
Protective Effects of Spirulina Against Lipid Micelles and Lipopolysaccharide-Induced Intestinal Epithelium Disruption in Caco-2 Cells: In Silico Molecular Docking Analysis of Phycocyanobilin

 ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Spirulina Aqueous Extract Preparation (SPAE)
2.3. Cell Culture and Treatments
- Control: Caco-2 cells in DMEM without treatment.
- LM: Caco-2 cells incubated with LM (4 days) without treatment.
- LM + SPAE: Caco-2 cells incubated with LM (4 days) and treated with SPAE (500 µg/mL) for 24 h.
- SPAE: Caco-2 cells in DMEM treated with SPAE (500 µg/mL) for 24 h.
- LPS: Caco-2 cells incubated with LPS (1 µg/mL) for 3 h.
- LPS + SPAE: Caco-2 cells incubated with LPS (1 µg/mL) for 3 h and treated with SPAE (500 µg/mL) for 24 h.
- LPS + LM: Caco-2 cells incubated with LM (4 days) and LPS (1 µg/mL) for 3 h.
- LPS + LM + SPAE: Caco-2 cells incubated with LM (4 days) and LPS (1 µg/mL) for 3 h and treated with SPAE (500 µg/mL) for 24 h.
2.4. Preparation of Lipid Micelles
2.5. Intestinal Permeability Measurements in Caco-2 Cells
- Papp = apparent permeability (cm/s)
