High Dose Fish Oil Added to Various Lipid Emulsions Normalizes Superoxide Dismutase 1 Activity in Home Parenteral Nutrition Patients

 , , ,
, , ,
Abstract
1. Introduction
2. Material and Methods
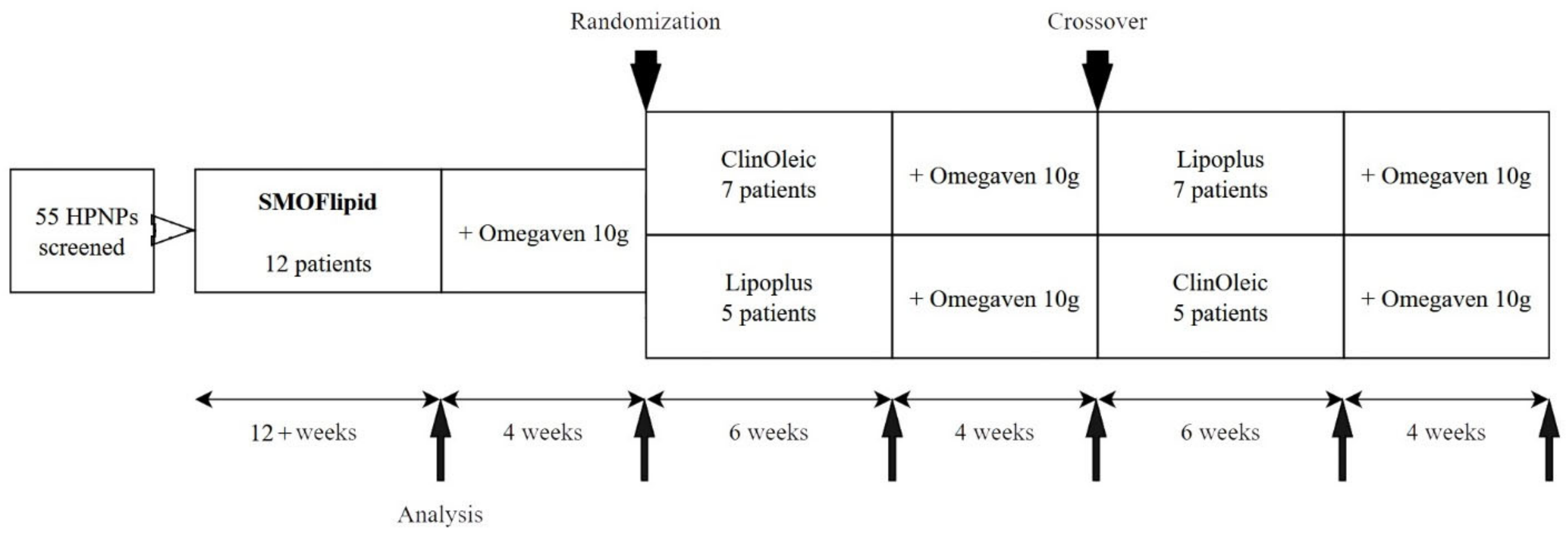
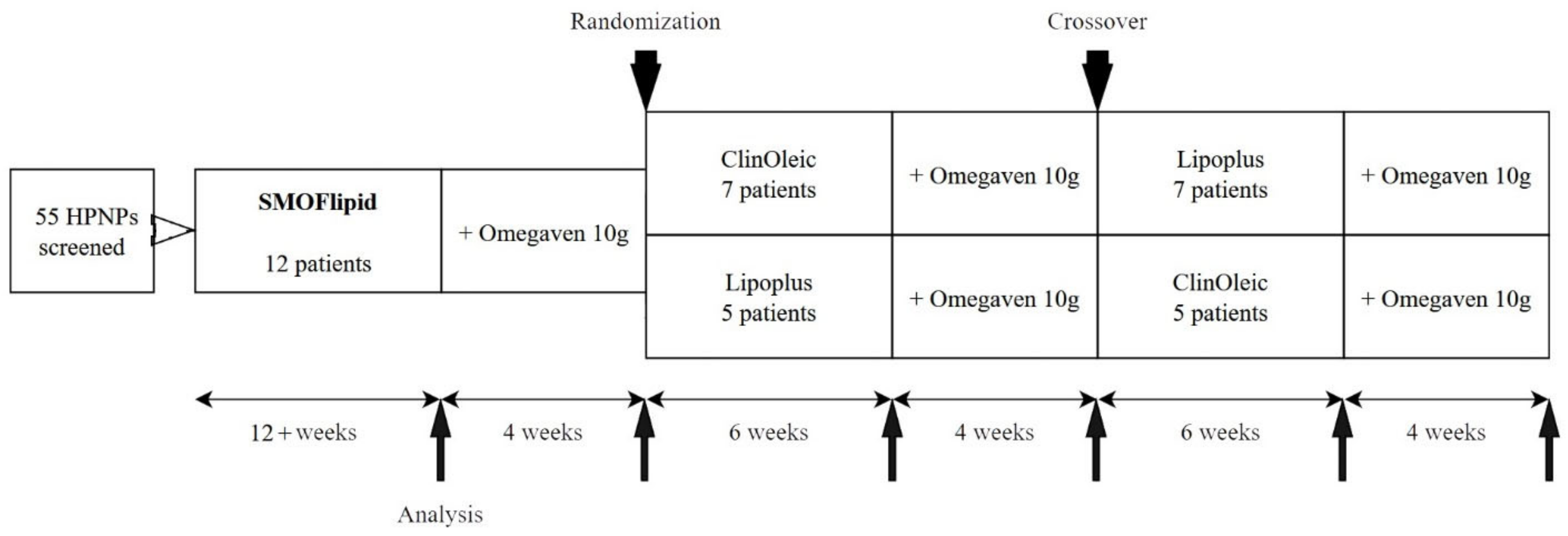
2.1. Design and Participants
2.2. FGF-19, Cholesten, and Bile Acid Measurement
2.3. Antioxidant Enzymes
2.4. Fatty Acid Analyses
2.5. Statistical Analyses
3. Results
3.1. Population Characteristics
3.2. Biochemical Parameters
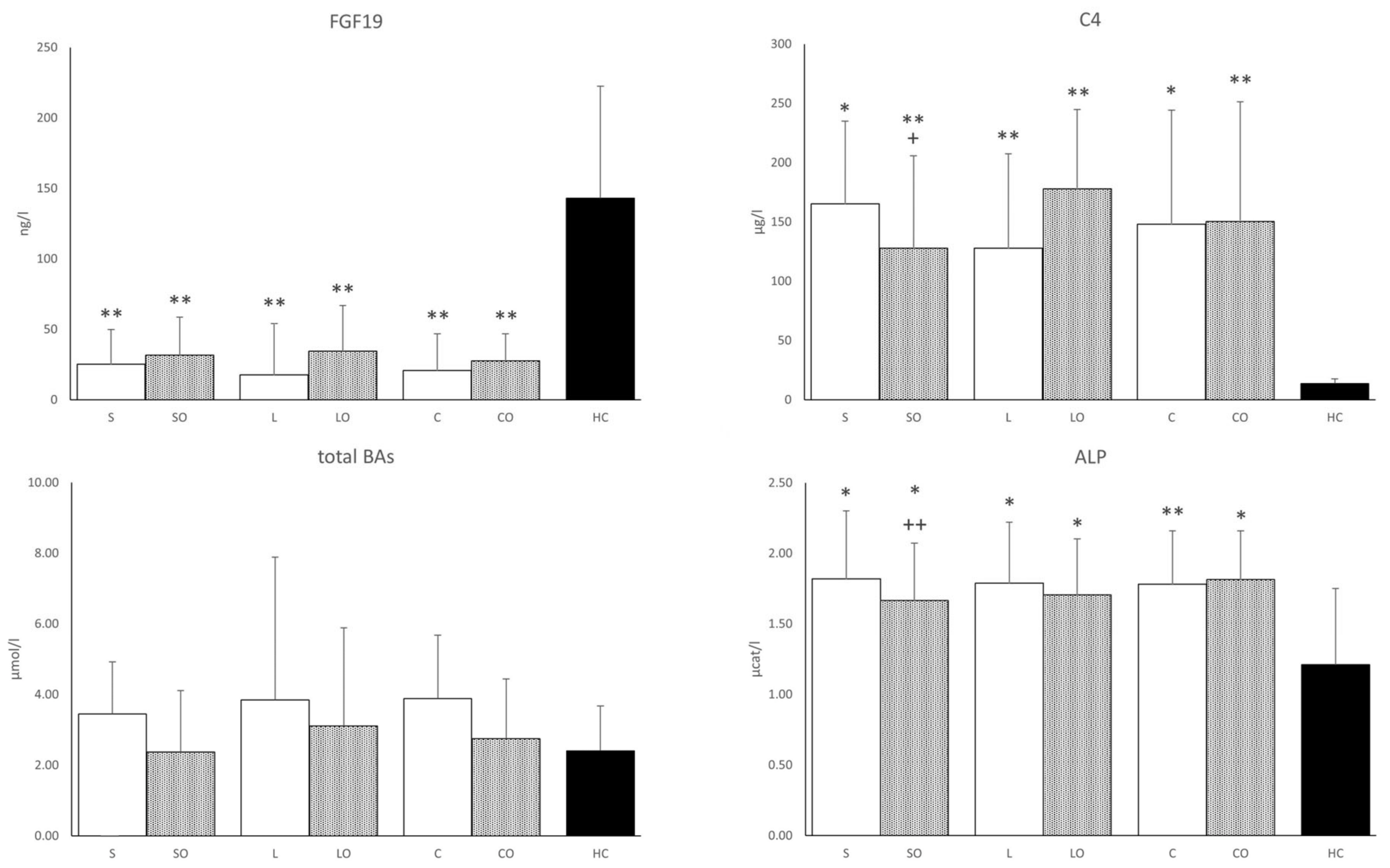
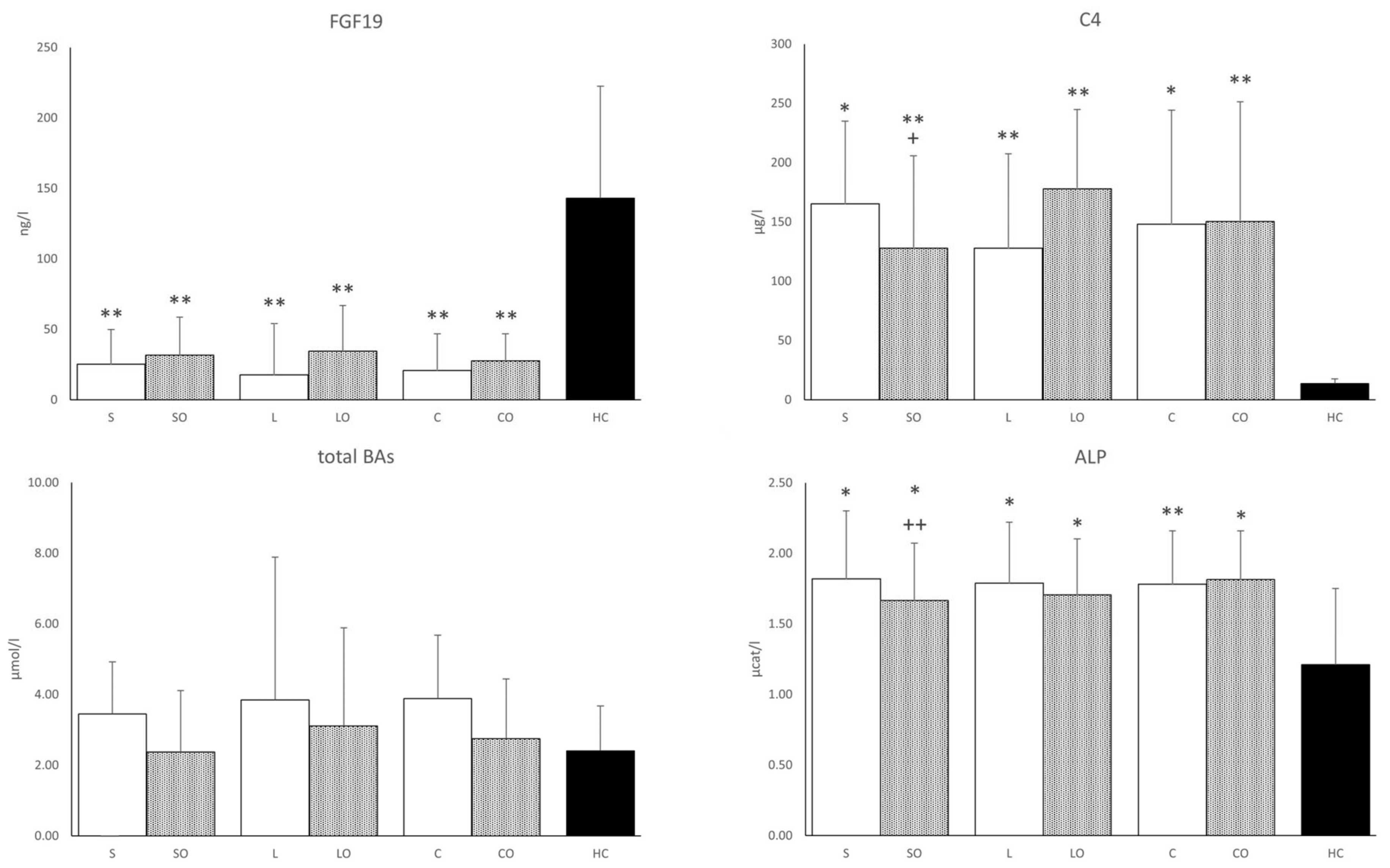
3.3. FGF19, Bile Acids, and Liver Function Tests
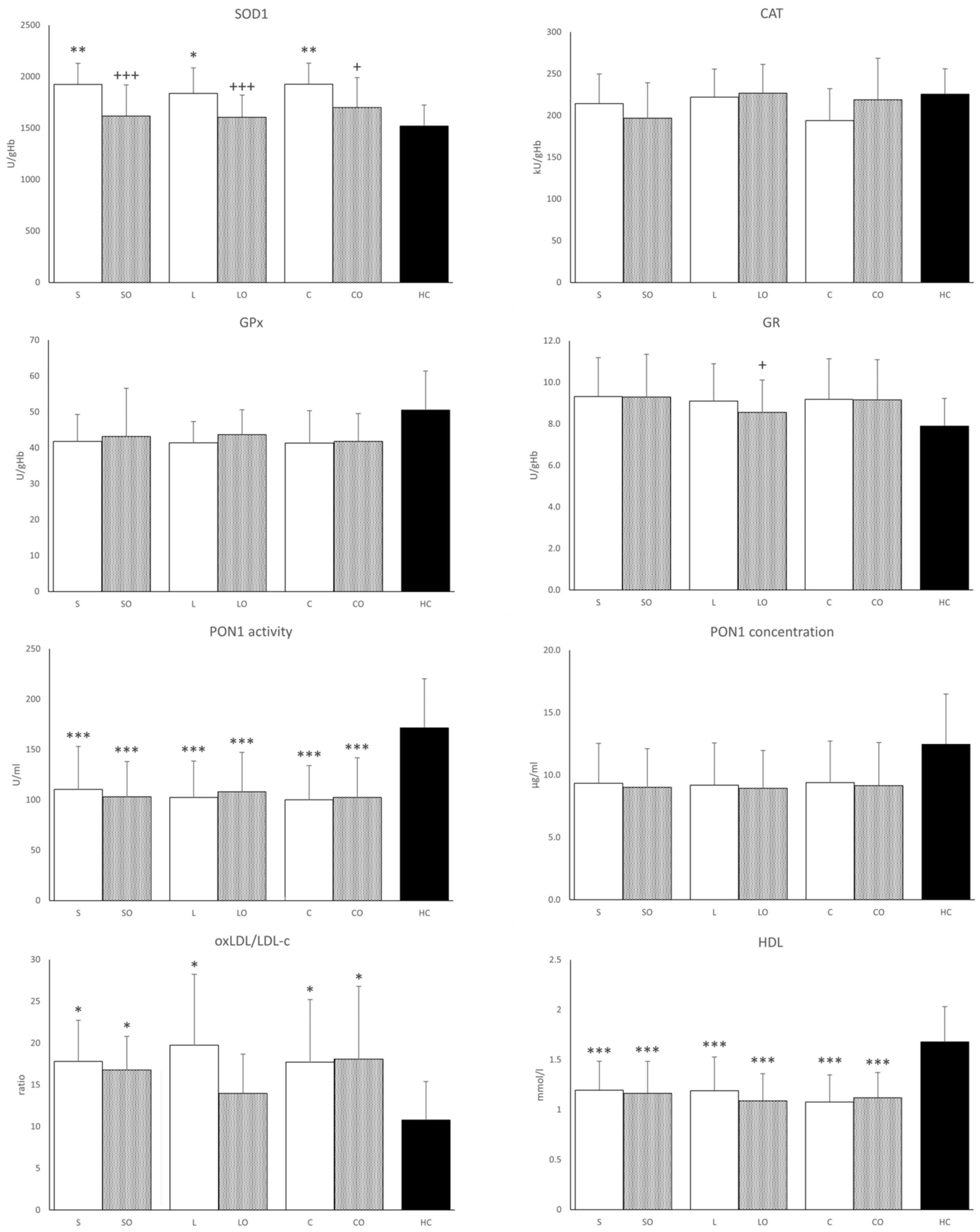
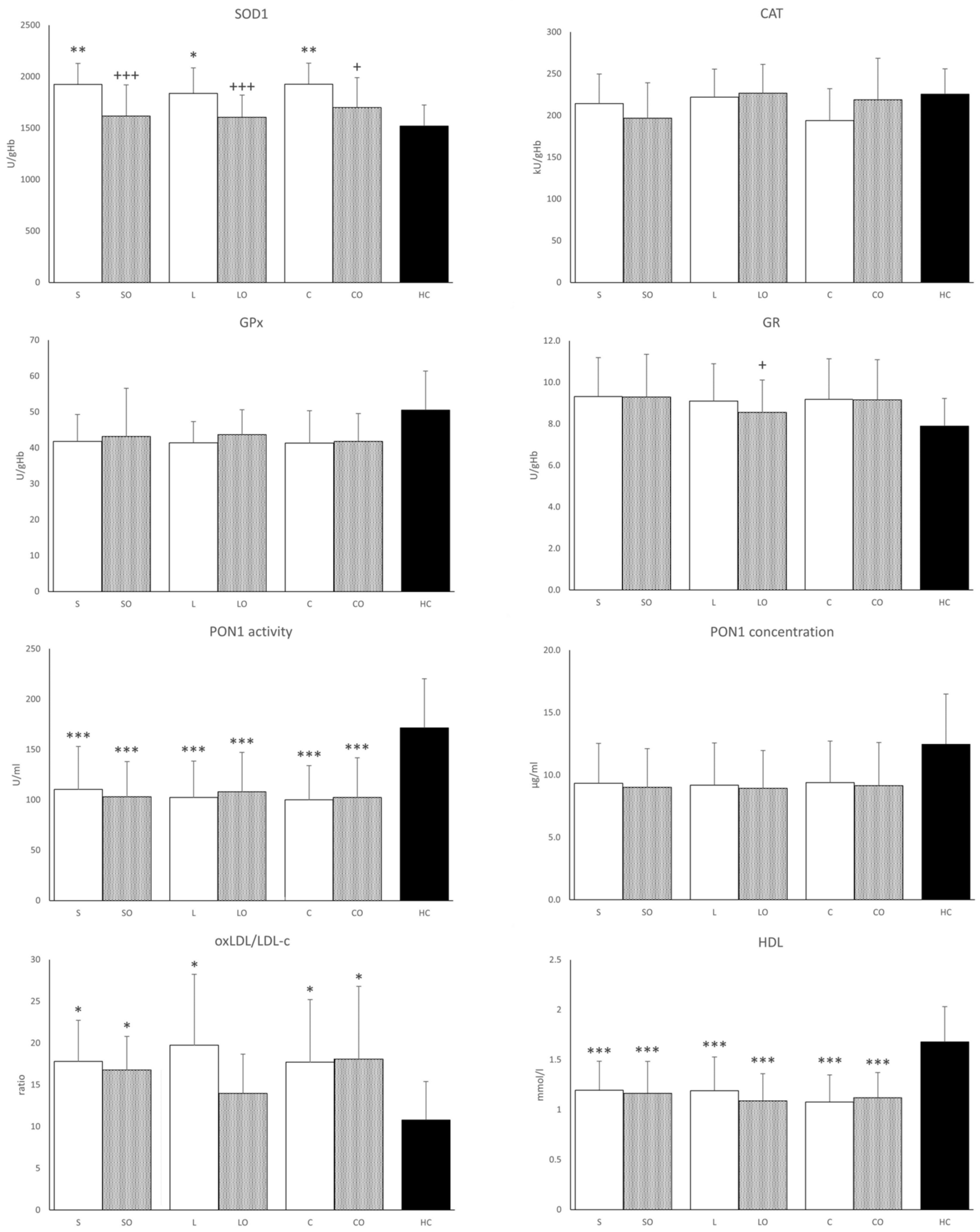
3.4. Antioxidant Enzymes
3.5. Fatty Acid Proportion in Plasma Phospholipids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cuerda, C.; Pironi, L.; Arends, J.; Bozzetti, F.; Gillanders, L.; Jeppesen, P.B.; Joly, F.; Kelly, D.; Lal, S.; Staun, M.; et al. ESPEN practical guideline: Clinical nutrition in chronic intestinal failure. Clin. Nutr. 2021, 40, 5196–5220. [Google Scholar] [CrossRef] [PubMed]
- Novak, F.; Vecka, M.; Meisnerova, E.; Sevela, S.; Vavrova, L.; Rychlikova, J.; Dolezalova, L.; Myslivcova, D.; Zak, A.; Vitek, L.; et al. Fish oil supplementation with various lipid emulsions suppresses in vitro cytokine release in home parenteral nutrition patients: A crossover study. Nutr. Res. 2019, 72, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Kosek, V.; Heczkova, M.; Novak, F.; Meisnerova, E.; Novakova, O.; Zelenka, J.; Bechynska, K.; Vrzacova, N.; Suttnar, J.; Hlavackova, A.; et al. The omega-3 Polyunsaturated Fatty Acids and Oxidative Stress in Long-Term Parenteral Nutrition Dependent Adult Patients: Functional Lipidomics Approach. Nutrients 2020, 12, 2351. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xiao, Y.; Liu, Y.; Tian, X.; Wang, W.; Jiang, L.; Wu, W.; Zhang, T.; Cai, W.; Wang, Y. Fish oil-based lipid emulsion alleviates parenteral nutrition-associated liver diseases and intestinal injury in piglets. J. Parenter. Enteral. Nutr. 2022, 46, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.G.; Ng, K.; Stoll, B.; De Pipaón, M.S. Impact of new-generation lipid emulsions on cellular mechanisms of parenteral nutrition-associated liver disease. Adv. Nutr. 2014, 5, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Vogt, M.; Bauer, M.K.; Ferrari, D.; Schulze-Osthoff, K. Oxidative stress and hypoxia/reoxygenation trigger CD95 (APO-1/Fas) ligand expression in microglial cells. FEBS Lett. 1998, 429, 67–72. [Google Scholar] [CrossRef]
