
Profiles of Serum Fatty Acids in Healthy Women on Different Types of Vegetarian Diets


 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Types and Anthropometric Data
- -
- Body mass (Jawon Medical X-contact 350, Jawon, Seongnam, Republic of Korea) and height (stadiometer Seca 213, Germany), used to calculate the body mass index (BMI) value;
- -
- The circumference of the waist and hips (tape measure Seca 201, Seca, Hamburg, Germany), used to calculate the waist-to-hip ratio (WHR);
- -
- Body composition (body fat content), estimated with the BIA method (Jawon Medical X-contact 350, Jawon, Seongnam, Republic of Korea).
2.2. Sample Collection and Lipid Analysis
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Djuricic, I.; Calder, P.C. Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef]
- Dąbrowski, G.; Konopka, I. Update on Food Sources and Biological Activity of Odd-Chain, Branched and Cyclic Fatty Acids––A Review. Trends Food Sci. Technol. 2022, 119, 514–529. [Google Scholar] [CrossRef]
- Czumaj, A.; Śledziński, T.; Mika, A. Branched-Chain Fatty Acids Alter the Expression of Genes Responsible for Lipid Synthesis and Inflammation in Human Adipose Cells. Nutrients 2022, 14, 2310. [Google Scholar] [CrossRef]
- Lee, H.Y.; Woo, J.; Chen, Z.Y.; Leung, S.F.; Peng, X.H. Serum Fatty Acid, Lipid Profile and Dietary Intake of Hong Kong Chinese Omnivores and Vegetarians. Eur. J. Clin. Nutr. 2000, 54, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Elorinne, A.L.; Alfthan, G.; Erlund, I.; Kivimäki, H.; Paju, A.; Salminen, I.; Turpeinen, U.; Voutilainen, S.; Laakso, J. Food and Nutrient Intake and Nutritional Status of Finnish Vegans and Non-Vegetarians. PLoS ONE 2016, 11, e0148235. [Google Scholar] [CrossRef]
- Espinosa-Marrón, A.; Laviada-Molina, H.; Moreno-Enríquez, A.; Sosa-Crespo, I.F.; Molina-Segui, F.; Fernanda Villaseñor-Espinosa, M.; Antonio Ciau-Mendoza, J.; Alfredo Araujo-León, J. Serum Fatty Acids Chemical Characterization after Prolonged Exposure to a Vegan Diet. J. Food Nutr. Res. 2019, 7, 742–750. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Dybvik, J.S.; Svendsen, M.; Aune, D. Vegetarian and Vegan Diets and the Risk of Cardiovascular Disease, Ischemic Heart Disease and Stroke: A Systematic Review and Meta-Analysis of Prospective Cohort Studies. Eur. J. Nutr. 2023, 62, 51–69. [Google Scholar] [CrossRef]
