
A Narrative Review on the Neuroprotective Potential of Brown Macroalgae in Alzheimer’s Disease

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
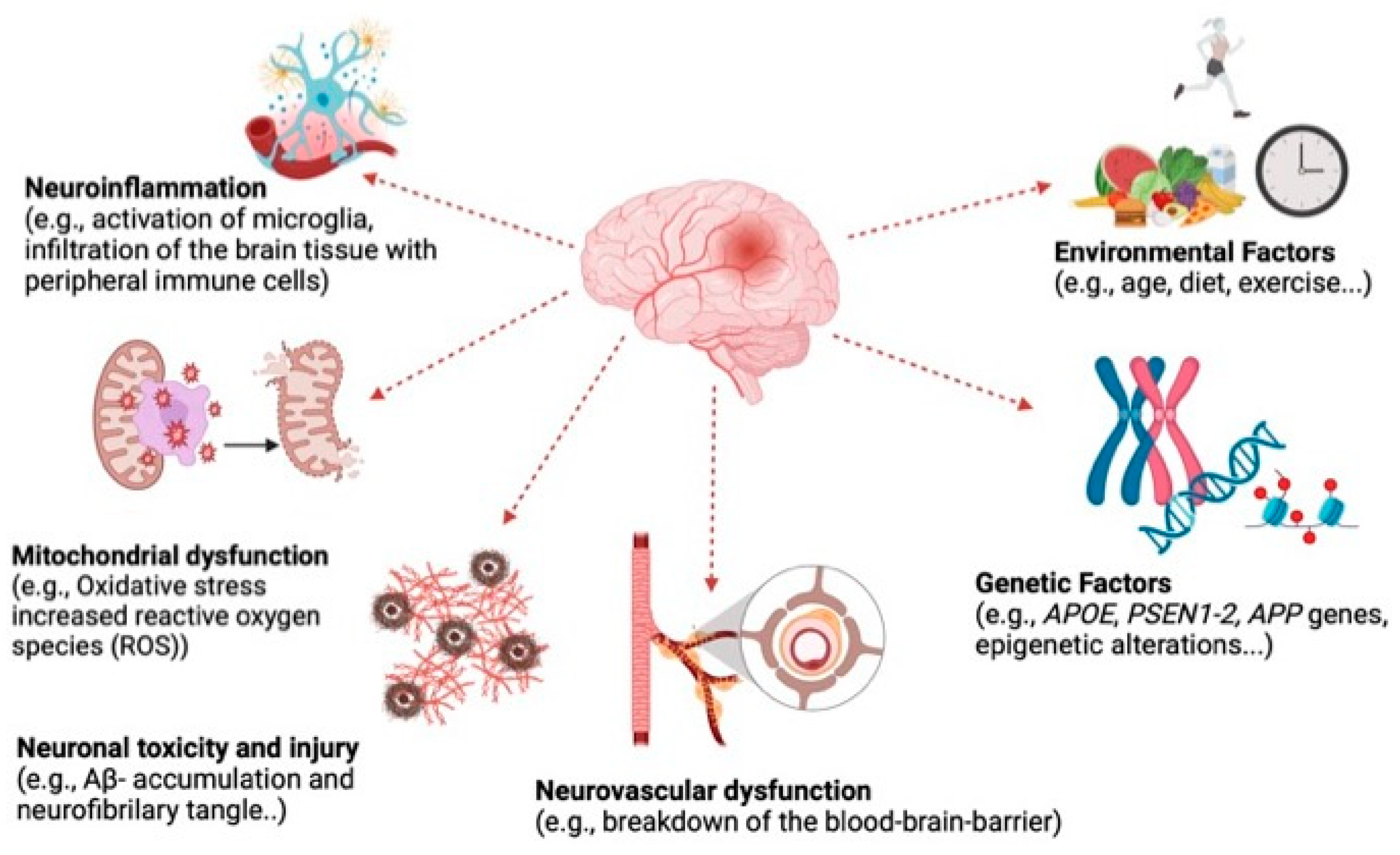
1. Introduction
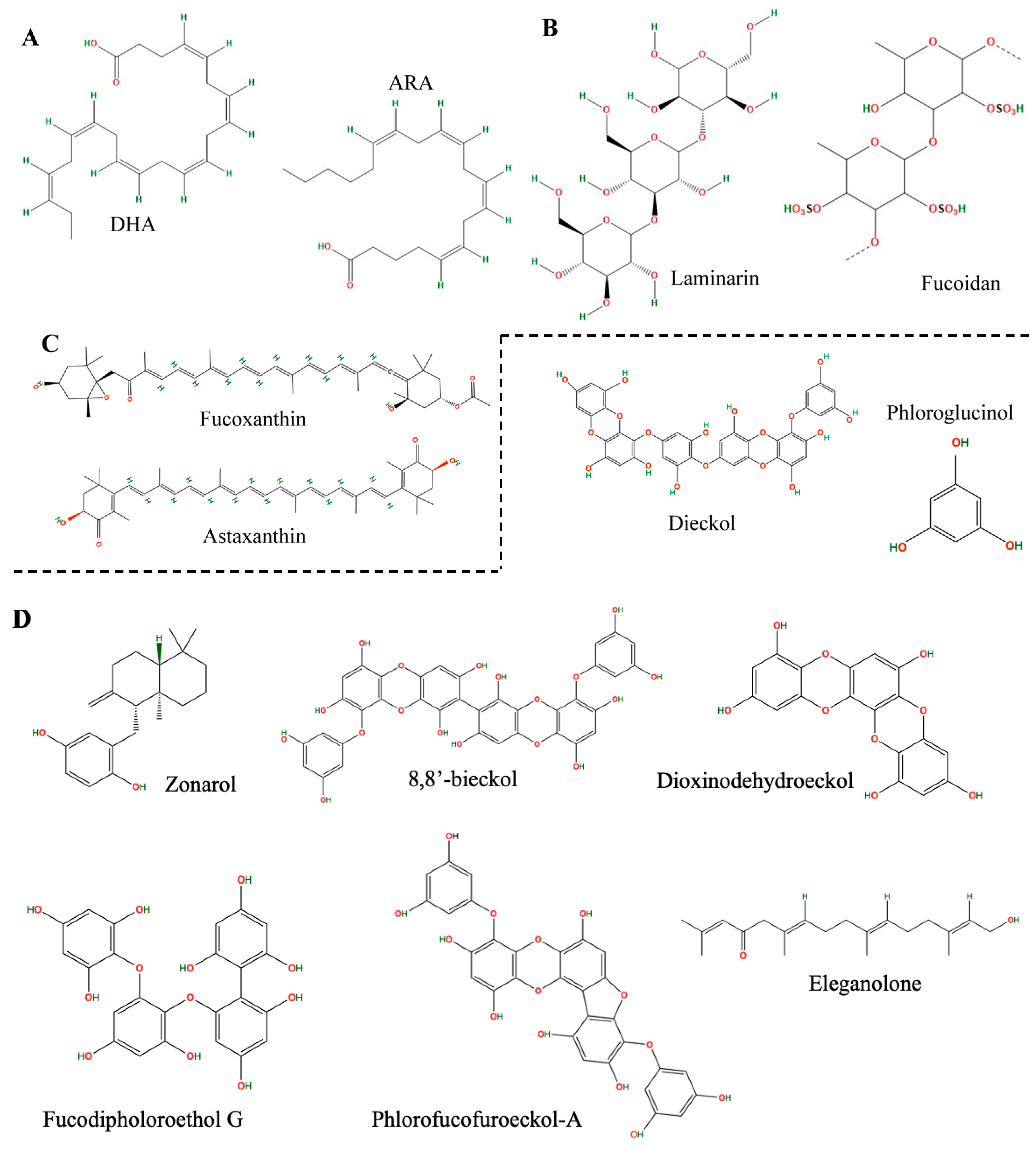
2. Neuroprotective Compound Extraction from Brown Seaweed
2.1. Conventional Solid–Liquid Extraction
2.2. Advanced Extraction Technologies for Brown Macroalgae Neuroprotection: A Greener Perspective
3. Chemical Characterization of Extracts from Brown Macroalgae
3.1. Liquid Chromatography
3.2. Other Techniques

{kind=link}
{kind=link}
| Family | Compounds | Brown Algae Species | Separation Technique—Detection Instrument (GC/LC Column) | Stationary Phase and Mobile Phases | Ionization Source | References |
|---|---|---|---|---|---|---|
| Lipids | Fatty acids, phospholipid, glyceroglycolipid, diglyceride and triglyceride. | Alaria esculenta, Ascophyllum nodosum, Fucus vesiculosus, Himanthalia elongata, Saccharina latissima, Sargassum muticum, and Sargassum fusiforme. | HPLC–Orbitrap–MS/MS (Poreshell 120 EC-C18) | RP. A: ACN/water 60:40 (0.1% FA and 10 mM NH4HCO2); B: IPA/ACN 90:10 (0.1% FA and 10 mM NH4HCO2). | ESI+; ESI- | [28] |
| Lipids | Sterols (saringosterol and fucosterol). | GC–MS (DB-XLB 122-1232) | - | EI | ||
| Polyphenol | Dieckol. | Ecklonia cava. | HPLC–DAD (C18 Kromasil 100-5) | RP. A: water; B: MeOH. | - | [41] |
| Amino acids, fatty acids, terpenoids, phenolics, aromatics, and cyclic ketones. | 6-aminocaproic acid, carnitine, 2-decenoic acid, docosahexaenoic acid, 5,8,11,14-eicosatetraynoic acid, 2,7,12,17-octadecanetetrol, 10,12-hexadecadienal, erucamide, 15-oxo-11,13-eicosadienoic acid, 16-hydroxypalmitic acid, 2-hydroxymyristic acid, 9,12,13-trihydroxy-15-octadecenoic acid, oleic acid, palmitic acid, 3,5-dibromo-4-hydroxybenzoic acid, 11-(2-hexyl-5-hydroxyphenoxy)-N- (2-hydroxyethyl) undecanamide, 6-gingerol, paradol, decanophenone, heptanophenone, haplamine, N-benzylformamide, 15-hydroxy-1-[2-(hydroxymethyl)-1-piperidinyl] prost-13-ene-1,9-dione, 8-[3-oxo-2-(2-penten-1-yl)-1-cyclopenten- 1-yl]octanoic acid, jasmone, laurendecumenyne A, coronaridine, ar-turmerone, retinaldehyde. | Laurencia snackeyi. | HPLC–DAD–Orbitrap MS/MS (Phenomenex C18) | RP A: water (0.1% FA); B: ACN (0.1% FA). | ESI+; ESI- | [42] |
| Palisadin A, aplysistatin and 5-acetoxypalisadin B. | NMR. | |||||
| Fatty acids, terpenoids | Vachanic acid methyl ester, (R)-ricinoleic acid, saikosaponin E, acanthoside D, and kansuinin D. | Petalonia binghamiae. | HPLC–DAD–QToF–MS (ZORBAX Eclipse Plus C18). | RP A: water (0.1% FA); B: CAN (0.1% FA). | ESI- | [43] |
| Phenolics | Total of 49 compounds: 18 phenolic acids, 22 flavonoids, 6 other polyphenols, 1 lignan and 1 stillbene. | Phyllospora comosa, Ecklonia radiata, Durvillaea sp., Sargassum sp., Cystophora sp. | LC–ESI–QTOF–MS/MS (Synergi Hydro-Reverse Phase 80°A). | RP A: water (0.1% FA); B: ACN/water (0.1% FA) (95:5). | ESI+; ESI- | [49] |
| Pigment | Fucoxanthin. | Sargassum mucclurei (1650.03 ± 7.10), S. binderi (296.07 ± 5.50), S. polycystum (3.82 ± 0.89), S. duplicatum (255.03 ± 5.71), S. denticarpum (24.25 ± 2.72), S. swartzii (217.60 ± 5.04), S. microcystum (161.52 ± 2.90), S. crassifolium (26.65 ± 2.86), S. oligocystum (2927.98 ± 8.01) *. | HPLC–DAD (Symmetry® C18). | RP ACN/MeOH (1:9, v/v). | - | [20] |
| Phenolics | Phlorotannins. | Cystoseira abies-marina. | HILIC × RP–DAD–MS/MS (lichrospher diol-5; Ascentis Express C18). | HILIC A: ACN/AcOH (98:2, v/v) B: MeOH/Water/AcOH (95:3:2, v/v/v). RP A: water (0.1% FA); B: can. | Ion trap with ESI- | [45] |
| Sulfate polysaccharide | Fucoidan. | Sargassum Horneri. | Fourier-transform infrared spectroscopy (FT-IR). | [47] | ||
| Polysaccharide | Alginate (through the measure of monosaccharides xylose, galactose, glucose, mannose, fructose, and rhamnose). | Padina pavonica, Sargassum cinereum, Turbinaria turbinata, and Dictyota dichotoma. | HPLC–UV–vis (C18). | RP A: water/ACN (90:10, v/v); B: ACN (0.045% KH2PO4–0.05% trimethylamine). | [50] | |
| Meroterpenoids | New sargasilols (9): (10′E)-10′-dehydroxy-11′,12′-dihydro-10′,11′-didehydro-12′-hydroperoxysargachromanol G; (10′E)-10′-dehydroxy-11′,12′-dihydro-10′,11′-dide-hydro-12′-hydroxysargachromanol G; 9′-deoxy-9′-oxo-11′,12′-dihydrosargachro-manol K; 9′-deoxysargachromanol K; 3′,4′-dihydro-4′,16′-didehydro-3′-oxosargachromanol E; 6′-hydroxy derivative of sargachromanol G; 15′-hydroxysargachromanol L; methylsargachro-manol E. | Sargassum siliquastrum. | NMR. | [51] |
4. Neuroprotection Assays of Brown Macroalgae Extracts
4.1. Cholinesterase Inhibitors
4.2. Amyloid-Beta (Aβ) Inhibition
4.3. Inhibition of Neurotoxic Effects in Cells
4.4. In Vivo Evaluation of Neuroprotective Effects
| In vitro | |||||
|---|---|---|---|---|---|
| Species | Extraction Approach | Possible Bioactive Compound | Pharmacological Markers/Biological Tests | Neuroprotective Effect | References |
| Sargassum Horneri | Enzyme-assisted extraction | Sulphated fucooligosaccharide | AChE and BChE | AChE (IC50: 4.0–14.4 µM) and BChE (IC50: 18.5–25.3 µM) inhibition | [47] |
| Ecklonia cava | Ethanol reflux extraction | Polyphenol and fucoidan | Mitochondrial reactive oxygen species (ROS) content amyloid-β production; tau hyperphosphorylation-mediated proteins; mitochondrial membrane potential (MMP, ΔΨm); adenosine triphosphate (ATP) content; mitochondria-mediated protein analysis (BAX, cytochrome C) | Reduced AChE activity; reduced mitochondrial ROS; ATP production and MMP restored down-regulating amyloid-β production (by JNK/IRS-1/IDE pathway); reduced tau hyperphosphorylation (by PI3K/Akt/GSK-3 pathway) | [8] |
| Ecklonia cava | Ethanol extraction | Phlorotannins | AChE and BChE | AChE (IC50: 0.9 ± 0.8 to 66.5 ± 0.4 μM) and BChE (IC50: 1.4 ± 3.8 to 25.2 ± 0.1 μM) inhibition | [56] |
| Ecklonia cava | Ethanol extraction | Eckol, dieckol, phlorofucofuroeckol-A (PFFA) and 974-A | H2O2, t-BHP, Aβ1–42 | Effective in ROS scavenging but not in protecting against oxidative stress-evoked neurotoxicity; PFFA and 974-Al; provided broad neuroprotective activity, including protection against oxidative stress and Aβ1–42. | [70] |
