A Novel Lactobacillus brevis Fermented with a Vegetable Substrate (AL0035) Counteracts TNBS-Induced Colitis by Modulating the Gut Microbiota Composition and Intestinal Barrier

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Preparation
2.2. Animals and Experimental Design
2.3. Sample Collection and Evaluation of the Macroscopic Score
2.4. Colonic Histopathology
2.5. Myeloperoxidase (MPO) Activity
2.6. Relative Quantitative Real-Time PCR
2.7. Western Blot Analysis
2.8. Stool DNA Extraction and Microbiota Analysis
2.9. Statistical Analysis
3. Results
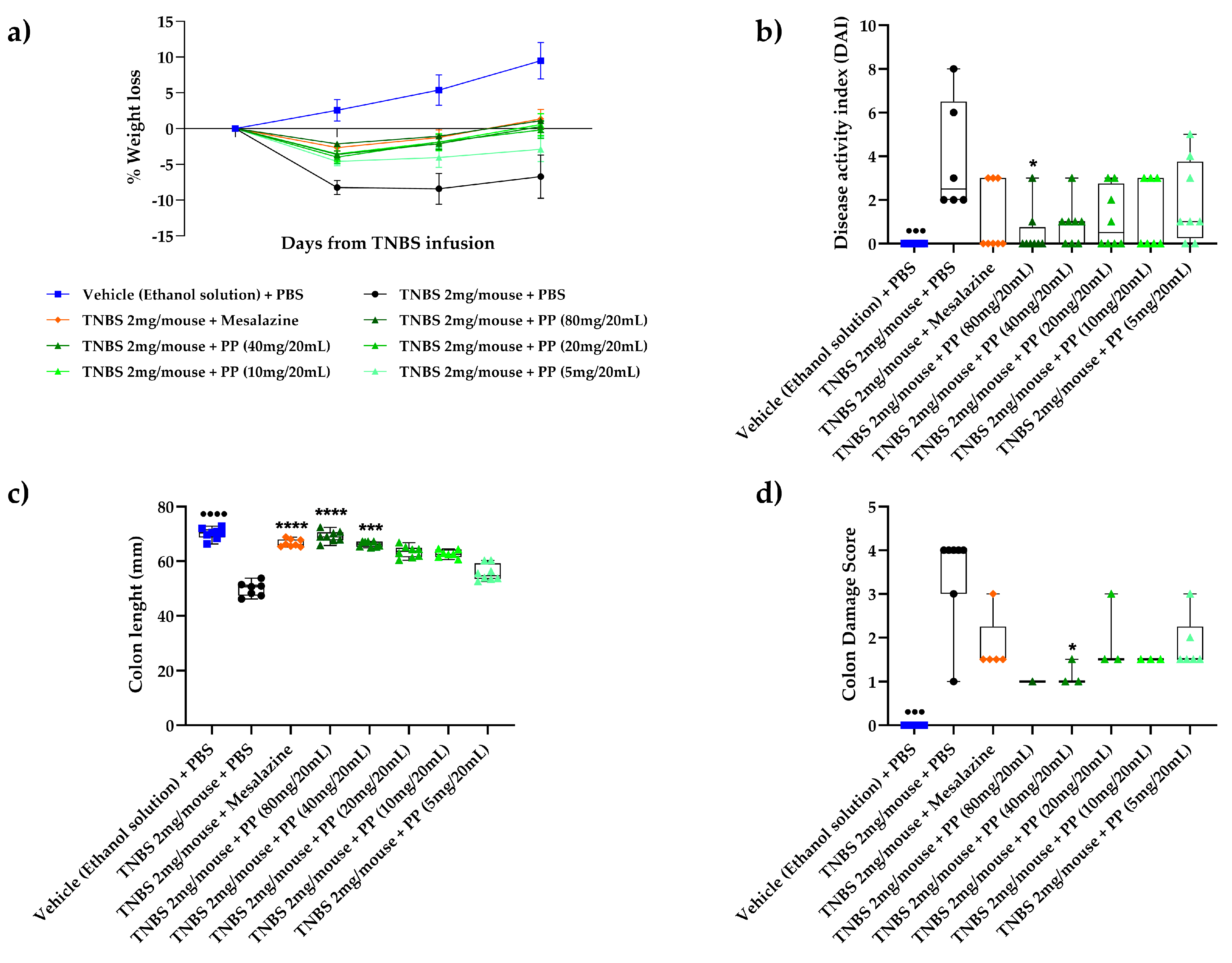
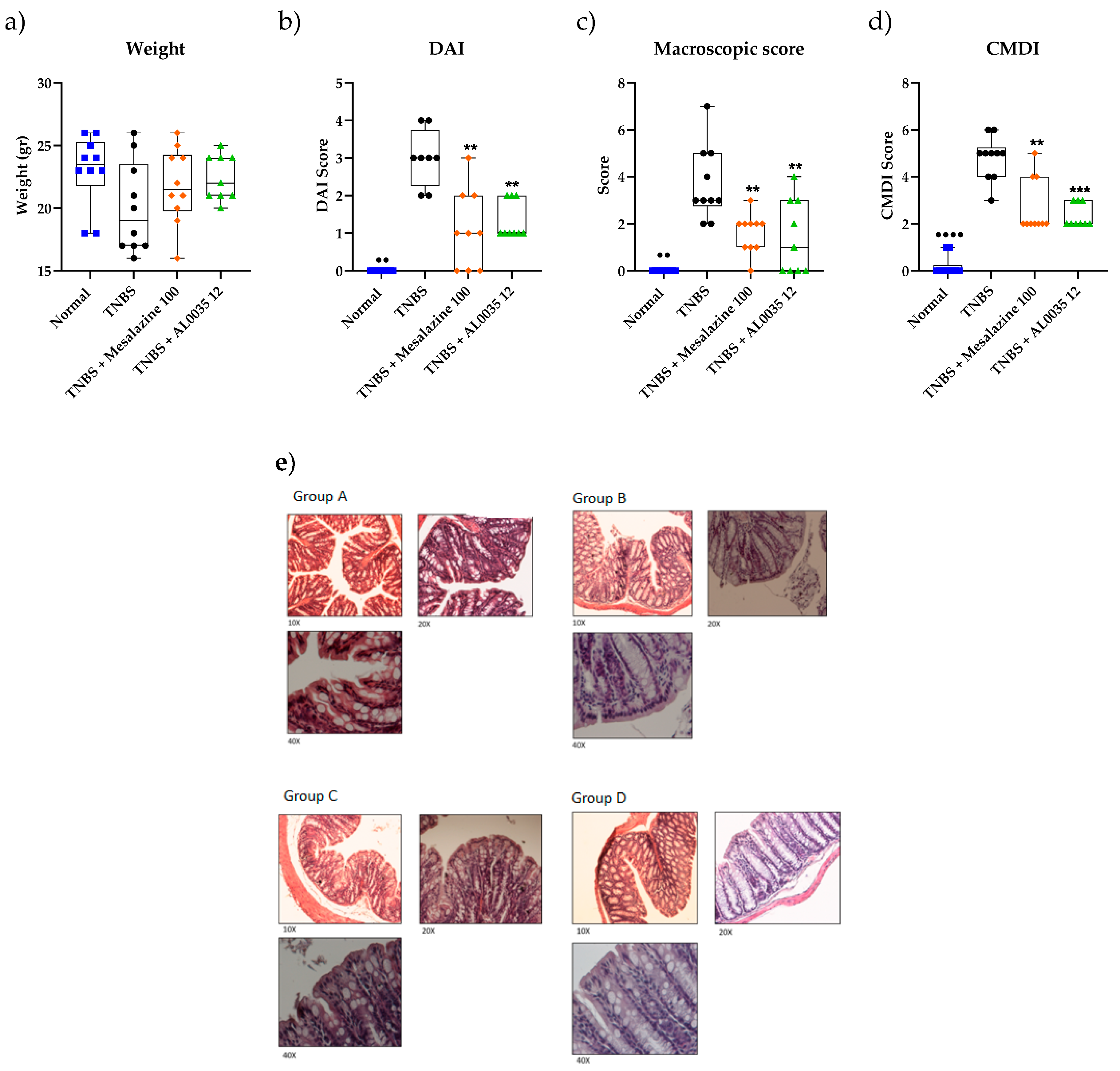
3.1. Lactobacillus brevis (AL0035) Alleviates Clinical Symptoms of TNBS-Induced Colitis in Mice
3.2. AL0035 Ameliorates Intestinal Injury in TNBS-Induced Colitis Mice
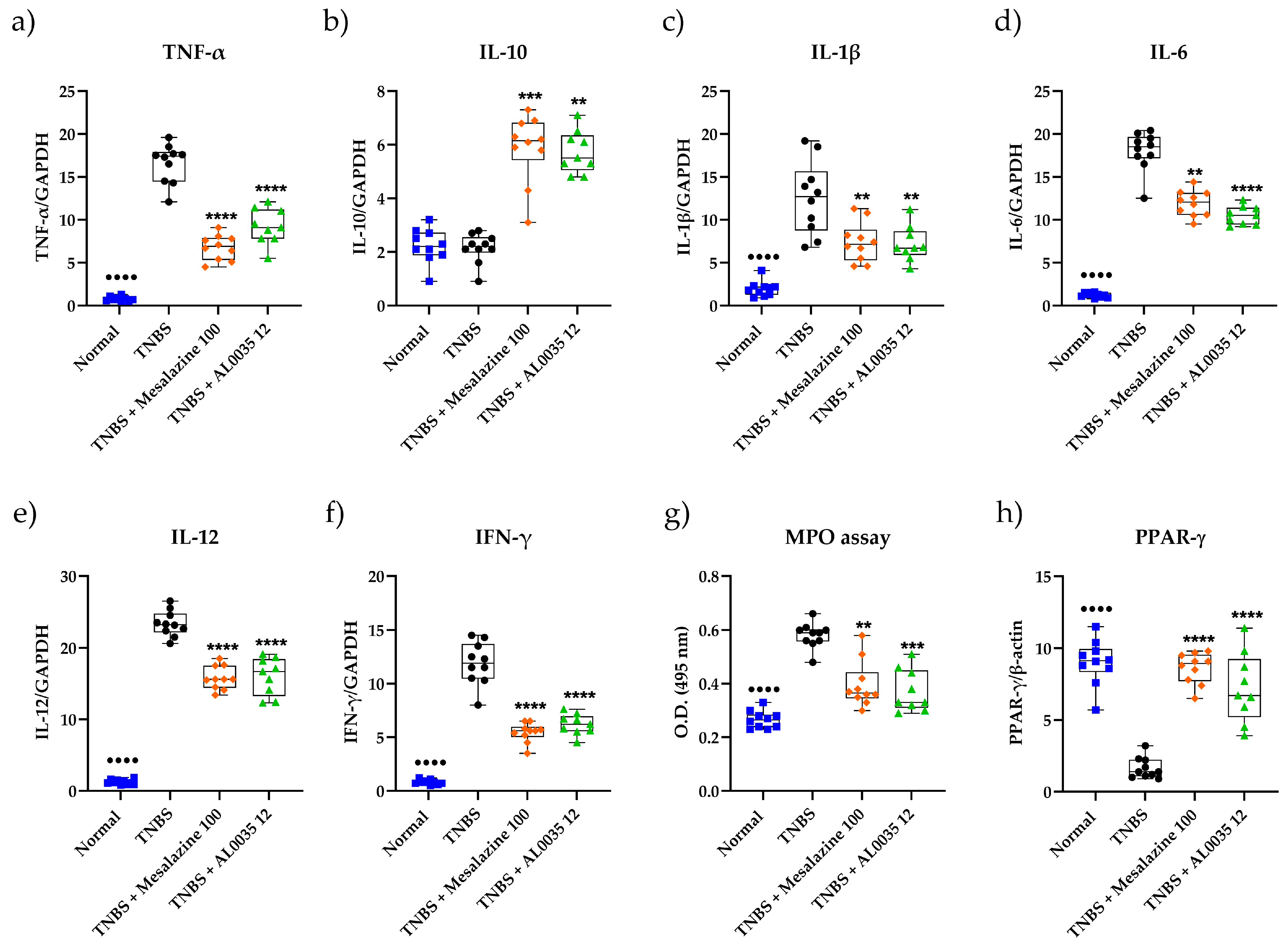
3.3. AL0035 Affects Inflammatory Cytokines, the PPAR-γ level and MPO Activity in the Colon
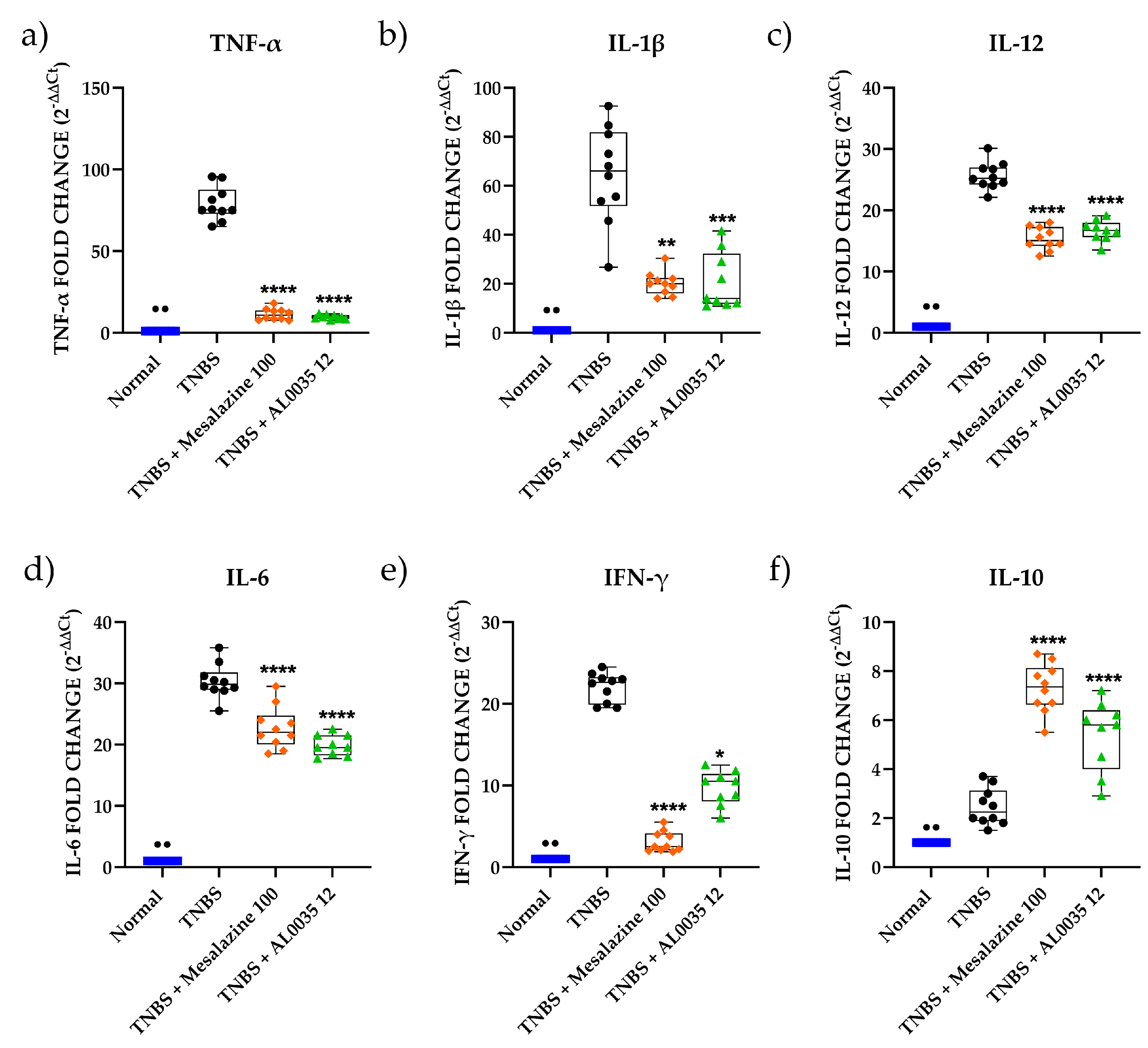
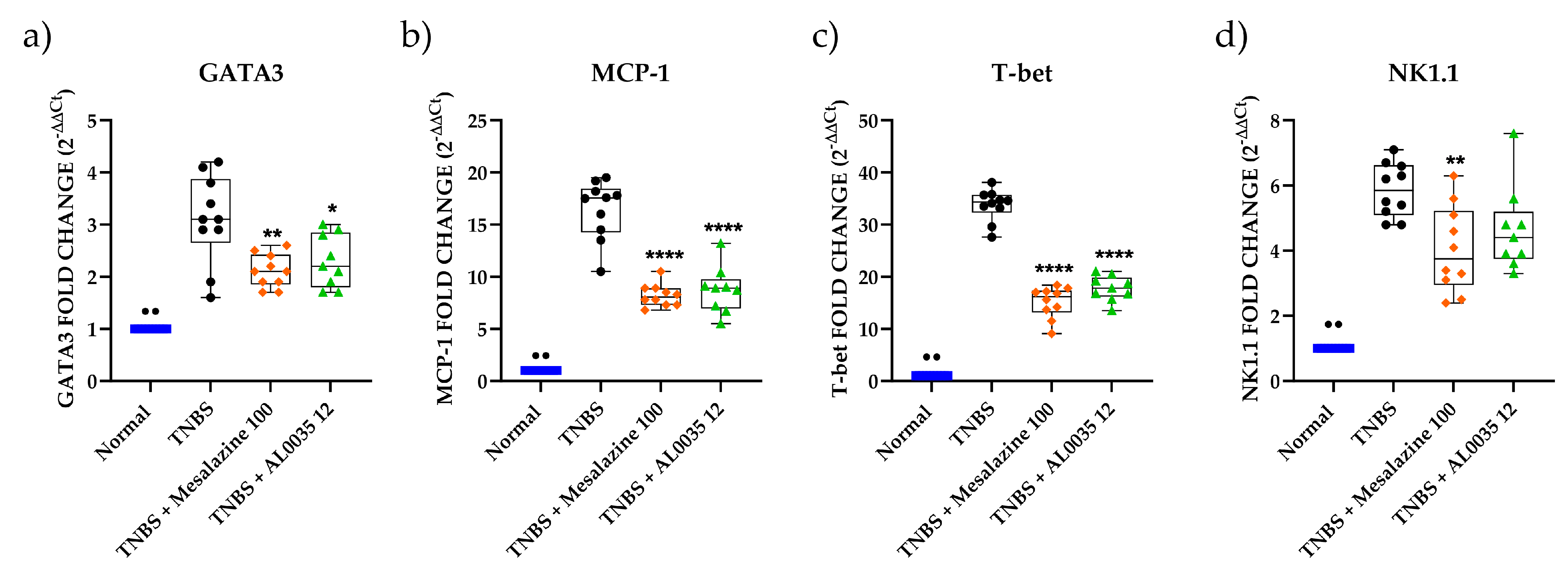
3.4. AL0035 Affects the Expression of Inflammatory Cytokines in TNBS-Treated Mice
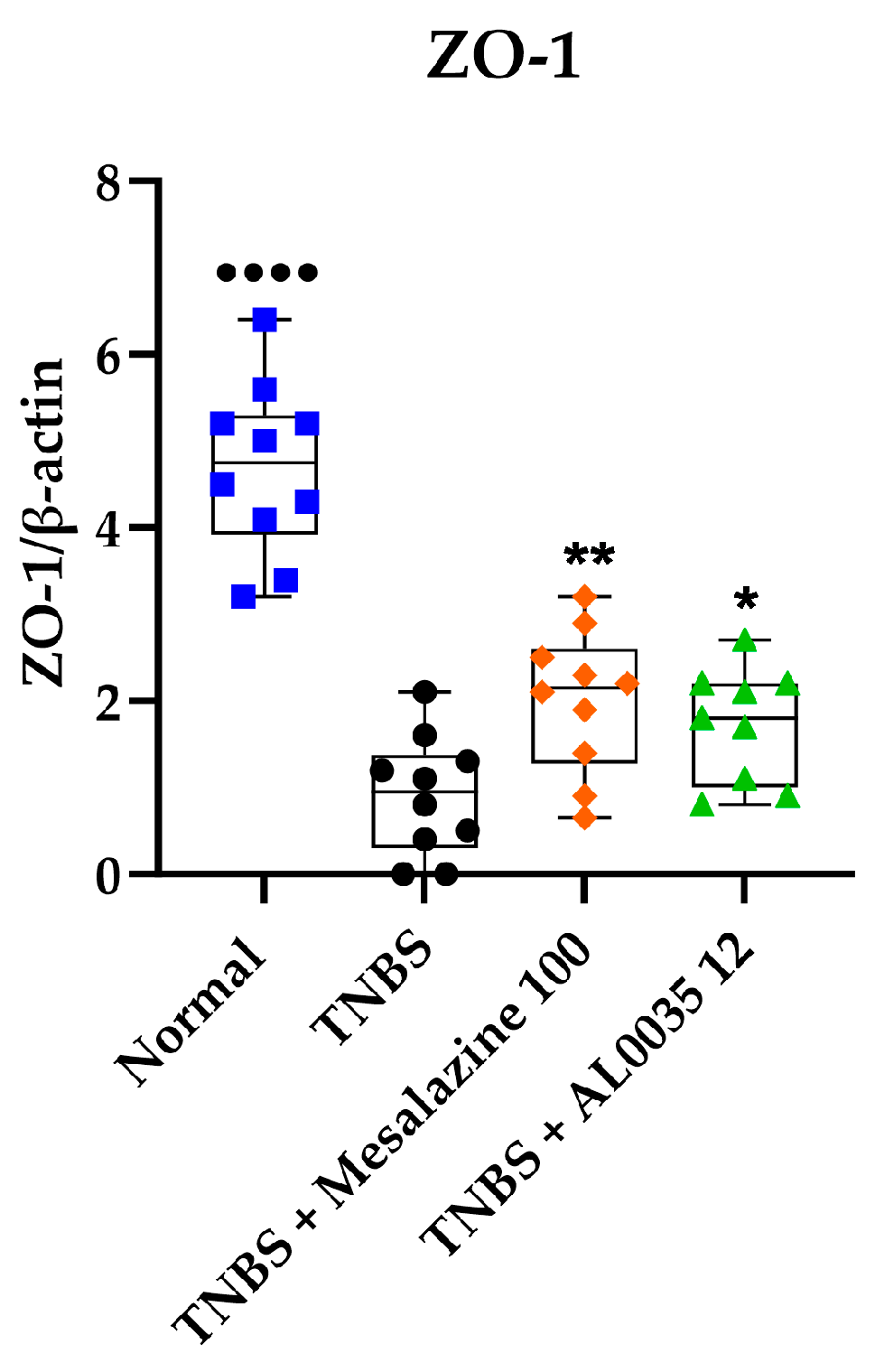
3.5. AL0035 Ameliorates the TNBS-Induced Tight Junction Protein Decrease
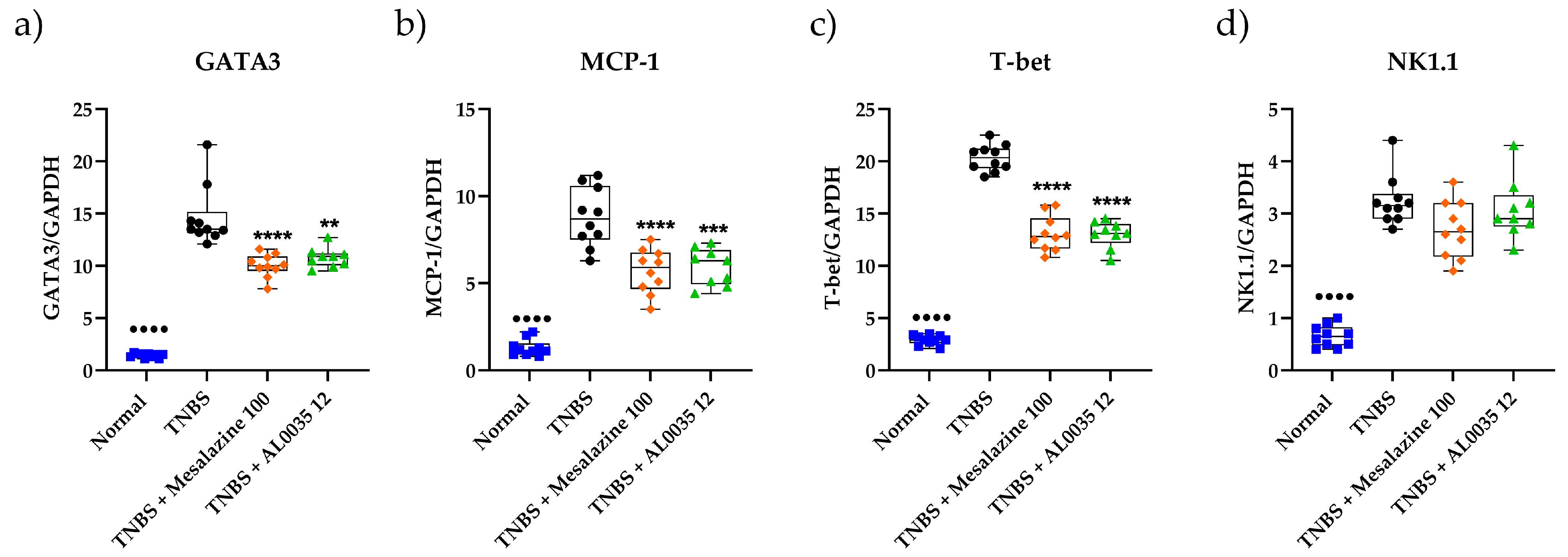
3.6. AL0035 Treatment Reduces Lymphocyte Markers Infiltration in the Colons of TNBS-Induced Colitis Mice
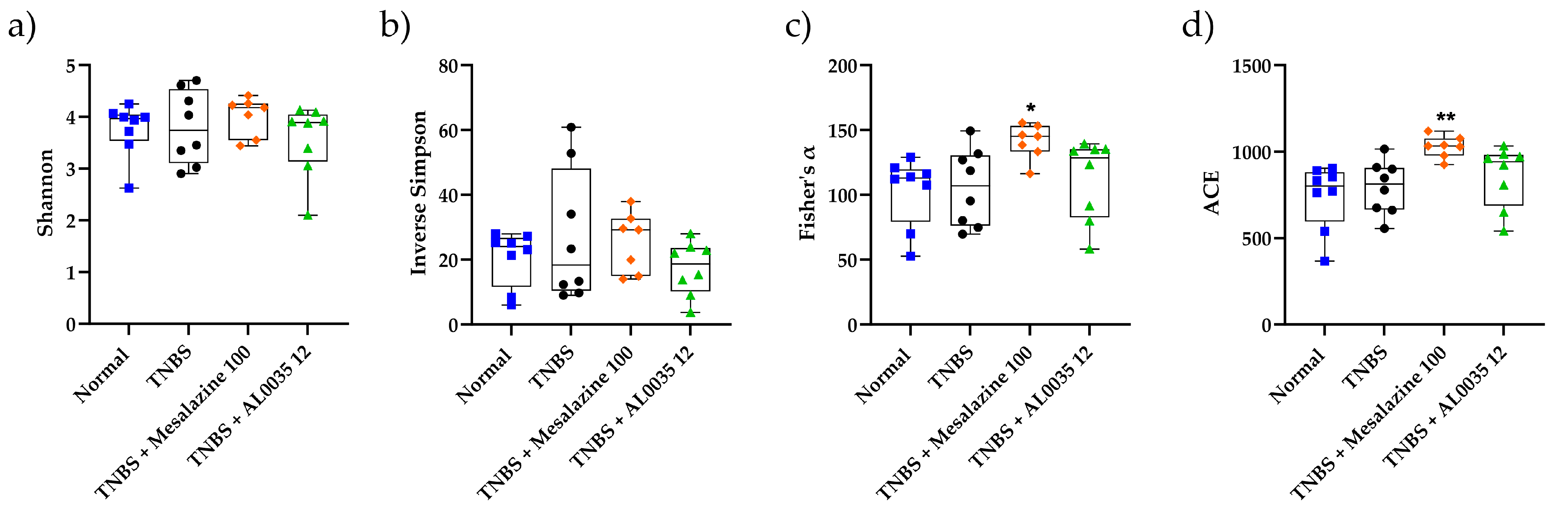
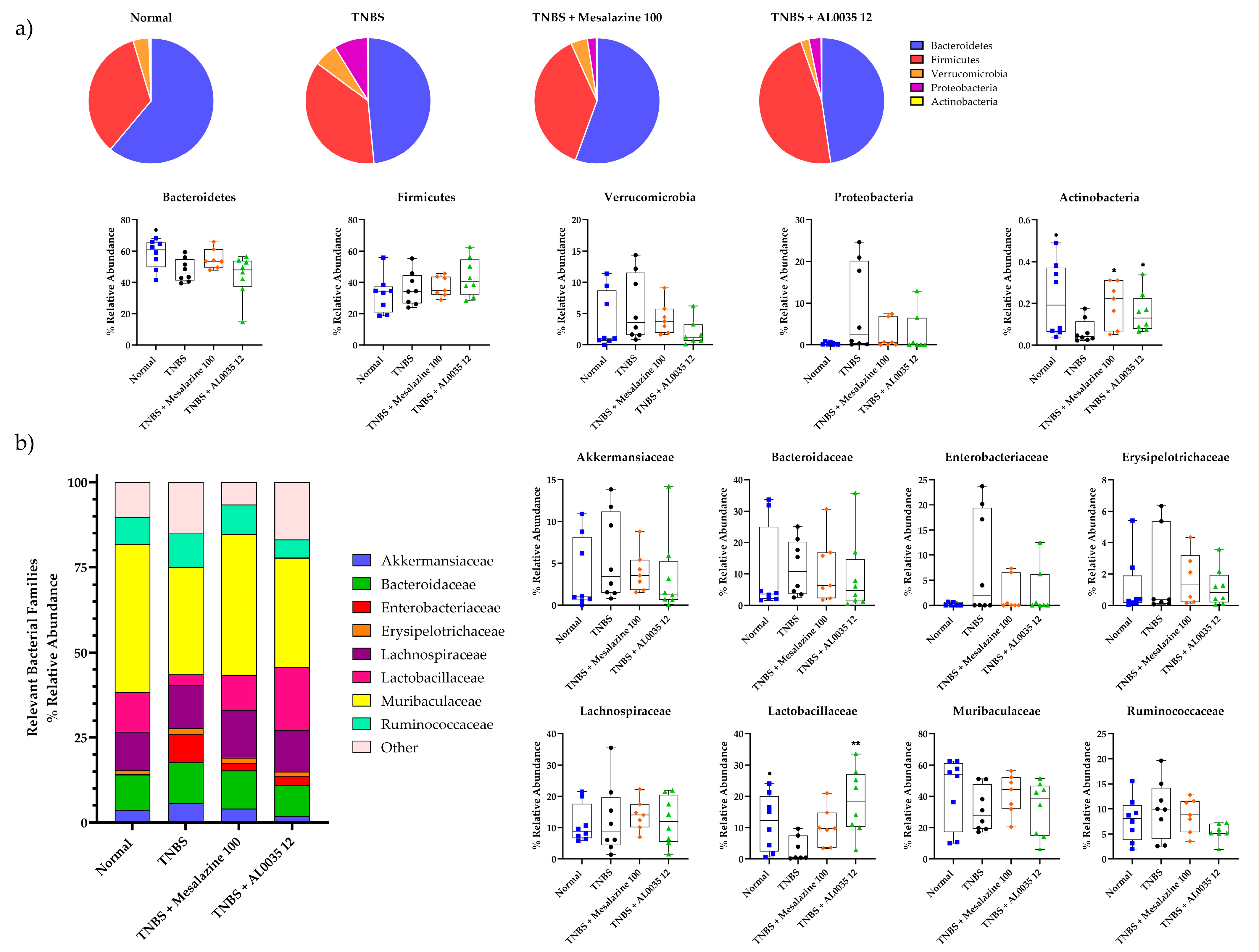
3.7. AL0035 Treatment Shows Eubiotic Properties on the Gut Microbiota of TNBS-Induced Colitis Mice
