
Effect of Blueberry Supplementation on a Diet-Induced Rat Model of Prediabetes—Focus on Hepatic Lipid Deposition, Endoplasmic Stress Response and Autophagy

 , , , , , ,
, , , , , ,  , , ,
, , ,  ,
,  , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Blueberry Juice Preparation
2.2. Blueberry Juice Characterization
2.2.1. Phytochemical Polyphenolic Composition
2.2.2. Total Phenolic Content
2.2.3. Antioxidant Capacity
2.2.4. Dietary Fiber
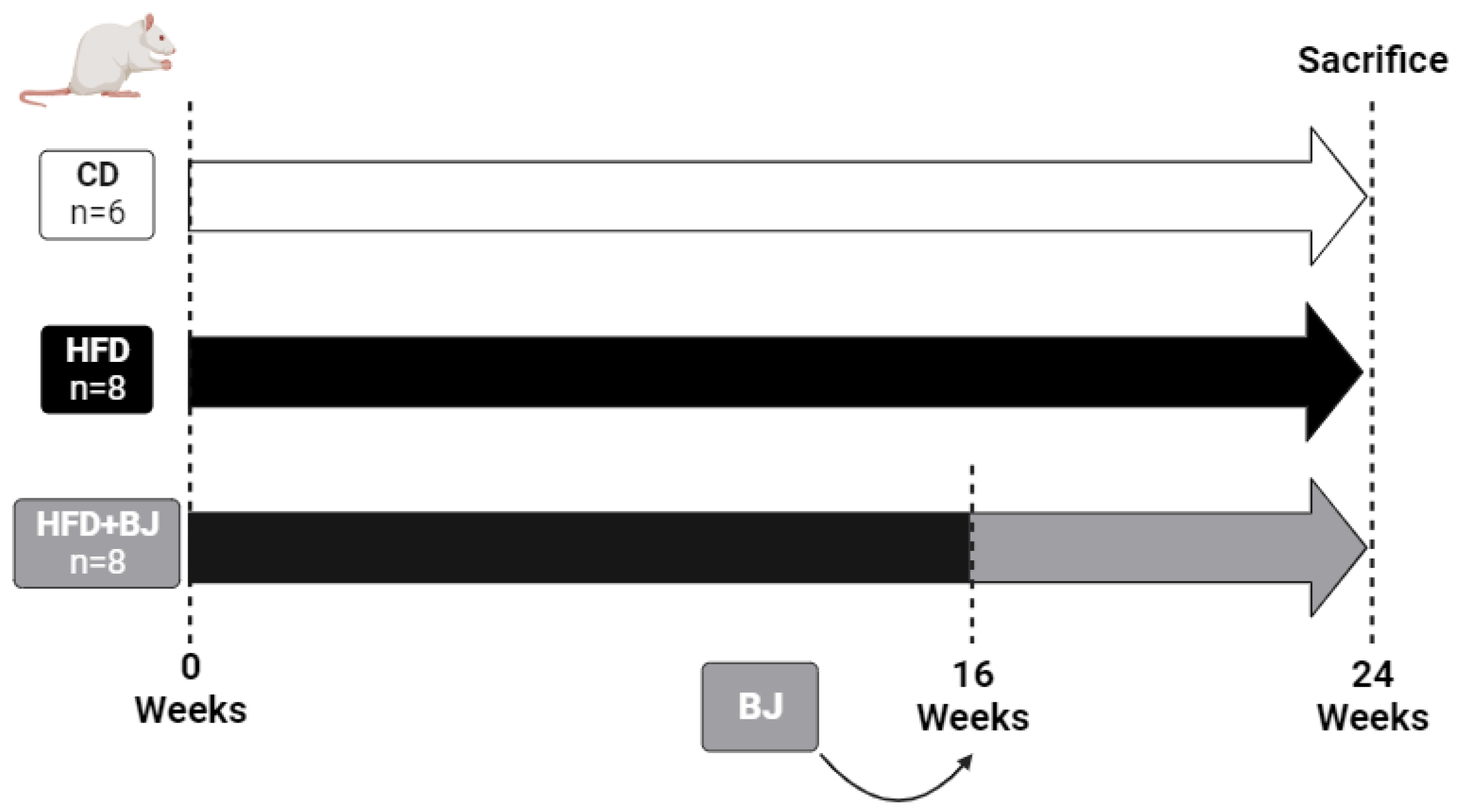
2.3. Experimental Design
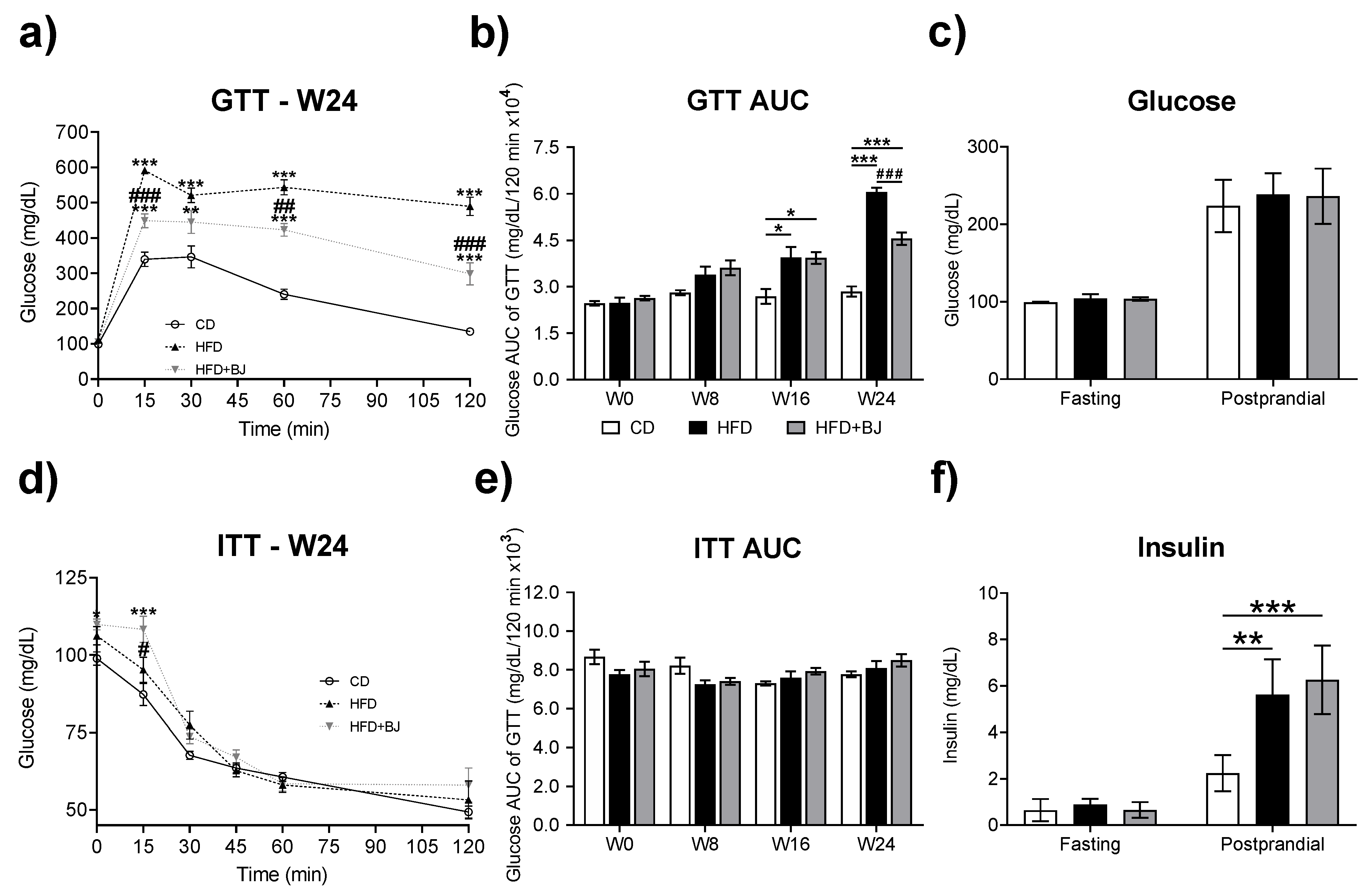
2.4. Glycemic and Insulinemic Profile
2.4.1. Glucose Tolerance Test (GTT) and Insulin Tolerance Test (ITT)
2.4.2. Fasting and Postprandial Glucose and Insulin
2.5. Blood and Tissue Collection
2.6. Serum Lipid Profile
2.7. Extraction and Quantification of Gut Microbiota in Feces
2.7.1. DNA Extraction from Stool
2.7.2. Real-Time PCR for Microbial Analysis of Stool
2.8. Fecal Short-Chain Fatty Acids Determination
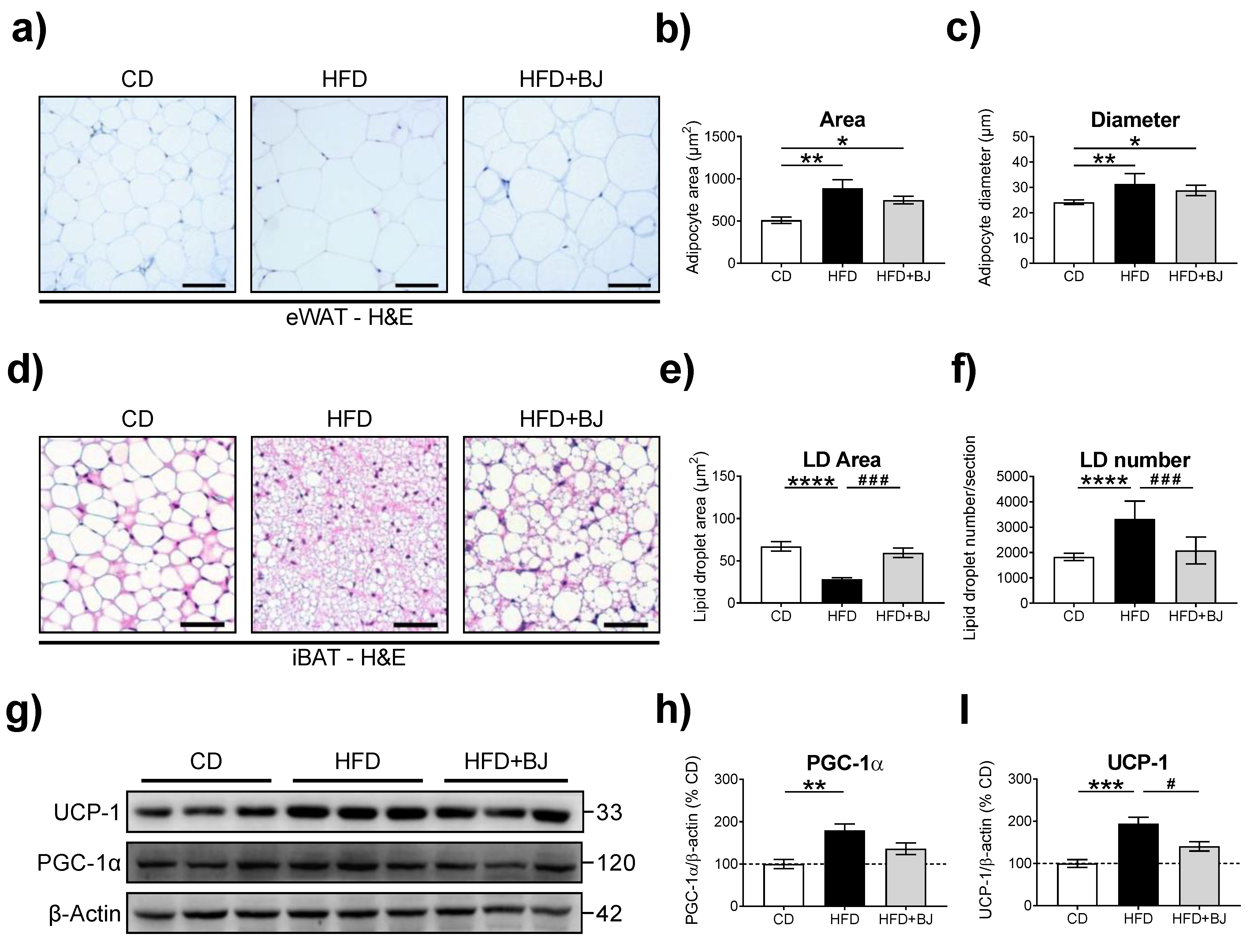
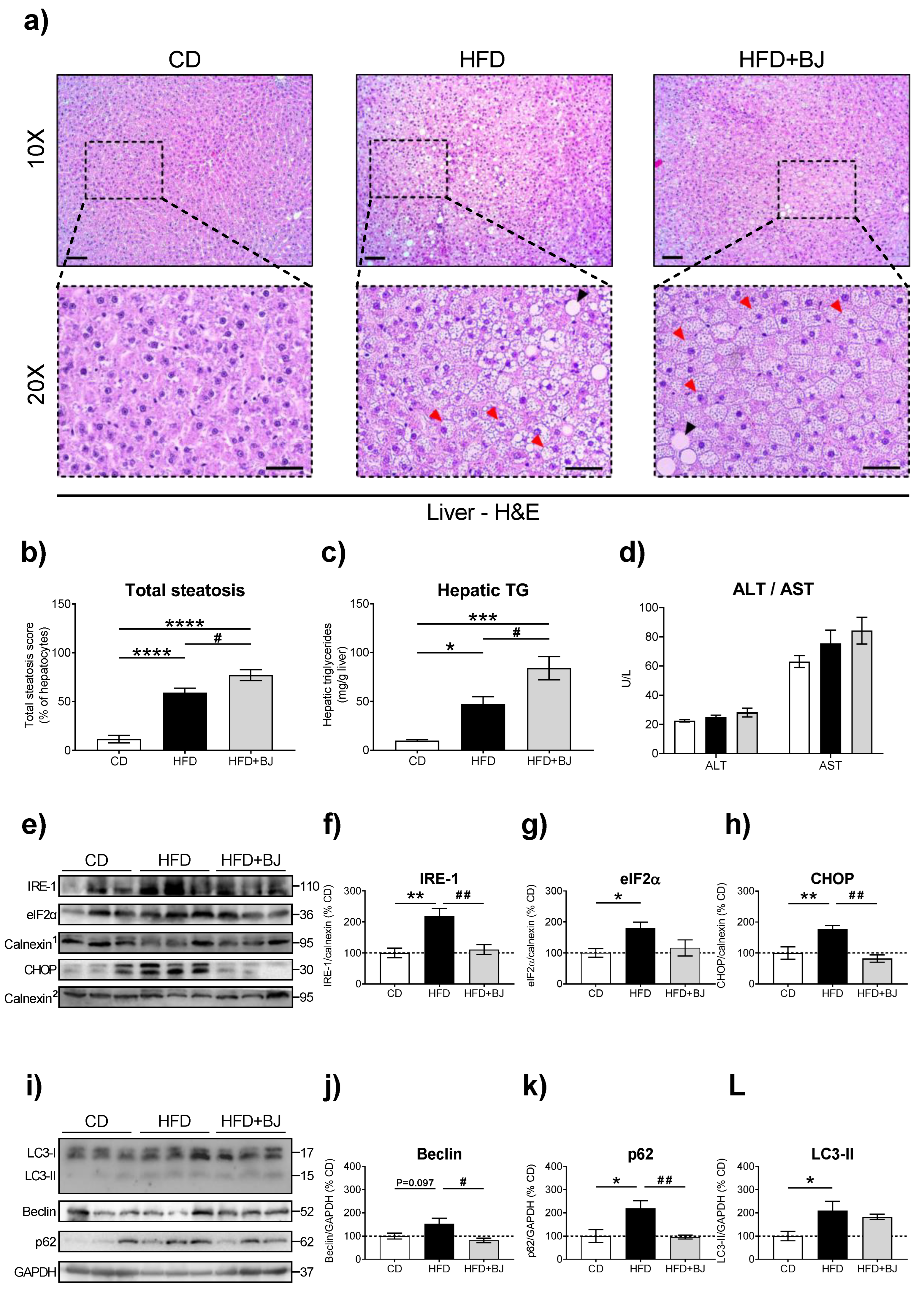
2.9. Liver, eWAT and iBAT Histomorphology
2.9.1. Hematoxylin and Eosin (H&E) Staining
2.9.2. Image Analysis and Data Quantification
Liver
eWAT
iBAT
2.10. Hepatic Triglycerides Quantification
2.11. Hepatic Enzymes Quantification
2.12. Protein Expression by Western Blotting
2.12.1. Protein Extraction and Quantification
2.12.2. Polyacrylamide Gel Electrophoresis and Immunodetection
2.13. Data Processing and Statistical Analysis
3. Results
3.1. Blueberry Juice Composition Characterization
3.2. Effects of BJ Nutraceutical Intervention on Glycemic and Insulinemic Profile
3.3. Effects of BJ Nutraceutical Intervention on Body and Tissue Weights
3.4. Effects of BJ Nutraceutical Intervention on Gut Microbiota
3.5. Effects of BJ Nutraceutical Intervention on Adiposity
3.6. Effects of BJ Nutraceutical Intervention on Hepatic Lipid Management
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tabák, A.G.; Herder, C.; Rathmann, W.; Brunner, E.J.; Kivimäki, M. Prediabetes: A High-Risk State for Diabetes Development. Lancet 2012, 379, 2279–2290. [Google Scholar] [CrossRef]
- Lee, K.-C.; Wu, P.-S.; Lin, H.-C. Pathogenesis and Treatment of Non-Alcoholic Steatohepatitis and Its Fibrosis. Clin. Mol. Hepatol. 2023, 29, 77–98. [Google Scholar] [CrossRef]
- Guillén, C.; Benito, M. MTORC1 Overactivation as a Key Aging Factor in the Progression to Type 2 Diabetes Mellitus. Front. Endocrinol. 2018, 9, 621. [Google Scholar] [CrossRef] [PubMed]
- Preguiça, I.; Alves, A.; Nunes, S.; Fernandes, R.; Gomes, P.; Viana, S.D.; Reis, F. Diet-Induced Rodent Models of Obesity-Related Metabolic Disorders-A Guide to a Translational Perspective. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2020, 21, e13081. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Sun, T.-Y.; He, Y.; Gou, W.; Zuo, L.-S.-Y.; Fu, Y.; Miao, Z.; Shuai, M.; Xu, F.; Xiao, C.; et al. Dietary Fruit and Vegetable Intake, Gut Microbiota, and Type 2 Diabetes: Results from Two Large Human Cohort Studies. BMC Med. 2020, 18, 371. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-Y.; Fang, J.-C.; Gao, Z.-H.; Zhang, C.; Xie, S.-Y. Higher Intake of Fruits, Vegetables or Their Fiber Reduces the Risk of Type 2 Diabetes: A Meta-Analysis. J. Diabetes Investig. 