- Koutentakis, M.; Surma, S.; Rogula, S.; Filipiak, K.J.; Gąsecka, A. The Effect of a Vegan Diet on the Cardiovascular System. J. Cardiovasc. Dev. Dis. 2023, 10, 94. [Google Scholar] [CrossRef]
- Perna, M.; Hewlings, S. Saturated Fatty Acid Chain Length and Risk of Cardiovascular Disease: A Systematic Review. Nutrients 2022, 15, 30. [Google Scholar] [CrossRef]
- Oppedisano, F.; Macrì, R.; Gliozzi, M.; Musolino, V.; Carresi, C.; Maiuolo, J.; Bosco, F.; Nucera, S.; Zito, M.C.; Guarnieri, L.; et al. The Anti-Inflammatory and Antioxidant Properties of n-3 PUFAs: Their Role in Cardiovascular Protection. Biomedicines 2020, 8, 306. [Google Scholar] [CrossRef]
- Harris, W.S. Achieving Optimal N-3 Fatty Acid Status: The Vegetarian’s Challenge… or Not. Am. J. Clin. Nutr. 2014, 100 (Suppl. 1), 449S–452S. [Google Scholar] [CrossRef]
- Rosell, M.S.; Lloyd-Wright, Z.; Appleby, P.N.; Sanders, T.A.; Allen, N.E.; Key, T.J. Long-Chain n-3 Polyunsaturated Fatty Acids in Plasma in British Meat-Eating, Vegetarian, and Vegan Men. Am. J. Clin. Nutr. 2005, 82, 327–334. [Google Scholar] [CrossRef]
- Sarter, B.; Kelsey, K.S.; Schwartz, T.A.; Harris, W.S. Blood Docosahexaenoic Acid and Eicosapentaenoic Acid in Vegans: Associations with Age and Gender and Effects of an Algal-Derived Omega-3 Fatty Acid Supplement. Clin. Nutr. 2015, 34, 212–218. [Google Scholar] [CrossRef]
- Dighriri, I.M.; Alsubaie, A.M.; Hakami, F.M.; Hamithi, D.M.; Alshekh, M.M.; Khobrani, F.A.; Dalak, F.E.; Hakami, A.A.; Alsueaadi, E.H.; Alsaawi, L.S.; et al. Effects of Omega-3 Polyunsaturated Fatty Acids on Brain Functions: A Systematic Review. Cureus 2022, 14, e30091. [Google Scholar] [CrossRef]
- Khalid, W.; Gill, P.; Arshad, M.S.; Ali, A.; Ranjha, M.M.A.N.; Mukhtar, S.; Afzal, F.; Maqbool, Z. Functional Behavior of DHA and EPA in the Formation of Babies Brain at Different Stages of Age, and Protect from Different Brain-Related Diseases. Int. J. Food Prop. 2022, 25, 1021–1044. [Google Scholar] [CrossRef]