3.8. Effectiveness of AL0035 Treatment Reagrding Microbial Abundance at the Phylum and Family Levels in TNBS-Induced Colitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chon, H.; Choi, B.; Jeong, G.; Lee, E.; Lee, S. Suppression of proinflammatory cytokine production by specific metabolites of Lactobacillus plantarum 10hk2 via inhibiting NF-κB and p38 MAPK expressions. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, e41–e49. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.T.; Odenwald, M.A.; Turner, J.R.; Zuo, L. Tight junction proteins occludin and ZO-1 as regulators of epithelial proliferation and survival. Ann. N. Y. Acad. Sci. 2022, 1514, 21–33. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.; Guo, C.; Li, X. Chitosan Ameliorates DSS-Induced Ulcerative Colitis Mice by Enhancing Intestinal Barrier Function and Improving Microflora. Int. J. Mol. Sci. 2019, 20, 5751. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Q.; Zhang, Y.; Ma, W.; Ning, K.; Xiang, J.; Cui, J.; Xiang, H. Combination of probiotics with different functions alleviate DSS-induced colitis by regulating intestinal microbiota, IL-10, and barrier function. Appl. Microbiol. Biotechnol. 2020, 104, 335–349. [Google Scholar] [CrossRef]
- Triantafyllidi, A.; Xanthos, T.; Papalois, A.; Triantafillidis, J.K. Herbal and plant therapy in patients with inflammatory bowel disease. Ann. Gastroenterol. 2015, 28, 210–220. [Google Scholar] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Vilchez-Padial, L.M.; Gil, A. Evidence of the Anti-Inflammatory Effects of Probiotics and Synbiotics in Intestinal Chronic Diseases. Nutrients 2017, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Bron, P.A.; Kleerebezem, M.; Brummer, R.; Cani, P.D.; Mercenier, A.; MacDonald, T.T.; Wells, J.M. Can probiotics modulate human disease by impacting intestinal barrier function? Br. J. Nutr. 2017, 117, 93–107. [Google Scholar] [CrossRef]