| Ecklonia cava | Ethanol extraction | Phlorotanninn and dieckol | AChE and BChE, AAPH and H2O2-induced oxidative stress in PC-12 and SH-SY5Y cells | AChE Inhibition (95.4%), BChE inhibition (74.7%), reduction in oxidative stress (26.3 to 51.1%) | [41] |
| Durvillaea incurvata | Ultrasound-assisted and conventional extraction | Crude ethanol extract | AChE and BChE | AChE (IC50: 48.55 ± 0.021 µg mL−1, 51.5% inhibition), BChE (IC50: 87.58 ± 0.044 µg mL−1, 32.8% inhibition) | [55] |
| Petalonia binghamiae | Sequential maceration | Achanic acid methyl ester, (R)-ricinoleic acid, saikosaponin E, acanthoside D and cansunin D | Glutamate-induced excitotoxicity, ROS scavenging, cell viability | Protection HT-22 cells from glutamate-induced excitotoxicity, increased cell viability and preserved cell morphology, reduced intracellular ROS production and increased HO-1 expression via Nrf2 activation. | [43] |
| Sargassum oligocystum Montagne | Maceration | Fucoxanthin | AChE, cytotoxic effect on C6 cells, neuroprotective effects against H2O2-induced oxidative stress and Aβ25–35-induced neurotoxicity, gene expression related to antioxidant enzymes (SOD, CAT, GPx), gene expression related to PI3K/Akt signaling (GSK-3β), ER stress and apoptosis-related gene expression (CHOP, Bax, caspase-3), gene expression related to ACh biosynthesis (ChAT, VAChT), protein translation (S6K1), autophagy regulation (p62, ATG5) | AChE (IC50: 130.12 ± 6.65 μg mL−1), protected C6 cells, viability increased to 91.23% at 100 μg mL−1 after H2O2 exposure, Increased cell survival rate significantly from 59.01% to 80.98% at 100 μg mL−1 after Aβ25–35 exposure; increased GPx activity by 105.81% at 100 μg mL−1 and CAT activity by 31.98% at 50 μg mL−1 after H2O2 exposure; increased mRNA expression of CAT and GPx; reversed the decrease in GSK3β induced by Aβ25–35; increased mRNA levels of ChAT and VAChT after Aβ25–35 treatment; inverted effect on S6K1 compared with galantamine; increased ATG5 mRNA levels and reduced p62 mRNA levels | [20] |
| Himanthalia elongata (L.) | Subcritical water extraction | Crude extract | •NO and O2•− scavenging activity, AChE and BChE | 40% AChE and 40% BChE inhibition, protects against oxidative and nitrosative stresses | [54] |
| Eisenia bicyclis (Kjellman) | Subcritical water extraction | Crude extract | •NO and O2•− scavenging activity, AChE and BChE | 50% AChE and 40% BChE inhibition, protects against oxidative and nitrosative stresses | [54] |
| Sargassum angustifolium | Extraction with acidic solution | Fucoidan | AChE, cytotoxic effects on NB4 cell line, Alterations in cell proliferation and cell cycle-related gene expression, Bcl-2 gene | AChE (IC50:1.20 µg mL−1); induction of p53, p21, and pro-apoptotic genes; inhibition of anti-apoptotic Bcl-2 gene | [16] |
| Padina tetrastromatica | Subcritical water hydrolysis | Low molecular weight peptides, flavonoids and phenolic compounds | AChE and α-amylase | AChE inhibition (IC50: 17.9 ± 0.1 to 65.9 ± 0.1 mg mL−1); α-amylase inhibition (2.4 ± 0.1 to 16.0 ± 0.5%) | [32] |
| Ishige foliacea | Maceration | Crude ethanol extract | AChE, BACE1, ROS scavenging | AChE inhibition (IC50: 205.1 μg mL−1), BACE1 (IC50: 266.8 μg mL−1), reduces H2O2 and Aβ-induced cell death in SH-SY5Y cells | [59] |
| Sargassum macrocarpum | Maceration | Terpenoid lactones | Human monoamine oxidases A and B | hMAO-A inhibition (42.18 ± 2.68% at 200 μM), any activity against hMAO-B | [62] |
| Dictyota coriacea | Maceration | Acetoxypachydiol | Keap1-Nrf2/HO-1, ROS scavenging | Reduces OGD/R and H2O2-induced cell death in SH-SY5Y cells, increased the mRNA and protein levels of Nrf2 and HO-1, decreased the protein level of Keap1, promoted the transport of Nrf2 to the cell nucleus | [65] |
| Padina gymnospora | Maceration | Crude extract and α-bisabolol | AChE, BACE1, ROS and RNS, apoptotic gene expression | Inhibition of cholinesterase (ChE) and β-secretase (BACE1) activity (specific inhibition values not stated); Reduction of ROS and RNS production; Attenuation of lipid and protein oxidation; Restoration of mitochondrial; Reduced Caspase-3 activation, increased Bcl-2 expression membrane potential | [66] |
| Fucus vesiculosus | Maceration | Intracellular and cell wall-bound phlorotannins | Aβ25–35-induced AD model (SH-SY5Y cells) | Protection at 5 μg mL−1 and 10 μg mL−1 | [67] |
| Pelvetia canaliculata | Maceration | Intracellular and cell wall-bound phlorotannins | Aβ25–35-induced AD model (SH-SY5Y cells) | Protection at all concentrations tested (1–10 µg mL−1) | [67] |
| Ecklonia maxima | Maceration | Phloroglucinol, catechin, epicatechin, biochanin A, vulgaxanthin and 7,2,4—trihydoxyisoflavanol | Cell viability; apoptosis (AO/EB staining); SOD, CAT, GSH, MDA, NO, AChE | Increased cell viability in HT-22 cells treated with ZnSO4; reduced apoptosis in Zn-treated cells; increased SOD and CAT activities; increased GSH levels; reduced MDA levels; decreased NO levels; reduced AChE activity | [68] |
| Gelidium pristoides | Maceration | Phloroglucinol, catechin, epicatechin, biochanin A, vulgaxanthin and 7,2,4—trihydoxyisoflavanol | Cell viability; apoptosis (AO/EB staining); SOD, CAT, GSH, MDA, NO, AChE | Increased cell viability in HT-22 cells treated with ZnSO4; reduced apoptosis in Zn-treated cells; increased SOD and CAT activities; increased GSH levels; reduced MDA levels; decreased NO levels; reduced AChE activity | [68] |
| Ecklonia radiata | Enzyme-assisted extraction | Crude extract (CE), phlorotannin (PT), poly-saccharide (PS), free sugar (FS) | Aβ1–42 aggregation, H2O2-induced cytotoxicity, cell viability | High neuroprotective activity (viability >92% at 3.125–100 μg mL−1); inhibition of Aβ1–42 aggregation; antioxidant activity at 12.5–50 μg mL−1; enhanced neurite outgrowth (More than 19%) | [69] |
| Ecklonia cava | Maceration | Dieckol | Cell viability, LDH, morphological assessment, ROS scavenging, mitochondrial function, ATP, mitochondrial membrane potential (ΔΨm), mitochondrial Ca2+ and ROS, Nrf2/HO-1 | Increased cell viability in primary cortical neurons and HT22 neurons; decreased LDH release indicating reduced cytotoxicity; improved neuronal morphology post-glutamate exposure; decreased intracellular ROS levels in both primary cortical neurons and HT22 cells; protected against glutamate-induced mitochondrial dysfunction; rescued ATP depletion in HT22 neurons; prevented ΔΨm disruption in HT22 neurons; attenuated mitochondrial Ca2+ overload in HT22 neurons; reduced mitochondrial ROS levels in HT22 neurons; increased HO-1 expression and Nrf2 nuclear translocation | [71] |
