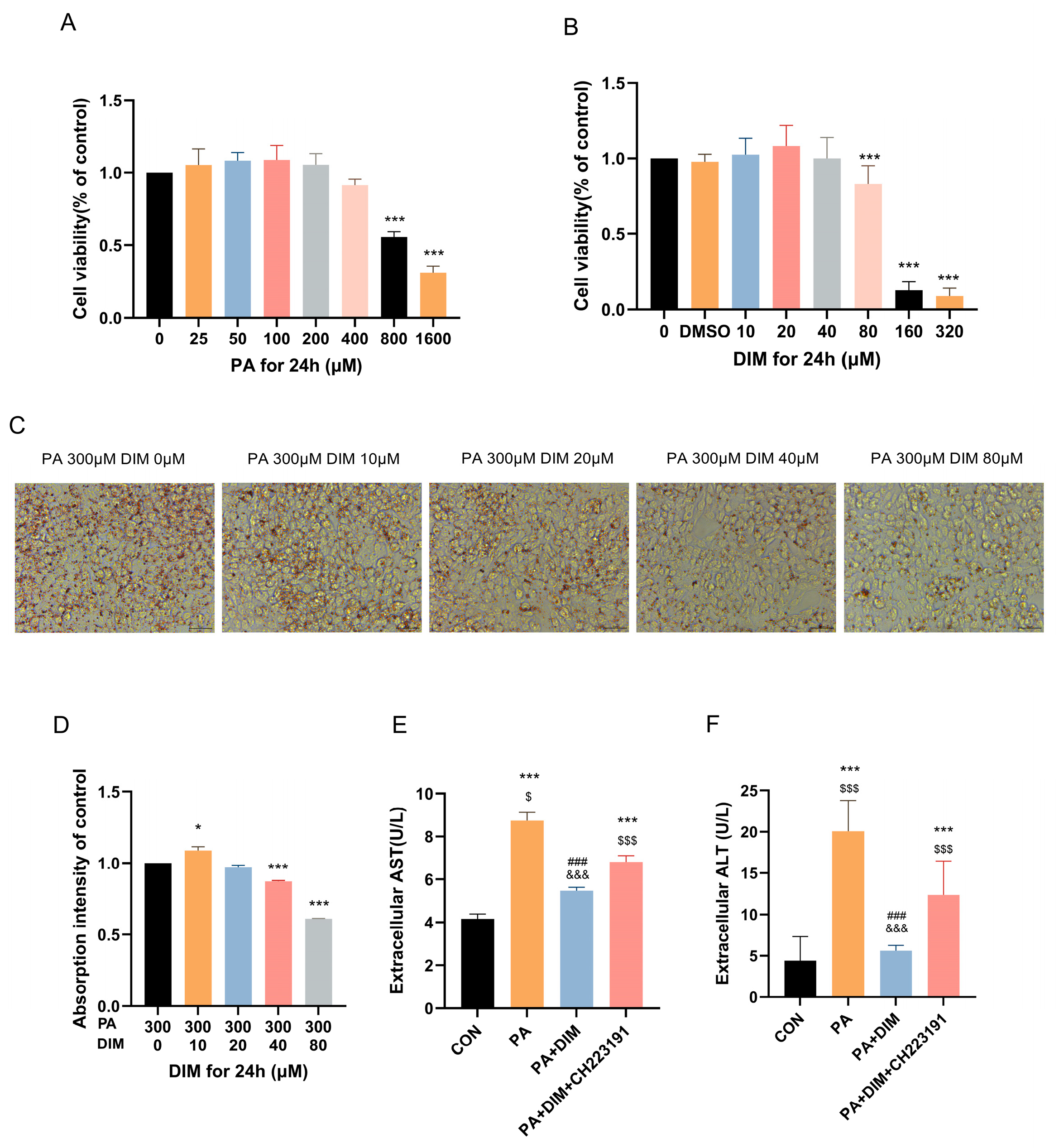
3.2. HepG2 Cells Accumulated Lipids as a Result of PA Activation, Which DIM Decreased
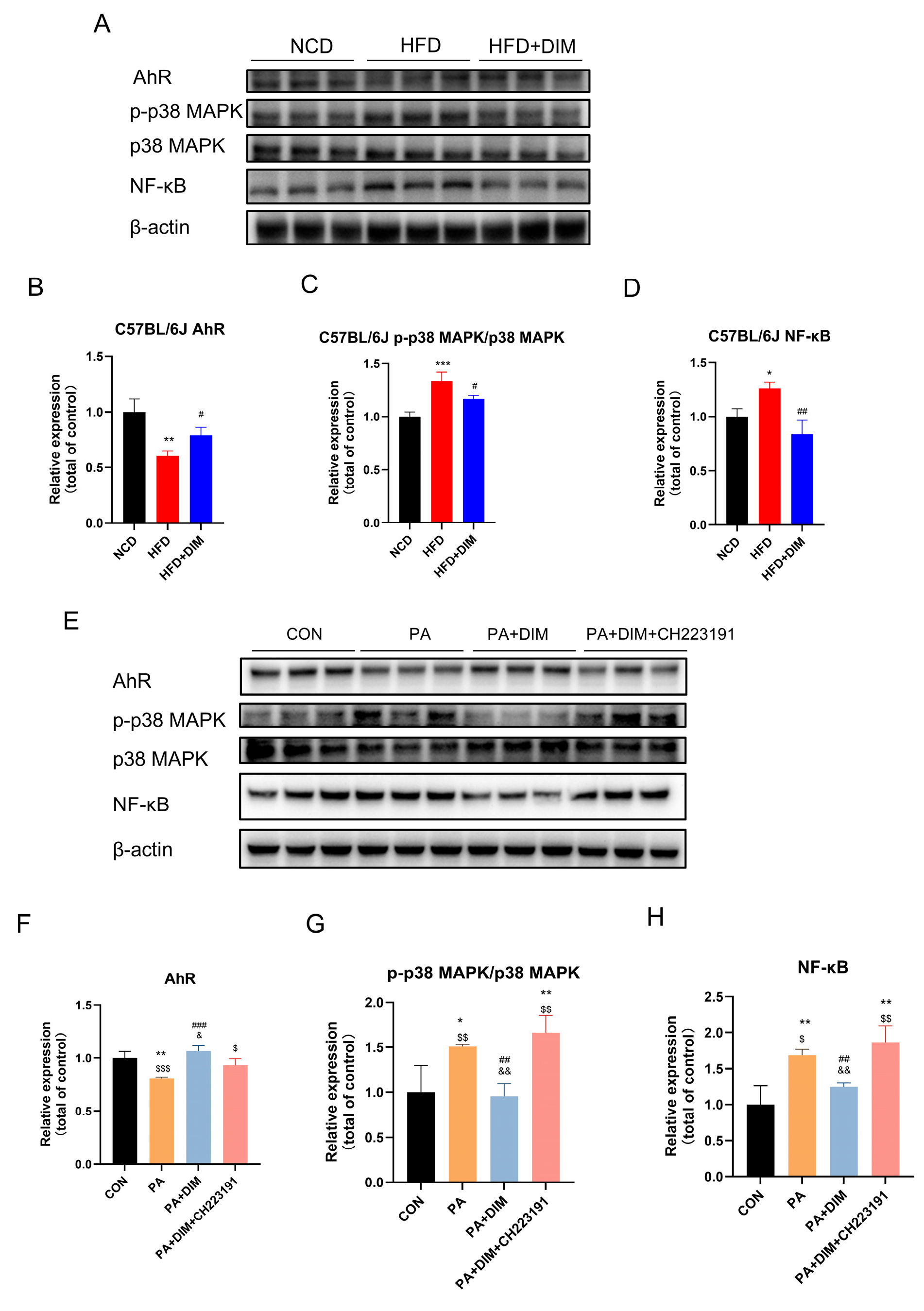
3.3. DIM Reduced p38MAPK Phosphorylation in Mice and HepG2 Cells
3.4. DIM Attenuated Lipid Transport in Mice and HepG2 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wai-Sun Wong, V.; Dufour, J.F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef]
- Li, J.; Zou, B.; Yeo, Y.H.; Feng, Y.; Xie, X.; Lee, D.H.; Fujii, H.; Wu, Y.; Kam, L.Y.; Ji, F.; et al. Prevalence, incidence, and outcome of non-alcoholic fatty liver disease in Asia, 1999–2019: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2019, 4, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yu, Y.; Cai, J.; Li, H. Emerging Molecular Targets for Treatment of Nonalcoholic Fatty Liver Disease. Trends Endocrinol. Metab. 2019, 30, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Dalbeni, A. Treatments for NAFLD: State of Art. Int. J. Mol. Sci. 2021, 22, 2350. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Li, Q.; He, Y.; Zhang, X.; Zhou, Z.; Gao, Y.; Fang, M.; Yu, Z.; Rodrigues, R.M.; Gao, Y.; et al. Therapeutic potential of traditional Chinese medicine for the treatment of NAFLD: A promising drug Potentilla discolor Bunge. Acta Pharm. Sin. B 2022, 12, 3529–3547. [Google Scholar] [CrossRef]
- Banerjee, S.; Kong, D.; Wang, Z.; Bao, B.; Hillman, G.G.; Sarkar, F.H. Attenuation of multi-targeted proliferation-linked signaling by 3,3′-diindolylmethane (DIM): From bench to clinic. Mutat. Res. 2011, 728, 47–66. [Google Scholar] [CrossRef]
- Lin, L.P.; Liu, D.; Qian, J.C.; Wu, L.; Zhao, Q.; Tan, R.X. Post-ingestion conversion of dietary indoles into anticancer agents. Natl. Sci. Rev. 2022, 9, nwab144. [Google Scholar] [CrossRef]
- Dong, L.; Xia, S.; Gao, F.; Zhang, D.; Chen, J.; Zhang, J. 3,3′-Diindolylmethane attenuates experimental arthritis and osteoclastogenesis. Biochem. Pharmacol. 2010, 79, 715–721. [Google Scholar] [CrossRef]
- Garcia-Villatoro, E.L.; Bomstein, Z.S.; Allred, K.F.; Callaway, E.S.; Safe, S.; Chapkin, R.S.; Jayaraman, A.; Allred, C.D. Involvement of Intestinal Epithelium Aryl Hydrocarbon Receptor Expression and 3, 3′-Diindolylmethane in Colonic Tertiary Lymphoid Tissue Formation. Int. J. Mol. Sci. 2024, 25, 10153. [Google Scholar] [CrossRef]