2016, 7, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Glechner, A.; Keuchel, L.; Affengruber, L.; Titscher, V.; Sommer, I.; Matyas, N.; Wagner, G.; Kien, C.; Klerings, I.; Gartlehner, G. Effects of Lifestyle Changes on Adults with Prediabetes: A Systematic Review and Meta-Analysis. Prim. Care Diabetes 2018, 12, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Costa, E.M.; Veiga, M.; Morais, R.M.; Calhau, C.; Pintado, M. Health Promoting Properties of Blueberries: A Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.; Vieira, P.; Gomes, P.; Viana, S.D.; Reis, F. Blueberry as an Attractive Functional Fruit to Prevent (Pre)Diabetes Progression. Antioxidants 2021, 10, 1162. [Google Scholar] [CrossRef] [PubMed]
- Stull, A.J. Blueberries’ Impact on Insulin Resistance and Glucose Intolerance. Antioxidants 2016, 5, 44. [Google Scholar] [CrossRef]
- Rada, P.; González-Rodríguez, Á.; García-Monzón, C.; Valverde, Á.M. Understanding Lipotoxicity in NAFLD Pathogenesis: Is CD36 a Key Driver? Cell Death Dis. 2020, 11, 802. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.-J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet–Induced Obesity via a PPARγ-Dependent Switch From Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef]
- Brahem, M.; Renard, C.M.G.C.; Eder, S.; Loonis, M.; Ouni, R.; Mars, M.; Le Bourvellec, C. Characterization and Quantification of Fruit Phenolic Compounds of European and Tunisian Pear Cultivars. Food Res. Int. 2017, 95, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.; Viana, S.D.; Preguiça, I.; Alves, A.; Fernandes, R.; Teodoro, J.S.; Figueirinha, A.; Salgueiro, L.; Silva, S.; Jarak, I.; et al. Blueberry Consumption Challenges Hepatic Mitochondrial Bioenergetics and Elicits Transcriptomics Reprogramming in Healthy Wistar Rats. Pharmaceutics 2020, 12, 1094. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of Total Phenolic Content and Other Oxidation Substrates in Plant Tissues Using Folin–Ciocalteu Reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Sánchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin-Ciocalteu Assay Revisited: Improvement of Its Specificity for Total Phenolic Content Determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M. Rice-Evans C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Burgeiro, A.; Cerqueira, M.G.; Varela-Rodríguez, B.M.; Nunes, S.; Neto, P.; Pereira, F.C.; Reis, F.; Carvalho, E. Glucose and Lipid Dysmetabolism in a Rat Model of Prediabetes Induced by a High-Sucrose Diet. Nutrients 2017, 9, 638. [Google Scholar] [CrossRef]
- Nunes, S.; Soares, E.; Fernandes, J.; Viana, S.; Carvalho, E.; Pereira, F.C.; Reis, F. Early Cardiac Changes in a Rat Model of Prediabetes: Brain Natriuretic Peptide Overexpression Seems to Be the Best Marker. Cardiovasc. Diabetol. 2013, 12, 44. [Google Scholar] [CrossRef]
- Madureira, A.R.; Nunes, S.; Campos, D.A.; Fernandes, J.C.; Marques, C.; Zuzarte, M.; Gullón, B.; Rodríguez-Alcalá, L.M.; Calhau, C.; Sarmento, B.; et al. Safety Profile of Solid Lipid Nanoparticles Loaded with Rosmarinic Acid for Oral Use: In Vitro and Animal Approaches. Int. J. Nanomed. 2016, 11, 3621–3640. [Google Scholar] [CrossRef]
