Bifidobacterium animalis Subsp. lactis PB200 Improves Intestinal Barrier Function and Flora Disturbance in Mice with Antibiotic-Induced Intestinal Injury


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacteria Culture
2.3. Animals and Experimental Design
2.4. Serum Cytokines Measurements
2.5. Histological Analysis of Colon Tissue
2.6. Western Blot Analysis
2.7. Microbial DNA Extraction and 16S rRNA Gene Sequencing
2.8. Bioinformatics Analysis
2.9. Fecal Short-Chain Fatty Acids Analysis
2.10. Statistical Analysis
3. Results
3.1. Effects of B. lactis on Body Weight and Immune Organ Indices in Mice with Antibiotic-Induced Intestinal Injury
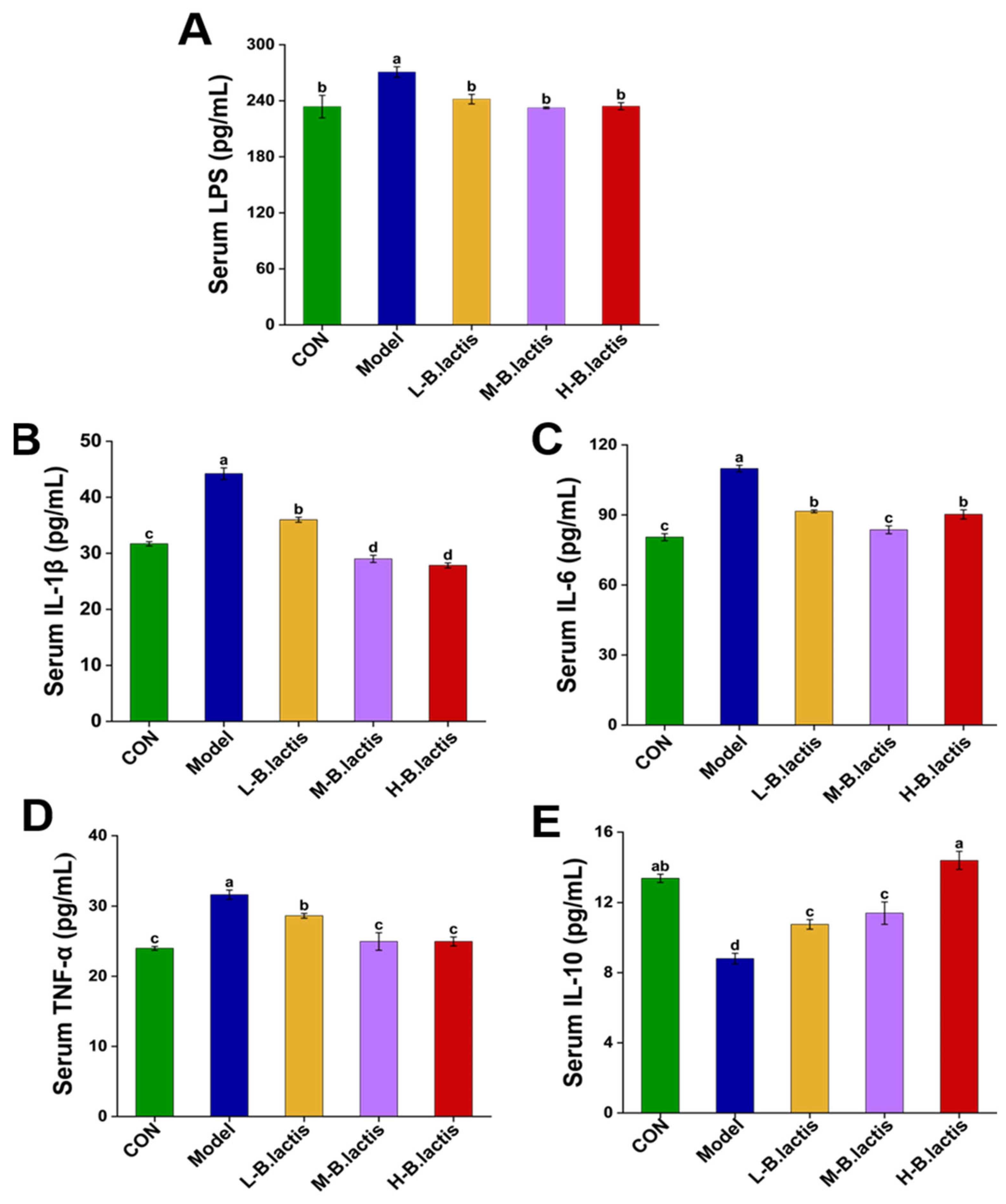
3.2. B. lactis Suppressed Inflammatory Response in Antibiotic-Induced Intestinal Injury Mice
3.3. Effects of B. lactis on Intestinal Barrier Integrity in Antibiotic-Induced Intestinal Injury Mice
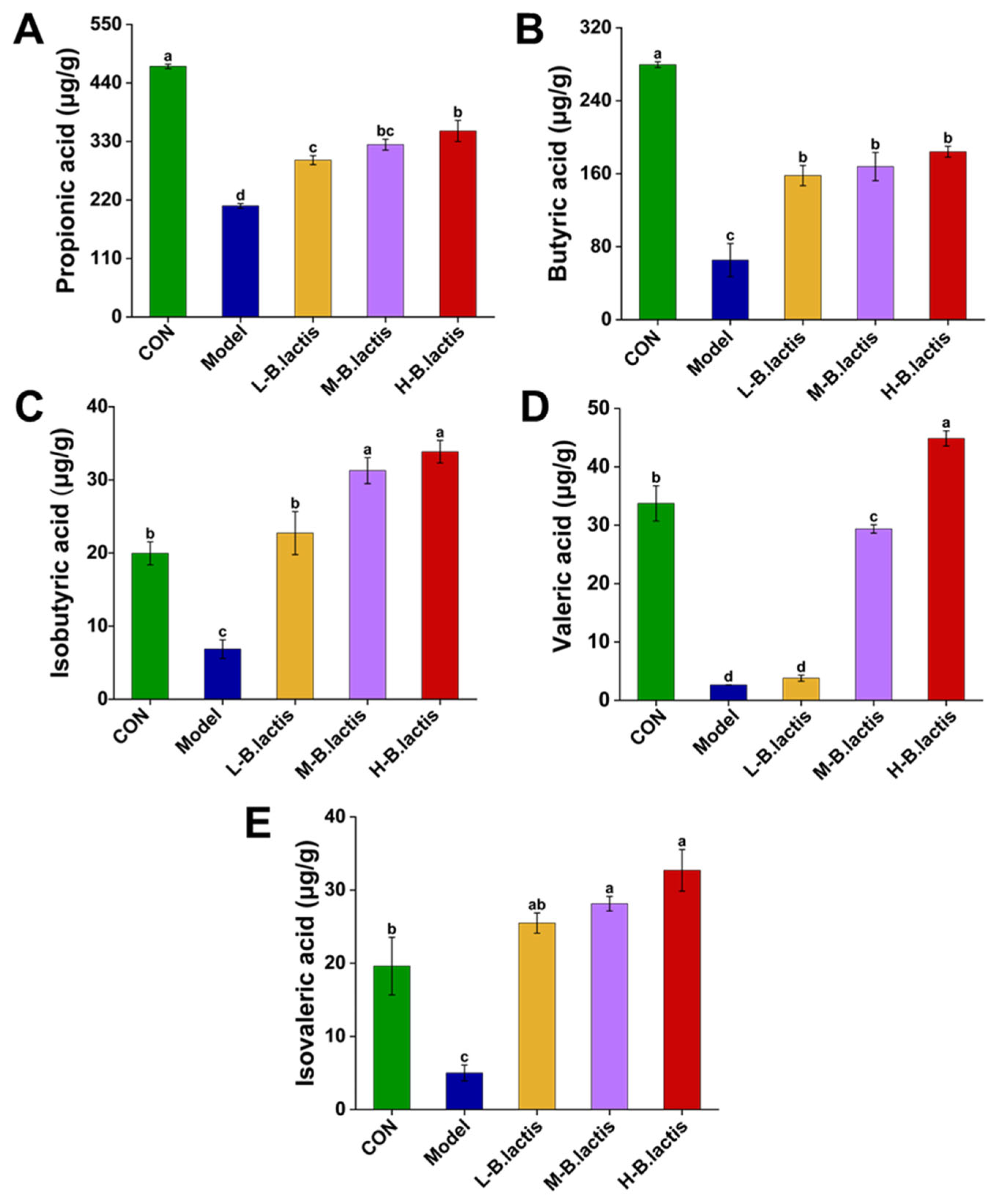
3.4. Effects of B. lactis on Fecal Short-Chain Fatty Acids (SCFAs) Levels in Antibiotic-Induced Intestinal Injury Mice
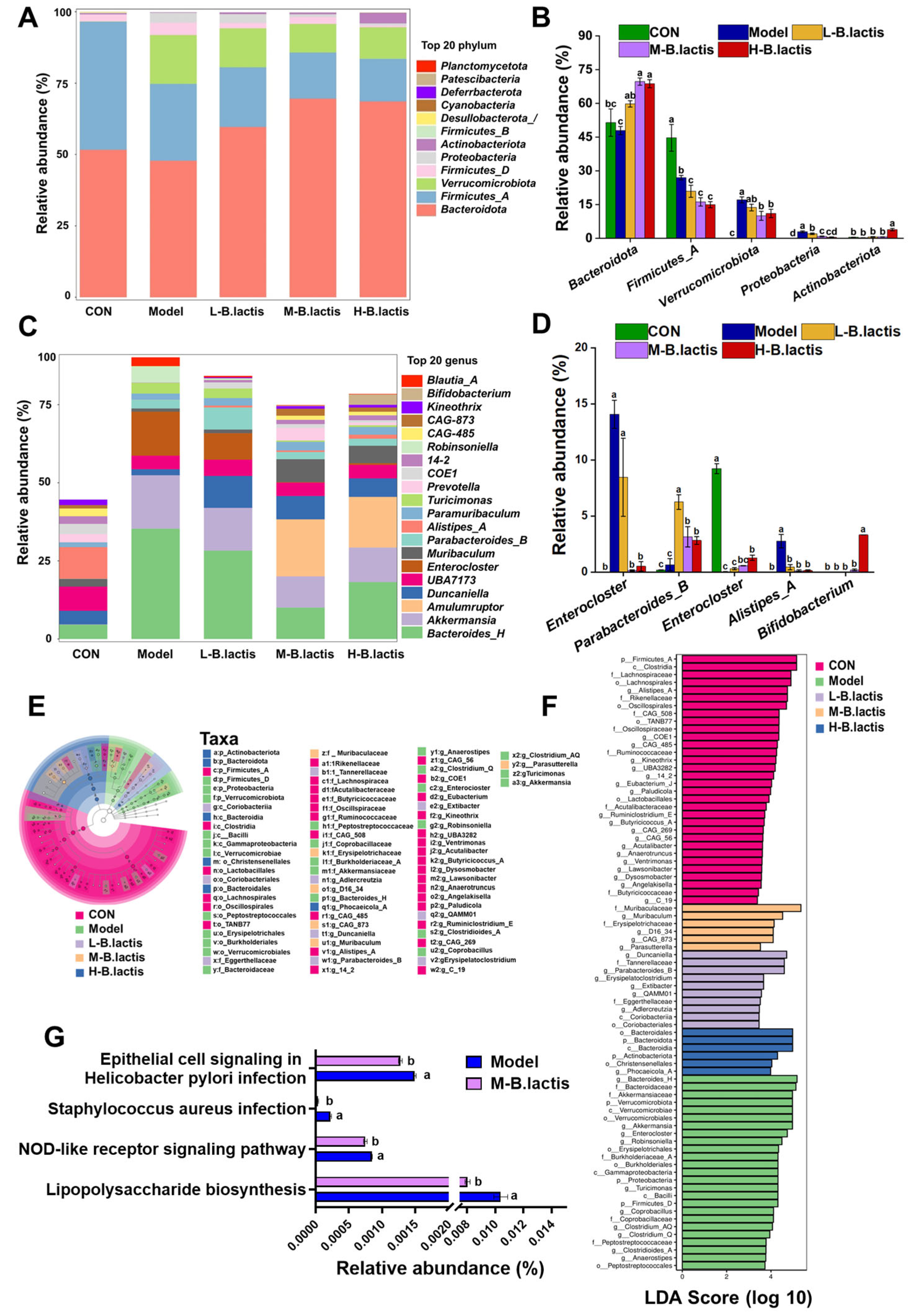
3.5. Effects of B. lactis on Microbiota Diversity in Mice with Antibiotic-Induced Intestinal Injury
3.5.1. Alpha Diversity
3.5.2. Beta Diversity
3.6. B. lactis Modulated Gut Microbiota Composition in Mice with Antibiotic-Induced Intestinal Injury
3.7. Correlation Analysis of Gut Microbiota with Representative Inflammatory Cytokines and Short-Chain Fatty Acids
4. Discussion
4.1. B. lactis PB200 Improved Intestinal Barrier Integrity and Gut Microbiota Dysbiosis in Antibiotic-Induced Intestinal Injury Mice
4.2. B. lactis PB200 May Restore Gut Homeostasis by Promoting SCFAs-Producing Bacteria and Enhancing Fecal SCFAs
4.3. B. lactis PB200 Had the Potential to Inhibit Inflammatory Pathways to Alleviate Intestinal Injury
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fishbein, S.R.S.; Mahmud, B.; Dantas, G. Antibiotic perturbations to the gut microbiome. Nat. Rev. Microbiol. 2023, 21, 772–788. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, S.; Farhadian, N.; Karimi, M.; Ebrahimi, M. Enhanced bactericidal effect of ceftriaxone drug encapsulated in nanostructured lipid carrier against gram-negative Escherichia coli bacteria: Drug formulation, optimization, and cell culture study. Antimicrob. Resist. Infect. Control 2020, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Liu, Y.; Zhang, W.; Lin, B.; Shen, W.; Li, Y.; He, Q.; Jin, J. Short-term use of ceftriaxone sodium leads to intestinal barrier disruption and ultrastructural changes of kidney in SD rats. Ren. Fail. 2023, 45, 2230322. [Google Scholar] [CrossRef] [PubMed]
- Cetinbas, M.; Thai, J.; Filatava, E.; Gregory, K.E.; Sadreyev, R.I. Long-term dysbiosis and fluctuations of gut microbiome in antibiotic treated preterm infants. iScience 2023, 26, 107995. [Google Scholar] [CrossRef]
- Ding, R.-X.; Goh, W.-R.; Wu, R.-N.; Yue, X.-Q.; Luo, X.; Khine, W.W.T.; Wu, J.-R.; Lee, Y.-K. Revisit gut microbiota and its impact on human health and disease. J. Food Drug Anal. 2019, 27, 623–631. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Wang, R.; Yu, Y.; Liu, X.; Tian, Z. Protective effect of sodium butyrate on intestinal barrier damage and uric acid reduction in hyperuricemia mice. Biomed. Pharmacother. 2023, 161, 114568. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Q.; Meng, M.; Chang, G.; Ma, N.; Shen, X. Butyrate Protects against γ-d-Glutamyl-meso-diaminopimelic Acid-Induced Inflammatory Response and Tight Junction Disruption through Histone Deacetylase 3 Inhibition in Bovine Mammary Epithelial Cells. J. Agric. Food Chem. 2023, 71, 14638–14648. [Google Scholar] [CrossRef]
- Li, P.; Li, M.; Song, Y.; Huang, X.; Wu, T.; Xu, Z.Z.; Lu, H. Green banana flour contributes to gut microbiota recovery and improves colonic barrier integrity in mice following antibiotic perturbation. Front. Immunol. 2022, 9, 832848. [Google Scholar] [CrossRef]
- Dethlefsen, L.; Huse, S.; Sogin, M.L.; Relman, D.A. The pervasive effects of an antibiotic on the human gut microbiota, as revealed by deep 16S rRNA sequencing. PLoS Biol. 2008, 6, 2383–2400. [Google Scholar] [CrossRef]
- Mishra, S.P.; Wang, B.; Jain, S.; Ding, J.; Rejeski, J.; Furdui, C.M.; Kitzman, D.W.; Taraphder, S.; Brechot, C.; Kumar, A.; et al. A mechanism by which gut microbiota elevates permeability and inflammation in obese/diabetic mice and human gut. Gut 2023, 72, 1848–1865. [Google Scholar] [CrossRef]