- Golberg, K.; Markus, V.; Kagan, B.E.; Barzanizan, S.; Yaniv, K.; Terali, K.; Kramarsky-Winter, E.; Marks, R.S.; Kushmaro, A. Anti-Virulence Activity of 3,3′-Diindolylmethane (DIM): A Bioactive Cruciferous Phytochemical with Accelerated Wound Healing Benefits. Pharmaceutics 2022, 14, 967. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Ahmad, A.; Bao, B.; Sarkar, F.H. Antioxidant function of isoflavone and 3,3′-diindolylmethane: Are they important for cancer prevention and therapy? Antioxid. Redox Signal 2013, 19, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Um, S.J.; Park, T. Indole-3-carbinol directly targets SIRT1 to inhibit adipocyte differentiation. Int. J. Obes. 2013, 37, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Deng, S.; Yang, M.; Bai, B.; Pan, Y.; Xie, G.; Zhao, D.; Wei, L. Indole-3-carbinol and its main derivative 3,3′-diindolylmethane: Regulatory roles and therapeutic potential in liver diseases. Biomed. Pharmacother. 2024, 180, 117525. [Google Scholar] [CrossRef]
- Baldelli, S.; Lombardo, M.; D’Amato, A.; Karav, S.; Tripodi, G.; Aiello, G. Glucosinolates in Human Health: Metabolic Pathways, Bioavailability, and Potential in Chronic Disease Prevention. Foods 2025, 14, 912. [Google Scholar] [CrossRef]
- Van der Heiden, E.; Bechoux, N.; Muller, M.; Sergent, T.; Schneider, Y.J.; Larondelle, Y.; Maghuin-Rogister, G.; Scippo, M.L. Food flavonoid aryl hydrocarbon receptor-mediated agonistic/antagonistic/synergic activities in human and rat reporter gene assays. Anal. Chim. Acta 2009, 637, 337–345. [Google Scholar] [CrossRef]
- Abd-Elmawla, M.A.; ElMonier, A.A.; Gad, E.S.; Khidr, H.Y.; Azzam, M.A. Implication of Let7b/AhR/ARNT/HMGB1/RAGE cascade in neuroplasticity disturbances induced by glucocorticoids and the promising reversible effect of 3,3 diindolymethane: Bidirectional crosstalk of Aryl hydrocarbon receptors. Biochem. Pharmacol. 2025, 232, 116692. [Google Scholar] [CrossRef] [PubMed]
- Carambia, A.; Schuran, F.A. The aryl hydrocarbon receptor in liver inflammation. Semin. Immunopathol. 2021, 43, 563–575. [Google Scholar] [CrossRef]
- Tan, Z.; Chang, X.; Puga, A.; Xia, Y. Activation of mitogen-activated protein kinases (MAPKs) by aromatic hydrocarbons: Role in the regulation of aryl hydrocarbon receptor (AHR) function. Biochem. Pharmacol. 2002, 64, 771–780. [Google Scholar] [CrossRef]
- Weiss, C.; Faust, D.; Durk, H.; Kolluri, S.K.; Pelzer, A.; Schneider, S.; Dietrich, C.; Oesch, F.; Gottlicher, M. TCDD induces c-jun expression via a novel Ah (dioxin) receptor-mediated p38-MAPK-dependent pathway. Oncogene 2005, 24, 4975–4983. [Google Scholar] [CrossRef]