- dQ/dt = permeability rate (amount permeated per minute over the 240 min duration)
- A = area of monolayer diffusion (cm2)
- C0 = initial concentration of the studied compounds.
2.6. Viability/CytotoxicityWST-1assay
2.7. RNA Extraction
2.8. cDNA Synthesis
2.9. Gene Expression Quantification by Real-Time qPCR (RT-qPCR)
2.10. Biochemical Analysis of Oxidative Stress Markers
2.10.1. Protein Assay
2.10.2. Oxidative Stress Assessment
2.10.3. Reactive Oxygen Species Measurement
2.11. Quantification of Cytokines by ELISA
2.12. Molecular Docking Study
2.13. Statistical Analysis
3. Results
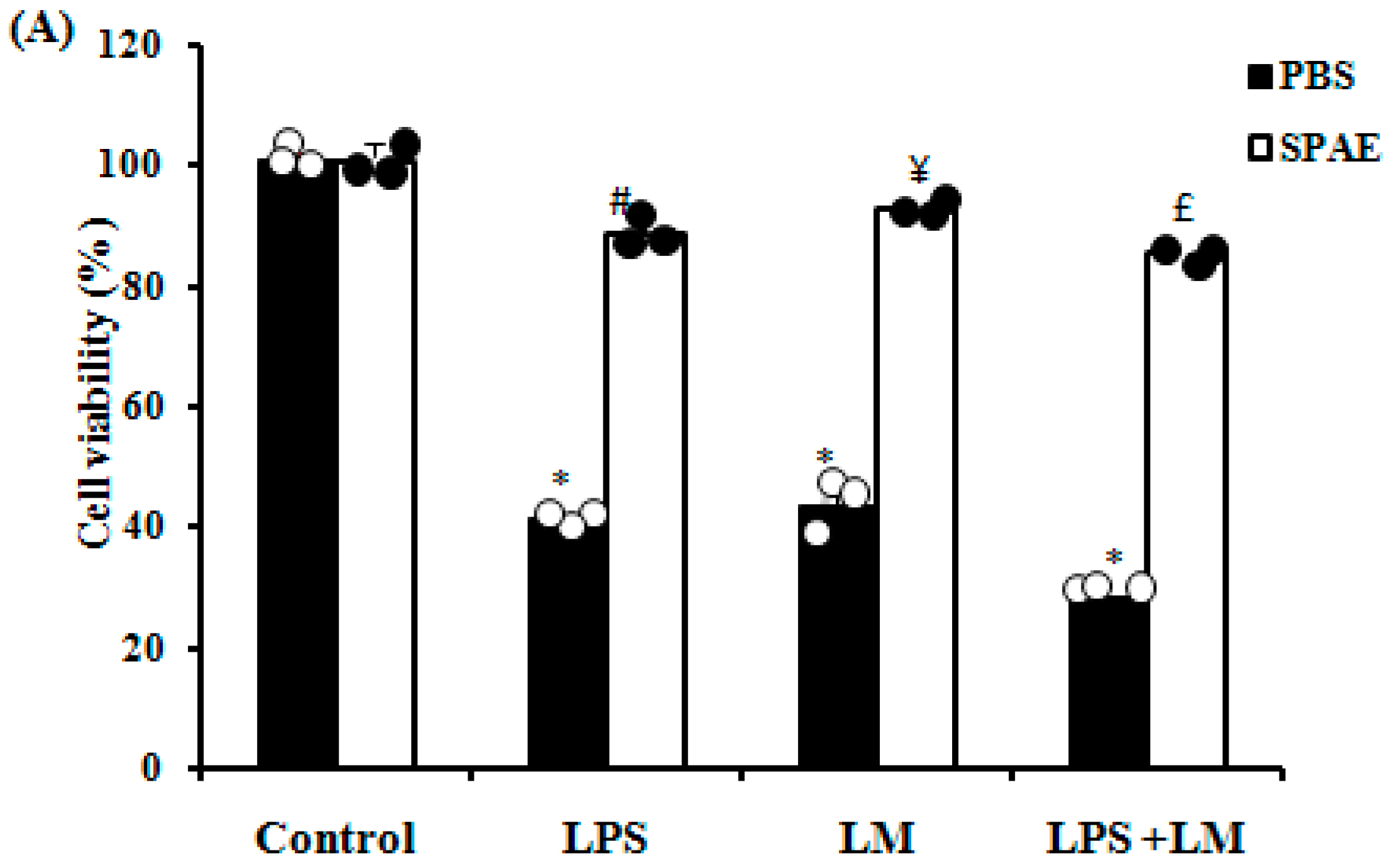
3.1. Effects of SPAE, LPS, and/or Lipid Micelles on Cell Viability
3.2. Effects of SPAE, LPS, and/or LM on Epithelial Barrier Permeability
