
Hydrophobic Components in Light-Yellow Pulp Sweet Potato (Ipomoea batatas (L.) Lam.) Tubers Suppress LPS-Induced Inflammatory Responses in RAW264.7 Cells via Activation of the Nrf2 Pathway

Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Sweet Potato Samples and Extracts
2.2. Reagents and Antibodies
2.3. Cell Culture and Stimulation Conditions
2.4. Cell Viability Assay
2.5. Enzyme-Linked Immunosorben Assay (ELISA)
2.6. Intracellular ROS Observation
2.7. RT-qPCR
2.8. Western Blotting
2.9. High-Performance Liquid Chromatography (HPLC)
2.10. DPPH Radical Scavenging Activity Assay
2.11. Total Polyphenol Content Assay
2.12. Total Flavonoid Content Assay
2.13. Statistical Analysis
3. Results
3.1. Sweet Potato Tuber Extracts Suppressed the Production of Inflammatory Cytokines and Gene Expression
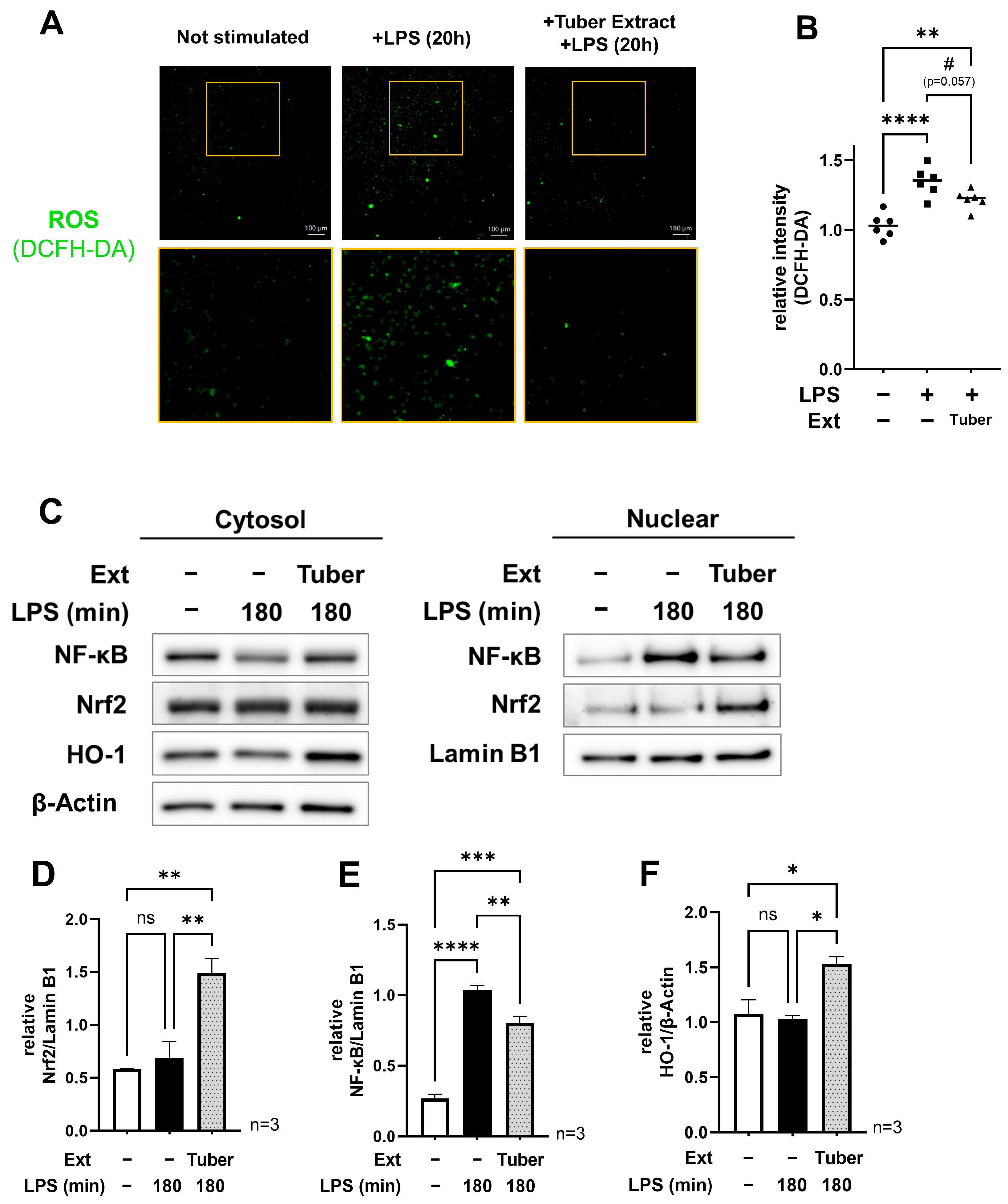
3.2. Tuber Extracts Induce Activation of the Nrf2 Oxidative Stress Response Pathway and Inhibit the Function of NF-κB
3.3. Nrf2 Inhibition Cancels the Anti-Inflammatory Effect of the Tuber Extracts
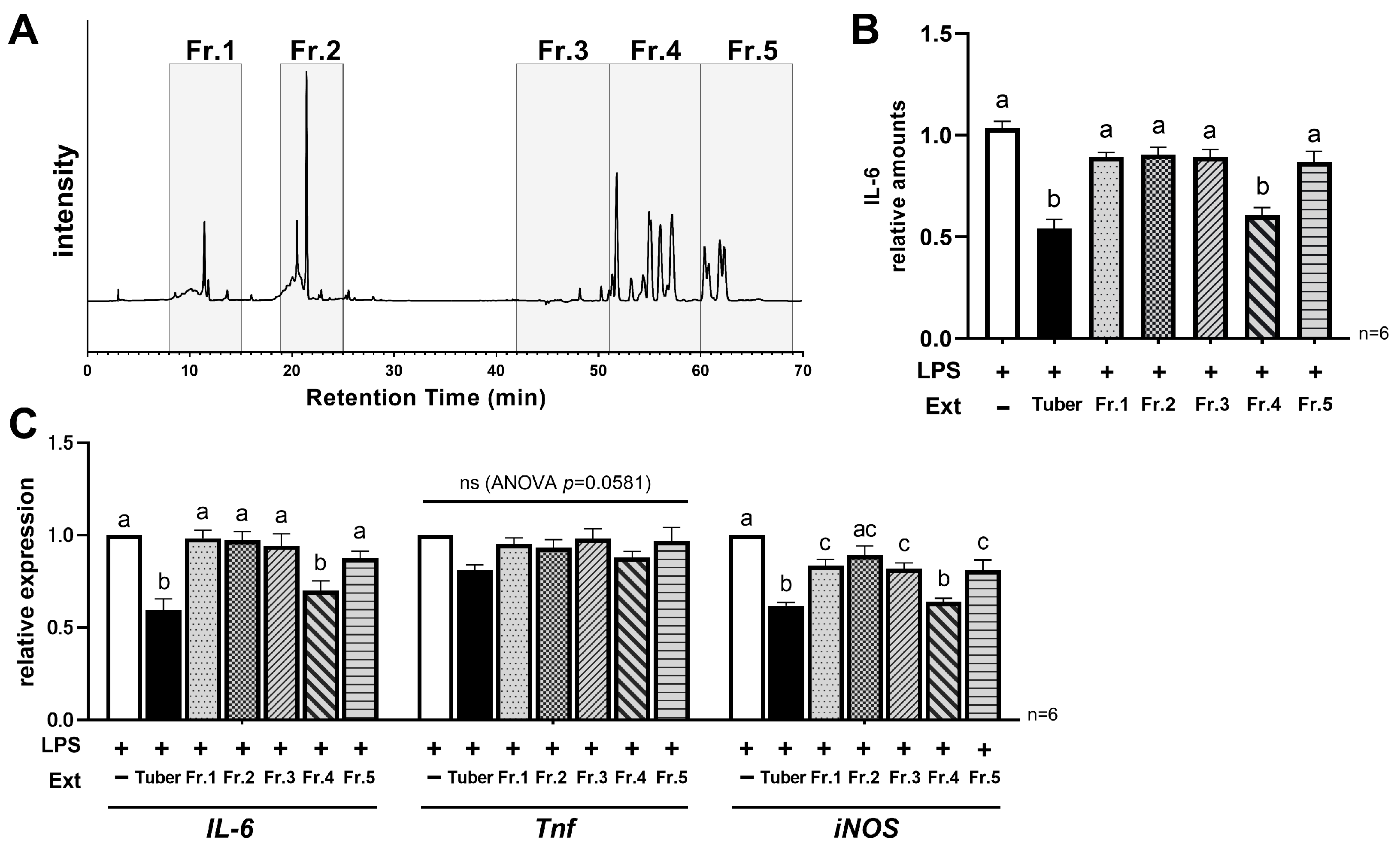
3.4. Hydrophobic Components in the Tuber Contribute to Its Anti-Inflammatory Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. S1), S4–S9. [Google Scholar] [CrossRef]
- Boutens, L.; Hooiveld, G.J.; Dhingra, S.; Cramer, R.A.; Netea, M.G.; Stienstra, R. Unique metabolic activation of adipose tissue macrophages in obesity promotes inflammatory responses. Diabetologia 2018, 61, 942–953. [Google Scholar] [CrossRef]
- Giallongo, C.; Tibullo, D.; Camiolo, G.; Parrinello, N.L.; Romano, A.; Puglisi, F.; Barbato, A.; Conticello, C.; Lupo, G.; Anfuso, C.D.; et al. TLR4 signaling drives mesenchymal stromal cells commitment to promote tumor microenvironment transformation in multiple myeloma. Cell Death. Dis. 2019, 10, 704. [Google Scholar] [CrossRef]
- Knekt, P.; Kumpulainen, J.; Jarvinen, R.; Rissanen, H.; Heliovaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food. Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Na, H.K.; Surh, Y.J. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem. Toxicol. 