- Shao, H.; Zhang, C.; Xiao, N.; Tan, Z. Gut microbiota characteristics in mice with antibiotic-associated diarrhea. BMC Microbiol. 2020, 20, 313. [Google Scholar] [CrossRef] [PubMed]
- Vasilescu, I.M.; Chifiriuc, M.C.; Pircalabioru, G.G.; Filip, R.; Bolocan, A.; Lazăr, V.; Diţu, L.M.; Bleotu, C. Gut dysbiosis and clostridioides difficile Infection in neonates and adults. Front. Microbiol. 2021, 12, 651081. [Google Scholar] [CrossRef] [PubMed]
- Ekstedt, N.; Jamioł-Milc, D.; Pieczyńska, J. Importance of gut microbiota in patients with inflammatory bowel disease. Nutrients 2024, 16, 2092. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Xu, B.; Liang, S.; Zhao, J.; Li, X.; Guo, J.; Xin, B.; Li, B.; Huo, G.; Ma, W. Bifidobacterium animalis subsp. lactis XLTG11 improves antibiotic-related diarrhea by alleviating inflammation, enhancing intestinal barrier function and regulating intestinal flora. Food Funct. 2022, 13, 6404–6418. [Google Scholar] [CrossRef]
- Merenstein, D.; Fraser, C.M.; Roberts, R.F.; Liu, T.; Grant-Beurmann, S.; Tan, T.P.; Smith, K.H.; Cronin, T.; Martin, O.A.; Sanders, M.E.; et al. Bifidobacterium animalis subsp. lactis BB-12 protects against antibiotic-Induced functional and compositional changes in human fecal microbiome. Nutrients 2021, 13, 2814. [Google Scholar] [CrossRef]
- Lin, Y.; Xie, Z.; Li, Z.; Yuan, C.; Zhang, C.; Li, Y.; Xie, K.; Wang, K. The microbiota-gut-brain axis: A crucial immunomodulatory pathway for Bifidobacterium animalis subsp. lactis’ resilience against LPS treatment in neonatal rats. Int. J. Biol. Macromol. 2024, 266, 131255. [Google Scholar] [CrossRef]
- Lin, C.; Lin, Y.; Wang, S.; Wang, J.; Mao, X.; Zhou, Y.; Zhang, H.; Chen, W.; Wang, G. Bifidobacterium animalis subsp. lactis boosts neonatal immunity: Unravelling systemic defences against Salmonella. Food Funct. 2024, 15, 236–254. [Google Scholar] [CrossRef]
- Ikuta, K.; Joho, D.; Kakeyama, M.; Matsumoto, M. Bifidobacterium animalis subsp. lactis and arginine mixture intake improves cognitive flexibility in mice. Front. Nutr. 2023, 10, 1164809. [Google Scholar] [CrossRef]