| Ecklonia radiata | Maceration | Dibenzodioxin-fucodiphloroethol | Cell viability, Aβ1–42 toxicity and aggregation, molecular docking, AChE, ROS scavenging | DFD (50 µM) does not induce toxicity in PC-12 cells; Rescued PC-12 cell viability at 1.0 and 1.5 µM Aβ1–42; Reduced Aβ1–42 aggregation; Attenuated ROS levels in PC-12 cells; AChE inhibition (IC50 = 41.09 µM); Binded to Aβ1–42 with a docking score of −43.28, forming hydrogen bonds with HIS14 and GLU11; Interacted with AChE with a docking score of −43.48, forming hydrogen bonds and π-π stacking; Binded to Aβ1–42 with a docking score of −43.28, forming hydrogen bonds with HIS14 and GLU11; Binded to Aβ1–42 pentamer with a docking score of −64.01, interacting with multiple residues | [28] |
| Sargassum horneri | Maceration | Fucosterol | Cell viability, ROS scavenging, Nrf2/HO-1, TNF-α/IFN-γ, NF-κB/MAPK | Biocompatible with HDF cells up to 120 μM; decreased in a dose-dependent manner the intracellular ROS production in HDFs; upregulated Nrf2 and HO-1 expression in HDF cells; down-regulated inflammatory mediators in TNF-α/IFN-γ-stimulated HDF cells; reduced phosphorylation of NF-κB and MAPK mediators in a dose-dependent manner; decreased molecules related to connective tissue degradation | [70] |
| Bifurcaria bifurcata | Maceration | Eleganolone | Cell viability, ROS scavenging, mitochondrial function, caspase-3, NF-κB, TNF-α, IL-6, IL-10 | Increased cell viability after 6-OHDA treatment; reduced ROS levels and H2O2 production in SH-SY5Y cells exposed to 6-OHDA; preserved mitochondrial membrane potential (MMP) and ATP levels in SH-SY5Y cells; reduced caspase-3 activity in SH-SY5Y cells exposed to 6-OHDA; inhibited NF-κB p65 translocation in SH-SY5Y cells after 6-OHDA exposure; reduced LPS-induced NO production; decreased TNF-α and IL-6 production in LPS-stimulated RAW 264.7 cells | [76] |
| Fucus guiryi | Maceration | Phlorotannins | AChE, BChE, MAO-A, MAO-B | AChE inhibition (IC50 μg DE mL−1): 969.51 ± 76.99; BChE inhibition (IC50 μg DE mL−1): 1065.29 ± 35.35; MAO-A inhibition (IC50 μg DE mL−1): 168.24 ± 5.40; MAO-B inhibition (IC50 μg DE mL−1): >500; tyrosinaz inhibition (IC50 μg DE mL−1): 47.99 ± 0.59 | [72] |
| Fucus serratus | Maceration | Phlorotannins | AChE, BChE, MAO-A, MAO-B | AChE inhibition (IC50 μg DE mL−1): 2709.58 ± 55.25; BChE inhibition (IC50 μg DE mL−1): 3539.79 ± 109.43; MAO-A inhibition (IC50 μg DE mL−1): 173.80 ± 25.52; MAO-B inhibition (IC50 μg DE mL−1): >500; tyrosinaz Inhibition (IC50 μg DE mL−1): 47.66 ± 2.84 | [72] |
| Fucus spiralis L. | Maceration | Phlorotannins | AChE, BChE, MAO-A, MAO-B | AChE inhibition (IC50 μg DE mL−1): >5000; BChE inhibition (IC50 μg DE mL−1): >5000; MAO-A inhibition (IC50 μg DE mL−1): 1929.65 ± 100.44; MAO-B inhibition (IC50 μg DE mL−1): >500; tyrosinaz inhibition (IC50 μg DE mL−1): 861.73 ± 7.37 | [72] |
| Fucus vesiculosus L. | Maceration | Phlorotannins | AChE, BChE, MAO-A, MAO-B | AChE inhibition (IC50 μg DE mL−1): >5000; BChE inhibition (IC50 μg DE mL−1): >5000; MAO-A inhibition (IC50 μg DE mL−1): >3000; MAO-B inhibition (IC50 μg DE mL−1): >500; tyrosinaz inhibition (IC50 μg DE mL−1): 2546.82 ± 98.00 | [72] |
| Padina australis | Maceration | Fucosterol | Cell viability, apoptosis, APP mRNA, intracellular Aβ, BACE1, ROS scavenging | Increased cell viability in SH-SY5Y cells treated with 2 μM Aβ; reduced apoptosis (30.93%); reduced APP mRNA levels; reduced intracellular Aβ levels; noncompetitive inhibitor on β-secretase (BACE1); reduced oxidative stress by increasing anti-oxidant enzyme activities | [75] |
| Dictyota coriacea | Maceration | Xenicane diterpenes | Cell viability, LDH, Nrf2/ARE | Increased cell viability in PC12 cells damaged by H2O2; reduced LDH levels released from PC12 cells; increased Nrf2 and HO-1 expression in PC12 cells; promoted nuclear translocation of Nrf2 | [77] |
| Dictyota spp. | Maceration | Hydroazulene diterpenes | Cell viability, Nrf2/ARE | Increased cell viability in H2O2-damaged PC12 cells at a concentration of 2 μM; antioxidant effect by activating the Nrf2/ARE signaling pathway | [78] |
| Dictyopteris polypodioides | Maceration | Meroterpenoids: yahazunol (1), zonarol (2), isozonarol (3), and four other meroterpenoids | NO production, iNOS, IL-6, and CCL2 mRNA expression, structure activity of compounds | Inhibited NO production in lipopolysaccharide (LPS)-stimulated RAW264 cells; inhibited iNOS, IL-6, and CCL2 mRNA expression. The hydroquinone unit is important in the anti-inflammatory activity of these sesquiterpenoids | [79] |
| Sargassum siliquastrum | Maceration | Meroterpenoids | BV-2 microglial cells, anti-inflammatory cytokines | Reduced the expression of anti-inflammatory cytokines (IL-1β, IL-6, TNF-α); inhibited LPS-induced NO production in BV-2 microglial cells by targeting IKK/IκB/NF-κB pathways | [51] |
| Fucus vesiculosus | Maceration | Fucoidan | CCK-8 (cell viability), LDH, Hoechst staining, MAP2 immunostaining, MitoSOX staining (for ROS), protein target identification (ATP5F1a) | Enhanced cell viability at concentrations of 5, 10, and 25 μM in MPP+-induced SH-SY5Y cells; reduced LDH release in MPP+-induced SH-SY5Y cells and primary neurons, indicating reduced cell damage. Decreased the proportion of apoptotic cells in MPP+-treated primary neurons; protected neurons from MPP+-induced axon loss and damage. Reduced mitochondrial ROS production in MPP+-treated SH-SY5Y cells. ATP5F1a knockdown reversed the neuroprotective effects of FvF, confirming its role in mitigating mitochondrial dysfunction and apoptosis | [80] |
| Fucus vesiculosus | Maceration | Fucoidan | Aβ1–42 aggregation and cytotoxicity, ROS Scavenging, neurite outgrowth | Protected against Aβ1–42-induced cytotoxicity; inhibited Aβ1–42 aggregation; slight protection against hydrogen peroxide-induced cytotoxicity; inhibited apoptosis induced by Aβ1–42 | [22] |