- Jarosz, J.C.M.; Rychlik, E.; Stoś, K.; Charzewska, J. Normy Żywienia Dla Populacji Polski i Ich Zastosowanie; Narodowy Instytut Zdrowia Publicznego—Państwowy Zakład Higieny: Warszawa, Poland, 2020; Available online: https://www.bing.com/search?q=J.+C.+M.+Jarosz%2C+E.+Rychlik%2C+K.+Sto%C5%9B%2C+J.+Charzewska+%E2%80%9CNormy+%C5%BCywienia+dla+populacji+Polski+i+ich+zastosowanie%2C%E2%80%9D+Narodowy+Instytut+Zdrowia+Publicznego+%E2%80%93+Pa%C5%84stwowy+Zak%C5%82ad+Higieny%2C+Warszawa%2C+2020&cvid=295ea4dcbec84d69b16a42ccc6868c6e&gs_lcrp=EgZjaHJvbWUyBggAEEUYOdIBBzc1MWowajmoAgCwAgA&FORM=ANAB01&PC=LCTS (accessed on 1 December 2023).
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, Interconversion, and Dose Response of n-3 Fatty Acids in Humans. Am. J. Clin. Nutr. 2006, 83, 1467S–1476S. [Google Scholar] [CrossRef]
- Brenna, J.T.; Salem, N.; Sinclair, A.J.; Cunnane, S.C. Alpha-Linolenic Acid Supplementation and Conversion to n-3 Long-Chain Polyunsaturated Fatty Acids in Humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Fokkema, M.R.; Brouwer, D.A.J.; Hasperhoven, M.B.; Martini, I.A.; Muskiet, F.A.J. Short-Term Supplementation of Low-Dose Gamma-Linolenic Acid (GLA), Alpha-Linolenic Acid (ALA), or GLA plus ALA Does Not Augment LCP Omega 3 Status of Dutch Vegans to an Appreciable Extent. Prostaglandins Leukot. Essent. Fat. Acids 2000, 63, 287–292. [Google Scholar] [CrossRef] [PubMed]
- García-Maldonado, E.; Alcorta, A.; Zapatera, B.; Vaquero, M.P. Changes in Fatty Acid Levels after Consumption of a Novel Docosahexaenoic Supplement from Algae: A Crossover Randomized Controlled Trial in Omnivorous, Lacto-Ovo Vegetarians and Vegans. Eur. J. Nutr. 2023, 62, 1691–1705. [Google Scholar] [CrossRef]
- Arshad, Z.; Rezapour-Firouzi, S.; Ebrahimifar, M.; Jarrahi, A.M.; Mohammadian, M. Association of Delta-6-Desaturase Expression with Aggressiveness of Cancer, Diabetes Mellitus, and Multiple Sclerosis: A Narrative Review. Asian Pac. J. Cancer Prev. 2019, 20, 1005–1018. [Google Scholar] [CrossRef]
- Gonzalez-Soto, M.; Mutch, D.M. Diet Regulation of Long-Chain PUFA Synthesis: Role of Macronutrients, Micronutrients, and Polyphenols on Δ-5/Δ-6 Desaturases and Elongases 2/5. Adv. Nutr. 2021, 12, 980–994. [Google Scholar] [CrossRef]