2008, 46, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, L.F.; Pedruzzi, L.M.; Stenvinkel, P.; Stockler-Pinto, M.B.; Daleprane, J.B.; Leite, M., Jr.; Mafra, D. Nutritional strategies to modulate inflammation and oxidative stress pathways via activation of the master antioxidant switch Nrf2. Biochimie 2013, 95, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi-Tago, M.; Nonaka, Y.; Tago, K.; Takeda, M.; Ishihara, Y.; Sakai, A.; Matsutaka, M.; Kobata, K.; Tamura, H. Pyrocatechol, a component of coffee, suppresses LPS-induced inflammatory responses by inhibiting NF-kappaB and activating Nrf2. Sci. Rep. 2020, 10, 2584. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell. Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Willis, D.; Moore, A.R.; Frederick, R.; Willoughby, D.A. Heme oxygenase: A novel target for the modulation of the inflammatory response. Nat. Med. 1996, 2, 87–90. [Google Scholar] [CrossRef]
- Liu, G.H.; Qu, J.; Shen, X. NF-kappaB/p65 antagonizes Nrf2-ARE pathway by depriving CBP from Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochim. Biophys. Acta 2008, 1783, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef] [PubMed]
- Oin, Y.; Naumovski, N.; Ranasheera, C.S.; D’Cunha, N.M. Nutrition-related health outcomes of sweet potato (Ipomoea batatas) consumption: A systematic review. Food Biosci. 2022, 50, 102208. [Google Scholar] [CrossRef]
- Islam, S.N.; Nusrat, T.; Begum, P.; Ahsan, M. Carotenoids and beta-carotene in orange fleshed sweet potato: A possible solution to vitamin A deficiency. Food Chem. 2016, 199, 628–631. [Google Scholar] [CrossRef]
- Chen, C.C.; Lin, C.; Chen, M.H.; Chiang, P.Y. Stability and Quality of Anthocyanin in Purple Sweet Potato Extracts. Foods 2019, 8, 393. [Google Scholar] [CrossRef] [PubMed]
- Sugata, M.; Lin, C.Y.; Shih, Y.C. Anti-Inflammatory and Anticancer Activities of Taiwanese Purple-Fleshed Sweet Potatoes (Ipomoea batatas L. Lam) Extracts. Biomed Res. Int. 2015, 2015, 768093. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.Y.; Park, W.S.; Kim, H.J.; Kim, H.S.; Kang, K.K.; Kwak, S.S.; Ahn, M.J. Protective Effect of Carotenoid Extract from Orange-Fleshed Sweet Potato on Gastric Ulcer in Mice by Inhibition of NO, IL-6 and PGE(2) Production. Pharmaceuticals 2021, 14, 1320. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Zhou, J.; Liu, W.; Tao, W.; He, J.; Jin, W.; Guo, H.; Yang, N.; Li, Y. The anti-inflammatory potential of protein-bound anthocyanin compounds from purple sweet potato in LPS-induced RAW264.7 macrophages. Food Res. Int. 2020, 137, 109647. [Google Scholar] [CrossRef]