| Padina australis | Maceration | Crude ethanol extract | Cell viability, NO, prostaglandin E2, ROS scavenging, iNOS and COX-2 expression, TNF-α and IL-6 secretion | No cytotoxicity at concentrations of 0.25–2.0 mg mL−1, reduced at higher concentrations (4.0–8.0 mg mL−1); reduced NO production compared with LPS-stimulated levels dose-dependently (0.5–2.0 mg mL−1); suppressed PGE2 production at concentrations of 0.5–1.0 mg mL−1, increased PGE2 levels at higher concentration (2.0 mg mL−1); reduced intracellular ROS generation compared with LPS-stimulated levels dose-dependently; down-regulated iNOS and COX-2 expression induced by LPS; inhibited TNF-α and IL-6 secretion compared with LPS-stimulated levels | [81] |
| In vivo | |||||
| Species | Possible Bioactive Compound | Pharmacological Markers/Biological Tests | Neuroprotective Effect | References | |
| Dictyopteris undulata | Zonarol | Brain tissue distribution after administration | Higher level in brain tissue than in other tissues | [82] | |
| Ecklonia cava | Crude water extracts | Reduction of pro-inflammatory cytokines, NF-κB, STAT3 phosphorylation | Reduced neuroinflammation in LPS-induced model | [83] | |
| Ishige foliacea | Crude ethanol extract | BDNF and TrkB expression, synaptic plasticity | Increased BDNF, TrkB-phosphorylated ERK expression | [59] | |
| Sargassum fusiformis | Fucoxanthin (Fx) | Reduction of NO, ROS, cell death | Reduced inflammatory responses | [9] | |
| Sargassum fusiformis | Phlorotannin and fucoidan mixture | Protection against Aβ-induced learning and memory impairments | Anti-amnesiac effect | [9] | |
| Sargassum ilicifolium and Padina tetrastromatica | Crude chloroform and ethanol extracts | Improvement of memory impairments, oxidative enzyme protection, AChE inhibition | Cognitive improvement | [84] | |
| Fucus vesiculosus | Fucoidan | Improvement of neuroinflammation, promotion of neurogenesis, reduced blood–brain and intestinal barrier permeability | Neuroprotective effects | [85] | |
| Sargassum wightii Greville | Fucoidan | Behavioral assays, oxidative stress markers, hyperphosphorylated tau protein, amyloidosis symptoms | Improvement of cognitive impairments and oxidative stress | [86] | |
| Padina gymnospora | Crude extract and α-bisabolol | Lifespan in CL2006 and CL4176 mutants | Lowering the expression of AD-related genes (ace-1, hsp-4) in C. Elegans | [66] | |
| Dictyota coriacea | Xenicane diterpenes | CIRI Model | Reduced brain infarct size and neurological deficit score in the transient middle cerebral artery occlusion (MCAO) rat model | [83] | |
| Dictyota spp. | Hydroazulene diterpenes | CIRI Model | Reduction in both brain infarct size and neurological deficit score in transient middle cerebral artery occlusion (MCAO) rat model | [78] | |
| Fucus vesiculosus | Fucoidan (FvF) | MPTP mouse model | Improved motor coordination and balance; slowed loss of TH markers; mitigated dopamine neuron loss in the substantia nigra pars compacta (SNpc). | [80] | |
| Laminaria japonica | Fucoidan | Adenine-induced CKD mice | Reduced signs associated with cognitive behavior; improvement in genes related to Alzheimer’s disease and memory; regulated of inflammatory response via microglia/macrophage polarization; ameliorated oxidative stress via Nrf2-HO-1 signaling pathway; improved cognitive impairments (new object recognition, object location, passive avoidance tests) | [87] | |
| Fucus vesiculosus | Fucoidan | Mongolian gerbils (Meriones unguiculatus) | Reduced tGCI-induced hyperactivity; reduced the loss of NeuN+ pyramidal cells in the CA1 region after tGCI; reduced the number of F-J B+ cells in the CA1 region; reduced the activation of GFAP+ astroglia and the production of reactive oxygen species; reduced the activation of Iba-1+ microglia and the production of reactive oxygen species; reduced HNE immunoreactivity and lipid peroxidation in CA1 pyramidal cells; reduced superoxide levels in CA1 pyramidal cells; increased SOD1 and SOD2 expression in CA1 pyramidal cells | [88] | |
| Laminaria digitata | Laminarin | Mongolian gerbils | Decreased superoxide anions; decreased IL-1β and TNF-α; increased SOD expression; increased IL-4 and IL-13 expression | [89] | |
| Petalonia binghamiae | Water extract | MCAO/R mice model | Decreased neuronal death in ischemic lesion; attenuated sensorimotor deficits; reduced ROS levels; decreased apoptosis in ischemic area; enhanced Nrf2 nuclear translocation; increased HO-1 expression | [43] | |
| Sargassum boveanum | Crude ethanolic extract | Sprague–Dawley rats | Increased antioxidant enzyme activities (SOD, GPx) and gene expressions (SOD, GPx, Nrf2, HO-1); decreased expression of antioxidant genes (SOD, GPx, Nrf2, HO-1); increased expression of inflammatory markers (TNF-α, NF-κB) | [90] | |
| Sargassum angustifolium | Hydroalcoholic, methanolic, and hexane extract | Reversal of memory impairment, passive avoidance test, Morris water maze test | Improvement of cognitive functions | [91] | |
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lamisa, A.B.; Ahammad, I.; Bhattacharjee, A.; Hossain, M.U.; Ishtiaque, A.; Chowdhury, Z.M.; Das, K.C.; Salimullah, M.; Keya, C.A. A Meta-Analysis of Bulk RNA-Seq Datasets Identifies Potential Biomarkers and Repurposable Therapeutics against Alzheimer’s Disease. Sci. Rep. 2024, 14, 24717. [Google Scholar] [CrossRef] [PubMed]
- Ashrafian, H.; Zadeh, E.H.; Khan, R.H. Review on Alzheimer’s Disease: Inhibition of Amyloid Beta and Tau Tangle Formation. Int. J. Biol. Macromol. 2021, 167, 382–394. [Google Scholar] [CrossRef] [PubMed]
- Alhazmi, H.A.; Albratty, M. An Update on the Novel and Approved Drugs for Alzheimer Disease. Saudi Pharm. J. 2022, 30, 1755–1764. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D. How Many Species of Algae Are There? A Reprise. Four Kingdoms, 14 Phyla, 63 Classes and Still Growing. J. Phycol. 2024, 60, 214–228. [Google Scholar] [CrossRef]
- El-Manaway, I.M.; Rashedy, S.H. The Ecology and Physiology of Seaweeds: An Overview. In Sustainable Global Resources of Seaweeds Volume 1: Bioresources, Cultivation, Trade and Multifarious Applications; Ranga Rao, A., Ravishankar, G.A., Eds.; Springer: Cham, Switzerland, 2022; Volume 1, pp. 3–16. [Google Scholar]