- Gonzalez-Soto, M.; Abdelmagid, S.A.; Ma, D.W.L.; El-Sohemy, A.; Mutch, D.M. Soy Consumption, but Not Dairy Consumption, Is Inversely Associated with Fatty Acid Desaturase Activity in Young Adults. Nutrients 2021, 13, 2817. [Google Scholar] [CrossRef]
- Fokkema, M.R.; Brouwer, D.A.J.; Hasperhoven, M.B.; Hettema, Y.; Bemelmans, W.J.E.; Muskiet, F.A.J. Polyunsaturated Fatty Acid Status of Dutch Vegans and Omnivores. Prostaglandins Leukot. Essent. Fat. Acids 2000, 63, 279–285. [Google Scholar] [CrossRef]
- Miles, F.L.; Lloren, J.I.C.; Haddad, E.; Jaceldo-Siegl, K.; Knutsen, S.; Sabate, J.; Fraser, G.E. Plasma, Urine, and Adipose Tissue Biomarkers of Dietary Intake Differ Between Vegetarian and Non-Vegetarian Diet Groups in the Adventist Health Study-2. J. Nutr. 2019, 149, 667–675. [Google Scholar] [CrossRef]
- Burns-Whitmore, B.; Froyen, E.; Heskey, C.; Parker, T.; Pablo, G.S. Alpha-Linolenic and Linoleic Fatty Acids in the Vegan Diet: Do They Require Dietary Reference Intake/Adequate Intake Special Consideration? Nutrients 2019, 11, 2365. [Google Scholar] [CrossRef] [PubMed]
- Pilorgé, E. Sunflower in the Global Vegetable Oil System: Situation, Specificities and Perspectives. OCL 2020, 27, 34. [Google Scholar] [CrossRef]
- Mann, N.; Pirotta, Y.; O’Connell, S.; Li, D.; Kelly, F.; Sinclair, A. Fatty Acid Composition of Habitual Omnivore and Vegetarian Diets. Lipids 2006, 41, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Dawczynski, C.; Weidauer, T.; Richert, C.; Schlattmann, P.; Dawczynski, K.; Kiehntopf, M. Nutrient Intake and Nutrition Status in Vegetarians and Vegans in Comparison to Omnivores—The Nutritional Evaluation (NuEva) Study. Front. Nutr. 2022, 9, 819106. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Magkos, F.; Zhou, D.; Eagon, J.C.; Fabbrini, E.; Okunade, A.L.; Klein, S. Adipose Tissue Monomethyl Branched-Chain Fatty Acids and Insulin Sensitivity: Effects of Obesity and Weight Loss. Obesity 2015, 23, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Mika, A.; Stepnowski, P.; Kaska, L.; Proczko, M.; Wisniewski, P.; Sledzinski, M.; Sledzinski, T. A Comprehensive Study of Serum Odd- and Branched-Chain Fatty Acids in Patients with Excess Weight. Obesity 2016, 24, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Ballantyne, C.M.; Nambi, V. Markers of Inflammation and Their Clinical Significance. Atheroscler. Suppl. 