- Guthrie, G.; Burrin, D. Impact of Parenteral Lipid Emulsion Components on Cholestatic Liver Disease in Neonates. Nutrients 2021, 13, 508. [Google Scholar] [CrossRef]
- Pironi, L.; Sasdelli, A.S. Intestinal Failure-Associated Liver Disease. Clin. Liver Dis. 2019, 23, 279–291. [Google Scholar] [CrossRef]
- Bond, A.; Huijbers, A.; Pironi, L.; Schneider, S.M.; Wanten, G.; Lal, S. Review article: Diagnosis and management of intestinal failure-associated liver disease in adults. Aliment. Pharmacol. Ther. 2019, 50, 640–653. [Google Scholar] [CrossRef]
- Secor, J.D.; Yu, L.; Tsikis, S.; Fligor, S.; Puder, M.; Gura, K.M. Current strategies for managing intestinal failure-associated liver disease. Expert. Opin. Drug Saf. 2021, 20, 307–320. [Google Scholar] [CrossRef]
- Mutanen, A.; Lohi, J.; Heikkila, P.; Jalanko, H.; Pakarinen, M.P. Loss of ileum decreases serum fibroblast growth factor 19 in relation to liver inflammation and fibrosis in pediatric onset intestinal failure. J. Hepatol. 2015, 62, 1391–1397. [Google Scholar] [CrossRef]
- van Erpecum, K.J.; Schaap, F.G. Intestinal failure to produce FGF19: A culprit in intestinal failure-associated liver disease? J. Hepatol. 2015, 62, 1231–1233. [Google Scholar] [CrossRef]
- Stofan, M.; Guo, G.L. Bile Acids and FXR: Novel Targets for Liver Diseases. Front. Med. 2020, 7, 544. [Google Scholar] [CrossRef]
- Lucchinetti, E.; Lou, P.H.; Wawrzyniak, P.; Wawrzyniak, M.; Scharl, M.; Holtzhauer, G.A.; Kramer, S.D.; Hersberger, M.; Rogler, G.; Zaugg, M. Novel Strategies to Prevent Total Parenteral Nutrition-Induced Gut and Liver Inflammation, and Adverse Metabolic Outcomes. Mol. Nutr. Food Res. 2021, 65, e1901270. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qin, C.; Dong, L.; Zhang, X.; Wu, Z.; Liu, L.; Yang, J.; Liu, L. Whole grain benefit: Synergistic effect of oat phenolic compounds and β-glucan on hyperlipidemia via gut microbiota in high-fat-diet mice. Food Funct. 2022, 13, 12686–12696. [Google Scholar] [CrossRef] [PubMed]
- Madnawat, H.; Welu, A.L.; Gilbert, E.J.; Taylor, D.B.; Jain, S.; Manithody, C.; Blomenkamp, K.; Jain, A.K. Mechanisms of Parenteral Nutrition-Associated Liver and Gut Injury. Nutr. Clin. Pract. 2020, 35, 63–71. [Google Scholar] [CrossRef]
- Wawrzyniak, P.; Noureddine, N.; Wawrzyniak, M.; Lucchinetti, E.; Krämer, S.D.; Rogler, G.; Zaugg, M.; Hersberger, M. Nutritional Lipids and Mucosal Inflammation. Mol. Nutr. Food Res. 2021, 65, e1901269. [Google Scholar] [CrossRef] [PubMed]