- Li, H.; Hong, W.; Jin, X.; Li, G.; Zhou, G.; Fan, L. The aryl hydrocarbon receptor is a novel negative regulator of interleukin-17-mediated signaling and inflammation in vitro. FEBS Lett. 2019, 593, 952–961. [Google Scholar] [CrossRef]
- Coulthard, L.R.; White, D.E.; Jones, D.L.; McDermott, M.F.; Burchill, S.A. p38(MAPK): Stress responses from molecular mechanisms to therapeutics. Trends Mol. Med. 2009, 15, 369–379. [Google Scholar] [CrossRef]
- Tang, P.; Low, H.B.; Png, C.W.; Torta, F.; Kumar, J.K.; Lim, H.Y.; Zhou, Y.; Yang, H.; Angeli, V.; Shabbir, A.; et al. Protective Function of Mitogen-Activated Protein Kinase Phosphatase 5 in Aging- and Diet-Induced Hepatic Steatosis and Steatohepatitis. Hepatol. Commun. 2019, 3, 748–762. [Google Scholar] [CrossRef] [PubMed]
- Darlyuk-Saadon, I.; Bai, C.; Heng, C.K.M.; Gilad, N.; Yu, W.P.; Lim, P.Y.; Cazenave-Gassiot, A.; Zhang, Y.; Wong, W.S.F.; Engelberg, D. Active p38alpha causes macrovesicular fatty liver in mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2018069118. [Google Scholar] [CrossRef] [PubMed]
- Voisset, E.; Oeztuerk-Winder, F.; Ruiz, E.J.; Ventura, J.J. p38alpha negatively regulates survival and malignant selection of transformed bronchioalveolar stem cells. PLoS ONE 2013, 8, e78911. [Google Scholar] [CrossRef]
- Jiang, Y.; Fang, Y.; Ye, Y.; Xu, X.; Wang, B.; Gu, J.; Aschner, M.; Chen, J.; Lu, R. Anti-Cancer Effects of 3, 3′-Diindolylmethane on Human Hepatocellular Carcinoma Cells Is Enhanced by Calcium Ionophore: The Role of Cytosolic Ca2+ and p38 MAPK. Front. Pharmacol. 2019, 10, 1167. [Google Scholar] [CrossRef]
- Rajoria, S.; Suriano, R.; Parmar, P.S.; Wilson, Y.L.; Megwalu, U.; Moscatello, A.; Bradlow, H.L.; Sepkovic, D.W.; Geliebter, J.; Schantz, S.P.; et al. 3,3′-diindolylmethane modulates estrogen metabolism in patients with thyroid proliferative disease: A pilot study. Thyroid 2011, 21, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Kotsopoulos, J.; Zhang, S.; Akbari, M.; Salmena, L.; Llacuachaqui, M.; Zeligs, M.; Sun, P.; Narod, S.A. BRCA1 mRNA levels following a 4–6-week intervention with oral 3,3′-diindolylmethane. Br. J. Cancer 2014, 111, 1269–1274. [Google Scholar] [CrossRef]
- Reed, G.A.; Sunega, J.M.; Sullivan, D.K.; Gray, J.C.; Mayo, M.S.; Crowell, J.A.; Hurwitz, A. Single-dose pharmacokinetics and tolerability of absorption-enhanced 3,3′-diindolylmethane in healthy subjects. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2619–2624. [Google Scholar] [CrossRef]