- Wan, A.H.L.; Davies, S.J.; Soler-Vila, A.; Fitzgerald, R.; Johnson, M.P. Macroalgae as a Sustainable Aquafeed Ingredient. Rev. Aquac. 2019, 11, 458–492. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, H.; Wang, Y.; Peng, Z.; Guo, Z.; Ma, Y.; Zhang, R.; Zhang, M.; Wu, Q.; Xiao, J.; et al. Effects of Different Extraction Methods on Contents, Profiles, and Antioxidant Abilities of Free and Bound Phenolics of Sargassum polycystum from the South China Sea. J. Food Sci. 2022, 87, 968–981. [Google Scholar] [CrossRef]
- Park, S.K.; Kang, J.Y.; Kim, J.M.; Yoo, S.K.; Han, H.J.; Chung, D.H.; Kim, D.O.; Kim, G.H.; Heo, H.J. Fucoidan-Rich Substances from Ecklonia cava Improve Trimethyltin-Induced Cognitive Dysfunction via Down-Regulation of Amyloid β Production/Tau Hyperphosphorylation. Mar. Drugs 2019, 17, 591. [Google Scholar] [CrossRef]
- Dai, Y.L.; Jiang, Y.F.; Lu, Y.A.; Yu, J.B.; Kang, M.C.; Jeon, Y.J. Fucoxanthin-Rich Fraction from Sargassum fusiformis Alleviates Particulate Matter-Induced Inflammation in Vitro and in Vivo. Toxicol. Rep. 2021, 8, 349–358. [Google Scholar] [CrossRef]
- Martić, A.; Čižmek, L.; Ul’yanovskii, N.V.; Paradžik, T.; Perković, L.; Matijević, G.; Vujović, T.; Baković, M.; Babić, S.; Kosyakov, D.S.; et al. Intra-Species Variations of Bioactive Compounds of Two Dictyota Species from the Adriatic Sea: Antioxidant, Antimicrobial, Dermatological, Dietary, and Neuroprotective Potential. Antioxidants 2023, 12, 857. [Google Scholar] [CrossRef]
- Ciko, A.M.; Jokić, S.; Šubarić, D.; Jerković, I. Overview on the Application of Modern Methods for the Extraction of Bioactive Compounds from Marine Macroalgae. Mar. Drugs 2018, 16, 348. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of Novel Extraction Technologies for Bioactives from Marine Algae. J. Agric. Food Chem. 2013, 61, 4667–4675. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.; Fernandes, M.; Lima, M.; Gomes, J.P.; Silva, F.; Castro, S.; Sampaio, F.; Gomes, A.C. Nanotechnology to the Rescue: Therapeutic Strategies Based on Brown Algae for Neurodegenerative Diseases. Appl. Sci. 2023, 13, 1883. [Google Scholar] [CrossRef]
- Aakre, I.; Solli, D.D.; Markhus, M.W.; Mæhre, H.K.; Dahl, L.; Henjum, S.; Alexander, J.; Korneliussen, P.A.; Madsen, L.; Kjellevold, M. Commercially Available Kelp and Seaweed Products—Valuable Iodine Source or Risk of Excess Intake? Food Nutr. Res. 2021, 65, 7584. [Google Scholar] [CrossRef] [PubMed]
- Byoung, W.C.; Ryu, G.; Soo, H.P.; Eun, S.K.; Shin, J.; Seok, S.R.; Hyeon, C.S.; Bong, H.L. Anticholinesterase Activity of Plastoquinones from Sargassum sagamianum: Lead Compounds for Alzheimer’s Disease Therapy. Phytother. Res. 2007, 21, 423–426. [Google Scholar] [CrossRef]
- Hosseinpouri, A.; Mohammadi, M.; Ehsandoost, E.; Sharafi-Badr, P.; Obeidi, N. Chemical Identification, Antioxidant, Cholinesterase Inhibitory, and Cytotoxic Properties of Fucoidan Extracted from Persian Gulf Sargassum angustifolium. Acta Oceanol. Sin. 2022, 41, 133–141. [Google Scholar] [CrossRef]
- Ktari, L.; Mdallel, C.; Aoun, B.; Chebil Ajjabi, L.; Sadok, S. Fucoxanthin and Phenolic Contents of Six Dictyotales from the Tunisian Coasts with an Emphasis for a Green Extraction Using a Supercritical CO2 Method. Front. Mar. Sci. 2021, 8, 647159. [Google Scholar] [CrossRef]
- Jiang, H.; Kong, L.; Tang, H.; Wang, Z.; Liu, C.; Zhang, J.; Chen, Y.; Shen, J.; Zhou, Y. Study on the Preparation and Enzyme Inhibitory Activity of Polyphenols from Sargassum pallidum. PLoS ONE 2024, 19, e0297434. [Google Scholar] [CrossRef]
- Ruiz-Domínguez, M.C.; Mendiola, J.A.; Sánchez-Martínez, J.D.; Bueno, M.; Cerezal-Mezquita, P.; Ibáñez, E. Evaluation of the Antioxidant and Neuroprotective Activity of the Seaweed Durvillaea antarctica (Cochayuyo) Extracts Using Pressurized Liquids. J. Appl. Phycol. 2023, 35, 835–847. [Google Scholar] [CrossRef]
- Hong, D.D.; Thom, L.T.; Ha, N.C.; Thu, N.T.H.; Hien, H.T.M.; Tam, L.T.; Dat, N.M.; Duc, T.M.; Van Tru, N.; Hang, N.T.M.; et al. Isolation of Fucoxanthin from Sargassum oligocystum Montagne, 1845 Seaweed in Vietnam and Its Neuroprotective Activity. Biomedicines 2023, 11, 2310. [Google Scholar] [CrossRef]
- Sánchez-Camargo, A.P.; Montero, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Application of Hansen Solubility Approach for the Subcritical and Supercritical Selective Extraction of Phlorotannins from Cystoseira abies-marina. RSC Adv. 2016, 6, 94884–94895. [Google Scholar] [CrossRef]
- Alghazwi, M.; Smid, S.; Karpiniec, S.; Zhang, W. Comparative Study on Neuroprotective Activities of Fucoidans from Fucus vesiculosus and Undaria pinnatifida. Int. J. Biol. Macromol. 2019, 122, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Ehrig, K.; Alban, S. Sulfated Galactofucan from the Brown Alga Saccharina latissima-Variability of Yield, Structural Composition and Bioactivity. Mar. Drugs 2015, 13, 76–101. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.; Barbero, G.F.; PiñEiro, Z.; Liazid, A.; Barroso, C.G.; Rostagno, M.A.; Prado, J.M.; Meireles, M.A.A. Chapter 2: Extraction of Natural Products: Principles and Fundamental Aspects. In Natural Product Extraction: Principles and Applications; Green Chemistry Series; Royal Society of Chemistry: London, UK, 2013; pp. 58–88. [Google Scholar] [CrossRef]
- Dertli, H.; Saloglu, D. A Valorization Approach of Food Industry Wastewater Using Microwave-Assisted Extraction. In Advanced Technologies in Wastewater Treatment: Food Processing Industry; Elsevier: Amsterdam, The Netherlands, 2023; Chapter 6; pp. 155–178. [Google Scholar] [CrossRef]
- Shinoda, S.; Tozawa, Y.; Kurimoto, S.; Shigemori, H.; Sekiguchi, M. Three New Meroterpenoids from Sargassum macrocarpum and Their Inhibitory Activity against Amyloid β Aggregation. J. Nat. Med. 2023, 77, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Johnston, M.R.; Zhang, W.; Smid, S.D. A Phlorotannin Isolated from Ecklonia radiata, Dibenzodioxin-Fucodiphloroethol, Inhibits Neurotoxicity and Aggregation of β-Amyloid. Phytomedicine Plus 2021, 1, 100125. [Google Scholar] [CrossRef]
