
Caffeine-Induced Upregulation of pas-1 and pas-3 Enhances Intestinal Integrity by Reducing Vitellogenin in Aged Caenorhabditis elegans Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Caenorhabditis elegans Strains and Caffeine Treatment
2.2. RNA Sequencing and Data Visualization
2.3. RNA Extraction and Quantitative Reverse Transcriptase-PCR (qRT-PCR)
2.4. RNA Interference (RNAi)
2.5. Intestinal Integrity Assays
2.6. Motility and Life Span Assays
2.7. Statistical Analysis
3. Results
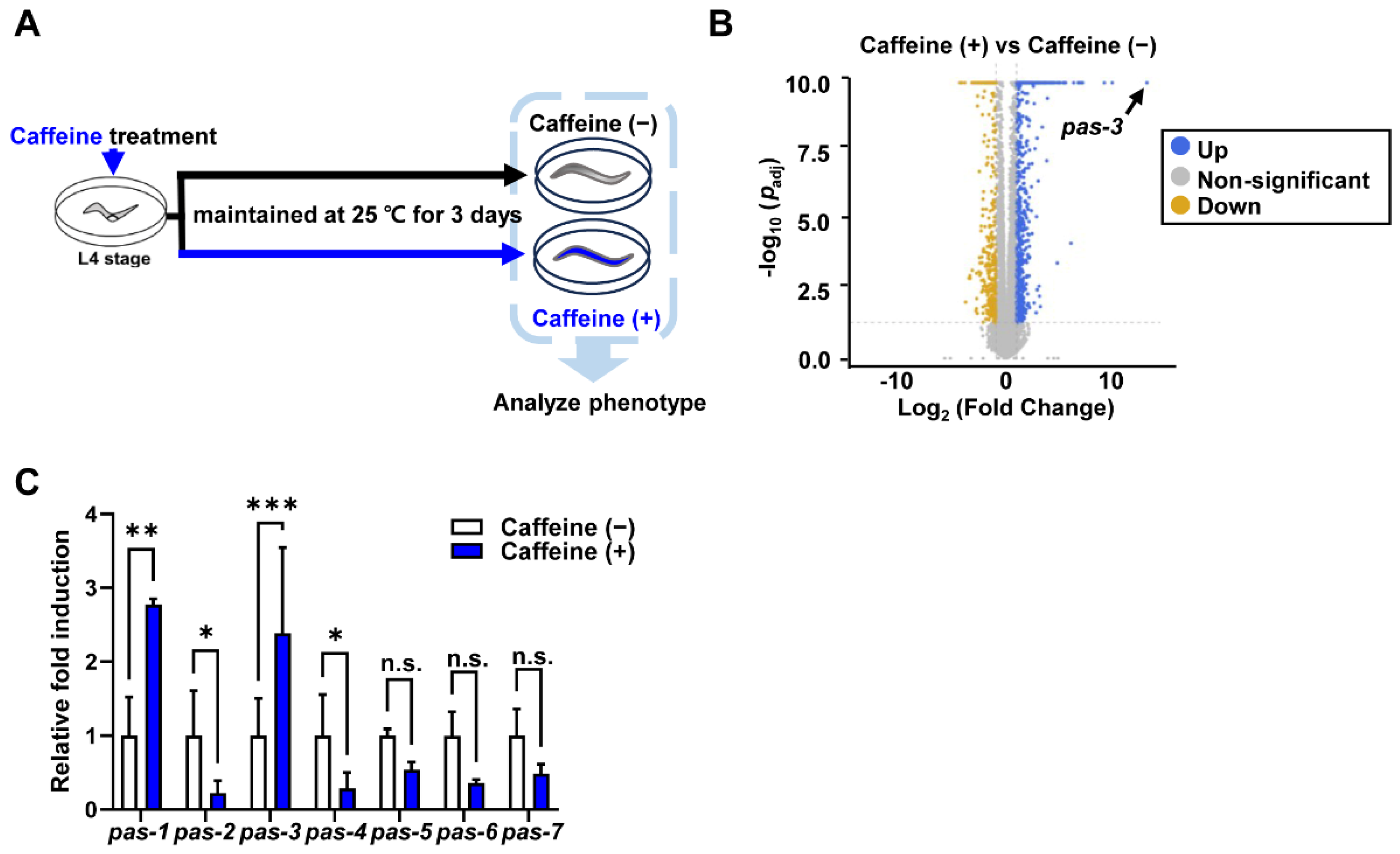
3.1. Caffeine Intake Significantly Increases Expression of Proteasome α-Subunit Genes, pas-1 and pas-3 in aged C. elegans
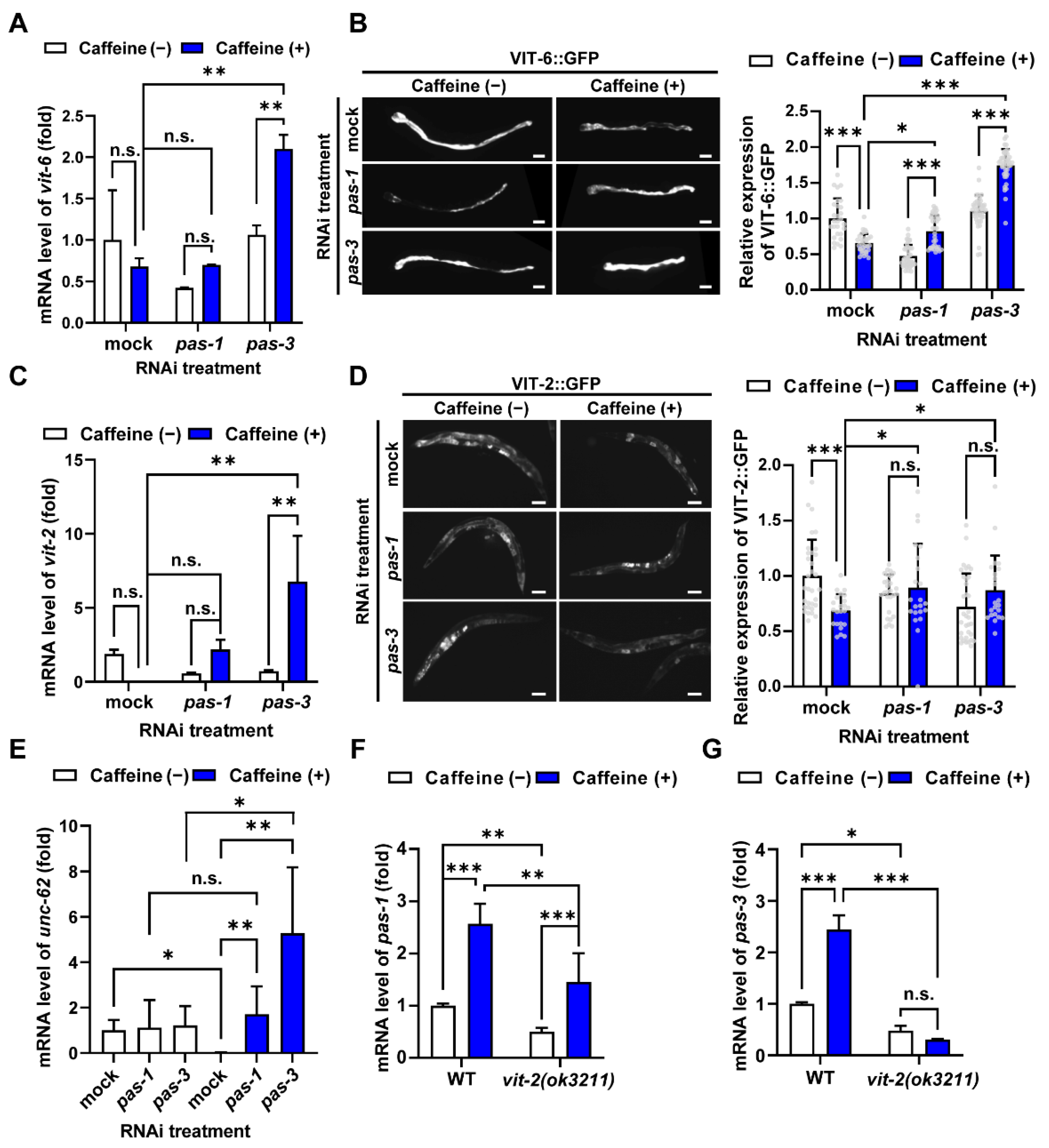
3.2. Caffeine Intake Reduces Vitellogenin Production by Modulating pas-1 and pas-3 During Aging in C. elegans
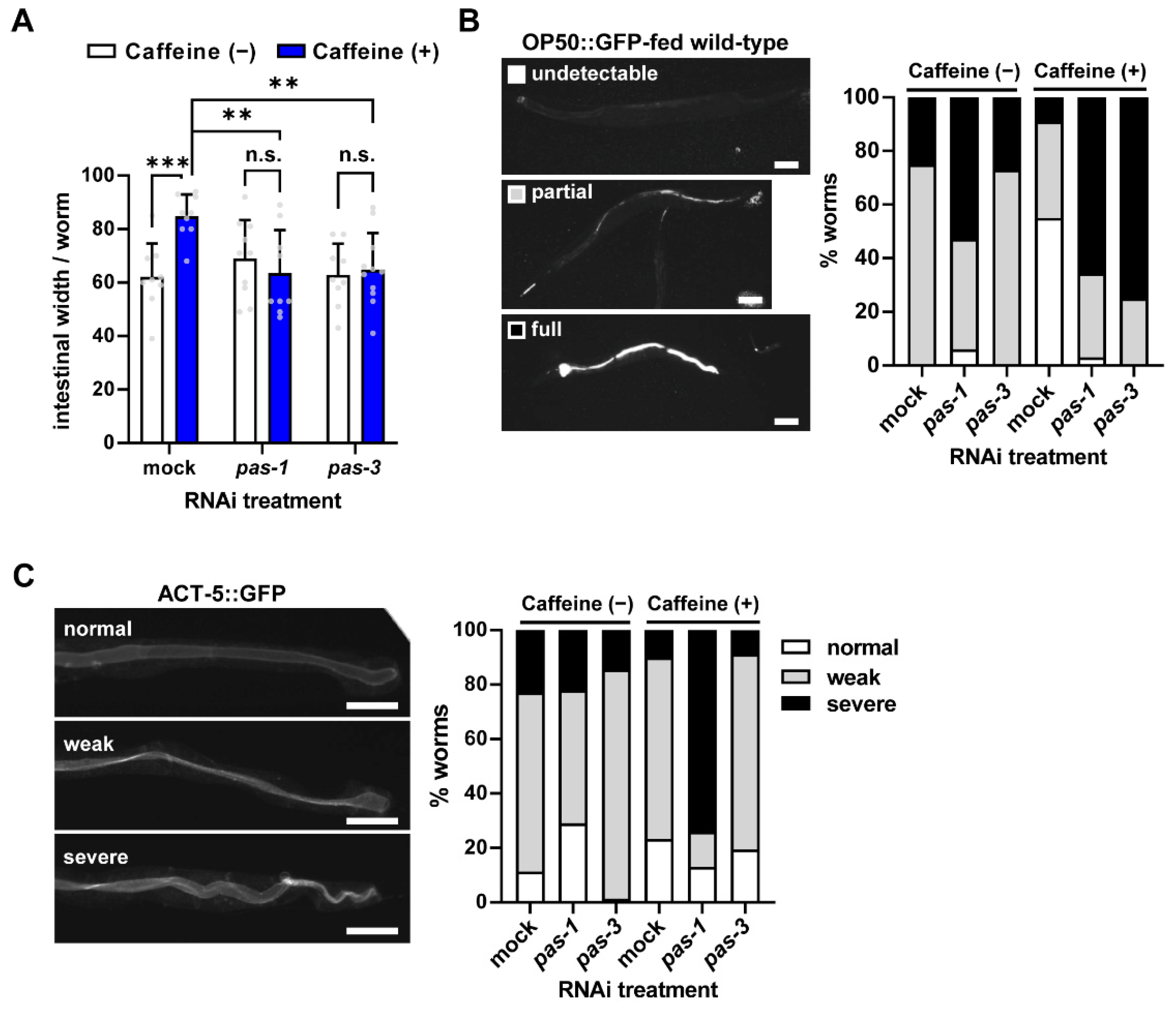
3.3. Caffeine Intake Improves Intestinal Integrity via pas-1 and pas-3 in Aged Adults
3.4. pas-1 Extends SKN-1-Mediated Lifespan During Aging in Caffeine-Fed Aged C. elegans
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grubben, M.J.; Van Den Braak, C.C.; Broekhuizen, R.; De Jong, R.; Van Rijt, L.; De Ruijter, E.; Peters, W.H.; Katan, M.B.; Nagengast, F.M. The Effect of Unfiltered Coffee on Potential Biomarkers for Colonic Cancer Risk in Healthy Volunteers: A Randomized Trial. Aliment. Pharmacol. Ther. 2000, 14, 1181–1190. [Google Scholar]
- Arnaud, M.J. The Pharmacology of Caffeine. Prog. Drug Res. 1987, 31, 273–313. [Google Scholar]
- Reyes, C.M.; Cornelis, M.C. Caffeine in the Diet: Country-Level Consumption and Guidelines. Nutrients 2018, 10, 1772. [Google Scholar] [CrossRef]
- Saraiva, S.M.; Jacinto, T.A.; Gonçalves, A.C.; Gaspar, D.; Silva, L.R. Overview of Caffeine Effects on Human Health and Emerging Delivery Strategies. Pharmaceuticals 2023, 16, 1067. [Google Scholar] [CrossRef]