2005, 6, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Menzel, J.; Jabakhanji, A.; Biemann, R.; Mai, K.; Abraham, K.; Weikert, C. Systematic Review and Meta-Analysis of the Associations of Vegan and Vegetarian Diets with Inflammatory Biomarkers. Sci. Rep. 2020, 10, 21736. [Google Scholar] [CrossRef] [PubMed]
- Jenko Pražnikar, Z.; Šik Novak, K.; Bogataj Jontez, N.; Petelin, A.; Mohorko, N.; Kenig, S. Inflammatory and Intestinal Permeability Biomarkers in Healthy Participants on Long Term Vegan, Vegetarian, Omnivore and Low-Carbohydrate High-Fat Diet. Sci. Rep. 2023, 13, 17286. [Google Scholar] [CrossRef]
- Řezanka, T.; Nedbalová, L.; Sigler, K. Odd-Numbered Very-Long-Chain Polyunsaturated Fatty Acids from the Dinoflagellate Amphidinium Carterae Identified by Atmospheric Pressure Chemical Ionization Liquid Chromatography-Mass Spectrometry. Phytochemistry 2008, 69, 2849–2855. [Google Scholar] [CrossRef]
- Gogga, P.; Śliwińska, A.; Aleksandrowicz-Wrona, E.; Małgorzewicz, S. Lipid Profile in Polish Women Following Lacto-Ovo-Vegetarian and Vegan Diets—Preliminary Study. Acta Biochim. Pol. 2021, 68, 751–755. [Google Scholar] [CrossRef]
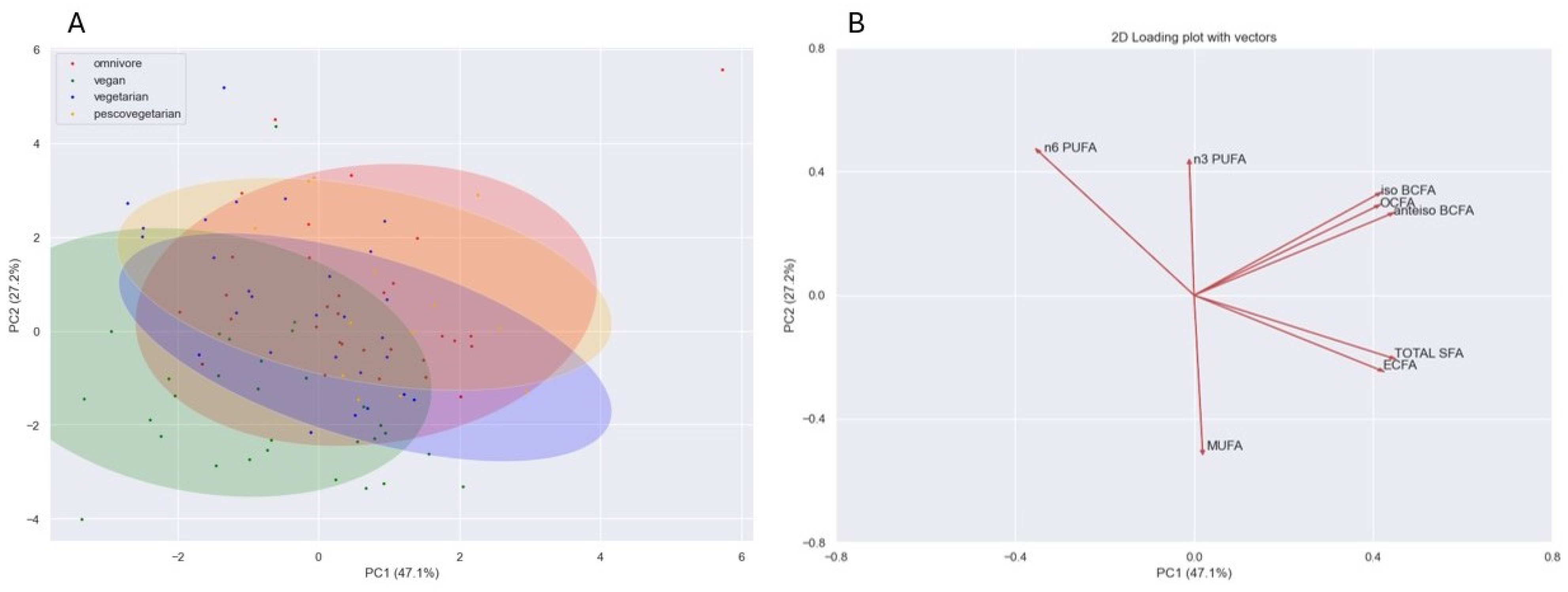
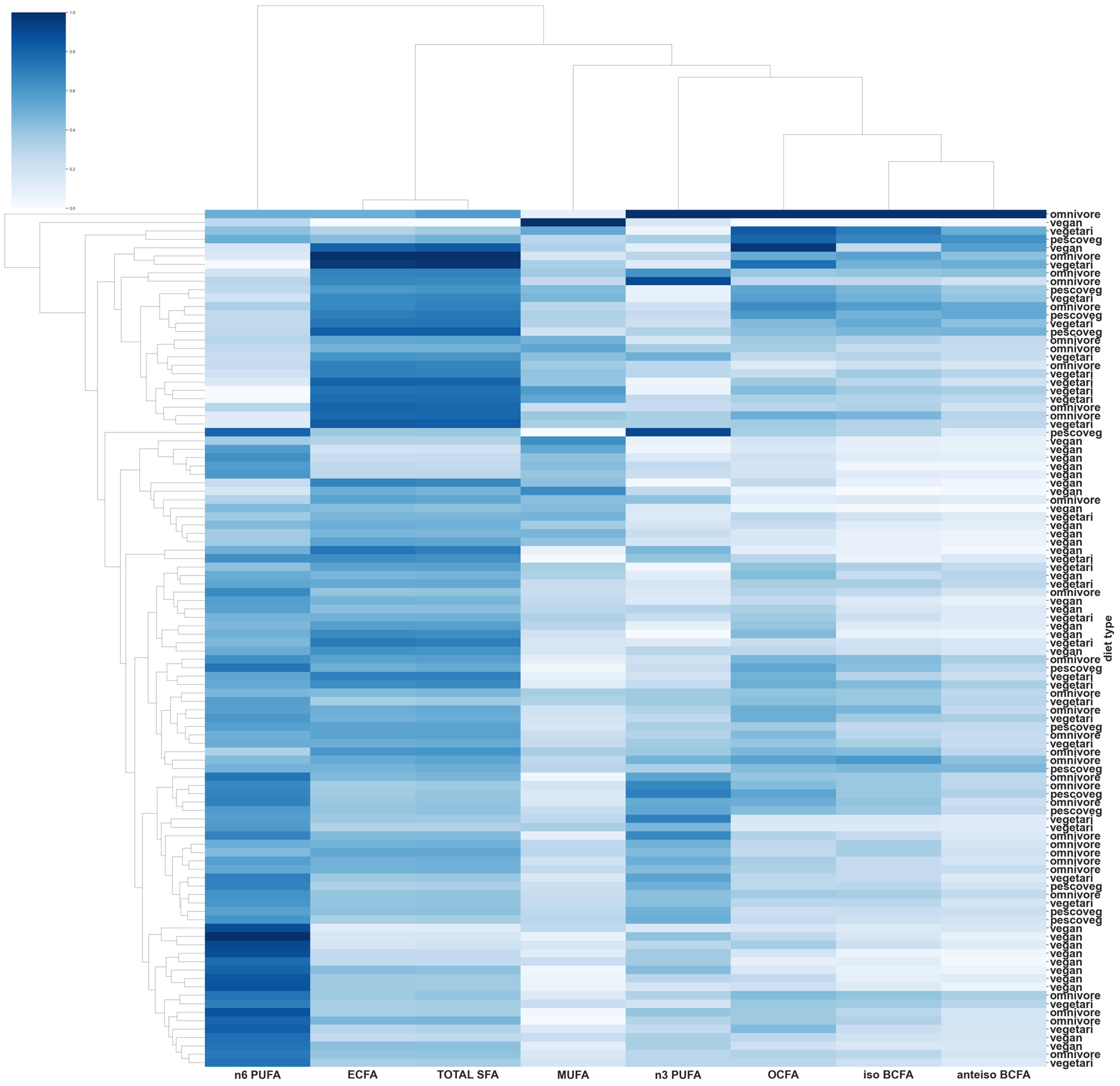

{kind=link}
{kind=link}
| Vegans (n = 30) | Vegetarians (n = 28) | Pescatarians (n = 13) | Omnivores (n = 31) | p | |
|---|---|---|---|---|---|
| Age [y] | 28.5 ± 6.3 | 27.2 ± 4.4 | 27.8 ± 7.7 | 35.6 ± 10.4 * | <0.001 |
| Body mass [kg] | 58.6 ± 6.1 | 62.4 ± 5.4 | 60.2 ± 6.4 | 61.1 ± 9.2 | 0.16 |
| BMI [kg/m2] | 21.0 ± 1.8 | 21.6 ± 1.7 | 22.0 ± 2.0 | 22.0 ± 1.9 | 0.19 |
| Body fat [%] | 24.8 ± 3.8 | 25.0 ± 3.8 | 24.5 ± 3.5 | 27.5 ± 6.2 | 0.08 |
| WHR | 0.77 ± 0.05 | 0.75 ± 0.04 | 0.76 ± 0.05 | 0.81 ± 0.04 * | <0.001 |
| FAs Levels [%] | Diet Type | Mean ± SD | n | p | |||||
|---|---|---|---|---|---|---|---|---|---|
| Vgn vs. Vgt | Vgn vs. Psc | Vgn vs. Omv | Vgt vs. Psc | Vgt vs. Omv | Psc vs. Omv | ||||
| ECSFAs | Vgn | 31.51 ± 3.46 | 30 | 0.004 | 0.260 | 0.030 | 0.054 | 0.400 | 0.230 |
| Vgt | 34.36 ± 3.34 | 28 | |||||||
| Psc | 32.52 ± 2.01 | 12 | |||||||
| Omv | 33.60 ± 2.16 | 30 | |||||||
| OCFAs | Vgn | 0.55 ± 0.11 | 29 | <0.001 | <0.001 | <0.001 | 0.020 | 0.490 | 0.090 |