- Martens, N.; Zhan, N.; Voortman, G.; Leijten, F.P.J.; van Rheenen, C.; van Leerdam, S.; Geng, X.; Huybrechts, M.; Liu, H.; Jonker, J.W.; et al. Activation of Liver X Receptors and Peroxisome Proliferator-Activated Receptors by Lipid Extracts of Brown Seaweeds: A Potential Application in Alzheimer’s Disease? Nutrients 2023, 15, 3004. [Google Scholar] [CrossRef]
- Priego-Capote, F.; De La Torre, M.D.P.D. Chapter 5: Accelerated Liquid Extraction. In Natural Product Extraction: Principles and Applications; Green Chemistry Series; Royal Society of Chemistry: London, UK, 2013; pp. 157–195. [Google Scholar] [CrossRef]
- Zohar, M.; Matzrafi, M.; Abu-Nassar, J.; Khoury, O.; Gaur, R.Z.; Posmanik, R. Subcritical Water Extraction as a Circular Economy Approach to Recover Energy and Agrochemicals from Sewage Sludge. J. Environ. Manag. 2021, 285, 112111. [Google Scholar] [CrossRef]
- Boucelkha, A.; Petit, E.; Elboutachfaiti, R.; Molinié, R.; Amari, S.; Yahaoui, R.Z. Production of Guluronate Oligosaccharide of Alginate from Brown Algae Stypocaulon scoparium Using an Alginate Lyase. J. Appl. Phycol. 2017, 29, 509–519. [Google Scholar] [CrossRef]
- Hans, N.; Solanki, D.; Nagpal, T.; Amir, H.; Naik, S.; Malik, A. Process Optimization and Characterization of Hydrolysate from Underutilized Brown Macroalgae (Padina tetrastromatica) after Fucoidan Extraction through Subcritical Water Hydrolysis. J. Environ. Manag. 2024, 349, 119497. [Google Scholar] [CrossRef]
- Pingret, D.; Fabiano-Tixier, A.S.; Chemat, F. Chapter 3: Ultrasound-Assisted Extraction. In Natural Product Extraction: Principles and Applications; Green Chemistry Series; Royal Society of Chemistry: London, UK, 2013; pp. 89–112. [Google Scholar] [CrossRef]
- Soares, C.; Paíga, P.; Marques, M.; Neto, T.; Carvalho, A.P.; Paiva, A.; Simões, P.; Costa, L.; Bernardo, A.; Fernández, N.; et al. Multi-Step Subcritical Water Extracts of Fucus vesiculosus L. and Codium tomentosum Stackhouse: Composition, Health-Benefits and Safety. Processes 2021, 9, 893. [Google Scholar] [CrossRef]
- Wee, A.S.; Nhu, T.D.; Khaw, K.Y.; Tang, K.S.; Yeong, K.Y. Linking Diabetes to Alzheimer’s Disease: Potential Roles of Glucose Metabolism and Alpha-Glucosidase. Curr. Neuropharmacol. 2022, 21, 2036–2048. [Google Scholar] [CrossRef]
- Cebrián-Lloret, V.; Cartan-Moya, S.; Martínez-Sanz, M.; Gómez-Cortés, P.; Calvo, M.V.; López-Rubio, A.; Martínez-Abad, A. Characterization of the Invasive Macroalgae Rugulopteryx Okamurae for Potential Biomass Valorisation. Food Chem. 2024, 440, 138241. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease. Mar. Drugs 2019, 17, 609. [Google Scholar] [CrossRef] [PubMed]
- Quitain, A.T.; Kai, T.; Sasaki, M.; Goto, M. Microwave-Hydrothermal Extraction and Degradation of Fucoidan from Supercritical Carbon Dioxide Deoiled Undaria pinnatifida. Ind. Eng. Chem. Res. 2013, 52, 7940–7946. [Google Scholar] [CrossRef]
- Lama-Muñoz, A.; Contreras, M.M. Extraction Systems and Analytical Techniques for Food Phenolic Compounds: A Review. Foods 2022, 11, 3671. [Google Scholar] [CrossRef]
- Anjana, K.; Arunkumar, K. Brown Algae Biomass for Fucoxanthin, Fucoidan and Alginate; Update Review on Structure, Biosynthesis, Biological Activities and Extraction Valorisation. Int. J. Biol. Macromol. 2024, 280, 135632. [Google Scholar] [CrossRef]
- Nho, J.A.; Shin, Y.S.; Jeong, H.R.; Cho, S.; Heo, H.J.; Kim, G.H.; Kim, D.O. Neuroprotective Effects of Phlorotannin-Rich Extract from Brown Seaweed Ecklonia cava on Neuronal PC-12 and SH-SY5Y Cells with Oxidative Stress. J. Microbiol. Biotechnol. 2020, 30, 359–367. [Google Scholar] [CrossRef]
- Palaniveloo, K.; Ong, K.H.; Satriawan, H.; Abdul Razak, S.; Suciati, S.; Hung, H.Y.; Hirayama, S.; Rizman-Idid, M.; Tan, J.K.; Yong, Y.S.; et al. In Vitro and in Silico Cholinesterase Inhibitory Potential of Metabolites from Laurencia snackeyi (Weber-van Bosse) M. Masuda. 3 Biotech 2023, 13, 337. [Google Scholar] [CrossRef]
- Eom, S.H.; Hong, G.L.; Kang, H.B.; Lee, N.S.; Kim, D.K.; Jeong, Y.G.; Kim, C.S.; Yoo, Y.C.; Lee, B.H.; Jung, J.Y.; et al. Neuroprotective Effects of Water Extract from Brown Algae Petalonia Binghamiae in an Experimental Model of Focal Cerebral Ischemia In Vitro and In Vivo. Curr. Issues Mol. Biol. 2023, 45, 8427–8443. [Google Scholar] [CrossRef]
- van den Hurk, R.S.; Pursch, M.; Stoll, D.R.; Pirok, B.W.J. Recent Trends in Two-Dimensional Liquid Chromatography. TrAC Trends Anal. Chem. 2023, 166, 117166. [Google Scholar] [CrossRef]
- Montero, L.; Herrero, M.; Ibáñez, E.; Cifuentes, A. Separation and Characterization of Phlorotannins from Brown Algae Cystoseira Abies-Marina by Comprehensive Two-Dimensional Liquid Chromatography. Electrophoresis 2014, 35, 1644–1651. [Google Scholar] [CrossRef]
- Li, S.; Hu, M.; Tong, Y.; Xia, Z.; Tong, Y.; Sun, Y.; Cao, J.; Zhang, J.; Liu, J.; Zhao, S.; et al. A Review of Volatile Compounds in Edible Macroalgae. Food Res. Int. 2023, 165, 112559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Zhang, H.Z.; Liu, Y.W.; Tang, M.; Jiang, Y.J.; Li, F.N.; Guan, L.P.; Jin, Q.H. Sulphated Fucooligosaccharide from Sargassum horneri: Structural Analysis and Anti-Alzheimer Activity. Neurochem. Res. 2024, 49, 1592–1602. [Google Scholar] [CrossRef] [PubMed]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-Proliferative Activity and Chemical Characterization by Comprehensive Two-Dimensional Liquid Chromatography Coupled to Mass Spectrometry of Phlorotannins from the Brown Macroalga Sargassum muticum Collected on North-Atlantic Coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Subbiah, V.; Duan, X.; Agar, O.T.; Dunshea, F.R.; Barrow, C.J.; Suleria, H.A.R. Comparative Study on the Effect of Different Drying Techniques on Phenolic Compounds in Australian Beach-Cast Brown Seaweeds. Algal Res. 2023, 72, 103140. [Google Scholar] [CrossRef]
- El-Sheekh, M.; Kassem, W.M.A.; Alwaleed, E.A.; Saber, H. Optimization and Characterization of Brown Seaweed Alginate for Antioxidant, Anticancer, Antimicrobial, and Antiviral Properties. Int. J. Biol. Macromol. 2024, 278, 134715. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, Z.; Zhang, J.; Tang, S.; Zhu, H.; Jiang, B.; Li, X.; Wang, J.; Sun, Z.; Zhao, M.; et al. Anti-Neuroinflammatory Meroterpenoids from a Chinese Collection of the Brown Alga Sargassum siliquastrum. J. Nat. Prod. 2023, 86, 1284–1293. [Google Scholar] [CrossRef]