- Min, H.; Kawasaki, I.; Gong, J.; Shim, Y.H. Caffeine Induces High Expression of cyp-35A Family Genes and Inhibits the Early Larval Development in Caenorhabditis elegans. Mol. Cells 2015, 38, 236–242. [Google Scholar]
- Nawrot, P.; Jordan, S.; Eastwood, J.; Rotstein, J.; Hugenholtz, A.; Feeley, M. Effects of Caffeine on Human Health. Food Addit. Contam. 2003, 20, 1–30. [Google Scholar]
- Gavrieli, A.; Yannakoulia, M.; Fragopoulou, E.; Margaritopoulos, D.; Chamberland, J.P.; Kaisari, P.; Kavouras, S.A.; Mantzoros, C.S. Caffeinated Coffee Does Not Acutely Affect Energy Intake, Appetite, or Inflammation but Prevents Serum Cortisol Concentrations from Falling in Healthy Men. J. Nutr. 2011, 141, 703–707. [Google Scholar]
- Min, H.; Youn, E.; Shim, Y.H. Maternal Caffeine Intake Disrupts Eggshell Integrity and Retards Larval Development by Reducing Yolk Production in a Caenorhabditis elegans Model. Nutrients 2020, 12, 1334. [Google Scholar] [CrossRef]
- Takahashi, K.; Yanai, S.; Shimokado, K.; Ishigami, A. Coffee Consumption in Aged Mice Increases Energy Production and Decreases Hepatic MTOR Levels. Nutrition 2017, 38, 1–8. [Google Scholar]
- Min, H.; Youn, E.; Shim, Y.H. Long-Term Caffeine Intake Exerts Protective Effects on Intestinal Aging by Regulating Vitellogenesis and Mitochondrial Function in an Aged Caenorhabditis elegans Model. Nutrients 2021, 13, 2517. [Google Scholar] [CrossRef]
- Min, H.; Lee, M.; Kang, S.; Shim, Y.H. Vitamin B12 Supplementation Improves Oocyte Development by Modulating Mitochondria and Yolk Protein in a Caffeine-Ingested Caenorhabditis elegans Model. Antioxidants 2024, 13, 53. [Google Scholar]
- Sheng, X.; Zhu, Y.; Zhou, J.; Yan, L.; Du, G.; Liu, Z.; Chen, H. Antioxidant Effects of Caffeic Acid Lead to Protection of Drosophila Intestinal Stem Cell Aging. Front. Cell Dev. Biol. 2021, 9, 735483. [Google Scholar]