- Queipo-Ortuño, M.I.; Seoane, L.M.; Murri, M.; Pardo, M.; Gomez-Zumaquero, J.M.; Cardona, F.; Casanueva, F.; Tinahones, F.J. Gut Microbiota Composition in Male Rat Models under Different Nutritional Status and Physical Activity and Its Association with Serum Leptin and Ghrelin Levels. PLoS ONE 2013, 8, e65465. [Google Scholar] [CrossRef] [PubMed]
- Delroisse, J.M.; Boulvin, A.L.; Parmentier, I.; Dauphin, R.D.; Vandenbol, M.; Portetelle, D. Quantification of Bifidobacterium spp. and Lactobacillus spp. in Rat Fecal Samples by Real-Time PCR. Microbiol. Res. 2008, 163, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Rinttilä, T.; Kassinen, A.; Malinen, E.; Krogius, L.; Palva, A. Development of an Extensive Set of 16S RDNA-Targeted Primers for Quantification of Pathogenic and Indigenous Bacteria in Faecal Samples by Real-Time PCR. J. Appl. Microbiol. 2004, 97, 1166–1177. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, T.; Watanabe, K.; Fujimoto, J.; Miyamoto, Y.; Takada, T.; Matsumoto, K.; Oyaizu, H.; Tanaka, R. Development of 16S rRNA-gene-targeted group-specific primers for the detection and identification of predominant bacteria in human feces. Appl. Environ. Microbiol. 2002, 68, 5445–5451. [Google Scholar] [CrossRef] [PubMed]
- Scortichini, S.; Chiara, M.; Silvi, S.; Fiorini, D. Development and Validation of a GC-FID Method for the Analysis of Short Chain Fatty Acids in Rat and Human Faeces and in Fermentation Fluids. J. Chromatogr. B 2020, 1143, 121972. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.; Viana, S.D.; Preguiça, I.; Alves, A.; Fernandes, R.; Teodoro, J.S.; Matos, P.; Figueirinha, A.; Salgueiro, L.; André, A.; et al. Blueberry Counteracts Prediabetes in a Hypercaloric Diet-Induced Rat Model and Rescues Hepatic Mitochondrial Bioenergetics. Nutrients 2021, 13, 4192. [Google Scholar] [CrossRef] [PubMed]
- Elks, C.M.; Terrebonne, J.D.; Ingram, D.K.; Stephens, J.M. Blueberries Improve Glucose Tolerance without Altering Body Composition in Obese Postmenopausal Mice. Obesity 2015, 23, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Marques, C.; Meireles, M.; Norberto, S.; Leite, J.; Freitas, J.; Pestana, D.; Faria, A.; Calhau, C. High-Fat Diet-Induced Obesity Rat Model: A Comparison between Wistar and Sprague-Dawley Rat. Adipocyte 2016, 5, 11–21. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human Gut Microbiota Changes Reveal the Progression of Glucose Intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef]
- Allin, K.H.; Tremaroli, V.; Caesar, R.; Jensen, B.A.H.; Damgaard, M.T.F.; Bahl, M.I.; Licht, T.R.; Hansen, T.H.; Nielsen, T.; Dantoft, T.M.; et al. Aberrant Intestinal Microbiota in Individuals with Prediabetes. Diabetologia 2018, 61, 810–820. [Google Scholar] [CrossRef]
- Wu, H.; Tremaroli, V.; Schmidt, C.; Lundqvist, A.; Olsson, L.M.; Krämer, M.; Gummesson, A.; Perkins, R.; Bergström, G.; Bäckhed, F. The Gut Microbiota in Prediabetes and Diabetes: A Population-Based Cross-Sectional Study. Cell Metab. 2020, 32, 379–390. [Google Scholar] [CrossRef] [PubMed]