- Jeong, J.; Lee, H.; Jang, S.; Han, M.J.; Kim, D. Lactobacillus plantarum C29 alleviates NF-κB activation and Th17/Treg imbalance in mice with TNBS-induced colitis. Food Agric. Immunol. 2018, 29, 577–589. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, H.; Jing, M.; Zhang, Z.; Zhang, G.; Sun, Y.; Shan, L.; Yu, P.; Wang, Y.; Xu, L. Andrographolide derivative AL-1 ameliorates TNBS-induced colitis in mice: Involvement of NF-κB and PPAR-γ signaling pathways. Sci. Rep. 2016, 2019, 29716. [Google Scholar] [CrossRef]
- Alex, P.; Zachos, N.C.; Nguyen, T.; Gonzales, L.; Chen, T.E.; Conklin, L.S.; Centola, M.; Li, X. Distinct cytokine patterns identified from multiplex profiles of murine DSS and TNBS-induced colitis. Inflamm. Bowel Dis. 2009, 15, 341–352. [Google Scholar] [CrossRef]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Morris, G.P.; Beck, P.L.; Herridge, M.S.; Depew, W.T.; Szewczuk, M.R.; Wallace, J.L. Hapten-induced model of chronic inflammation and ulceration in the rat colon. Gastroenterology 1989, 96, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Siegmund, B.; Fantuzzi, G.; Rieder, F.; Gamboni-Robertson, F.; Lehr, H.A.; Hartmann, G.; Dinarello, C.A.; Endres, S.; Eigler, A. Neutralization of interleukin-18 reduces severity in murine colitis and intestinal IFN-gamma and TNF-alpha production. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R1264–R1273. [Google Scholar] [CrossRef] [PubMed]