- González, S.; Salazar, N.; Ruiz-Saavedra, S.; Gómez-Martín, M.; de los Reyes-Gavilán, C.G.; Gueimonde, M. Long-Term Coffee Consumption Is Associated with Fecal Microbial Composition in Humans. Nutrients 2020, 12, 1287. [Google Scholar] [CrossRef]
- Baspinar, B.; Eskici, G.; Ozcelik, A.O. How Coffee Affects Metabolic Syndrome and Its Components. Food Funct. 2017, 8, 2089–2101. [Google Scholar]
- O’Keefe, J.H.; DiNicolantonio, J.J.; Lavie, C.J. Coffee for Cardioprotection and Longevity. Prog. Cardiovasc. Dis. 2018, 61, 38–42. [Google Scholar]
- Gkegkes, I.D.; Minis, E.E.; Iavazzo, C. Effect of Caffeine Intake on Postoperative Ileus: A Systematic Review and Meta-Analysis. Dig. Surg. 2020, 37, 22–31. [Google Scholar]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of Diet on the Gut Microbiome and Implications for Human Health. J. Transl. Med. 2017, 15, 73. [Google Scholar]
- Li, Z.; Zhang, Z.; Ren, Y.; Wang, Y.; Fang, J.; Yue, H.; Ma, S.; Guan, F. Aging and Age-related Diseases: From Mechanisms to Therapeutic Strategies. Biogerontology 2021, 22, 165–187. [Google Scholar]
- Lee, K.X.; Quek, K.F.; Ramadas, A. Dietary and Lifestyle Risk Factors of Obesity Among Young Adults: A Scoping Review of Observational Studies. Curr. Nutr. Rep. 2023, 12, 733–743. [Google Scholar]
- Zhang, K.; Ma, Y.; Luo, Y.; Song, Y.; Xiong, G.; Ma, Y.; Sun, X.; Kan, C. Metabolic Diseases and Healthy Aging: Identifying Environmental and Behavioral Risk Factors and Promoting Public Health. Front. Public Health 2023, 11, 1253506. [Google Scholar]
- Surugiu, R.; Iancu, M.A.; Vintilescu, Ș.B.; Stepan, M.D.; Burdusel, D.; Genunche-Dumitrescu, A.V.; Dogaru, C.A.; Dumitra, G.G. Molecular Mechanisms of Healthy Aging: The Role of Caloric Restriction, Intermittent Fasting, Mediterranean Diet, and Ketogenic Diet—A Scoping Review. Nutrients 2024, 16, 2878. [Google Scholar] [CrossRef]