- Rogulska, J.; Osowska, S.; Kunecki, M.; Sobocki, J.; Ładyżyński, P.; Giebułtowicz, J. Antioxidant balance in plasma of patients on home parenteral nutrition: A pilot study comparing three different lipid emulsions. Clin. Nutr. 2021, 40, 3950–3958. [Google Scholar] [CrossRef]
- Zaloga, G.P. Narrative Review of n-3 Polyunsaturated Fatty Acid Supplementation upon Immune Functions, Resolution Molecules and Lipid Peroxidation. Nutrients 2021, 13, 662. [Google Scholar] [CrossRef]
- Lenicek, M.; Vecka, M.; Zizalova, K.; Vitek, L. Comparison of simple extraction procedures in liquid chromatography-mass spectrometry based determination of serum 7α-hydroxy-4-cholesten-3-one, a surrogate marker of bile acid synthesis. J. Chromatogr. B Analyt Technol. Biomed. Life Sci. 2016, 1033–1034, 317–320. [Google Scholar] [CrossRef]
- Vavrova, L.; Rychlikova, J.; Mrackova, M.; Novakova, O.; Zak, A.; Novak, F. Increased inflammatory markers with altered antioxidant status persist after clinical recovery from severe sepsis: A correlation with low HDL cholesterol and albumin. Clin. Exp. Med. 2016, 16, 557–569. [Google Scholar] [CrossRef]
- Kodydková, J.; Vávrová, L.; Zeman, M.; Jirák, R.; Macásek, J.; Stanková, B.; Tvrzická, E.; Zák, A. Antioxidative enzymes and increased oxidative stress in depressive women. Clin. Biochem. 2009, 42, 1368–1374. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Vecka, M.; Tvrzická, E.; Stanková, B.; Zák, A. Effect of column and software on gas chromatographic determination of fatty acids. J. Chromatogr. B Analyt Technol. Biomed. Life Sci. 2002, 770, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Klek, S.; Chambrier, C.; Singer, P.; Rubin, M.; Bowling, T.; Staun, M.; Joly, F.; Rasmussen, H.; Strauss, B.J.; Wanten, G.; et al. Four-week parenteral nutrition using a third generation lipid emulsion (SMOFlipid)—A double-blind, randomised, multicentre study in adults. Clin. Nutr. 2013, 32, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Sands, S.A.; Windsor, S.L.; Ali, H.A.; Stevens, T.L.; Magalski, A.; Porter, C.B.; Borkon, A.M. Omega-3 fatty acids in cardiac biopsies from heart transplantation patients: Correlation with erythrocytes and response to supplementation. Circulation 2004, 110, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Heshmati, J.; Morvaridzadeh, M.; Maroufizadeh, S.; Akbari, A.; Yavari, M.; Amirinejad, A.; Maleki-Hajiagha, A.; Sepidarkish, M. Omega-3 fatty acids supplementation and oxidative stress parameters: A systematic review and meta-analysis of clinical trials. Pharmacol. Res. 2019, 149, 104462. [Google Scholar] [CrossRef]