- Obermeier, F.; Kojouharoff, G.; Hans, W.; Scholmerich, J.; Gross, V.; Falk, W. Interferon-gamma (IFNgamma)- and tumour necrosis factor (TNF)-induced nitric oxide as toxic effector molecule in chronic dextran sulphate sodium (DSS)-induced colitis in mice. Clin. Exp. Immunol. 1999, 116, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.W.; Barrett, K.E. Mast cells are not essential to inflammation in murine model of colitis. Dig. Dis. Sci. 1994, 39, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.A.; et al. Improved Bacterial 16S rRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems 2015, 1, e00009-15. [Google Scholar] [CrossRef]
- Comeau, A.M.; Douglas, G.M.; Langille, M.G. Microbiome Helper: A Custom and Streamlined Workflow for Microbiome Research. mSystems. 2017, 2, e00127-16. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lei, J.; Hu, G.; Chen, X.; Liu, Z.; Yang, J. Matrine Mediates Inflammatory Response via Gut Microbiota in TNBS-Induced Murine Colitis. Front. Physiol. 2019, 10, 28. [Google Scholar] [CrossRef]
- Cota, D.; Mishra, S.; Shengule, S. Beneficial role of Terminalia arjuna hydroalcoholic extract in colitis and its possible mechanism. J. Ethnopharmacol. 2019, 230, 117–125. [Google Scholar] [CrossRef]
- Choi, K.; Cho, S.; Kook, S.; Chun, S.; Bhattarai, G.; Poudel, S.B.; Kim, M.K.; Lee, K.Y.; Lee, J.C. Intestinal anti-inflammatory activity of the seeds of Raphanus sativus L. in experimental ulcerative colitis models. J. Ethnopharmacol. 2016, 179, 55–65. [Google Scholar] [CrossRef]
- Biasi, F.; Astegiano, M.; Maina, M.; Leonarduzzi, G.; Poli, G. Polyphenol Supplementation as a Complementary Medicinal Approach to Treating Inflammatory Bowel Disease. Curr. Med. Chem. 2011, 18, 4851–4865. [Google Scholar] [CrossRef]
- Chi, J.H.; Kim, Y.H.; Sohn, D.H.; Seo, G.S.; Lee, S.H. Ameliorative effect of Alnus japonica ethanol extract on colitis through the inhibition of inflammatory responses and attenuation of intestinal barrier disruption in vivo and in vitro. Biomed. Pharmacother. 2018, 108, 1767–1774. [Google Scholar] [CrossRef]