- DeFuria, J.; Bennett, G.; Strissel, K.J.; Perfield, J.W., 2nd; Milbury, P.E.; Greenberg, A.S.; Obin, M.S. Dietary Blueberry Attenuates Whole-Body Insulin Resistance in High Fat-Fed Mice by Reducing Adipocyte Death and Its Inflammatory Sequelae. J. Nutr. 2009, 139, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Roopchand, D.E.; Kuhn, P.; Rojo, L.E.; Lila, M.A.; Raskin, I. Blueberry Polyphenol-Enriched Soybean Flour Reduces Hyperglycemia, Body Weight Gain and Serum Cholesterol in Mice. Pharmacol. Res. 2013, 68, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Han, X.; Tan, H.; Huang, W.; You, Y.; Zhan, J. Blueberry Extract Improves Obesity through Regulation of the Gut Microbiota and Bile Acids via Pathways Involving FXR and TGR5. iScience 2019, 19, 676–690. [Google Scholar] [CrossRef]
- Romestaing, C.; Piquet, M.A.; Bedu, E.; Rouleau, V.; Dautresme, M.; Hourmand-Ollivier, I.; Filippi, C.; Duchamp, C.; Sibille, B. Long Term Highly Saturated Fat Diet Does Not Induce NASH in Wistar Rats. Nutr. Metab. 2007, 4, 4. [Google Scholar] [CrossRef]
- Saito, M. Brown Adipose Tissue as a Regulator of Energy Expenditure and Body Fat in Humans. Diabetes Metab. J. 2013, 37, 22–29. [Google Scholar] [CrossRef]
- Wood Dos Santos, T.; Cristina Pereira, Q.; Teixeira, L.; Gambero, A.; A Villena, J.; Lima Ribeiro, M. Effects of Polyphenols on Thermogenesis and Mitochondrial Biogenesis. Int. J. Mol. Sci. 2018, 19, 2757. [Google Scholar] [CrossRef]
- Fedorenko, A.; Lishko, P.V.; Kirichok, Y. Mechanism of Fatty-Acid-Dependent UCP1 Uncoupling in Brown Fat Mitochondria. Cell 2012, 151, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Loftus, H.; McAinch, A.J.; Su, X.Q. Blueberry as a Source of Bioactive Compounds for the Treatment of Obesity, Type 2 Diabetes and Chronic Inflammation. J. Funct. Foods 2017, 30, 16–29. [Google Scholar] [CrossRef]
- Seymour, E.M.; Tanone, I.I.; Urcuyo-Llanes, D.E.; Lewis, S.K.; Kirakosyan, A.; Kondoleon, M.G.; Kaufman, P.B.; Bolling, S.F. Blueberry Intake Alters Skeletal Muscle and Adipose Tissue Peroxisome Proliferator-Activated Receptor Activity and Reduces Insulin Resistance in Obese Rats. J. Med. Food 2011, 14, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Vendrame, S.; Daugherty, A.; Kristo, A.S.; Klimis-Zacas, D. Wild Blueberry (Vaccinium Angustifolium)-Enriched Diet Improves Dyslipidaemia and Modulates the Expression of Genes Related to Lipid Metabolism in Obese Zucker Rats. Br. J. Nutr. 2014, 111, 194–200. [Google Scholar] [CrossRef]
- Granato, D.; Mocan, A.; Câmara, J.S. Is a Higher Ingestion of Phenolic Compounds the Best Dietary Strategy? A Scientific Opinion on the Deleterious Effects of Polyphenols in Vivo. Trends Food Sci. Technol. 2020, 98, 162–166. [Google Scholar] [CrossRef]
- Martin, K.R.; Appel, C.L. Polyphenols as Dietary Supplements: A Double-Edged Sword. Nutr. Diet. Suppl. 2009, 2020, 1–12. [Google Scholar] [CrossRef]
- Mao, X.; Gu, C.; Chen, D.; Yu, B.; He, J. Oxidative Stress-Induced Diseases and Tea Polyphenols. Oncotarget 2017, 8, 81649–81661. [Google Scholar] [CrossRef] [PubMed]
- Isbrucker, R.A.; Edwards, J.A.; Wolz, E.; Davidovich, A.; Bausch, J. Safety Studies on Epigallocatechin Gallate (EGCG) Preparations. Part 2: Dermal, Acute and Short-Term Toxicity Studies. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2006, 44, 636–650. [Google Scholar] [CrossRef]