- Drozdowski, L.; Thomson, A.B. Aging and the Intestine. World J. Gastroenterol. 2006, 12, 7578–7584. [Google Scholar]
- McGee, M.D.; Weber, D.; Day, N.; Vitelli, C.; Crippen, D.; Herndon, L.A.; Hall, D.H.; Melov, S. Loss of Intestinal Nuclei and Intestinal Integrity in Aging C. elegans. Aging Cell 2011, 10, 699–710. [Google Scholar]
- Perez, M.F.; Lehner, B. Vitellogenins—Yolk Gene Function and Regulation in Caenorhabditis elegans. Front. Physiol. 2019, 10, 1067. [Google Scholar]
- Baker, M.E. Is Vitellogenin an Ancestor of Apolipoprotein B-100 of Human Low-Density Lipoprotein and Human Lipoprotein Lipase? Biochem. J. 1988, 255, 1057–1060. [Google Scholar]
- Kimbleand, J.; Sharrock, W.J. Tissue-Specific Synthesis of Yolk Proteins in Caenorhabditis elegans. Dev. Biol. 1983, 96, 189–196. [Google Scholar]
- Ezcurra, M.; Benedetto, A.; Sornda, T.; Gilliat, A.F.; Au, C.; Zhang, Q.; van Schelt, S.; Petrache, A.L.; Wang, H.; de la Guardia, Y.; et al. C. elegans Eats Its Own Intestine to Make Yolk Leading to Multiple Senescent Pathologies. Curr. Biol. 2018, 28, 2544–2556.e5. [Google Scholar]
- Sornda, T.; Ezcurra, M.; Kern, C.; Galimov, E.R.; Au, C.; De La Guardia, Y.; Gems, D. Production of YP170 Vitellogenins Promotes Intestinal Senescence in Caenorhabditis elegans. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 1180–1188. [Google Scholar] [CrossRef]
- Song, R.; Hu, M.; Qin, X.; Qiu, L.; Wang, P.; Zhang, X.; Liu, R.; Wang, X. The Roles of Lipid Metabolism in the Pathogenesis of Chronic Diseases in the Elderly. Nutrients 2023, 15, 3433. [Google Scholar] [CrossRef]
- Libina, N.; Berman, J.R.; Kenyon, C. Tissue-Specific Activities of C. elegans DAF-16 in the Regulation of Lifespan. Cell 2003, 115, 489–502. [Google Scholar]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Hoogewijs, D.; Houthoofd, K.; Matthijssens, F.; Vandesompele, J.; Vanfleteren, J.R. Selection and Validation of a Set of