- Song, L.; Xie, W.; Liu, Z.; Guo, D.; Zhao, D.; Qiao, X.; Wang, L.; Zhou, H.; Cui, W.; Jiang, Y.; et al. Oral delivery of a Lactococcus lactis strain secreting bovine lactoferricin–lactoferrampin alleviates the development of acute colitis in mice. Appl. Microbiol. Biotechnol. 2019, 103, 6169–6186. [Google Scholar] [CrossRef] [PubMed]
- Koeninger, L.; Armbruster, N.S.; Brinch, K.S.; Kjaerulf, S.; Andersen, B.; Langnau, C.; Autenrieth, S.E.; Schneidawind, D.; Stange, E.F.; Malek, N.P. Human β-Defensin 2 Mediated Immune Modulation as Treatment for Experimental Colitis. Front. Immunol. 2020, 11, 93. [Google Scholar] [CrossRef]
- Burrello, C.; Garavaglia, F.; Cribiu, F.M.; Ercoli, G.; Lopez, G.; Troisi, J.; Colucci, A.; Guglietta, S.; Carloni, S.; Guglielmetti, S.; et al. Therapeutic faecal microbiota transplantation controls intestinal inflammation through IL10 secretion by immune cells. Nat. Commun. 2018, 9, 5184. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Liu, M.; Li, Y.; Yu, H.; Wang, D.; Chen, Q.; Chen, Y.; Zhang, Y.; Wang, T. Flavonoids from Citrus aurantium ameliorate TNBS-induced ulcerative colitis through protecting colonic mucus layer integrity. Eur. J. Pharmacol. 2019, 857, 172456. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, Y.; Wan, P.; Chen, D.; Ding, Y.; Ran, L.; Mi, J.; Lu, L.; Zhang, Z.J.; Li, Z.; et al. Gut microbiota modulation and anti-inflammatory properties of anthocyanins from the fruits of Lycium ruthenicum Murray in dextran sodium sulfate-induced colitis in mice. Free. Radic. Biol. Med. 2019, 136, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Choi, H.; Kim, D. The mixture of Anemarrhena asphodeloides and Coptis chinensis attenuates high-fat diet-induced colitis in mice. Am. J. Chin. Med. 2017, 45, 1033–1046. [Google Scholar] [CrossRef]