- Huang, X.; Wang, J.; Liu, R.; Yang, C.; Shao, Y.; Wang, X.; Yi, H.; Lu, Y. An effective potential Bifidobacterium animalis F1-7 delivery strategy: Supramolecular hydrogel—Sodium alginate/tryptophan-sulfobutylether-β-cyclodextrin (Alg/Trp-SBE-β-CD). Food Hydrocoll. 2024, 155, 110217. [Google Scholar] [CrossRef]
- Yan, X.; Yan, J.; Xiang, Q.; Wang, F.; Dai, H.; Huang, K.; Fang, L.; Yao, H.; Wang, L.; Zhang, W. Fructooligosaccharides protect against OVA-induced food allergy in mice by regulating the Th17/Treg cell balance using tryptophan metabolites. Food Funct. 2021, 12, 3191–3205. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Peng, C.; Zhang, W.; Zhang, T.; He, Q.; Kwok, L.-Y.; Zhang, H. Postbiotic administration ameliorates colitis and inflammation in rats possibly through gut microbiota modulation. J. Agric. Food Chem. 2024, 72, 9054–9066. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Chen, C.C.; Lin, Y.T.; Wu, W.K.; Chang, L.C.; Lai, C.H.; Wu, M.S.; Kuo, C.H. Evaluation and optimization of sample handling methods for quantification of short-chain fatty acids in human fecal samples by GC-MS. J. Proteome Res. 2019, 18, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Barkhouse, A.; Hackenberger, D.; Wright, G.D. Antibiotic resistance: A key microbial survival mechanism that threatens public health. Cell Host Microbe 2024, 32, 837–851. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, B.; Mu, L.; Wang, H.; Luo, J.; Yang, Y.; Yang, H.; Li, M.; Zhou, L.; Tao, C. Long-term exposure to ceftriaxone sodium induces alteration of gut microbiota accompanied by abnormal behaviors in mice. Front. Cell. Infect. Microbiol. 2020, 10, 258. [Google Scholar] [CrossRef]
- Zhu, L.; Li, J.; Wei, C.; Luo, T.; Deng, Z.; Fan, Y.; Zheng, L. A polysaccharide from Fagopyrum esculentum Moench bee pollen alleviates microbiota dysbiosis to improve intestinal barrier function in antibiotic-treated mice. Food Funct. 2020, 11, 10519–10533. [Google Scholar] [CrossRef]