- Galati, G.; Lin, A.; Sultan, A.M.; O’Brien, P.J. Cellular and in Vivo Hepatotoxicity Caused by Green Tea Phenolic Acids and Catechins. Free Radic. Biol. Med. 2006, 40, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-Y.; Han, J.; Cao, S.Y.; Hong, T.; Zhuo, D.; Shi, J.; Liu, Z.; Cao, W. Hepatic Autophagy Is Suppressed in the Presence of Insulin Resistance and Hyperinsulinemia: Inhibition of FoxO1-Dependent Expression of Key Autophagy Genes by Insulin. J. Biol. Chem. 2009, 284, 31484–31492. [Google Scholar] [CrossRef]
- Da Cruz, L.L.; Vesentini, G.; Sinzato, Y.K.; Villaverde, A.I.S.B.; Volpato, G.T.; Damasceno, D.C. Effects of High-Fat Diet-Induced Diabetes on Autophagy in the Murine Liver: A Systematic Review and Meta-Analysis. Life Sci. 2022, 309, 121012. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, J.; Wang, X.; Li, J.; Chen, J.; Wang, N.; Zhang, M.; Feng, Y.; Guo, H. 2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-Glucoside Induces Autophagy of Liver by Activating PI3K/Akt and Erk Pathway in Prediabetic Rats. BMC Complement. Med. Ther. 2020, 20, 177. [Google Scholar] [CrossRef]
- Leng, Y.-P.; Qiu, N.; Fang, W.; Zhang, M.; He, Z.-M.; Xiong, Y. Involvement of Increased Endogenous Asymmetric Dimethylarginine in the Hepatic Endoplasmic Reticulum Stress of Type 2 Diabetic Rats. PLoS ONE 2014, 9, e97125. [Google Scholar] [CrossRef]
- Pandey, V.K.; Mathur, A.; Khan, M.F.; Kakkar, P. Activation of PERK-EIF2α-ATF4 Pathway Contributes to Diabetic Hepatotoxicity: Attenuation of ER Stress by Morin. Cell. Signal. 2019, 59, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, Y.; Zhang, H.; Wang, Y.; Wu, C.; Huang, W. Malvidin Induces Hepatic Stellate Cell Apoptosis via the Endoplasmic Reticulum Stress Pathway and Mitochondrial Pathway. Food Sci. Nutr. 2020, 8, 5095–5106. [Google Scholar] [CrossRef] [PubMed]
- Kandeil, M.A.; Hashem, R.M.; Mahmoud, M.O.; Hetta, M.H.; Tohamy, M.A. Zingiber Officinale Extract and Omega-3 Fatty Acids Ameliorate Endoplasmic c Rat Model. J. Food Biochem. 2019, 43, e13076. [Google Scholar] [CrossRef]
- Zhuge, Q.; Zhang, Y.; Liu, B.; Wu, M. Blueberry Polyphenols Play a Preventive Effect on Alcoholic Fatty Liver Disease C57BL/6 J Mice by Promoting Autophagy to Accelerate Lipolysis to Eliminate Excessive TG Accumulation in Hepatocytes. Ann. Palliat. Med. 2020, 9, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Guo, H.; Yang, H.; Wang, D.; Gao, P.; Wei, W. Pterostilbene, An Active Constituent of Blueberries, Suppresses Proliferation Potential of Human Cholangiocarcinoma via Enhancing the Autophagic Flux. Front. Pharmacol. 2019, 10, 1238. [Google Scholar] [CrossRef]
- Singh, A.K.; Cabral, C.; Kumar, R.; Ganguly, R.; Rana, H.K.; Gupta, A.; Lauro, M.R.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial E Ff Ects of Dietary Polyphenols on Gut Microbiota and Strategies to Improve. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids in Control of Body Weight and Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef]
- Rosendo-Silva, D.; Viana, S.; Carvalho, E.; Reis, F.; Matafome, P. Are Gut Dysbiosis, Barrier Disruption, and Endotoxemia Related to Adipose Tissue Dysfunction in Metabolic Disorders? Overview of the Mechanisms Involved. Intern. Emerg. Med. 2023, 18, 1287–1302. [Google Scholar] [CrossRef]