- Tannock, G.W. Molecular methods for exploring the intestinal ecosystem. Br. J. Nutr. 2002, 87, 199–201. [Google Scholar] [CrossRef]
- Zhu, Y.; Xu, Y.; Wang, X.; Rao, L.; Yan, X.; Gao, R.; Shen, T.; Zhou, Y.; Kong, C.; Zhou, L. Probiotic Cocktail Alleviates Intestinal Inflammation Through Improving Gut Microbiota and Metabolites in Colitis Mice. Front. Cell Infect. Microbiol. 2022, 12, 886061. [Google Scholar] [CrossRef]
- González-Lozano, E.; García-García, J.; Gálvez, J.; Hidalgo-García, L.; Rodríguez-Nogales, A.; Rodríguez-Cabezas, M.E.; Sánchez, M. Novel Horizons in Postbiotics: Lactobacillaceae Extracellular Vesicles and Their Applications in Health and Disease. Nutrients 2022, 14, 5296. [Google Scholar] [CrossRef] [PubMed]
- Štofilová, J.; Kvaková, M.; Kamlárová, A.; Hijová, E.; Bertková, I.; Guľašová, Z. Probiotic-Based Intervention in the Treatment of Ulcerative Colitis: Conventional and New Approaches. Biomedicines 2022, 10, 2236. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Zhang, X.; Tong, L.; Liu, Q.; Liang, X.; Bu, Y.; Gong, P.; Liu, T.; Zhang, L.; Xia, Y.; et al. Effect of Extracellular Vesicles Derived From Lactobacillus plantarum Q7 on Gut Microbiota and Ulcerative Colitis in Mice. Front. Immunol. 2021, 12, 777147. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DAI Index | Weight Loss (%) | Stool Consistency | Occult/Gross Bleeding |
|---|---|---|---|
| 0 | None | Normal | No bleeding |
| 1 | 1–5 | ||