- Boccatonda, A.; Andreetto, L.; D’Ardes, D.; Cocco, G.; Rossi, I.; Vicari, S.; Schiavone, C.; Cipollone, F.; Guagnano, M.T. From NAFLD to MAFLD: Definition, Pathophysiological Basis and Cardiovascular Implications. Biomedicines 2023, 11, 883. [Google Scholar] [CrossRef]
- Hammoutene, A.; Laouirem, S.; Albuquerque, M.; Colnot, N.; Brzustowski, A.; Valla, D.; Provost, N.; Delerive, P.; Paradis, V.; QUID-NASH Research Group. A new NRF2 activator for the treatment of human metabolic dysfunction-associated fatty liver disease. JHEP Rep. 2023, 5, 100845. [Google Scholar] [CrossRef]
- Luo, J.; Chen, M.; Ji, H.; Su, W.; Song, W.; Zhang, D.; Su, W.; Liu, S. Hypolipidemic and Anti-Obesity Effect of Anserine on Mice Orally Administered with High-Fat Diet via Regulating SREBP-1, NLRP3, and UCP-1. Mol. Nutr. Food Res. 2024, 68, e2300471. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.Y.; Ge, L.H.; Zhang, K.; Chen, H.F.; Zhan, X.X.; Yang, Y.; Dang, Q.L.; Zheng, Y.; Zhou, H.B.; Lyu, J.X.; et al. Assessment of mitochondrial function in metabolic dysfunction-associated fatty liver disease using obese mouse models. Zool. Res. 2020, 41, 539–551. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R. The past and present of serum aminotransferases and the future of liver injury biomarkers. EXCLI J. 2016, 15, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Stockinger, B.; Shah, K.; Wincent, E. AHR in the intestinal microenvironment: Safeguarding barrier function. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 559–570. [Google Scholar] [CrossRef]
- Yao, Y.; Luo, Z.P.; Li, H.W.; Wang, S.X.; Wu, Y.C.; Hu, Y.; Hu, S.; Yang, C.C.; Yang, J.F.; Wang, J.P.; et al. P38gamma modulates the lipid metabolism in non-alcoholic fatty liver disease by regulating the JAK-STAT signaling pathway. FASEB J. 2023, 37, e22716. [Google Scholar] [CrossRef]
- Wu, Y.K.; Ren, Z.N.; Zhu, S.L.; Wu, Y.Z.; Wang, G.; Zhang, H.; Chen, W.; He, Z.; Ye, X.L.; Zhai, Q.X. Sulforaphane ameliorates non-alcoholic fatty liver disease in mice by promoting FGF21/FGFR1 signaling pathway. Acta Pharmacol. Sin. 2022, 43, 1473–1483. [Google Scholar] [CrossRef]