- Sovran, B.; Hugenholtz, F.; Elderman, M.; Van Beek, A.A.; Graversen, K.; Huijskes, M.; Boekschoten, M.V.; Savelkoul, H.F.J.; De Vos, P.; Dekker, J.; et al. Age-associated impairment of the mucus barrier function is associated with profound changes in microbiota and immunity. Sci. Rep. 2019, 9, 1437. [Google Scholar] [CrossRef]
- Zhou, F.; Lin, Y.; Chen, S.; Bao, X.; Fu, S.; Lv, Y.; Zhou, M.; Chen, Y.; Zhu, B.; Qian, C.; et al. Ameliorating role of Tetrastigma hemsleyanum polysaccharides in antibiotic-induced intestinal mucosal barrier dysfunction in mice based on microbiome and metabolome analyses. Int. J. Biol. Macromol. 2023, 241, 124419. [Google Scholar] [CrossRef]
- Yoon, H.; Schaubeck, M.; Lagkouvardos, I.; Blesl, A.; Heinzlmeir, S.; Hahne, H.; Clavel, T.; Panda, S.; Ludwig, C.; Kuster, B.; et al. Increased pancreatic protease activity in response to antibiotics impairs gut barrier and triggers colitis. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 370–388.e373. [Google Scholar] [CrossRef]
- Hu, R.; Yang, T.; Ai, Q.; Shi, Y.; Ji, Y.; Sun, Q.; Tong, B.; Chen, J.; Wang, Z. Autoinducer-2 promotes the colonization of Lactobacillus rhamnosus GG to improve the intestinal barrier function in a neonatal mouse model of antibiotic-induced intestinal dysbiosis. J. Transl. Med. 2024, 22, 177. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, J.; Zhang, H.; Lee, Y.-K.; Zhai, Q.; Chen, W. Roles of intestinal bacteroides in human health and diseases. Crit. Rev. Food Sci. Nutr. 2021, 61, 3518–3536. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Magdy Wasfy, R.; Mbaye, B.; Borentain, P.; Tidjani Alou, M.; Murillo Ruiz, M.L.; Caputo, A.; Andrieu, C.; Armstrong, N.; Million, M.; Gerolami, R. Ethanol-producing enterocloster bolteae is enriched in chronic hepatitis b-associated gut dysbiosis: A case-control culturomics study. Microorganisms 2023, 11, 2437. [Google Scholar] [CrossRef] [PubMed]
- Fidelle, M.; Rauber, C.; Alves Costa Silva, C.; Tian, A.-L.; Lahmar, I.; de La Varende, A.-L.M.; Zhao, L.; Thelemaque, C.; Lebhar, I.; Messaoudene, M.; et al. A microbiota-modulated checkpoint directs immunosuppressive intestinal T cells into cancers. Science 2023, 380, eabo2296. [Google Scholar] [CrossRef]