| Vgt | 0.68 ± 0.10 | 26 | |||||||
| Psc | 0.77 ± 0.16 | 13 | |||||||
| Omv | 0.71 ± 0.11 | 30 | |||||||
| Iso-BCFAs | Vgn | 0.09 ± 0.03 | 28 | <0.001 | <0.001 | <0.001 | 0.060 | 0.080 | 0.440 |
| Vgt | 0.19 ± 0.06 | 27 | |||||||
| Psc | 0.22 ± 0.05 | 13 | |||||||
| Omv | 0.22 ± 0.06 | 30 | |||||||
| Anteiso-BCFAs | Vgn | 0.09 ± 0.03 | 28 | <0.001 | <0.001 | <0.001 | 0.020 | 0.990 | 0.010 |
| Vgt | 0.18 ± 0.06 | 28 | |||||||
| Psc | 0.22 ± 0.08 | 13 | |||||||
| Omv | 0.17 ± 0.04 | 29 | |||||||
| Total BCFAs | Vgn | 0.19 ± 0.06 | 28 | <0.001 | <0.001 | <0.001 | 0.160 | 0.600 | 0.340 |
| Vgt | 0.38 ± 0.11 | 27 | |||||||
| Psc | 0.47 ± 0.15 | 13 | |||||||
| Omv | 0.41 ± 0.10 | 30 | |||||||
| SFAs | Vgn | 32.31 ± 3.57 | 30 | 0.001 | 0.110 | 0.010 | 0.080 | 0.440 | 0.280 |
| Vgt | 35.48 ± 3.41 | 28 | |||||||
| Psc | 33.76 ± 2.17 | 12 | |||||||
| Omv | 34.76 ± 2.19 | 30 | |||||||
| MUFAs | Vgn | 31.72 ± 3.42 | 29 | 0.830 | 0.150 | 0.230 | 0.110 | 0.180 | 0.750 |
| Vgt | 31.92 ± 2.85 | 28 | |||||||
| Psc | 30.40 ± 2.37 | 13 | |||||||
| Omv | 30.67 ± 2.55 | 31 | |||||||
| n-3 PUFAs | Vgn | 0.87 ± 0.37 | 30 | 0.310 | <0.001 | <0.001 | 0.010 | 0.010 | 1.000 |
| Vgt | 1.00 ± 0.48 | 28 | |||||||
| Psc | 1.27 ± 0.41 | 12 | |||||||
| Omv | 1.35 ± 0.41 | 29 | |||||||
| n-6 PUFAs | Vgn | 34.61 ± 4.60 | 30 | 0.040 | 0.560 | 0.200 | 0.110 | 0.360 | 0.430 |
| Vgt | 31.59 ± 4.70 | 28 | |||||||
| Psc | 33.85 ± 3.59 | 13 | |||||||
| Omv | 32.80 ± 4.11 | 31 | |||||||
| EPA | Vgn | 0.21 ± 0.10 | 30 | 0.660 | 0.020 | <0.001 | 0.040 | 0.002 | 0.210 |
| Vgt | 0.23 ± 0.11 | 27 | |||||||
| Psc | 0.30 ± 0.10 | 11 | |||||||
| Omv | 0.34 ± 0.12 | 30 | |||||||
| DHA | Vgn | 0.22 ± 0.17 | 30 | 0.050 | <0.001 | <0.001 | <0.001 | 0.050 | 0.120 |
| Vgt | 0.48 ± 0.27 | 28 | |||||||
| Psc | 0.74 ± 0.36 | 13 | |||||||
| Omv | 0.63 ± 0.23 | 29 | |||||||
| LA | Vgn | 31.12 ± 3.93 | 30 | 0.020 | 0.770 | 0.030 | 0.040 | 0.910 | 0.040 |
| Vgt | 27.90 ± 0.39 | 28 | |||||||
| Psc | 30.73 ± 1.41 | 9 | |||||||
| Omv | 28.05 ± 3.99 | 31 | |||||||
| ARA | Vgn | 2.54 ± 0.99 | 30 | 0.740 | 0.130 | <0.001 | 0.210 | <0.001 | 0.056 |
| Vgt | 2.62 ± 0.98 | 28 | |||||||
| Psc | 3.04 ± 1.09 | 9 | |||||||
| Omv | 3.66 ± 0.90 | 31 | |||||||