- Liu, L.; Jin, R.; Hao, J.; Zeng, J.; Yin, D.; Yi, Y.; Zhu, M.; Mandal, A.; Hua, Y.; Ng, C.K.; et al. Consumption of the Fish Oil High-Fat Diet Uncouples Obesity and Mammary Tumor Growth through Induction of Reactive Oxygen Species in Protumor Macrophages. Cancer Res. 2020, 80, 2564–2574. [Google Scholar] [CrossRef] [PubMed]
- Miloudi, K.; Comte, B.; Rouleau, T.; Montoudis, A.; Levy, E.; Lavoie, J.C. The mode of administration of total parenteral nutrition and nature of lipid content influence the generation of peroxides and aldehydes. Clin. Nutr. 2012, 31, 526–534. [Google Scholar] [CrossRef]
- Shimizu, M.; Sato, R. Endocrine Fibroblast Growth Factors in Relation to Stress Signaling. Cells 2022, 11, 505. [Google Scholar] [CrossRef]
- Vakili, L.; Navab, K.D.; Shabihkhani, M.; Pourtabatabaei, N.; Vazirian, S.; Barseghian, Z.; Seyedali, S.; Hough, G. Systemic inflammation, intestine, and paraoxonase-1. Adv. Exp. Med. Biol. 2014, 824, 83–88. [Google Scholar] [CrossRef]
- Boehm, D.; Krzystek-Korpacka, M.; Neubauer, K.; Matusiewicz, M.; Berdowska, I.; Zielinski, B.; Paradowski, L.; Gamian, A. Paraoxonase-1 status in Crohn’s disease and ulcerative colitis. Inflamm. Bowel Dis. 2009, 15, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, G.; Bacchetti, T. Effect of dietary lipids on paraoxonase-1 activity and gene expression. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, B.; Volkova, N.; Aviram, M. Postprandial serum triacylglycerols and oxidative stress in mice after consumption of fish oil, soy oil or olive oil: Possible role for paraoxonase-1 triacylglycerol lipase-like activity. Nutrition 2006, 22, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Freese, R.; Alfthan, G.; Jauhiainen, M.; Basu, S.; Erlund, I.; Salminen, I.; Aro, A.; Mutanen, M. High intakes of vegetables, berries, and apples combined with a high intake of linoleic or oleic acid only slightly affect markers of lipid peroxidation and lipoprotein metabolism in healthy subjects. Am. J. Clin. Nutr. 2002, 76, 950–960. [Google Scholar] [CrossRef] [PubMed]
- Boutte, H.J., Jr.; Chen, J.; Wylie, T.N.; Wylie, K.M.; Xie, Y.; Geisman, M.; Prabu, A.; Gazit, V.; Tarr, P.I.; Levin, M.S.; et al. Fecal microbiome and bile acid metabolome in adult short bowel syndrome. Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 322, G154–G168. [Google Scholar] [CrossRef] [PubMed]