- Singh, A.; Venkannagari, S.; Oh, K.H.; Zhang, Y.Q.; Rohde, J.M.; Liu, L.; Nimmagadda, S.; Sudini, K.; Brimacombe, K.R.; Gajghate, S.; et al. Small Molecule Inhibitor of NRF2 Selectively Intervenes Therapeutic Resistance in KEAP1-Deficient NSCLC Tumors. ACS Chem. Biol. 2016, 11, 3214–3225. [Google Scholar] [CrossRef]
- Makori, S.; Mu, T.-H.; Sun, H.-N. Total Polyphenol Content, Antioxidant Activity, and Individual Phenolic Composition of Different Edible Parts of 4 Sweet Potato Cultivars. Nat. Prod. Commun. 2020, 15. [Google Scholar] [CrossRef]
- Ishiguro, K.; Yoshinaga, M.; Kai, Y.; Maoka, T.; Yoshimoto, M. Composition, content and antioxidative activity of the carotenoids in yellow-fleshed sweetpotato (Ipomoea batatas L.). Breed. Sci. 2010, 60, 324–329. [Google Scholar] [CrossRef]
- Wang, A.; Li, R.; Ren, L.; Gao, X.; Zhang, Y.; Ma, Z.; Ma, D.; Luo, Y. A comparative metabolomics study of flavonoids in sweet potato with different flesh colors (Ipomoea batatas (L.) Lam). Food Chem. 2018, 260, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Nagane, M.; Aihara, N.; Kamiie, J.; Miyanabe, M.; Hiraki, S.; Luo, X.; Nakanishi, I.; Shoji, Y.; Matsumoto, K.I.; et al. Lipid-soluble polyphenols from sweet potato exert antitumor activity and enhance chemosensitivity in breast cancer. J. Clin. Biochem. Nutr. 2021, 68, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Kitajima, M.; Nishida, K.; Serada, S.; Fujimoto, M.; Naka, T.; Fujii-Kuriyama, Y.; Sakamato, S.; Ito, T.; Handa, H.; et al. NQO1 inhibits the TLR-dependent production of selective cytokines by promoting IkappaB-zeta degradation. J. Exp. Med. 2018, 215, 2197–2209. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, F.; Lu, S.; Ren, L.; Bian, S.; Liu, M.; Zhao, D.; Wang, S.; Wang, J. Ginseng root extract attenuates inflammation by inhibiting the MAPK/NF-kappaB signaling pathway and activating autophagy and p62-Nrf2-Keap1 signaling in vitro and in vivo. J. Ethnopharmacol. 2022, 283, 114739. [Google Scholar] [CrossRef]