3.3. Effects of SPAE, LPS, and/or Lipid Micelles on Intercellular Tight Junction Proteins
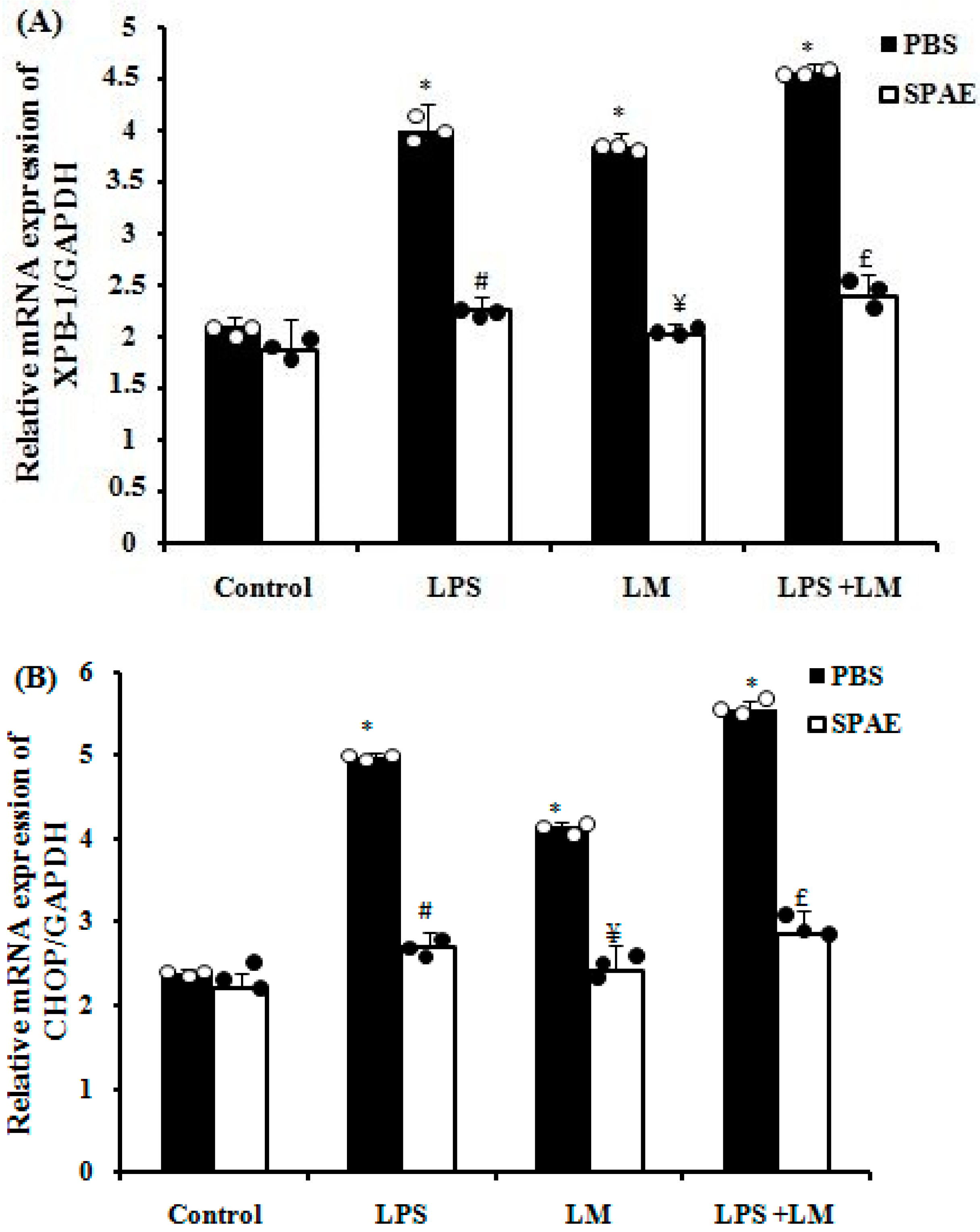
3.4. LPS and/or LM-Induced Increase in Endoplasmic Reticulum (ER) Stress Markers Expression in Caco-2 Cells Is Prevented by SPAE
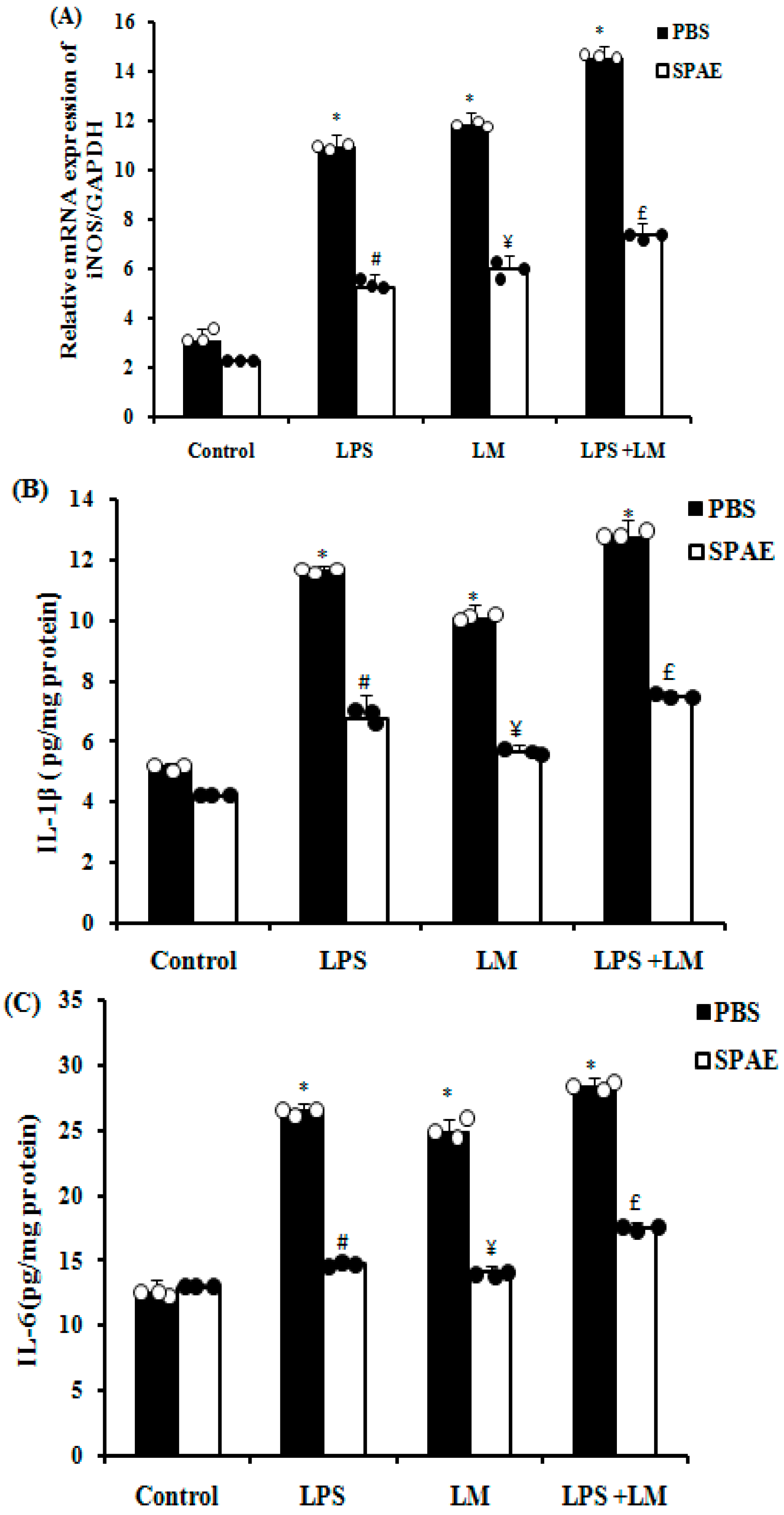
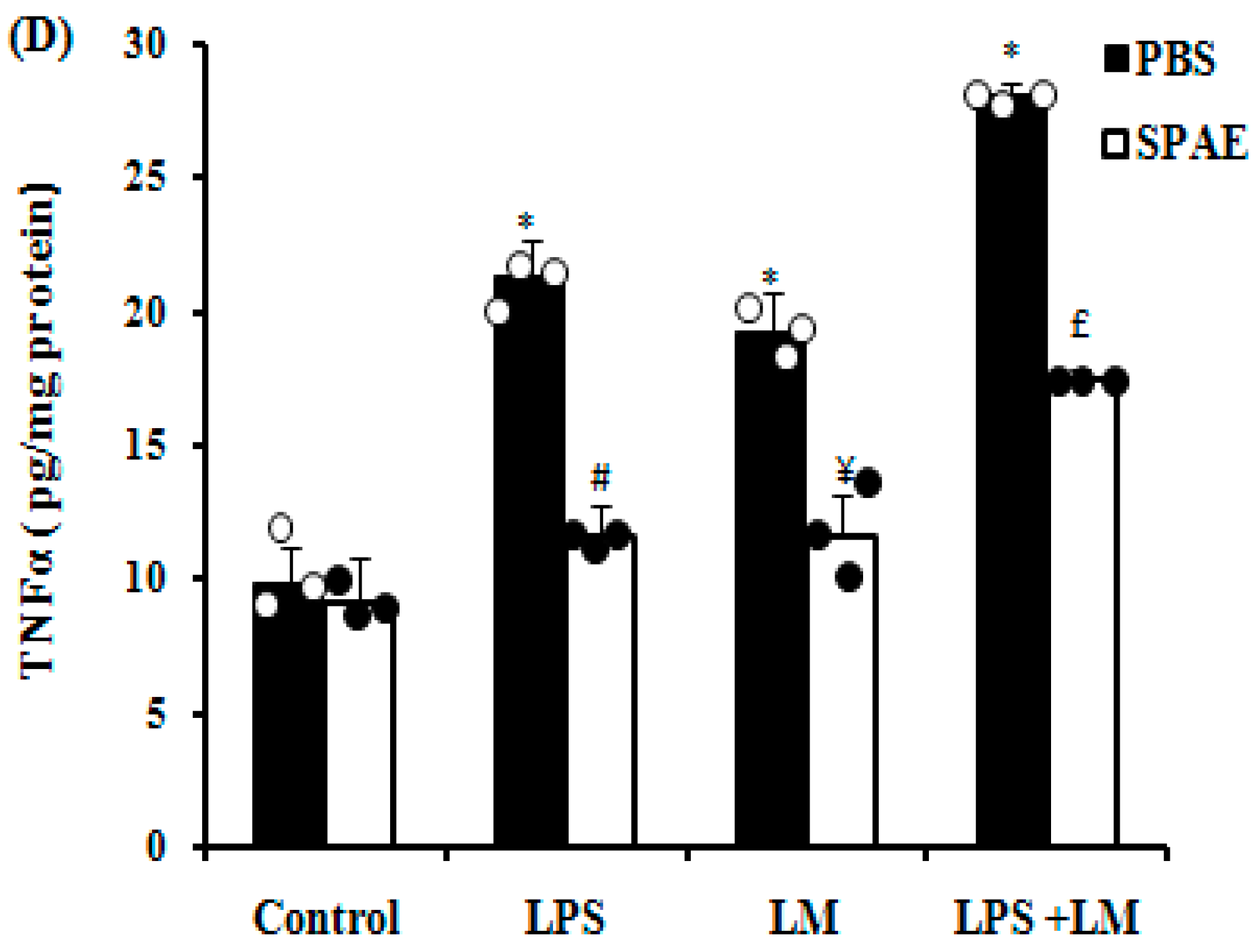