- Dai, X.; Guo, Z.; Chen, D.; Li, L.; Song, X.; Liu, T.; Jin, G.; Li, Y.; Liu, Y.; Ajiguli, A.; et al. Maternal Sucralose Intake Alters Gut Microbiota of Offspring and Exacerbates Hepatic Steatosis in Adulthood. Gut Microbes 2020, 11, 1043–1063. [Google Scholar] [CrossRef]
- Guo, J.; Shi, C.-X.; Zhang, Q.-Q.; Deng, W.; Zhang, L.-Y.; Chen, Q.; Zhang, D.-M.; Gong, Z.-J. Interventions for Non-Alcoholic Liver Disease: A Gut Microbial Metabolites Perspective. Ther. Adv. Gastroenterol. 2022, 15, 17562848221138676. [Google Scholar] [CrossRef]
- Shah, S.; Fillier, T.; Pham, T.H.; Thomas, R.; Cheema, S.K. Intraperitoneal Administration of Short-Chain Fatty Acids Improves Lipid Metabolism of Long-Evans Rats in a Sex-Specific Manner. Nutrients 2021, 13, 892. [Google Scholar] [CrossRef]
- Kalt, W.; Ryan, D.A.J.; Duy, J.C.; Prior, R.L.; Ehlenfeldt, M.K.; Vander Kloet, S.P. Interspecific Variation in Anthocyanins, Phenolics, and Antioxidant Capacity among Genotypes of Highbush and Lowbush Blueberries (Vaccinium Section Cyanococcus spp.). J. Agric. Food Chem. 2001, 49, 4761–4767. [Google Scholar] [CrossRef]
- Lila, M.A. The Nature-versus-Nurture Debate on Bioactive Phytochemicals: The Genome versus Terroir. J. Sci. Food Agric. 2006, 86, 2510–2515. [Google Scholar] [CrossRef]
- Rodriguez-Mateos, A.; Cifuentes-Gomez, T.; Tabatabaee, S.; Lecras, C.; Spencer, J.P.E. Procyanidin, Anthocyanin, and Chlorogenic Acid Contents of Highbush and Lowbush Blueberries. J. Agric. Food Chem. 2012, 60, 5772–5778. [Google Scholar] [CrossRef]
- Yousef, G.G.; Brown, A.F.; Funakoshi, Y.; Mbeunkui, F.; Grace, M.H.; Ballington, J.R.; Loraine, A.; Lila, M.A. Efficient Quantification of the Health-Relevant Anthocyanin and Phenolic Acid Profiles in Commercial Cultivars and Breeding Selections of Blueberries (Vaccinium spp.). J. Agric. Food Chem. 2013, 61, 4806–4815. [Google Scholar] [CrossRef]
- Weaver, C.M.; Ferruzzi, M.G.; Maiz, M.; Cladis, D.P.; Nakatsu, C.H.; McCabe, G.P.; Lila, M.A. Crop, Host, and Gut Microbiome Variation Influence Precision Nutrition: An Example of Blueberries. Antioxidants 2023, 12, 1136. [Google Scholar] [CrossRef] [PubMed]
- Gezer, C. Stress Response of Dietary Phytochemicals in a Hormetic Manner for Health and Longevity. In Gene Expression and Regulation in Mammalian Cells; Uchiumi, F., Ed.; IntechOpen: Rijeka, Croatia, 2018. [Google Scholar]
- Son, T.G.; Camandola, S.; Mattson, M.P. Hormetic Dietary Phytochemicals. NeuroMol. Med. 2008, 10, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Mattson, M.P. How Does Hormesis Impact Biology, Toxicology, and Medicine? Aging Mech. Dis. 2017, 3, 13. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Group | Primer Sequence (5′-3′) | Genomic DNA Standard | PCR Product Size (bp) | AT (°C) | Ref. |
|---|---|---|---|---|---|
| Firmicutes | AGC TGA CGA CAA CCA TGC AC ATG TGG TTT AAT TCG AAG CA | Lactobacillus gasseri ATCC 33323 | 126 | 45 | [21] |
| Lactobacillus | AGC AGT AGG GAA TCT TCC A CAC CGC TAC ACA TGG AG | Lactobacillus gasseri ATCC 33323 | 351 | 55 | [22] |
| Enterococcus | ACT CGT TGT ACT TCC CTT GT CCC TTA TTG TTA GTT GCC ATC ATT | Enterococcus gilvus ATCC BAA-350 | 155 | 50 | [23] |
| Clostridium leptum | CTT CCT CCG TTT TGT CAA GCA CAA GCA GTG GAG T | Clostridium leptum ATCC 29065 | 239 | 45 | [23] |