| ALA | Vgn | 0.15 ± 0.06 | 29 | 0.110 | 0.860 | 0.280 | 0.120 | 0.010 | 0.240 |
| Vgt | 0.11 ± 0.06 | 27 | |||||||
| Psc | 0.14 ± 0.05 | 13 | |||||||
| Omv | 0.17 ± 0.08 | 30 | |||||||
| FA Intake [g] | Diet Type | Mean ± SD | n | p | |||||
|---|---|---|---|---|---|---|---|---|---|
| Vgn vs. Vgt | Vgn vs. Psc | Vgn vs. Omv | Vgt vs. Psc | Vgt vs. Omv | Psc vs. Omv | ||||
| SFAs | Vgn | 15.09 ± 7.05 | 30 | 0.049 | 0.010 | 0.030 | 0.430 | 0.760 | 0.600 |
| Vgt | 19.26 ± 5.01 | 24 | |||||||
| Psc | 21.02 ± 8.19 | 12 | |||||||
| Omv | 19.92 ± 6.67 | 25 | |||||||
| MUFAs | Vgn | 15.09 ± 7.05 | 30 | 0.070 | 0.030 | 0.001 | 0.600 | 0.110 | 0.250 |
| Vgt | 19.06 ± 5.01 | 24 | |||||||
| Psc | 22.85 ± 6.51 | 11 | |||||||
| Omv | 25.19 ± 6.49 | 27 | |||||||
| Total PUFAs | Vgn | 15.58 ± 7.33 | 30 | 0.040 | 0.080 | 0.050 | 0.610 | 0.820 | 0.750 |
| Vgt | 11.75 ± 2.81 | 23 | |||||||
| Psc | 12.66 ± 4.16 | 11 | |||||||
| Omv | 12.14 ± 3.99 | 27 | |||||||
| n-3 PUFAs | Vgn | 1.53 ± 0.98 | 28 | 0.570 | 0.030 | 0.770 | 0.010 | 0.420 | 0.040 |
| Vgt | 1.33 ± 0.77 | 23 | |||||||
| Psc | 2.35 ± 1.44 | 11 | |||||||
| Omv | 1.63 ± 1.26 | 26 | |||||||
| n-6 PUFAs | Vgn | 8.91 ± 5.11 | 27 | 0.380 | 0.880 | 0.120 | 0.340 | 0.430 | 0.100 |
| Vgt | 7.62 ± 4.09 | 24 | |||||||
| Psc | 9.13 ± 3.01 | 11 | |||||||
| Omv | 6.44 ± 4.89 | 26 | |||||||
| EPA + DHA * | Vgn | 0.00 ± 0.00 | 30 | 0.500 | 0.120 | 0.001 | 1.000 | 0.400 | 1.000 |
| Vgt | 0.02 ± 0.04 | 23 | |||||||
| Psc | 0.03 ± 0.43 | 12 | |||||||
| Omv | 0.10 ± 0.19 | 24 | |||||||
| ALA | Vgn | 1.73 ± 1.12 | 29 | 0.910 | 0.004 | 0.650 | 0.010 | 0.710 | 0.010 |
| Vgt | 1.78 ± 1.05 | 24 | |||||||
| Psc | 3.00 ± 2.45 | 12 | |||||||
| Omv | 1.93 ± 0.91 | 27 | |||||||
| LA | Vgn | 11.13 ± 6.27 | 30 | 0.280 | 0.270 | 0.260 | 0.920 | 0.880 | 0.950 |
| Vgt | 9.58 ± 2.20 | 22 | |||||||
| Psc | 9.45 ± 2.93 | 10 | |||||||
| Omv | 9.36 ± 3.20 | 27 | |||||||
| n-6/n-3 | Vgn | 6.53 ± 2.42 | 26 | 0.120 | 0.060 | 0.030 | 0.670 | 0.450 | 0.700 |
| Vgt | 5.30 ± 2.19 | 20 | |||||||
| Psc | 4.97 ± 1.67 | 8 | |||||||
| Omv | 4.67 ± 2.02 | 26 | |||||||
| Nutrient Intake | Diet Type | Me ± IQR | Mean ± SD | n | p | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Vgn vs. Vgt | Vgn vs. Psc | Vgn vs. Omv | Vgt vs. Psc | Vgt vs. Omv | Psc vs. Omv | |||||