3.5. Effects of SPAE, LPS, and/or LM on Inducible Nitric Oxide Synthase (iNOS) and Pro-Inflammatory Cytokine Expression
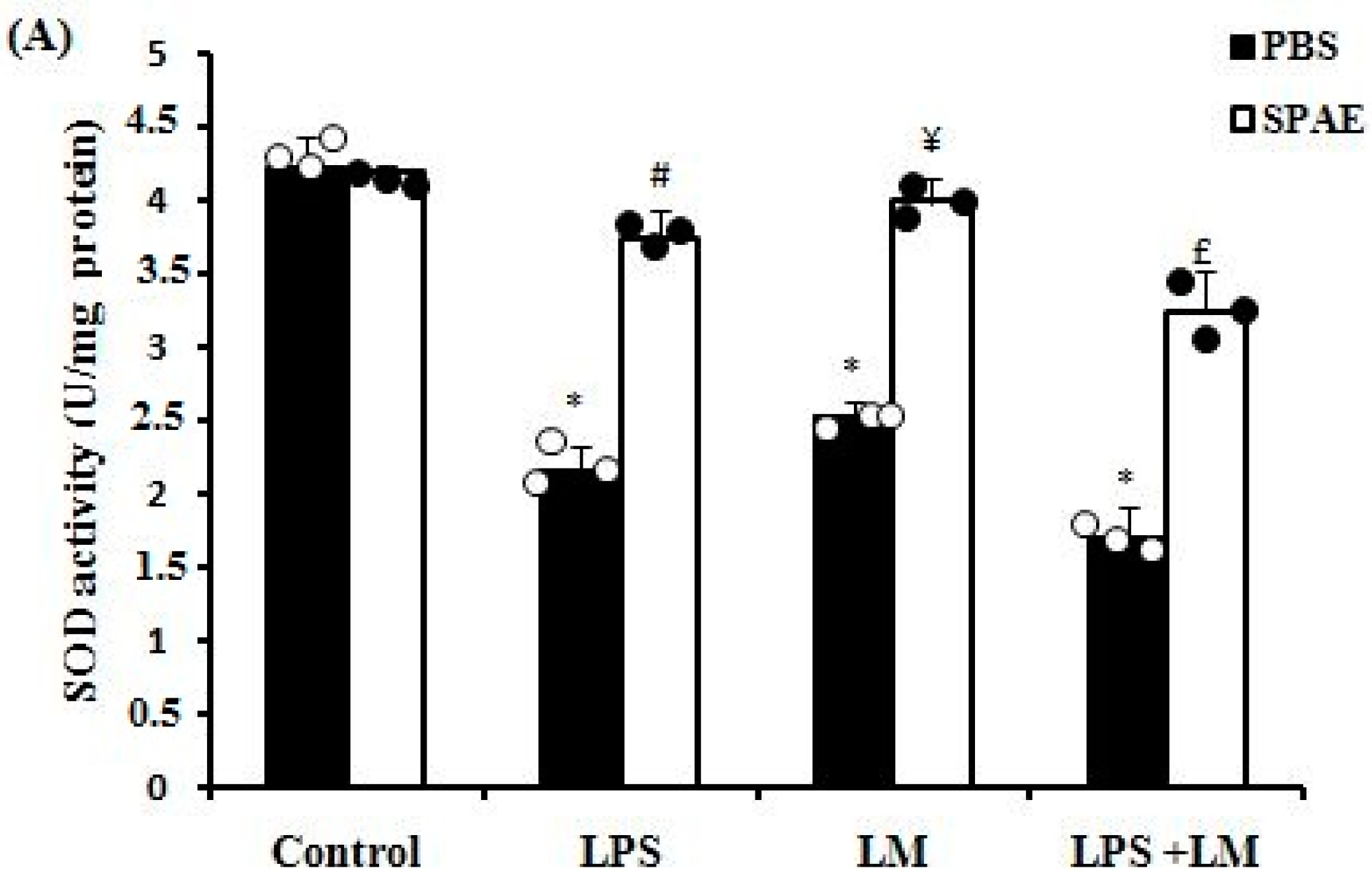
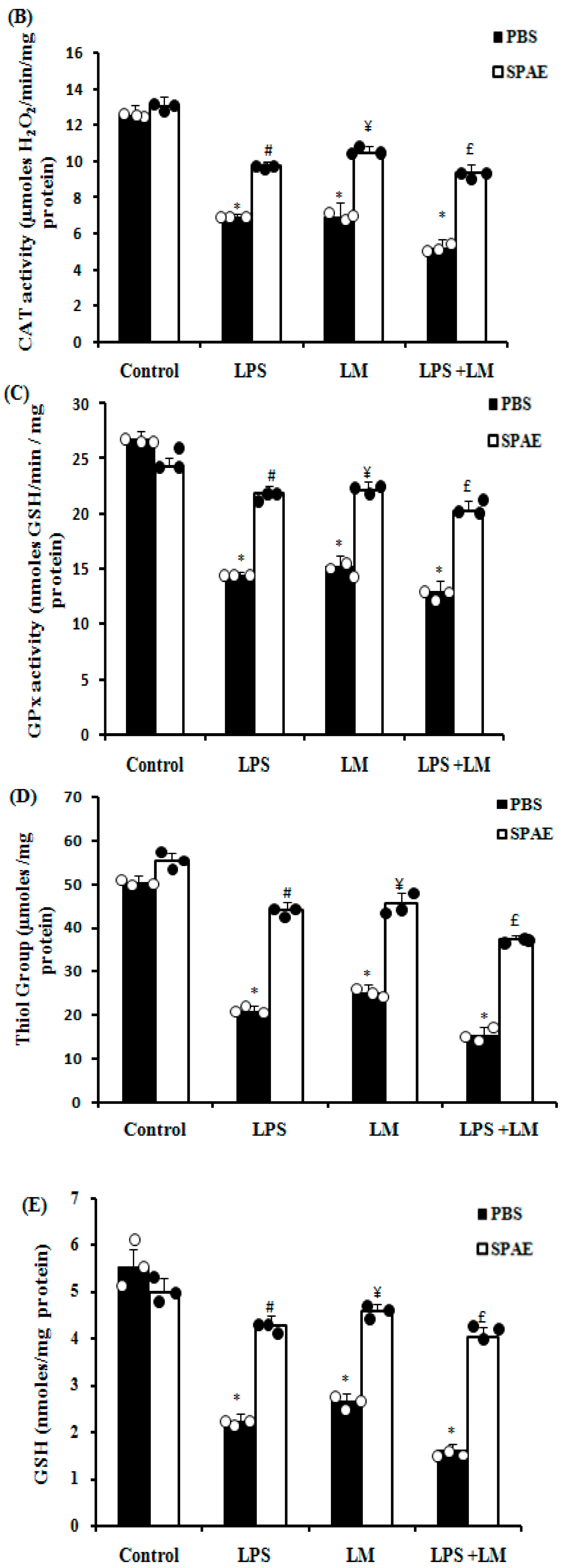
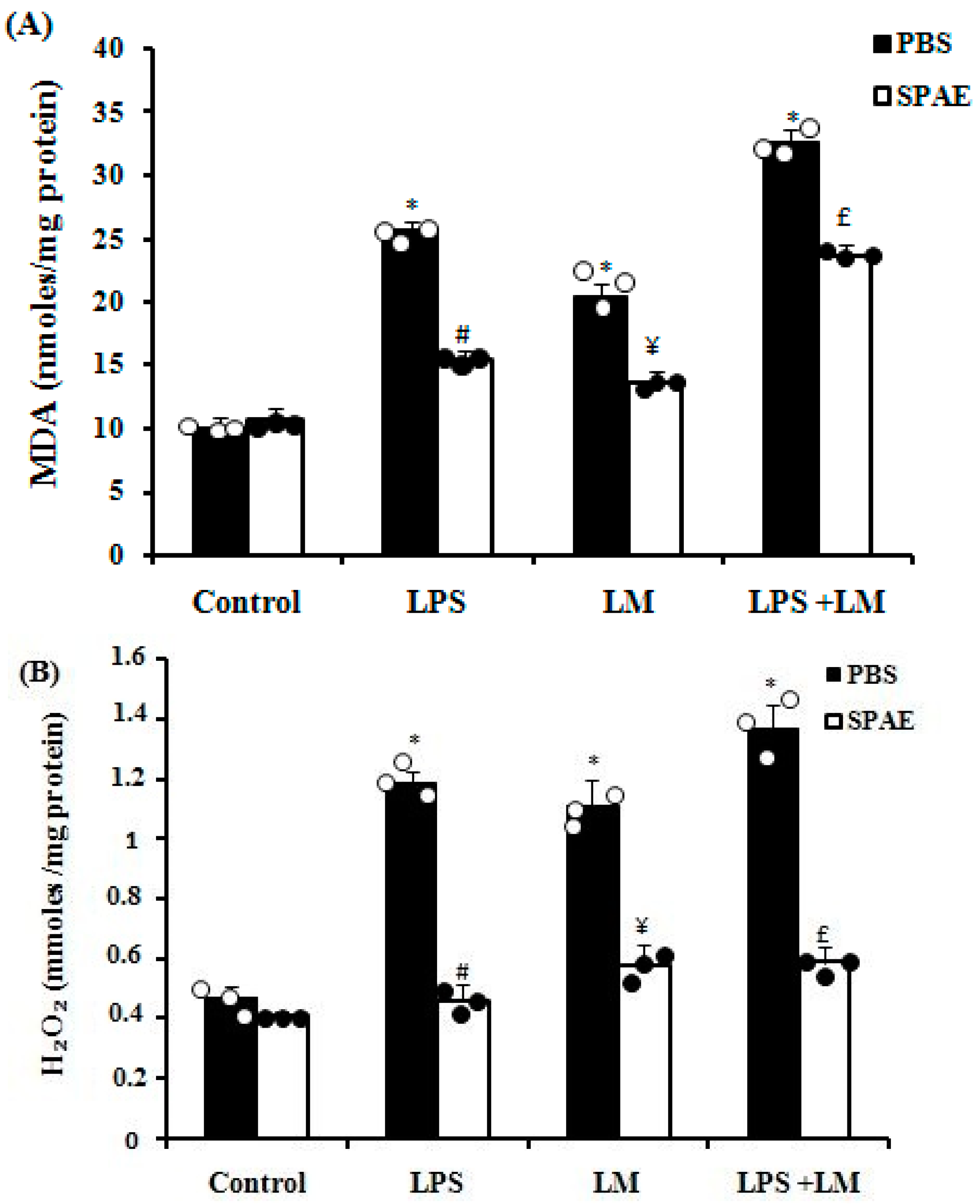
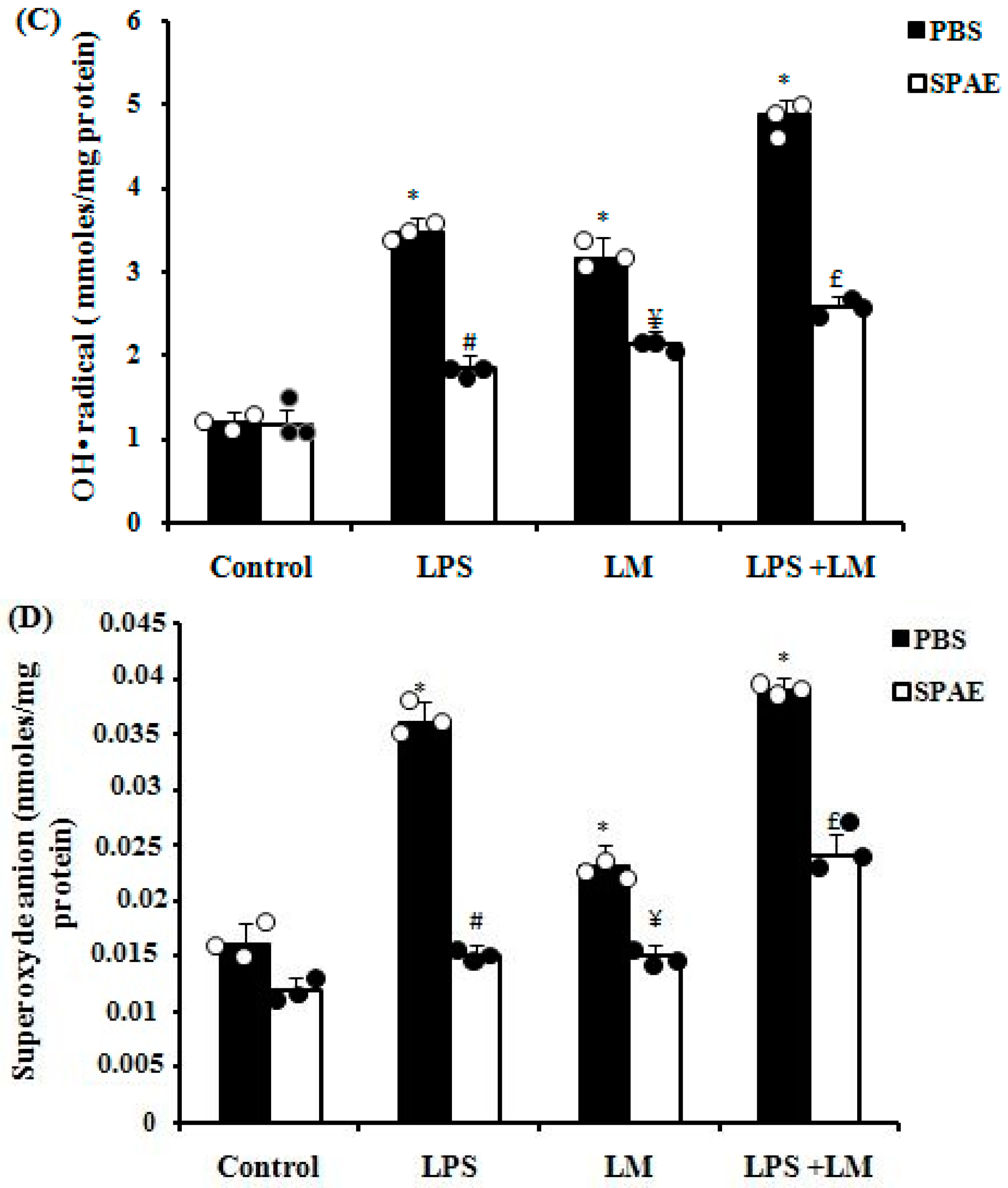
3.6. Effects of SPAE, LPS, and/or LM on Oxidative Parameters
3.6.1. Enzymatic Antioxidants (SOD, CAT, and GPx)