Reliable Reference Genes for Quantitative Sod Gene Expression Analysis in C. elegans. BMC Mol. Biol. 2008, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Maeda, I.; Kohara, Y.; Yamamoto, M.; Sugimoto, A. Large-Scale Analysis of Gene Function in Caenorhabditis elegans by High-Throughput RNAi. Curr. Biol. 2001, 11, 171–176. [Google Scholar] [CrossRef]
- Takahashi, M.; Iwasaki, H.; Inoue, H.; Takahashi, K. Reverse Genetic Analysis of the Caenorhabditis elegans 26S Proteasome Subunits by RNA Interference. Biol. Chem. 2002, 383, 1263–1266. [Google Scholar] [CrossRef]
- Walker, A.C.; Bhargava, R.; Vaziriyan-Sani, A.S.; Pourciau, C.; Donahue, E.T.; Dove, A.S.; Gebhardt, M.J.; Ellward, G.L.; Romeo, T.; Czyż, D.M. Colonization of the Caenorhabditis elegans gut with human enteric bacterial pathogens leads to proteostasis disruption that is rescued by butyrate. PLoS Pathog. 2021, 17, e1009510. [Google Scholar] [CrossRef]
- Davy, A.; Bello, P.; Thierry-Mieg, N.; Vaglio, P.; Hitti, J.; Doucette-Stamm, L.; Thierry-Mieg, D.; Reboul, J.; Boulton, S.; Walhout, A.J.; et al. A Protein-Protein Interaction Map of the Caenorhabditis elegans 26S Proteasome. EMBO Rep. 2001, 2, 821–828. [Google Scholar] [CrossRef]
- Papaevgeniou, N.; Chondrogianni, N. The Ubiquitin Proteasome System in Caenorhabditis elegans and Its Regulation. Redox Biol. 2014, 2, 333–347. [Google Scholar] [CrossRef]
- Vilchez, D.; Saez, I.; Dillin, A. The Role of Protein Clearance Mechanisms in Organismal Ageing and Age-Related Diseases. Nat. Commun. 2014, 5, 5659. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. Proteostasis and Aging. Nat. Med. 2015, 21, 1406–1415. [Google Scholar] [CrossRef]
- Dowen, R.H. CEH-60/PBX and UNC-62/MEIS Coordinate a Metabolic Switch That Supports Reproduction in C. elegans. Dev. Cell 2019, 49, 235–250.e7. [Google Scholar] [CrossRef]