- Koul, B.; Farooq, U.; Yadav, D.; Song, M. Phytochemicals: A Promising Alternative for the Prevention of Alzheimer’s Disease. Life 2023, 13, 999. [Google Scholar] [CrossRef]
- Pedro, B.; Guedes, L.; André, R.; Gaspar, H.; Vaz, P.; Ascensão, L.; Melo, R.; Luísa Serralheiro, M. Undaria pinnatifida (U. pinnatifida) Bioactivity: Antioxidant, Gastro-Intestinal Motility, Cholesterol Biosynthesis and Liver Cell Lines Proteome. J. Funct. Foods 2021, 83, 104567. [Google Scholar] [CrossRef]
- Gomes, I.; Rodrigues, H.; Rodrigues, C.; Marques, M.; Paíga, P.; Paiva, A.; Simões, P.; Fernandes, V.C.; Vieira, M.; Delerue-Matos, C.; et al. Evaluation of the Biological Potential of Himanthalia Elongata (L.) S.F.Gray and Eisenia Bicyclis (Kjellman) Setchell Subcritical Water Extracts. Foods 2022, 11, 746. [Google Scholar] [CrossRef]
- Muñoz-Molina, N.; Parada, J.; Simirgiotis, M.; Montecinos-González, R. The Potential of Using Cochayuyo (Durvillaea incurvata) Extract Obtained by Ultrasound-Assisted Extraction to Fight against Aging-Related Diseases. Foods 2024, 13, 269. [Google Scholar] [CrossRef]
- Park, S.R.; Kim, Y.H.; Yang, S.Y. Enzyme Kinetics and Molecular Docking Investigation of Acetylcholinesterase and Butyrylcholinesterase Inhibitors from the Marine Alga Ecklonia cava. Nat. Product. Sci. 2023, 29, 182–192. [Google Scholar] [CrossRef]
- Dhanabalan, A.K.; Kumar, P.; Vasudevan, S.; Chworos, A.; Velmurugan, D. Identification of a Novel Drug Molecule for Neurodegenerative Disease from Marine Algae through in-silico Analysis. J. Biomol. Struct. Dyn. 2024, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chagas Monteiro, K.L.; dos Santos Alcântara, M.G.; Lins Freire, N.M.; Brandão, E.M.; do Nascimento, V.L.; dos Santos Viana, L.M.; de Aquino, T.M.; da Silva-Júnior, E.F. BACE-1 Inhibitors Targeting Alzheimer’s Disease. Curr. Alzheimer Res. 2023, 20, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.E.; Son, H.J.; Lim, D.W.; Yoon, M.; Lee, J.; Kim, Y.T.; Han, D.; Lee, C.; Um, M.Y. Memory-Enhancing Effects of Ishige foliacea Extract: In Vitro and in Vivo Study. J. Food Biochem. 2020, 44, e13162. [Google Scholar] [CrossRef] [PubMed]
- Dhanabalan, A.K.; Vasudevan, S.; Velmurugan, D.; Khan, M.S. Identification of Potential Marine Bioactive Compounds from Brown Seaweeds towards BACE1 Inhibitors: Molecular Docking and Molecular Dynamics Simulations Approach. In Silico Pharmacol. 2024, 12, 40. [Google Scholar] [CrossRef]
- Dezsi, L.; Vecsei, L. Monoamine Oxidase B Inhibitors in Parkinson’s Disease. CNS Neurol. Disord. Drug Targets 2017, 16, 425–439. [Google Scholar] [CrossRef]
- Kwon, J.; Lee, K.; Hwang, H.; Kim, S.H.; Park, S.E.; Durai, P.; Park, K.; Kim, H.S.; Jang, D.S.; Choi, J.S.; et al. New Monocyclic Terpenoid Lactones from a Brown Algae Sargassum macrocarpum as Monoamine Oxidase Inhibitors. Plants 2022, 11, 1998. [Google Scholar] [CrossRef]
- Tuppo, E.E.; Arias, H.R. The Role of Inflammation in Alzheimer’s Disease. Int. J. Biochem. Cell Biol. 2005, 37, 289–305. [Google Scholar] [CrossRef]
- Dhapola, R.; Beura, S.K.; Sharma, P.; Singh, S.K.; HariKrishnaReddy, D. Oxidative Stress in Alzheimer’s Disease: Current Knowledge of Signaling Pathways and Therapeutics. Mol. Biol. Rep. 2024, 51, 48. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, G.; Jin, S.; Jian, R.; Zou, Z.; Wang, C.; Zhang, Y.; Zhao, M.; Zhu, H.; Yan, P. Neuroprotective Effect of Acetoxypachydiol Against Oxidative Stress Through Activation of the Keap1-Nrf2/HO-1 Pathway. BMC Complement. Med. Ther. 2024, 24, 175. [Google Scholar] [CrossRef]
- Shanmuganathan, B.; Sathya, S.; Balasubramaniam, B.; Balamurugan, K.; Devi, K.P. Amyloid-β Induced Neuropathological Actions Are Suppressed by Padina gymnospora (Phaeophyceae) and Its Active Constituent α-Bisabolol in Neuro2a Cells and Transgenic Caenorhabditis elegans Alzheimer’s Model. Nitric Oxide 2019, 91, 52–66. [Google Scholar] [CrossRef]
- Meshalkina, D.; Tsvetkova, E.; Orlova, A.; Islamova, R.; Grashina, M.; Gorbach, D.; Babakov, V.; Francioso, A.; Birkemeyer, C.; Mosca, L.; et al. First Insight into the Neuroprotective and Antibacterial Effects of Phlorotannins Isolated from the Cell Walls of Brown Algae Fucus vesiculosus and Pelvetia canaliculata. Antioxidants 2023, 12, 696. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Neuroprotective Effects of Some Seaweeds against Zn—Induced Neuronal Damage in HT-22 Cells via Modulation of Redox Imbalance, Inhibition of Apoptosis and Acetylcholinesterase Activity. Metab. Brain Dis. 2019, 34, 1615–1627. [Google Scholar] [CrossRef] [PubMed]
- Alghazwi, M.; Charoensiddhi, S.; Smid, S.; Zhang, W. Impact of Ecklonia radiata Extracts on the Neuroprotective Activities against Amyloid Beta (Aβ1–42) Toxicity and Aggregation. J. Funct. Foods 2020, 68, 103893. [Google Scholar] [CrossRef]
- Shrestha, S.; Choi, J.S.; Zhang, W.; Smid, S.D. Neuroprotective Activity of Macroalgal Fucofuroeckols against Amyloid β Peptide-Induced Cell Death and Oxidative Stress. Int. J. Food Sci. Technol. 2022, 57, 4286–4295. [Google Scholar] [CrossRef]
- Cui, Y.; Amarsanaa, K.; Lee, J.H.; Rhim, J.K.; Kwon, J.M.; Kim, S.H.; Park, J.M.; Jung, S.C.; Eun, S.Y. Neuroprotective Mechanisms of Dieckol against Glutamate Toxicity through Reactive Oxygen Species Scavenging and Nuclear Factor-like 2/Heme Oxygenase-1 Pathway. Korean J. Physiol. Pharmacol. 2019, 23, 121–130. [Google Scholar] [CrossRef]
- Barbosa, M.; Valentão, P.; Ferreres, F.; Gil-Izquierdo, Á.; Andrade, P.B. In Vitro Multifunctionality of Phlorotannin Extracts from Edible Fucus species on Targets Underpinning Neurodegeneration. Food Chem. 2020, 333, 127456. [Google Scholar] [CrossRef]
- Lee, J.; Jun, M. Dual BACE1 and Cholinesterase Inhibitory Effects of Phlorotannins from Ecklonia cava—An In Vitro and in Silico Study. Mar. Drugs 2019, 17, 91. [Google Scholar] [CrossRef]
- Kirindage, K.G.I.S.; Jayasinghe, A.M.K.; Han, E.J.; Jee, Y.; Kim, H.J.; Do, S.G.; Fernando, I.P.S.; Ahn, G. Fucosterol Isolated from Dietary Brown Alga Sargassum horneri Protects TNF-α/IFN-γ-Stimulated Human Dermal Fibroblasts Via Regulating Nrf2/HO-1 and NF-ΚB/MAPK Pathways. Antioxidants 2022, 11, 1429. [Google Scholar] [CrossRef]