- Poupon, R. Liver alkaline phosphatase: A missing link between choleresis and biliary inflammation. Hepatology 2015, 61, 2080–2090. [Google Scholar] [CrossRef] [PubMed]
- Pike, A.F.; Kramer, N.I.; Blaauboer, B.J.; Seinen, W.; Brands, R. A novel hypothesis for an alkaline phosphatase ‘rescue’ mechanism in the hepatic acute phase immune response. Biochim. Biophys. Acta 2013, 1832, 2044–2056. [Google Scholar] [CrossRef]
- Tuin, A.; Huizinga-Van der Vlag, A.; van Loenen-Weemaes, A.M.; Meijer, D.K.; Poelstra, K. On the role and fate of LPS-dephosphorylating activity in the rat liver. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G377–G385. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| HPNPs | HCs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sex F/M | Age (years) | BMI | Infusions/Week | Infusion a (mL) | Energy b (kJ) | Enteral c (%) | Classification, IF d SBS/Other | Sex F/M | Age (years) | BMI |
| 8/4 | 58.3 ± 17.6 e | 23.4 ± 3.7 | 5.8 ± 1.0 | 2910 ± 1200 | 72 ± 32 | 40 ± 16 | 10/2 | 8/4 | 61 ± 17 | 25.1 ± 3.0 |
| Smoflipid | Lipoplus | ClinOleic | Healthy Controls | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Baseline | p | +Omegaven | p | Baseline | p | +Omegaven | p | Baseline | p | +Omegaven | p | ||
| TAG | (mmol/L) | 1.21 ± 0.44 | § 0.0297 | 1.21 ± 0.5 | 1.34 ± 0.6 | 1.54 ± 0.8 | 1.56 ± 0.6 | 1.43 ± 0.6 | 1.16 ± 0.5 | |||||
| TC | (mmol/L) | 4.30 ± 1.0 | * 0.0138 | 4.36 ± 1.0 | * 0.0039 | 4.22 ± 1.0 | * 0.0119 | * 0.0039 | * 0.0083 | 3.98 ± 1.0 | * 0.0039 | 4.12 ± 0.8 | * 0.0081 | 5.60 ± 1.1 |
| LDL-C | (mmol/L) | 2.57 ± 0.9 | 2.65 ± 1.0 | 2.43 ± 0.9 | 2.54 ± 1.0 | 2.20 ± 0.9 | 2.41 ± 0.8 | 3.4 ± 1.0 | ||||||
| Apo A1 | (g/L) | 1.43 ± 0.3 | 1.28 ± 0.3 | * 0.0002 | 1.29 ± 0.4 | * 0.0002 | 1.26 ± 0.4 | * 0.0002 | 1.31 ± 0.3 | * 0.0001 | 1.19 ± 0.3 | * 0.0001 | 1.67 ± 0.3 | |
| Apo B | (g/L) | 0.91 ± 0.34 | 0.87 ± 0.35 | * 0.0002 | 0.89 ± 0.33 | * 0.0002 | 0.93 ± 0.41 | * 0.0002 | 0.69 ± 0.24 | * 0.0001 | 1.07 ± 0.46 | * 0.0002 | 1.09 ± 0.39 | |
| Lp(a) | (g/L) | 0.21 ± 0.21 | 0.21 ± 0.19 | 0.3 ± 0.29 | 0.28 ± 0.23 | 0.27 ± 0.25 | 0.2 ± 0.16 | 0.14 ± 0.27 | ||||||
| SAA | (mg/L) | 24.80 ± 15.6 | 33.57 ± 18.6 | 25.55 ± 20.4 | 27.98 ± 14.2 | 31.50 ± 21.0 | 34.12 ± 15.6 | 19.96 ± 24.3 | ||||||
| HCY | (μmol/mL) | 13.3 ± 3.4 | 14.2 ± 4.6 | 13 ± 4 | 13.6 ± 4.9 | 13.5 ± 2.8 | 14.8 ± 4.6 | 13 ± 2.5 | ||||||
| Ca | (mmol/L) | 2.24 ± 0.1 | * 0.0285 | 2.20 ± 0.1 | * 0.0103 | 2.24 ± 0.1 | * 0.0372 | 2.24 ± 0.1 | * 0.0228 | 2.22 ± 0.1 | * 0.0221 | 2.27 ± 0.1 | * 0.0437 | 2.37 ± 0.06 |