- Tang, W.; Yao, X.; Xia, F.; Yang, M.; Chen, Z.; Zhou, B.; Liu, Q. Modulation of the gut microbiota in rats by Hugan Qingzhi tablets during the treatment of high-fat-diet-induced nonalcoholic fatty liver disease. Oxid. Med. Cell. Longev. 2018, 2018, 7261619. [Google Scholar] [CrossRef]
- Hiippala, K.; Kainulainen, V.; Suutarinen, M.; Heini, T.; Bowers, J.R.; Jasso-Selles, D.; Lemmer, D.; Valentine, M.; Barnes, R.; Engelthaler, D.M.; et al. Isolation of anti-inflammatory and epithelium reinforcing Bacteroides and Parabacteroides Spp. from a healthy fecal donor. Nutrients 2020, 12, 935. [Google Scholar] [CrossRef]
- Gu, H.; Tao, E.; Fan, Y.; Long, G.; Jia, X.; Yuan, T.; Chen, L.; Shu, X.; Zheng, W.; Jiang, M. Effect of β-lactam antibiotics on the gut microbiota of term neonates. Ann. Clin. Microbiol. Antimicrob. 2024, 23, 69. [Google Scholar] [CrossRef]
- Wu, S.; Wu, Q.; Wang, J.; Li, Y.; Chen, B.; Zhu, Z.; Huang, R.; Chen, M.; Huang, A.; Xie, Y.J.J.o.A.; et al. Novel selenium peptides obtained from selenium-enriched cordyceps militaris alleviate neuroinflammation and gut microbiota dysbacteriosis in LPS-injured mice. J. Agric. Food Chem. 2022, 70, 3194–3206. [Google Scholar] [CrossRef]
- Grazul, H.; Kanda, L.L.; Gondek, D. Impact of probiotic supplements on microbiome diversity following antibiotic treatment of mice. Gut Microbes 2016, 7, 101–114. [Google Scholar] [CrossRef]
- Niu, H.; Zhou, X.; Gong, P.; Jiao, Y.; Zhang, J.; Wu, Y.; Lyu, L.; Liang, C.; Chen, S.; Han, X.; et al. Effect of Lactobacillus rhamnosus MN-431 Producing Indole Derivatives on Complementary Feeding-Induced Diarrhea Rat Pups Through the Enhancement of the Intestinal Barrier Function. Mol. Nutr. Food Res. 2022, 66, e2100619. [Google Scholar] [CrossRef]
- Zarrinpar, A.; Chaix, A.; Xu, Z.Z.; Chang, M.W.; Marotz, C.A.; Saghatelian, A.; Knight, R.; Panda, S. Antibiotic-induced microbiome depletion alters metabolic homeostasis by affecting gut signaling and colonic metabolism. Nat. Commun. 2018, 9, 2872. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qiu, X.; Yang, Y.; Wang, J.; Wang, Q.; Liu, J.; Yang, F.; Liu, Z.; Qi, R. Alteration of gut microbiome and metabolome by Clostridium butyricum can repair the intestinal dysbiosis caused by antibiotics in mice. iScience 2023, 26, 106190. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fang, H.; Chen, H.; Liu, X.; Zhao, J.; Stanton, C.; Ross, R.P.; Chen, W.; Yang, B. Bifidobacterium inhibits the progression of colorectal tumorigenesis in mice through fatty acid isomerization and gut microbiota modulation. Gut Microbes 2025, 17, 2464945. [Google Scholar] [CrossRef] [PubMed]
- Eloe-Fadrosh, E.A.; Brady, A.; Crabtree, J.; Drabek, E.F.; Ma, B.; Mahurkar, A.; Ravel, J.; Haverkamp, M.; Fiorino, A.M.; Botelho, C.; et al. Functional dynamics of the gut microbiome in elderly people during probiotic consumption. mBio 2015, 6, 10-1128. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The genus alistipes: Gut bacteria with emerging implications to inflammation, cancer, and mental health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Older, E.A.; Mitchell, M.K.; Campbell, A.; Lian, X.; Madden, M.; Wang, Y.; van de Wal, L.E.; Zaw, T.; VanderVeen, B.N.; Tatum, R.; et al. Human gut commensal Alistipes timonensis modulates the host lipidome and delivers anti-inflammatory outer membrane vesicles to suppress colitis in an Il10 -deficient mouse model. bioRxiv 2024. [Google Scholar] [CrossRef]
- Bein, A.; Zilbershtein, A.; Golosovsky, M.; Davidov, D.; Schwartz, B. LPS induces hyper-permeability of intestinal epithelial cells. J. Cell. Physiol. 2017, 232, 381–390. [Google Scholar] [CrossRef]
- Roger, T.; Froidevaux, C.; Le Roy, D.; Reymond, M.K.; Chanson, A.-L.; Mauri, D.; Burns, K.; Riederer, B.M.; Akira, S.; Calandra, T. Protection from lethal gram-negative bacterial sepsis by targeting Toll-like receptor 4. Proc. Natl. Acad. Sci. USA 2009, 106, 2348–2352. [Google Scholar] [CrossRef]
- Caetano-Silva, M.E.; Shrestha, A.; Duff, A.F.; Kontic, D.; Brewster, P.C.; Kasperek, M.C.; Lin, C.H.; Wainwright, D.A.; Hernandez-Saavedra, D.; Woods, J.A.; et al. Aging amplifies a gut microbiota immunogenic signature linked to heightened inflammation. Aging Cell 2024, 23, e14190. [Google Scholar] [CrossRef]