- Gan, S.Y.; Wong, L.Z.; Wong, J.W.; Tan, E.L. Fucosterol Exerts Protection against Amyloid β-Induced Neurotoxicity, Reduces Intracellular Levels of Amyloid β and Enhances the MRNA Expression of Neuroglobin in Amyloid β-Induced SH-SY5Y Cells. Int. J. Biol. Macromol. 2019, 121, 207–213. [Google Scholar] [CrossRef]
- Silva, J.; Alves, C.; Pinteus, S.; Susano, P.; Simões, M.; Guedes, M.; Martins, A.; Rehfeldt, S.; Gaspar, H.; Goettert, M.; et al. Disclosing the Potential of Eleganolone for Parkinson’s Disease Therapeutics: Neuroprotective and Anti-Inflammatory Activities. Pharmacol. Res. 2021, 168, 105589. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, G.; Fang, C.; Jing, C.; Tang, S.; Li, G.; Wang, C.; Zhu, H.; Zhao, M.; Sun, Z.; et al. Antioxidant and Neuroprotective Xenicane Diterpenes from the Brown Alga Dictyota coriacea. ACS Omega 2023, 8, 8034–8044. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xi, Y.; Li, G.; Zheng, Y.; Wang, Z.; Wang, J.; Fang, C.; Sun, Z.; Hu, L.; Jiang, W.; et al. Hydroazulene Diterpenes from A Dictyota Brown Alga and Their Antioxidant and Neuroprotective Effects against Cerebral Ischemia-Reperfusion Injury. J. Nat. Prod. 2021, 84, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, M.; Matsuda, A.; Shiiba, N.; Tsuruta, T.; Endo, H.; Nishikawa, K.; Morimoto, Y. Structure-Activity Relationship of Anti-Inflammatory Meroterpenoids Isolated from Dictyopteris polypodioides in RAW264 Cells. Biosci. Biotechnol. Biochem. 2024, 88, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Li, G.; Liu, Y.; Yang, L.; Zhang, Y.; Zhang, Y.; Ding, J.; Lu, M.; Yu, G.; Hu, G. Fucoidan from Fucus vesiculosus Prevents the Loss of Dopaminergic Neurons by Alleviating Mitochondrial Dysfunction through Targeting ATP5F1a. Carbohydr. Polym. 2023, 303, 120470. [Google Scholar] [CrossRef]
- Subermaniam, K.; Lew, S.Y.; Yow, Y.Y.; Lim, S.H.; Yu, W.S.; Lim, L.W.; Wong, K.H. Malaysian Brown Macroalga Padina australis Mitigates Lipopolysaccharide-Stimulated Neuroinflammation in BV2 Microglial Cells. Iran. J. Basic. Med. Sci. 2023, 26, 669. [Google Scholar] [CrossRef]
- Fei, J.; Yamada, S.; Satoh, T.; Koyama, T. Pharmacokinetic Analysis of Zonarol, a Marine Algal Hydroquinone, in Mice Using HPLC with Fluorescence Detection. Antibiotics 2023, 12, 1013. [Google Scholar] [CrossRef]
- Jo, S.L.; Yang, H.; Jeong, K.J.; Lee, H.W.; Hong, E.J. Neuroprotective Effects of Ecklonia cava in a Chronic Neuroinflammatory Disease Model. Nutrients 2023, 15, 2007. [Google Scholar] [CrossRef]
- Yende, S.R.; Arora, S.K.; Ittadwar, A.M. Antioxidant and Cognitive Enhancing Activities of Sargassum ilicifolium and Padina tetrastromatica in Scopolamine Treated Mice. J. Biol. Act. Prod. Nat. 2021, 11, 11–21. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Duan, L.; Li, X.; Yang, W.; Huang, T.; Kong, M.; Guan, F.; Ma, S. Fucoidan Ameliorates LPS-Induced Neuronal Cell Damage and Cognitive Impairment in Mice. Int. J. Biol. Macromol. 2022, 222, 759–771. [Google Scholar] [CrossRef]
- Ramu, S.; Anbu, J.; Ammunje, D.N.; Krishnaraj, K. Fucoidan Isolated from Sargassum wightii Greville Ameliorates Intracerebro-Ventricular Streptozotocin Induced Cognitive Deficits, Oxidative Stress and Amyloidosis in Wistar Rats. Bioact. Carbohydr. Diet. Fibre 2022, 27, 100309. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, Z.; Feng, X.; Deng, J.; He, C.; Li, R.; Zhao, Y.; Ge, Y.; Zhang, Y.; Song, C.; et al. The Emerging Evidence for a Protective Role of Fucoidan from Laminaria Japonica in Chronic Kidney Disease-Triggered Cognitive Dysfunction. Mar. Drugs 2022, 20, 258. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ahn, J.H.; Song, M.; Kim, D.W.; Lee, T.K.; Lee, J.C.; Kim, Y.M.; Kim, J.D.; Cho, J.H.; Hwang, I.K.; et al. Pretreated Fucoidan Confers Neuroprotection against Transient Global Cerebral Ischemic Injury in the Gerbil Hippocampal CA1 Area via Reducing of Glial Cell Activation and Oxidative Stress. Biomed. Pharmacother. 2019, 109, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Ahn, J.H.; Park, C.W.; Kim, B.; Park, Y.E.; Lee, J.C.; Park, J.H.; Yang, G.E.; Shin, M.C.; Cho, J.H.; et al. Pre-Treatment with Laminarin Protects Hippocampal CA1 Pyramidal Neurons and Attenuates Reactive Gliosis Following Transient Forebrain Ischemia in Gerbils. Mar. Drugs 2020, 18, 52. [Google Scholar] [CrossRef] [PubMed]
- Janahmadi, Z.; Jaberie, H.; Esmaili, A.; Nabipour, I. Hepatoprotective Effects of Brown Algae Sargassum Boveanum on Bile Duct-Ligated Cholestasis in Rats Are Mediated by Modulating NF-ΚB/TNF-α and Nrf2/HO-1 Gene Expression. Avicenna J. Phytomed. 2023, 13, 513–530. [Google Scholar] [CrossRef]
- Hassanzadeh, A.; Yegdaneh, A.; Rabbani, M. Effects of Hydroalcoholic, Methanolic, and Hexane Extracts of Brown Algae Sargassum Angustifolium on Scopolamine-Induced Memory Impairment and Learning Deficit in Rodents. Res Pharm Sci 2023, 18, 292–302. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cokdinleyen, M.; dos Santos, L.C.; de Andrade, C.J.; Kara, H.; Colás-Ruiz, N.R.; Ibañez, E.; Cifuentes, A. A Narrative Review on the Neuroprotective Potential of Brown Macroalgae in Alzheimer’s Disease. Nutrients 2024, 16, 4394. https://doi.org/10.3390/nu16244394
Cokdinleyen M, dos Santos LC, de Andrade CJ, Kara H, Colás-Ruiz NR, Ibañez E, Cifuentes A. A Narrative Review on the Neuroprotective Potential of Brown Macroalgae in Alzheimer’s Disease. Nutrients. 2024; 16(24):4394. https://doi.org/10.3390/nu16244394
Chicago/Turabian StyleCokdinleyen, Melis, Luana Cristina dos Santos, Cristiano José de Andrade, Huseyin Kara, Nieves R. Colás-Ruiz, Elena Ibañez, and Alejandro Cifuentes. 2024. "A Narrative Review on the Neuroprotective Potential of Brown Macroalgae in Alzheimer’s Disease" Nutrients 16, no. 24: 4394. https://doi.org/10.3390/nu16244394
APA StyleCokdinleyen, M., dos Santos, L. C., de Andrade, C. J., Kara, H., Colás-Ruiz, N. R., Ibañez, E., & Cifuentes, A. (2024). A Narrative Review on the Neuroprotective Potential of Brown Macroalgae in Alzheimer’s Disease. Nutrients, 16(24), 4394. https://doi.org/10.3390/nu16244394