- Jiang, J.; Yan, L.; Shi, Z.; Wang, L.; Shan, L.; Efferth, T. Hepatoprotective and anti-inflammatory effects of total flavonoids of Qu Zhi Ke (peel of Citrus changshan-huyou) on non-alcoholic fatty liver disease in rats via modulation of NF-kappaB and MAPKs. Phytomedicine 2019, 64, 153082. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, M.; Wang, C.; Guan, S.; Wang, L.; Cong, B.; Zhu, W.; Xu, Y. Low-molecular-weight fucoidan bidirectionally regulates lipid uptake and cholesterol efflux through the p38 MAPK phosphorylation. Int. J. Biol. Macromol. 2022, 220, 371–384. [Google Scholar] [CrossRef]
- Liu, Q.; Fan, J.; Bai, J.; Peng, L.; Zhang, T.; Deng, L.; Wang, G.; Zhao, Y.; Nong, J.; Zhang, M.; et al. IL-34 promotes foam cell formation by enhancing CD36 expression through p38 MAPK pathway. Sci. Rep. 2018, 8, 17347. [Google Scholar] [CrossRef]
- Rada, P.; Gonzalez-Rodriguez, A.; Garcia-Monzon, C.; Valverde, A.M. Understanding lipotoxicity in NAFLD pathogenesis: Is CD36 a key driver? Cell Death Dis. 2020, 11, 802. [Google Scholar] [CrossRef]
- Li, C.; Song, Z.; Gao, P.; Duan, W.; Liu, X.; Liang, S.; Gong, Q.; Guo, J. Transaldolase inhibits CD36 expression by modulating glutathione-p38 signaling, exerting protective effects against macrophage foam cell formation. Acta Biochim. Biophys. Sin. 2023, 55, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Herrmann, T.; Seessle, J.; Liebisch, G.; Merle, U.; Stremmel, W.; Chamulitrat, W. Role of fatty acid transport protein 4 in metabolic tissues: Insights into obesity and fatty liver disease. Biosci. Rep. 2022, 42, BSR20211854. [Google Scholar] [CrossRef]
- Gocebe, D.; Jansakun, C.; Zhang, Y.; Staffer, S.; Tuma-Kellner, S.; Altamura, S.; Muckenthaler, M.U.; Merle, U.; Herrmann, T.; Chamulitrat, W. Myeloid-specific fatty acid transport protein 4 deficiency induces a sex-dimorphic susceptibility for nonalcoholic steatohepatitis in mice fed a high-fat, high-cholesterol diet. Am. J. Physiol. Gastrointest. Liver Physiol. 2023, 324, G389–G403. [Google Scholar] [CrossRef] [PubMed]
- Krammer, J.; Digel, M.; Ehehalt, F.; Stremmel, W.; Fullekrug, J.; Ehehalt, R. Overexpression of CD36 and acyl-CoA synthetases FATP2, FATP4 and ACSL1 increases fatty acid uptake in human hepatoma cells. Int. J. Med. Sci. 2011, 8, 599–614. [Google Scholar] [CrossRef]
- You, H.; Wen, X.; Wang, X.; Zhu, C.; Chen, H.; Bu, L.; Zhang, J.; Qu, S. Derlin-1 ameliorates nonalcoholic hepatic steatosis by promoting ubiquitylation and degradation of FABP1. Free Radic. Biol. Med. 2023, 207, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Chang, C.C.; Wang, C.P.; Hung, W.C.; Tsai, I.T.; Tang, W.H.; Wu, C.C.; Wei, C.T.; Chung, F.M.; Lee, Y.J.; et al. Circulating fatty acid-binding protein 1 (FABP1) and nonalcoholic fatty liver disease in patients with type 2 diabetes mellitus. Int. J. Med. Sci. 2020, 17, 182–190. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, K.; Zhang, W.; Guo, W.; Wang, Y.; Zan, L.; Yang, W. Fatty acid-binding protein 1 increases steer fat deposition by facilitating the synthesis and secretion of triacylglycerol in liver. PLoS ONE 2019, 14, e0214144. [Google Scholar] [CrossRef]
- En Li Cho, E.; Ang, C.Z.; Quek, J.; Fu, C.E.; Lim, L.K.E.; Heng, Z.E.Q.; Tan, D.J.H.; Lim, W.H.; Yong, J.N.; Zeng, R.; et al. Global prevalence of non-alcoholic fatty liver disease in type 2 diabetes mellitus: An updated systematic review and meta-analysis. Gut 2023, 72, 2138–2148. [Google Scholar] [CrossRef]
- Deng, M.; Wen, Y.; Yan, J.; Fan, Y.; Wang, Z.; Zhang, R.; Ren, L.; Ba, Y.; Wang, H.; Lu, Q.; et al. Comparative effectiveness of multiple different treatment regimens for nonalcoholic fatty liver disease with type 2 diabetes mellitus: A systematic review and Bayesian network meta-analysis of randomised controlled trials. BMC Med. 2023, 21, 447. [Google Scholar] [CrossRef]
- Abdelmalek, M.F.; Harrison, S.A.; Sanyal, A.J. The role of glucagon-like peptide-1 receptor agonists in metabolic dysfunction-associated steatohepatitis. Diabetes Obes. Metab. 2024, 26, 2001–2016. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Das, S.; Samajdar, S.S.; Joshi, S.R. Role of semaglutide in the treatment of nonalcoholic fatty liver disease or non-alcoholic steatohepatitis: A systematic review and meta-analysis. Diabetes Metab. Syndr. 2023, 17, 102849. [Google Scholar] [CrossRef] [PubMed]
- Haigh, L.; Kirk, C.; El Gendy, K.; Gallacher, J.; Errington, L.; Mathers, J.C.; Anstee, Q.M. The effectiveness and acceptability of Mediterranean diet and calorie restriction in non-alcoholic fatty liver disease (NAFLD): A systematic review and meta-analysis. Clin. Nutr. 2022, 41, 1913–1931. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo, S.; Gomez-Martinez, S.; Diaz, L.E.; Nova, E.; Urrialde, R.; Marcos, A. Potential Effects of Sucralose and Saccharin on Gut Microbiota: A Review. Nutrients 2022, 14, 1682. [Google Scholar] [CrossRef] [PubMed]
- Brawner, K.M.; Yeramilli, V.A.; Duck, L.W.; Van Der Pol, W.; Smythies, L.E.; Morrow, C.D.; Elson, C.O.; Martin, C.A. Depletion of dietary aryl hydrocarbon receptor ligands alters microbiota composition and function. Sci. Rep. 2019, 9, 14724. [Google Scholar] [CrossRef]