| Protein [% of energy] | Vgn | 11.3 ± 2.5 | 29 | 1.00 | 0.15 | <0.001 | 0.47 | <0.001 | 0.09 | |
| Vgt | 12.1 ± 11.9 | 25 | ||||||||
| Psc | 13.9 ± 5.2 | 12 | ||||||||
| Omv | 19.0 ± 11.1 | 27 | ||||||||
| Fats [% of energy] | Vgn | 30.0 ± 8.7 | 29 | 1.00 | 1.00 | 0.006 | 1.00 | <0.001 | 0.38 | |
| Vgt | 28.5 ± 26.0 | 25 | ||||||||
| Psc | 30.3 ± 4.8 | 11 | ||||||||
| Omv | 35.0 ± 6.6 | 27 | ||||||||
| Carbohydrates [% of energy] | Vgn | 56.4 ± 6.5 | 27 | <0.001 | 0.15 | 0.18 | 1.00 | 0.29 | 1.00 | |
| Vgt | 49.0 ± 46.7 | 25 | ||||||||
| Pcs | 50.9 ± 6.9 | 12 | ||||||||
| Omv | 51.8 ± 12.4 | 26 | ||||||||
| Fiber [g] | Vgn | 44.6 ± 15.7 | 30 | <0.001 | 0.32 | 0.03 | 0.49 | 0.60 | 0.97 | |
| Vgt | 28.3 ± 10.7 | 24 | ||||||||
| Psc | 36.6 ± 9.6 | 12 | ||||||||
| Omv | 34.5 ± 13.0 | 27 | ||||||||
| Total energy [kcal] | Vgn | 2007 ± 506 | 30 | 0.74 | 0.97 | 0.91 | 0.99 | 0.98 | 1.00 | |
| Vgt | 1896 ± 385 | 24 | ||||||||
| Psc | 1944 ± 257 | 11 | ||||||||
| Omv | 1938 ± 304 | 27 | ||||||||
| Diet Type | r | p | n |
|---|---|---|---|
| Vegan | 0.07 | 0.71 | 28 |
| Vegetarian | −0.29 | 0.16 | 24 |
| Pescatarian | −0.20 | 0.53 | 12 |
| Omnivore | 0.19 | 0.36 | 25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gogga, P.; Mika, A.; Janczy, A.; Sztendel, A.; Sledzinski, T.; Małgorzewicz, S. Profiles of Serum Fatty Acids in Healthy Women on Different Types of Vegetarian Diets. Nutrients 2024, 16, 516. https://doi.org/10.3390/nu16040516
Gogga P, Mika A, Janczy A, Sztendel A, Sledzinski T, Małgorzewicz S. Profiles of Serum Fatty Acids in Healthy Women on Different Types of Vegetarian Diets. Nutrients. 2024; 16(4):516. https://doi.org/10.3390/nu16040516
Chicago/Turabian StyleGogga, Patrycja, Adriana Mika, Agata Janczy, Alicja Sztendel, Tomasz Sledzinski, and Sylwia Małgorzewicz. 2024. "Profiles of Serum Fatty Acids in Healthy Women on Different Types of Vegetarian Diets" Nutrients 16, no. 4: 516. https://doi.org/10.3390/nu16040516
APA StyleGogga, P., Mika, A., Janczy, A., Sztendel, A., Sledzinski, T., & Małgorzewicz, S. (2024). Profiles of Serum Fatty Acids in Healthy Women on Different Types of Vegetarian Diets. Nutrients, 16(4), 516. https://doi.org/10.3390/nu16040516