| Roseburia | CGG CAC CGA AGA GCA AT TAC TGC ATT GGA AAC TGT CG | Roseburia hominis A2-183 | 230 | 50 | [23] |
| Bacteroidetes | AGC TGA CGA CAA CCA TGC AG CAT GTG GTT TAA TTC GAT | Bacteroides vulgatus ATCC 8482 | 126 | 45 | [21] |
| Prevotella | GGT CGG GTT GCA GAC C CAC RGT AAA CGA TGG ATG CC | Prevotella nigrescens ATCC 33563 | 513 | 50 | [24] |
| Bacteroides | CCA GTA TCA ACT GCA ATT TTA ATA GCC TTT CGA AAG RAA GAT | Bacteroides vulgatus ATCC 8482 | 495 | 45 | [24] |
| Bifidobacterium | CCA GTA TCA ACT GCA ATT TTA ATA GCC TTT CGA AAG RAA GAT | Bifidobacterium longum subsp. Infantis ATCC 15697 | 244 | 50 | [21] |
| Antibody | Reference | Source | Animal Origin | Blocking Solution | Solution | Dilution | Incubation Time |
|---|---|---|---|---|---|---|---|
| Primary Antibody | |||||||
| Anti-PGC1α | Ab191838 | Abcam | Rabbit | 5% Milk in TBS-T | 1% Milk in TBS-T | 1:1500 | 48 h |
| Anti-UCP1 | Ab23841 | Abcam | Rabbit | 5% Milk in TBS-T | 1% Milk in TBS-T | 1:1500 | 24 h |
| Anti-eIF2α | 5324 | Cell Signaling Technology | Rabbit | 5% BSA in TBS-T | 1% BSA in TBS-T | 1:2000 | 24 h |
| Anti-IRE1α | 3294 | Cell Signaling Technology | Rabbit | 5% BSA in TBS-T | 1% BSA in TBS-T | 1:1000 | 24 h |
| Anti-CHOP | 2895 | Cell Signaling Technology | Mouse | 5% BSA in TBS-T | 1% BSA in TBS-T | 1:1000 | 24 h |
| Anti-SQSTMI/p62 | 5114S | Cell Signaling Technology | Rabbit | 5% BSA in TBS-T | 1% BSA in TBS-T | 1:1000 | 24 h |
| Anti-LC3 | PA1-16931 | Thermo Fisher Scientific | Rabbit | 5% BSA in TBS-T | 1% BSA in TBS-T | 1:1000 | 24 h |
| Loading Controls | |||||||
| Anti-β-actin | AB0145-200 | Sicgen | Goat | 5% BSA in TBS-T | 1% BSA in TBS-T | 1:1000 | 24 h |
| Anti-Calnexin | AB0041-200 | Sicgen | Goat | 5% BSA in TBS-T | 1% BSA in TBS-T | 1:1000 | 24 h |
| Anti-GAPDH | AB0049-200 | Sicgen | Goat | 5% BSA in TBS-T | 1% BSA in TBS-T | 1:5000 | 24 h |
| Secondary Antibodies | |||||||
| Anti-Rabbit | R-0572-050 | Advansta | Goat | Same as respective antibodies | 1:10,000 | 1 h | |
| Anti-Mouse | R-05071-500 | Advansta | Goat | 1:10,000 | 1 h | ||
| Anti-Goat | AB1011-1000 | Sicgen | Goat | 1:10,000 | 1 h | ||
| CD | HFD | HFD + BJ | |
|---|---|---|---|
| Body and Tissue Weights | |||
| Δ BW (g) | 121.600 ± 4.202 | 202.286 ± 23.550 * | 204.250 ± 18.890 * |
| Δ BW (%) | 32.600 ± 1.166 | 53.143 ± 6.212 * | 50.857 ± 3.575 * |
| eWAT (g) | 10.047 ± 0.544 | 23.936 ± 1.762 *** | 22.928 ± 2.221 *** |
| eWAT/BW | 2.044 ± 0.135 | 4.733 ± 0.374 **** | 4.199 ± 0.289 *** |
| iBAT (g) | 0.707 ± 0.011 | 0.925 ± 0.062 | 0.947 ± 0.075 * |
| iBAT/BW | 0.146 ± 0.004 | 0.173 ± 0.014 | 0.175 ± 0.012 |
| Liver (g) | 15.780 ± 0.028 | 15.387 ± 0.853 | 16.359 ± 0.973 |
| Liver/BW | 3.384 ± 0.097 | 2.550 ± 0.138 *** | 2.831 ± 0.080 ** |
| Food and Beverage Intake | |||
| Food (g/week) | 145.833 ± 2.027 | 104.127 ± 4.400 **** | 105.797 ± 4.162 **** |
| Beverage (mL/week) | 198.750 ± 2.294 | 176.453 ± 5.435 ** | 228.047 ± 8.098 #### |
| Calories (Kcal/week) | |||
| Carbohydrates | 311.916 ± 4.334 | 199.185 ± 8.412 **** | 210.557 ± 7.634 **** |
| Lipids | 39.506 ± 0.551 | 218.272 ± 9.220 **** | 221.773 ± 8.725 **** |
| Proteins | 107.953 ± 1.500 | 71.942 ± 3.039 **** | 73.095 ± 2.874 **** |
| Total | 459.375 ± 6.385 | 489.399 ± 20.670 | 505.425 ± 19.020 |