- Inzaghi, E.; Pampanini, V.; Deodati, A.; Cianfarani, S. The Effects of Nutrition on Linear Growth. Nutrients 2022, 14, 1752. [Google Scholar] [CrossRef]
- Indrio, F.; Neu, J.; Pettoello-Mantovani, M.; Marchese, F.; Martini, S.; Salatto, A.; Aceti, A. Development of the Gastrointestinal Tract in Newborns as a Challenge for an Appropriate Nutrition: A Narrative Review. Nutrients 2022, 14, 1405. [Google Scholar] [CrossRef]
- Norman, K.; Hass, U.; Pirlich, M. Malnutrition in Older Adults-Recent Advances and Remaining Challenges. Nutrients 2021, 13, 2764. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item (mmol/L) | NCD | HFD | HFD + DIM |
|---|---|---|---|
| TG | 0.37 ± 0.36 | 0.56 ± 0.62 * | 0.43 ± 0.30 # |
| TC | 3.17 ± 1.48 | 4.58 ± 0.40 ** | 4.02 ± 0.33 |
| FFA | 0.58 ± 0.26 | 0.66 ± 0.26 | 0.55 ± 0.59 |
| LDL-c | 0.39 ± 0.25 | 0.84 ± 0.10 * | 0.56 ± 0.56 # |
| HDL-c | 8.37 ± 0.23 | 8.16 ± 0.30 | 9.47 ± 0.33 *## |
| Item (mmol/L) | NCD | HFD | HFD + DIM |
|---|---|---|---|
| TG | 17.06 ± 4.75 | 52.55 ± 8.78 ** | 24.92 ± 2.98 ## |
| TC | 11.88 ± 0.62 | 13.09 ± 0.29 | 10.14 ± 0.47 *# |
| FFA | 1.01 ± 1.77 | 1.29 ± 0.32 | 0.51 ± 0.77 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, J.; Fang, H.; Lin, Y.; Yao, Y.; Liu, Y.; Zhong, Y.; Li, X.; Sun, S.; Huang, B.; Yang, G.; et al. 3,3′-Diindolylmethane Ameliorates Metabolism Dysfunction-Associated Fatty Liver Disease via AhR/p38 MAPK Signaling. Nutrients 2025, 17, 1681. https://doi.org/10.3390/nu17101681
Su J, Fang H, Lin Y, Yao Y, Liu Y, Zhong Y, Li X, Sun S, Huang B, Yang G, et al. 3,3′-Diindolylmethane Ameliorates Metabolism Dysfunction-Associated Fatty Liver Disease via AhR/p38 MAPK Signaling. Nutrients. 2025; 17(10):1681. https://doi.org/10.3390/nu17101681
Chicago/Turabian StyleSu, Jiewen, Heng Fang, Yunfeng Lin, Yilu Yao, Yanxi Liu, Yuquan Zhong, Xudong Li, Siyu Sun, Bing Huang, Guangyu Yang, and et al. 2025. "3,3′-Diindolylmethane Ameliorates Metabolism Dysfunction-Associated Fatty Liver Disease via AhR/p38 MAPK Signaling" Nutrients 17, no. 10: 1681. https://doi.org/10.3390/nu17101681
APA StyleSu, J., Fang, H., Lin, Y., Yao, Y., Liu, Y., Zhong, Y., Li, X., Sun, S., Huang, B., Yang, G., Li, W., Zhang, Y., Li, J., Wu, J., Liu, W., Hu, Q., & Zhu, W. (2025). 3,3′-Diindolylmethane Ameliorates Metabolism Dysfunction-Associated Fatty Liver Disease via AhR/p38 MAPK Signaling. Nutrients, 17(10), 1681. https://doi.org/10.3390/nu17101681