| Fe | (μmol/L) | 14.22 ± 7.47 | § 0.043 | 13.72 ± 5.32 | 15.09 ± 7.58 | 15.61 ± 6.5 | 17.89 ± 10.25 | 14.37 ± 6.5 | 22.2 ± 10.1 | |||||
| Transferrin | (mg/L) | 3.47 ± 0.81 | * 0.0204 | 3.45 ± 0.85 | * 0.0102 | 3.46 ± 0.78 | * 0.0122 | 3.5 ± 0.79 | * 0.0148 | 3.31 ± 0.74 | * 0.0201 | 3.25 ± 0.79 | * 0.0122 | 2.45 ± 0.34 |
| Ferritin | (μg/L) | 151 ± 220.2 | 130.7 ± 167.9 | 145.7 ± 185.6 | 152.5 ± 185.3 | 152.8 ± 232.2 | 220.1 ± 223.4 | 94.5 ± 69 | ||||||
| Hemoglobin | (g/L) | 131 ± 11 | 130 ± 13 | 132 ± 11 | 131 ± 15 | 128 ± 14 | 131 ± 14 | 144 ± 9 | ||||||
| Smoflipid | Lipoplus | ClinOleic | Healthy Controls | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Baseline | p | +Omegaven | p | Baseline | p | +Omegaven | p | Baseline | p | +Omegaven | p | |||
| Sum of primary BAs | (μmol/L) | 3.96 [2.94; 4.9] | 2.4 [2.14; 4.63] | 4.42 [3.14; 6.63] | 3.65 [2.39; 5.02] | 4.18 [2.63; 6.33] | 3.8 [2.4; 5] | 1.48 [1.3; 3.25] | ||||||
| Sum of secondary BAs | (μmol/L) | 0.45 [0.17; 0.94] | 0.31 [0.17; 0.94] | 0.55 [0.17; 1.44] | 0.43 [0.17; 0.97] | 0.38 [0.17; 0.92] | 0.23 [0.17; 0.85] | 0.65 [0.44; 1.17] | ||||||
| Primary/secondary BAs ratio | 6.6 [2.9; 30.6] | * 0.033 | 6.6 [2.7; 32] | * 0.106 | 6.9 [2.7; 31.4] | * 0.054 | 9.2 [2.3; 53.1] | * 0.039 | 7.8 [3.2; 47.7] | * 0.007 | 9.1 [3; 31.7] | * 0.014 | 2.7 [1.5; 4.8] | |
| Conjugated/unconjugated BAs ratio | 13.6 [0.1;51.8] | 7.4 [0.1; 48.4] | 5.9 [0.1; 43.3] | 12.4 [0.1; 91.6] | 2.5 [0.3; 66] | 7 [0.1; 59] | 5.4 [0.9; 16.5] | |||||||
| ALT | (μcat/L) | 0.61 ± 0.35 | 0.7 ± 0.44 | 0.89 ± 0.75 | 0.9 ± 0.81 | 0.91 ± 0.94 | 0.69 ± 0.43 | 0.36 ± 0.08 | ||||||
| AST | (μcat/L) | 0.48 ± 0.17 | 0.53 ± 0.19 | 0.58 ± 0.22 | 0.6 ± 0.27 | 0.61 ± 0.34 | 0.55 ± 0.14 | 0.41 ± 0.07 | ||||||
| GGT | (μcat/L) | 0.6 ± 0.54 | 0.65 ± 0.54 | 0.73 ± 0.57 | 0.74 ± 0.44 | 0.82 ± 0.72 | 0.83 ± 0.48 | 0.31 ± 0.19 | ||||||
| Total bilirubin | (μmol/L) | 13.3 ± 9.8 | 15.6 ± 13.2 | 17.9 ± 20.4 | 18.3 ± 17.7 | 19.4 ± 24.2 | 16.6 ± 14.3 | 10.4 ± 4.5 | ||||||
| FGF19 vs. SOD1 c | 0.82 | # 0.001 | 0.38 | # 0.228 | 0.7 | # 0.011 | 0.47 | # 0.88 | 0.76 | # 0.004 | 0.39 | # 0.211 | ||
| ALP vs. C4 c | 0.43 | # 0.166 | 0.47 | # 0.122 | 0.79 | # 0.002 | 0.58 | # 0.047 | 0.72 | # 0.008 | 0.71 | # 0.009 | ||
| ALP vs. p/s BAs c | 0.65 | # 0.021 | 0.57 | # 0.053 | 0.51 | # 0.088 | 0.64 | # 0.026 | 0.69 | # 0.012 | 0.63 | # 0.028 | ||
| Smoflipid | Lipoplus | ClinOleic | Healthy Controls | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acids, Mol% | Baseline | p | +Omegaven | p | Baseline | p | +Omegaven | p | Baseline | p | +Omegaven | p | |