- Macqueen, A.J.; Baggett, J.J.; Perumov, N.; Bauer, R.A.; Januszewski, T.; Schriefer, L.; Waddle, J.A. ACT-5 Is an Essential Caenorhabditis elegans Actin Required for Intestinal Microvilli Formation. Mol. Biol. Cell 2005, 16, 3247–3259. [Google Scholar] [PubMed]
- Anderson, R.T.; Bradley, T.A.; Smith, D.M. Hyperactivation of the Proteasome in Caenorhabditis elegans Protects against Proteotoxic Stress and Extends Lifespan. J. Biol. Chem. 2022, 298, 102415. [Google Scholar] [CrossRef] [PubMed]
- Tavirani, M.R.; Farahani, M.; Tavirani, M.R.; Razzaghi, Z.; Arjmand, B.; Khodadoost, M. Introducing Coffee as a Complementary Agent Beside Metformin Against Type 2 Diabetes. Res. J. Pharmacogn. 2024, 11, 31–40. [Google Scholar]
- Lim, H.H.; Kim, O.Y. Association of Serum Apolipoprotein B with the Increased Risk of Diabetes in Korean Men. Clin. Nutr. Res. 2016, 5, 204–212. [Google Scholar] [CrossRef]
- Hashmi, S.; Wang, Y.; Parhar, R.S.; Collison, K.S.; Conca, W.; Al-Mohanna, F. A C. elegans Model to Study Human Metabolic Regulation. Nutr. Metab. 2013, 10, 31. [Google Scholar]
- Mikkonen, E.; Haglund, C.; Holmberg, C.I. Immunohistochemical Analysis Reveals Variations in Proteasome Tissue Expression in C. elegans. PLoS ONE 2017, 12, e0183403. [Google Scholar] [CrossRef]
- Li, X.; Matilainen, O.; Jin, C.; Glover-Cutter, K.M.; Holmberg, C.I.; Blackwell, T.K. Specific SKN-1/NrF Stress Responses to Perturbations in Translation Elongation and Proteasome Activity. PLoS Genet. 2011, 7, e1002119. [Google Scholar]
- Lynn, D.A.; Dalton, H.M.; Sowa, J.N.; Wang, M.C.; Soukas, A.A.; Curran, S.P.; Ruvkun, G. Omega-3 and -6 Fatty Acids Allocate Somatic and Germline Lipids to Ensure Fitness during Nutrient and Oxidative Stress in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2015, 112, 15378–15383. [Google Scholar]