| 2 | 5–10 | Loose stools | Hemoccult-positive (slight bleeding) |
| 3 | 10–15 | ||
| 4 | >15 | Diarrhea | Gross bleeding |
| Colon Damage | Score |
|---|---|
| No damage | 0 |
| Hyperemia without ulcers | 1 |
| Hyperemia and wall thickening without ulcers | 2 |
| One ulceration site without wall thickening | 3 |
| Two or more ulceration sites | 4 |
| 0.5 cm extension of inflammation or major damage | 5 |
| 1 cm extension of inflammation or severe damage | 6–10 (the score was increased by 1 for every 0.5 cm of damage up to a maximal score of 10) |
| Infiltration of Inflammatory Cells | Score |
| Presence of occasional inflammatory cells in the lamina propria | 0 |
| Increased numbers of inflammatory cells in the lamina propria | 1 |
| Confluence of inflammatory cells, extending into the submucosa | 2 |
| Transmural extension of the infiltrate | 3 |
| Tissue damage | Score |
| No mucosal damage | 0 |
| Discrete lymphoepithelial lesions | 1 |
| Surface mucosal erosion or focal ulceration | 2 |
| Extensive mucosal damage and extension into deeper structures of the bowel wall | 3 |
| Gene Name | Sequence |
|---|---|
| FoxP3 | AGACCCCTGTGCTCCAAGTG CAGACTCCATTTGCCAGCAG |
| GATA3 | GAACCGCCCCTTATCAAG CAGGATGTCCCTGCTCTCCTT |
| NK1.1 | TCCCTTCTCACCACCAGTTA CAGTCTTGTGGGCACTCTAA |
| T-bet | AATCGACAACAACCCCTTTG AACTGTGTTCCCGAGGTGTC |
| IL-10 | ATTTGAATTCCCTGGGTGAGAAG CACAGGGGAGAAATCGATGAC |
| IL-6 | AGGATACCACTCCCAACAGACCT CAAGTGCATCATCGTTGTTCATAC |
| MCP-1 | TGATCCCAATGAGTAGGCTGGAG ATGTCTGGACCCATTCCTTCTTG |
| INF-γ | CAATGAACGCTACACACTGC CCACATCTATGCCACTTGAG |
| TNF-α | CCCCAAAGGGATGAGAAGTTC TGAGGGTCTGGGCCATAGAA |
| IL-1β | TCAGGCAGGCAGTATCACTC CTAATGGGAACGTCACACC |