| 16:00 | 31.40 ± 1.61 | * 0.0002 | 31.52 ± 2.8 | * 0.0002 | 30.61 ± 1.2 | * 0.0022 | 30.27 ± 1.6 | * 0.0061 | 30.27 ± 1.7 | * 0.0033 | 29.97 ± 0.95 | * 0.0043 | 27.89 ± 1.6 |
| 16:1n-7 | 0.61 ± 0.2 | 0.62 ± 0.2 | 0.64 ± 0.3 | 0.63 ± 0.3 | 0.65 ± 0.3 | 0.61 ± 0.2 | 0.59 ± 0.2 | ||||||
| 18:00 | 12.70 ± 1.7 | 12.92 ± 1.8 | 12.91 ± 1.4 | 13.28 ± 1.42 | 12.73 ± 1.2 | 12.83 ± 1.0 | 13.88 ± 1.2 | ||||||
| 18:1n-9 | 11.5 ± 1.2 | 11.28 ± 1.5 | 11.73 ± 2.0 | 11.42 ± 2.3 | 11.89 ± 1.7 | 12.65 ± 1.9 | * 0.0286 | 10.38 ± 1.1 | |||||
| 18:2n-6 | 16.67 ± 2.2 | * 0.0006 | 15.13 ± 2.2 | * 0.0001; | 17.76 ± 3.3 | * 0.0013 | 16.57 ± 3.6 | * 0.0007 | 17.38 ± 3.1 | * 0.0013 | 16.92 ± 3.0 | * 0.0008 | 21.84 ± 3.1 |
| + 0.033 | + 0.0356 | ||||||||||||
| 18:3n-3 | 0.16 ± 0.04 | * 0.0199 | 0.15 ± 0.1 | * 0.0095 | 0.16 ± 0.04 | * 0.0284 | 0.17 ± 0.1 | * 0.0369 | 0.18 ± 0.06 | * 0.0391 | 0.14 ± 0.01 | * 0.0047 | 0.22 ± 0.04 |
| 20:3n-6 | 2.75 ± 1.1 | § 0.0093 | 2.08 ± 1.0 | * 0.0327 | 2.83 ± 1.1 | 2.14 ± 0.8 | * 0.0344 | 3.69 ± 1.3 | 2.16 ± 0.8 | * 0.0261 | 3.284 ± 0.6 | ||
| + 0.0083 | + 0.0191 | + 0.0004 | |||||||||||
| 20:4n-6 | 8.66 ± 2.0 | * 0.0002 | 7.20 ± 1.7 | * 0.0001 | 8.36 ± 1.5 | * 0.0002 | 7.48 ± 1.6 | * 0.0001 | 9.57 ± 2.1 | * 0.0016 | 7.64 ± 1.6 | * 0.0001 | 11.89 ± 1.4 |
| + 0.00001 | + 0.0002 | + 0.0004 | |||||||||||
| 20:5n-3 | 3.34 ± 1.0 | * 0.0021 | 5.18 ± 1.8 | * 0.0001 | 3.08 ± 1.5 | * 0.0041 | 4.96 ± 2.07 | * 0.0001 | 2.18 ± 1.0 | 4.55 ± 2.0 | * 0.0001 | 1.1 ± 0.4 | |
| § 0.0042 | + 0.0002 | + 0.0001 | + 0.0003 | ||||||||||
| 22:5n-3 | 1.52 ± 0.3 | * 0.0037 | 1.66 ± 0.4 | * 0.0003 | 1.475 ± 0.3 | * 0.0072 | 1.53 ± 0.4 | * 0.0044 | 1.37 ± 0.3 | * 0.0342 | 1.26 ± 0.23 | + 0.0043 | 1.038 ± 0.07 |
| 22:6n-3 | 6.98 ± 1.3 | * 0.0002 | 8.70 ± 2.9 | * 0.0001 | 6.68 ± 1.4 | * 0.0002 | 7.93 ± 1.5 | * 0.0001 | 6.09 ± 1.9 | * 0.0009 | 7.4 ± 1.5 | * 0.0001 | 3.58 ± 1.0 |
| + 0.0001 | + 0.001 | ||||||||||||
| Ʃ SFA | 45.22 ± 1.4 | * 0.007 | 45.66 ± 2.6 | * 0.0012 | 44.43 ± 2.1 | 44.48 ± 1.6 | 43.89 ± 1.2 | 43.69 ± 0.8 | 42.83 ± 0.9 | ||||
| Ʃ MUFA | 14.19 ± 1.38 | 13.91 ± 1.62 | 14.45 ± 2.46 | 14.11 ± 2.54 | 14.74 ± 1.88 | 15.47 ± 1.92 | * 0.0322 | 12.96 ± 1.2 | |||||
| Ʃ n-6 PUFA | 28.81 ± 2.2 | * 0.0001 | 25.13 ± 3.4 | * 0.0001 | 29.81 ± 3.2 | * 0.0001 | 26.92 ± 3.6 | * 0.0001 | 31.61 ± 3.0 | * 0.0001 | 27.53 ± 3.7 | * 0.0001 | 38.16 ± 2.2 |
| § 0.0025 | + 0.0001 | + 0.00001 | + 0.0001 | ||||||||||