| CD | HFD | HFD + BJ | |
|---|---|---|---|
| Gut Microbiota Composition (Log10 copies/ng of DNA) | |||
| Firmicutes | 4.882 ± 0.547 | 6.678 ± 0.247 ** | 4.550 ± 0.157 ## |
| Bacteroidetes | 4.228 ± 0.252 | 4.146 ± 0.129 | 4.504 ± 0.511 |
| Firmicutes/Bacteroidetes | 1.276 ± 0.035 | 1.590 ± 0.062 * | 1.078 ± 0.095 ### |
| Bifidobacterium spp. | 3.230 ± 0.594 | 1.768 ± 0.130 | 1.452 ± 0.153* |
| Lactobacillus spp. | 4.828 ± 0.245 | 3.828 ± 0.100 ** | 3.899 ± 0.183* |
| Prevotella spp. | 3.203 ± 0.417 | 1.437 ± 0.119 ** | 2.139 ± 0.228 |
| Bacteroides spp. | 2.977 ± 0.062 | 2.902 ± 0.332 | 2.086 ± 0.230 |
| Clostridium Leptum | 3.165 ± 0.114 | 3.308 ± 0.260 | 2.680 ± 0.130 |
| Enterococcus spp. | 2.583 ± 0.177 | 2.813 ± 0.161 | 3.068 ± 0.233 |
| Roseburia | 2.234 ± 0.306 | 2.038 ± 0.175 | 1.764 ± 0.453 |
| Short-Chain Fatty Acids (µmol/g) | |||
| C2 Acetic | 113.230 ± 23.730 | 56.566 ± 9.231 * | 52.059 ± 6.625 * |
| C3 Propionic | 10.225 ± 2.441 | 7.095 ± 1.631 | 3.449 ± 0.554 * |
| C4 Butyric | 6.182 ± 1.323 | 4.129 ± 1.186 | 2.450 ± 0.478 * |
| iC4 Isobutyric | 0.574 ± 0.050 | 0.291 ± 0.114 | 0.183 ± 0.033 * |
| iC5 Isovaleric | 0.858 ± 0.088 | 0.246 ± 0.034 * | 0.220 ± 0.018 ** |
| C5 Valeric | 0.820 ± 0.048 | 0.541 ± 0.124 | 0.308 ± 0.046 *** |
| C6 Caproic | 0.929 ± 0.148 | 0.430 ± 0.062 ** | 0.379 ± 0.039 ** |
| Serum (mg/dL) | CD | HFD | HFD + BJ |
|---|---|---|---|
| Triglycerides | 237.000 ± 27.180 | 135.333 ± 17.920 | 242.500 ± 29.420 # |
| LDL-c | 4.000 ± 0.548 | 7.714 ± 0.865 * | 7.625 ± 0.800 * |
| HDL-c | 24.200 ± 1.828 | 27.857 ± 2.539 | 31.000 ± 1.604 |
| Total cholesterol | 74.800 ± 6.070 | 80.857 ± 5.869 | 95.500 ± 4.424 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, G.; Vieira, P.; Alves, A.; Nunes, S.; Preguiça, I.; Martins-Marques, T.; Ribeiro, T.; Girão, H.; Figueirinha, A.; Salgueiro, L.; et al. Effect of Blueberry Supplementation on a Diet-Induced Rat Model of Prediabetes—Focus on Hepatic Lipid Deposition, Endoplasmic Stress Response and Autophagy. Nutrients 2024, 16, 513. https://doi.org/10.3390/nu16040513
Ferreira G, Vieira P, Alves A, Nunes S, Preguiça I, Martins-Marques T, Ribeiro T, Girão H, Figueirinha A, Salgueiro L, et al. Effect of Blueberry Supplementation on a Diet-Induced Rat Model of Prediabetes—Focus on Hepatic Lipid Deposition, Endoplasmic Stress Response and Autophagy. Nutrients. 2024; 16(4):513. https://doi.org/10.3390/nu16040513
Chicago/Turabian StyleFerreira, Gonçalo, Pedro Vieira, André Alves, Sara Nunes, Inês Preguiça, Tânia Martins-Marques, Tânia Ribeiro, Henrique Girão, Artur Figueirinha, Lígia Salgueiro, and et al. 2024. "Effect of Blueberry Supplementation on a Diet-Induced Rat Model of Prediabetes—Focus on Hepatic Lipid Deposition, Endoplasmic Stress Response and Autophagy" Nutrients 16, no. 4: 513. https://doi.org/10.3390/nu16040513
APA StyleFerreira, G., Vieira, P., Alves, A., Nunes, S., Preguiça, I., Martins-Marques, T., Ribeiro, T., Girão, H., Figueirinha, A., Salgueiro, L., Pintado, M., Gomes, P., Viana, S., & Reis, F. (2024). Effect of Blueberry Supplementation on a Diet-Induced Rat Model of Prediabetes—Focus on Hepatic Lipid Deposition, Endoplasmic Stress Response and Autophagy. Nutrients, 16(4), 513. https://doi.org/10.3390/nu16040513