- Zhao, L.; Xie, Q.; Etareri Evivie, S.; Liu, D.; Dong, J.; Ping, L.; Liu, F.; Li, B.; Huo, G. Bifidobacterium dentium N8 with potential probiotic characteristics prevents LPS-induced intestinal barrier injury by alleviating the inflammatory response and regulating the tight junction in Caco-2 cell monolayers. Food Funct. 2021, 12, 7171–7184. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, Y.; Wang, Y.; Wang, P.; Song, H.; Wang, F. Antibiotics induced intestinal tight junction barrier dysfunction is associated with microbiota dysbiosis, activated NLRP3 inflammasome and autophagy. PLoS ONE 2019, 14, e0218384. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Chen, Y.; Xu, M. The critical role of short-chain fatty acids in health and disease: A subtle focus on cardiovascular disease-NLRP3 inflammasome-angiogenesis axis. Clin. Immunol. 2022, 238, 109013. [Google Scholar] [CrossRef] [PubMed]
- Galvão, I.; de Carvalho, R.V.H.; Vago, J.P.; Silva, A.L.N.; Carvalho, T.G.; Antunes, M.M.; Ribeiro, F.M.; Menezes, G.B.; Zamboni, D.S.; Sousa, L.P.; et al. The role of annexin A1 in the modulation of the NLRP3 inflammasome. Immunology 2020, 160, 78–89. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Gong, H.; Zou, Y.; Zhang, H.; Mao, X. Bifidobacterium animalis Subsp. lactis PB200 Improves Intestinal Barrier Function and Flora Disturbance in Mice with Antibiotic-Induced Intestinal Injury. Nutrients 2025, 17, 1610. https://doi.org/10.3390/nu17101610
Wang G, Gong H, Zou Y, Zhang H, Mao X. Bifidobacterium animalis Subsp. lactis PB200 Improves Intestinal Barrier Function and Flora Disturbance in Mice with Antibiotic-Induced Intestinal Injury. Nutrients. 2025; 17(10):1610. https://doi.org/10.3390/nu17101610
Chicago/Turabian StyleWang, Ganen, Han Gong, Yang Zou, Haijiao Zhang, and Xueying Mao. 2025. "Bifidobacterium animalis Subsp. lactis PB200 Improves Intestinal Barrier Function and Flora Disturbance in Mice with Antibiotic-Induced Intestinal Injury" Nutrients 17, no. 10: 1610. https://doi.org/10.3390/nu17101610
APA StyleWang, G., Gong, H., Zou, Y., Zhang, H., & Mao, X. (2025). Bifidobacterium animalis Subsp. lactis PB200 Improves Intestinal Barrier Function and Flora Disturbance in Mice with Antibiotic-Induced Intestinal Injury. Nutrients, 17(10), 1610. https://doi.org/10.3390/nu17101610