| Ʃ n-3 PUFA | 11.78 ± 2.2 | * 0.0002 | 15.3 ± 4.2 | * 0.0001 | 11.31 ± 2.1 | * 0.0002 | 14.49 ± 2.9 | * 0.0001 | 9.76 ± 2.8 | * 0.0019 | 13.31 ± 3.1 | * 0.0001 | 6.05 ± 1.4 |
| § 0.0082 | + 0.0062 | + 0.00001 | + 0.00001 | ||||||||||
| n-6 + n-3 | 40.58 ± 2.0 | * 0.0028 | 40.43 ± 2.9 | * 0.0022 | 41.11 ± 2.5 | * 0.0073 | 41.41 ± 2.6 | * 0.0036 | 41.37 ± 2.2 | * 0.0088 | 40.84 ± 2.1 | * 0.0046 | 44.21 ± 1.4 |
| SFA+MUFA/PUFA | 1.46 ± 0.12 | * 0.0058 | 1.48 ± 0.17 | * 0.0034 | 1.42 ± 0.13 | * 0.0124 | 1.45 ± 0.13 | * 0.0153 | 1.44 ± 0.14 | * 0.0058 | 1.42 ± 0.15 | * 0.0089 | 1.26 ± 0.07 |
| 20:4/20:5 | 2.88 ± 1.2 | * 0.0001 | 1.57 ± 0.7 | * 0.0001 | 3.35 ± 1.6 | * 0.0001 | 1.71 ± 0.6 | * 0.0001 | 5.44 ± 2.8 | * 0.0001 | 2.03 ± 1.0 | * 0.0001 | 3.56 ± 1.4 |
| § 0.0052 | + 0.0001 | + 0.0016 | + 0.0006 | ||||||||||
| 20:4/22:6 | 1.27 ± 0.4 | * 0.0001 | 0.9 ± 0.3 | * 0.0001 | 1.3 ± 0.4 | * 0.0001 | 0.96 ± 0.2 | * 0.0001 | 1.77 ± 0.9 | * 0.0001 | 1.07 ± 0.3 | * 0.0001 | 3.52 ± 0.9 |
| § 0.0149 | + 0.0001 | + 0.0002 | + 0.0028 | ||||||||||
| 22:6/18:3 | 31.42 ± 15.1 | * 0.0101 | 44.98 ± 24.0 | * 0.0001 | 29.99 ± 16.6 | * 0.0105 | 35.74 ± 16.3 | * 0.0027 | 22.73 ± 12.7 | 34.01 ± 10.0 | * 0.0045 | 11.46 ± 3.6 | |
| § 0.0009 | + 0.0005 | ||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sevela, S.; Meisnerova, E.; Vecka, M.; Vavrova, L.; Rychlikova, J.; Lenicek, M.; Vitek, L.; Novakova, O.; Novak, F. High Dose Fish Oil Added to Various Lipid Emulsions Normalizes Superoxide Dismutase 1 Activity in Home Parenteral Nutrition Patients. Nutrients 2024, 16, 485. https://doi.org/10.3390/nu16040485
Sevela S, Meisnerova E, Vecka M, Vavrova L, Rychlikova J, Lenicek M, Vitek L, Novakova O, Novak F. High Dose Fish Oil Added to Various Lipid Emulsions Normalizes Superoxide Dismutase 1 Activity in Home Parenteral Nutrition Patients. Nutrients. 2024; 16(4):485. https://doi.org/10.3390/nu16040485
Chicago/Turabian StyleSevela, Stanislav, Eva Meisnerova, Marek Vecka, Lucie Vavrova, Jana Rychlikova, Martin Lenicek, Libor Vitek, Olga Novakova, and Frantisek Novak. 2024. "High Dose Fish Oil Added to Various Lipid Emulsions Normalizes Superoxide Dismutase 1 Activity in Home Parenteral Nutrition Patients" Nutrients 16, no. 4: 485. https://doi.org/10.3390/nu16040485
APA StyleSevela, S., Meisnerova, E., Vecka, M., Vavrova, L., Rychlikova, J., Lenicek, M., Vitek, L., Novakova, O., & Novak, F. (2024). High Dose Fish Oil Added to Various Lipid Emulsions Normalizes Superoxide Dismutase 1 Activity in Home Parenteral Nutrition Patients. Nutrients, 16(4), 485. https://doi.org/10.3390/nu16040485