- Blackwell, T.K.; Steinbaugh, M.J.; Hourihan, J.M.; Ewald, C.Y.; Isik, M. SKN-1/Nrf, Stress Responses, and Aging in Caenorhabditis elegans. Free Radic. Biol. Med. 2015, 88, 290–301. [Google Scholar]
- Balklava, Z.; Pant, S.; Fares, H.; Grant, B.D. Genome-Wide Analysis Identifies a General Requirement for Polarity Proteins in Endocytic Traffic. Nat. Cell Biol. 2007, 9, 1066–1073. [Google Scholar]
- Saez, I.; Vilchez, D. The Mechanistic Links Between Proteasome Activity, Aging and Age-Related Diseases. Curr. Genom. 2014, 15, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Tomaru, U.; Takahashi, S.; Ishizu, A.; Miyatake, Y.; Gohda, A.; Suzuki, S.; Ono, A.; Ohara, J.; Baba, T.; Murata, S.; et al. Decreased Proteasomal Activity Causes Age-Related Phenotypes and Promotes the Development of Metabolic Abnormalities. Am. J. Pathol. 2012, 180, 963–972. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.; Lee, J.; Kim, D.; Min, H.; Shim, Y.-H. Caffeine-Induced Upregulation of pas-1 and pas-3 Enhances Intestinal Integrity by Reducing Vitellogenin in Aged Caenorhabditis elegans Model. Nutrients 2024, 16, 4298. https://doi.org/10.3390/nu16244298
Lee M, Lee J, Kim D, Min H, Shim Y-H. Caffeine-Induced Upregulation of pas-1 and pas-3 Enhances Intestinal Integrity by Reducing Vitellogenin in Aged Caenorhabditis elegans Model. Nutrients. 2024; 16(24):4298. https://doi.org/10.3390/nu16244298
Chicago/Turabian StyleLee, Mijin, Jea Lee, Dongyeon Kim, Hyemin Min, and Yhong-Hee Shim. 2024. "Caffeine-Induced Upregulation of pas-1 and pas-3 Enhances Intestinal Integrity by Reducing Vitellogenin in Aged Caenorhabditis elegans Model" Nutrients 16, no. 24: 4298. https://doi.org/10.3390/nu16244298
APA StyleLee, M., Lee, J., Kim, D., Min, H., & Shim, Y.-H. (2024). Caffeine-Induced Upregulation of pas-1 and pas-3 Enhances Intestinal Integrity by Reducing Vitellogenin in Aged Caenorhabditis elegans Model. Nutrients, 16(24), 4298. https://doi.org/10.3390/nu16244298