| IL-12 | CAGAAGCTAACCATCTCCTGGTTTG TCCGGAGTAATTTGGTGCTTCACAC |
| GAPDH | AACTTTGGCATTGTGGAAGG CACATTGGGGGTAGGAACAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vesci, L.; Tundo, G.; Soldi, S.; Galletti, S.; Stoppoloni, D.; Bernardini, R.; Modolea, A.B.; Luberto, L.; Marra, E.; Giorgi, F.; et al. A Novel Lactobacillus brevis Fermented with a Vegetable Substrate (AL0035) Counteracts TNBS-Induced Colitis by Modulating the Gut Microbiota Composition and Intestinal Barrier. Nutrients 2024, 16, 937. https://doi.org/10.3390/nu16070937
Vesci L, Tundo G, Soldi S, Galletti S, Stoppoloni D, Bernardini R, Modolea AB, Luberto L, Marra E, Giorgi F, et al. A Novel Lactobacillus brevis Fermented with a Vegetable Substrate (AL0035) Counteracts TNBS-Induced Colitis by Modulating the Gut Microbiota Composition and Intestinal Barrier. Nutrients. 2024; 16(7):937. https://doi.org/10.3390/nu16070937
Chicago/Turabian StyleVesci, Loredana, Grazia Tundo, Sara Soldi, Serena Galletti, Daniela Stoppoloni, Roberta Bernardini, Anamaria Bianca Modolea, Laura Luberto, Emanuele Marra, Fabrizio Giorgi, and et al. 2024. "A Novel Lactobacillus brevis Fermented with a Vegetable Substrate (AL0035) Counteracts TNBS-Induced Colitis by Modulating the Gut Microbiota Composition and Intestinal Barrier" Nutrients 16, no. 7: 937. https://doi.org/10.3390/nu16070937
APA StyleVesci, L., Tundo, G., Soldi, S., Galletti, S., Stoppoloni, D., Bernardini, R., Modolea, A. B., Luberto, L., Marra, E., Giorgi, F., & Marini, S. (2024). A Novel Lactobacillus brevis Fermented with a Vegetable Substrate (AL0035) Counteracts TNBS-Induced Colitis by Modulating the Gut Microbiota Composition and Intestinal Barrier. Nutrients, 16(7), 937. https://doi.org/10.3390/nu16070937