- Kim, H.N.; Park, G.H.; Park, S.B.; Kim, J.D.; Eo, H.J.; Son, H.J.; Song, J.H.; Jeong, J.B. Sageretia thea Inhibits Inflammation through Suppression of NF- kappa B and MAPK and Activation of Nrf2/HO-1 Signaling Pathways in RAW264.7 Cells. Am. J. Chin. Med. 2019, 47, 385–403. [Google Scholar] [CrossRef]
- Park, E.J.; Kim, Y.M.; Park, S.W.; Kim, H.J.; Lee, J.H.; Lee, D.U.; Chang, K.C. Induction of HO-1 through p38 MAPK/Nrf2 signaling pathway by ethanol extract of Inula helenium L. reduces inflammation in LPS-activated RAW 264.7 cells and CLP-induced septic mice. Food Chem. Toxicol. 2013, 55, 386–395. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | |
|---|---|---|
| IL-6 | Forward: | AGAGGATACCACTCCCAACAGA |
| Reverse: | CTGCAAGTGCATCATCGTTGTTC | |
| Tnf | Forward: | ATTCGAGTGACAAGCCTGTAG |
| Reverse: | TGAAGAGAACCTGGGAGTAGAC | |
| iNOS (Nos2) | Forward: | AGGCTGGAAGCTGTAACAAAGG |
| Reverse: | GCTGAAACATTTCCTGTGCTGTG | |
| Actb | Forward: | ACTATTGGCAACGAGCGGTTC |
| Reverse: | TCAGCAATGCCTGGGTACATG | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, Y.; Suto, M.; Umebara, I.; Masutomi, H.; Ishihara, K. Hydrophobic Components in Light-Yellow Pulp Sweet Potato (Ipomoea batatas (L.) Lam.) Tubers Suppress LPS-Induced Inflammatory Responses in RAW264.7 Cells via Activation of the Nrf2 Pathway. Nutrients 2024, 16, 563. https://doi.org/10.3390/nu16040563
Matsumoto Y, Suto M, Umebara I, Masutomi H, Ishihara K. Hydrophobic Components in Light-Yellow Pulp Sweet Potato (Ipomoea batatas (L.) Lam.) Tubers Suppress LPS-Induced Inflammatory Responses in RAW264.7 Cells via Activation of the Nrf2 Pathway. Nutrients. 2024; 16(4):563. https://doi.org/10.3390/nu16040563
Chicago/Turabian StyleMatsumoto, Yuma, Mari Suto, Io Umebara, Hirofumi Masutomi, and Katsuyuki Ishihara. 2024. "Hydrophobic Components in Light-Yellow Pulp Sweet Potato (Ipomoea batatas (L.) Lam.) Tubers Suppress LPS-Induced Inflammatory Responses in RAW264.7 Cells via Activation of the Nrf2 Pathway" Nutrients 16, no. 4: 563. https://doi.org/10.3390/nu16040563
APA StyleMatsumoto, Y., Suto, M., Umebara, I., Masutomi, H., & Ishihara, K. (2024). Hydrophobic Components in Light-Yellow Pulp Sweet Potato (Ipomoea batatas (L.) Lam.) Tubers Suppress LPS-Induced Inflammatory Responses in RAW264.7 Cells via Activation of the Nrf2 Pathway. Nutrients, 16(4), 563. https://doi.org/10.3390/nu16040563