3.6.2. Non-Enzymatic Antioxidants (G-SH and GSH)
3.6.3. Lipid Peroxidation
3.6.4. Effects of SPAE, LPS, and/or LM on Reactive Oxygen Species (ROS) Production in Cultured Caco-2 Cells
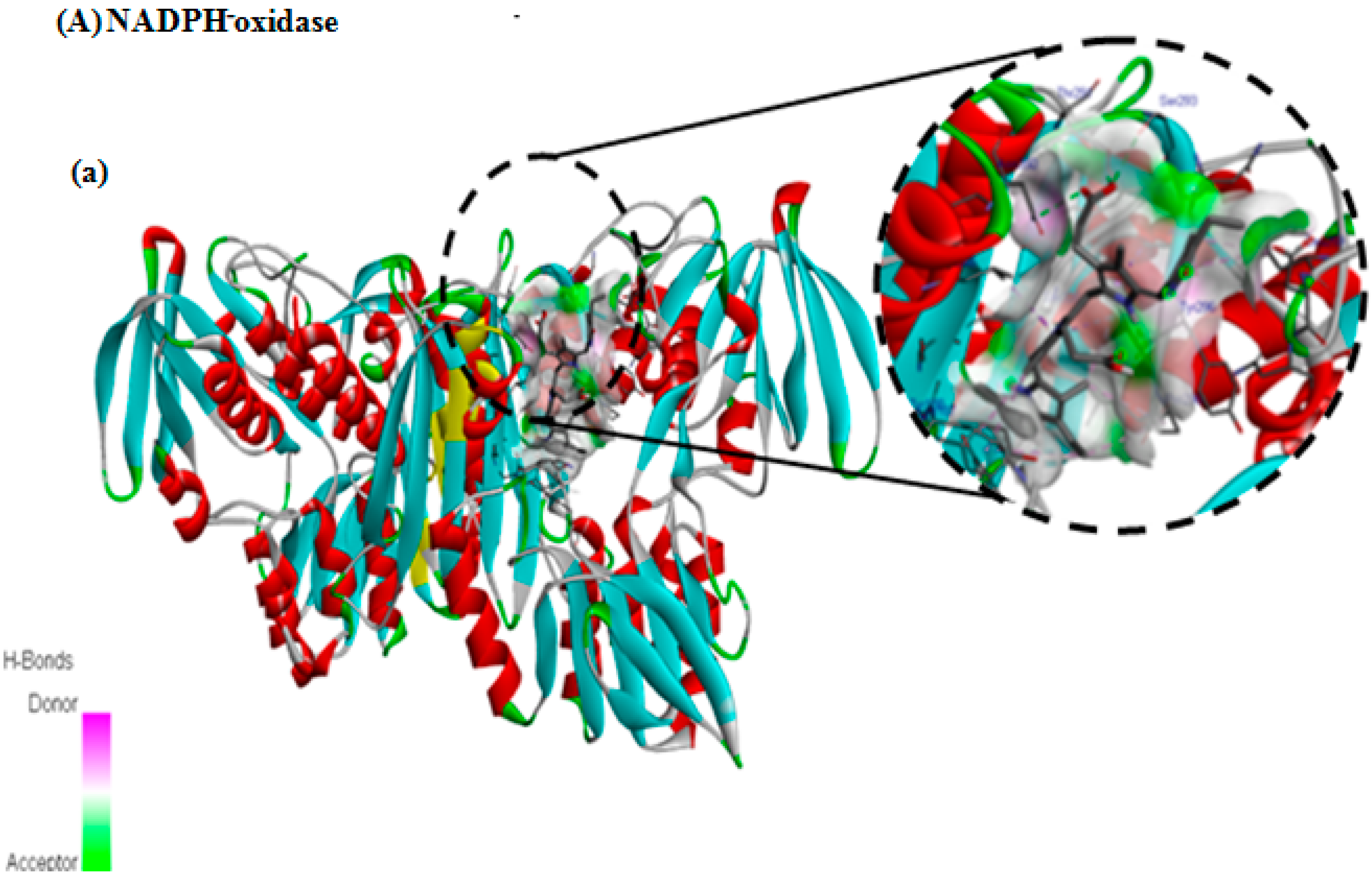
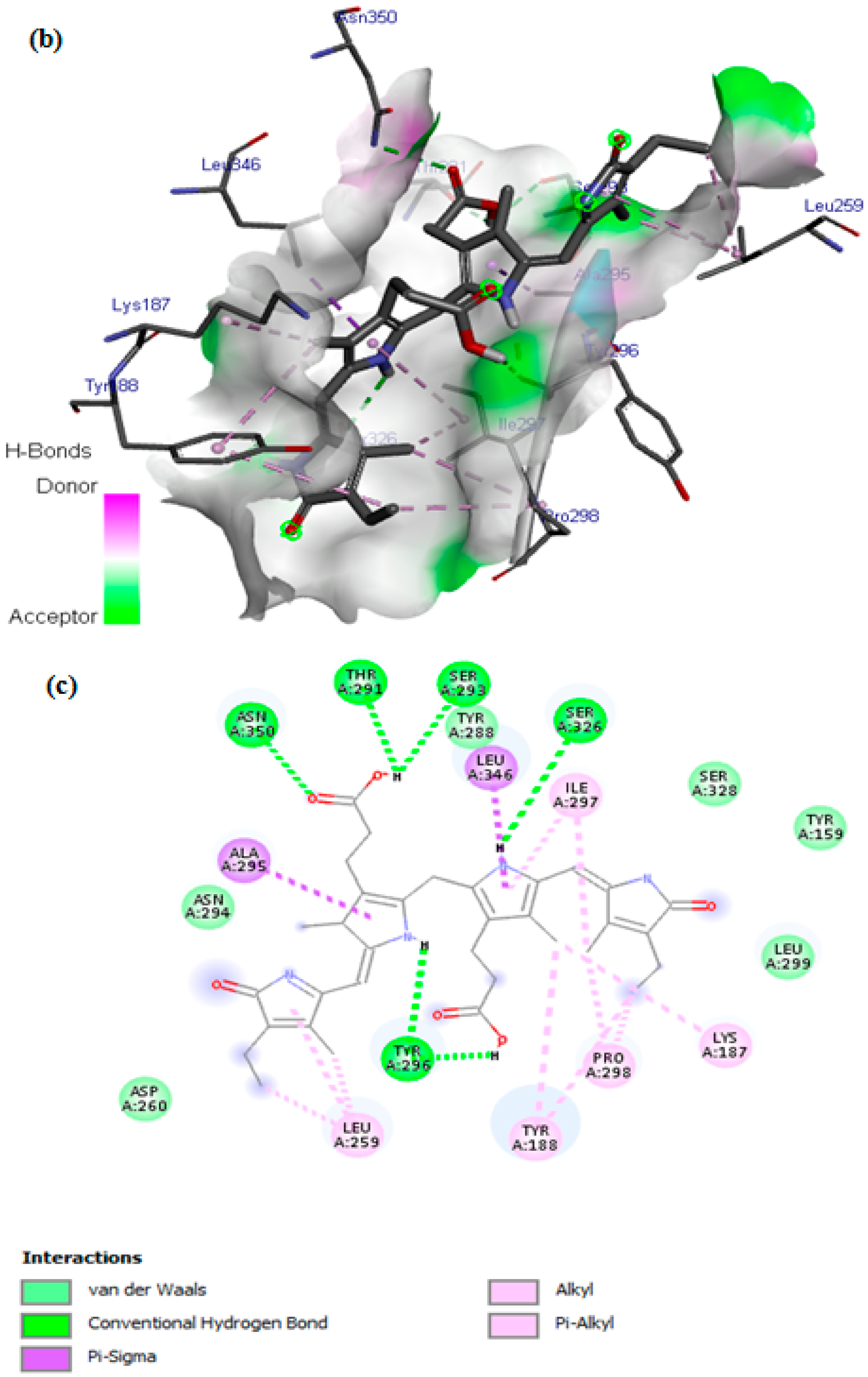
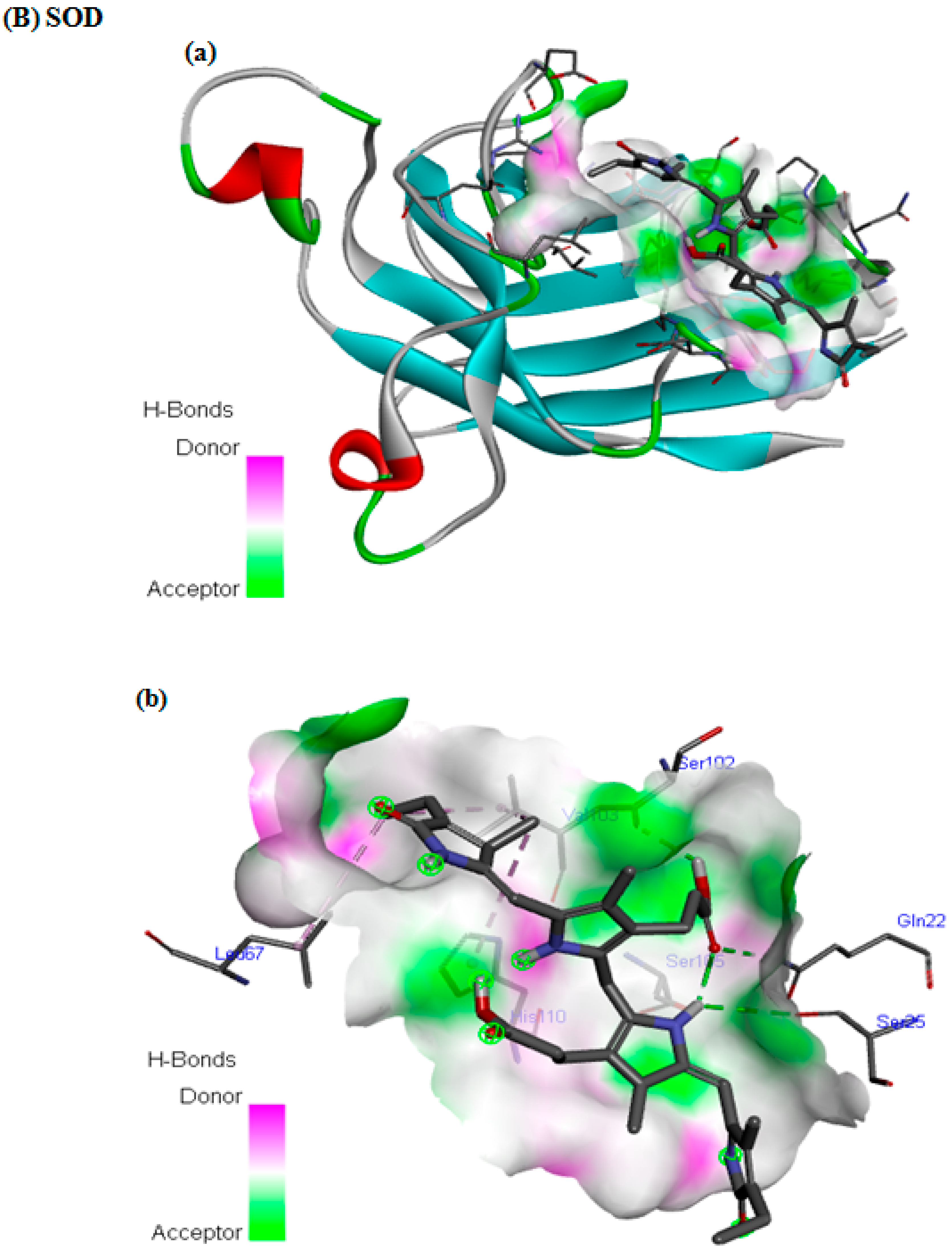
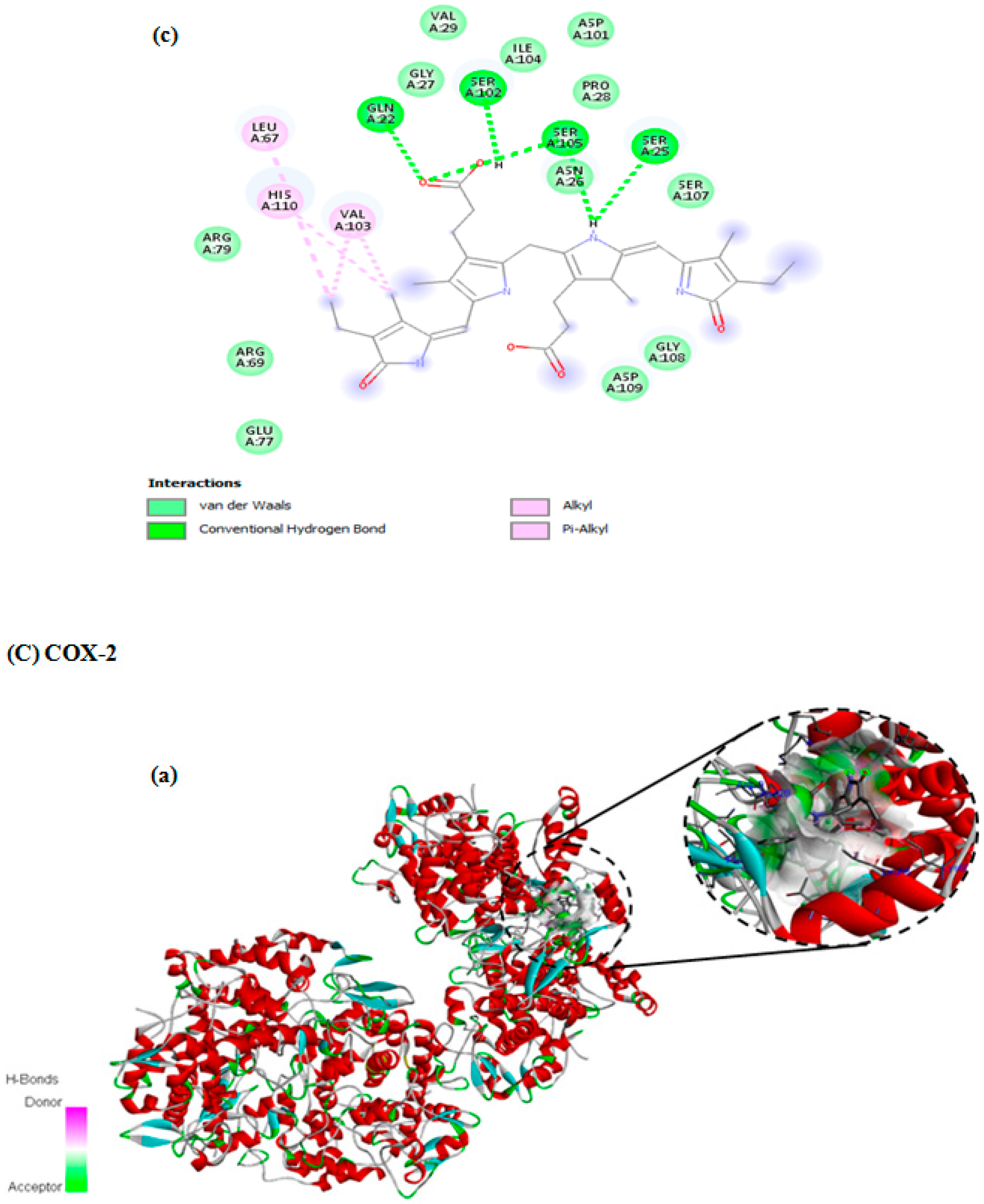
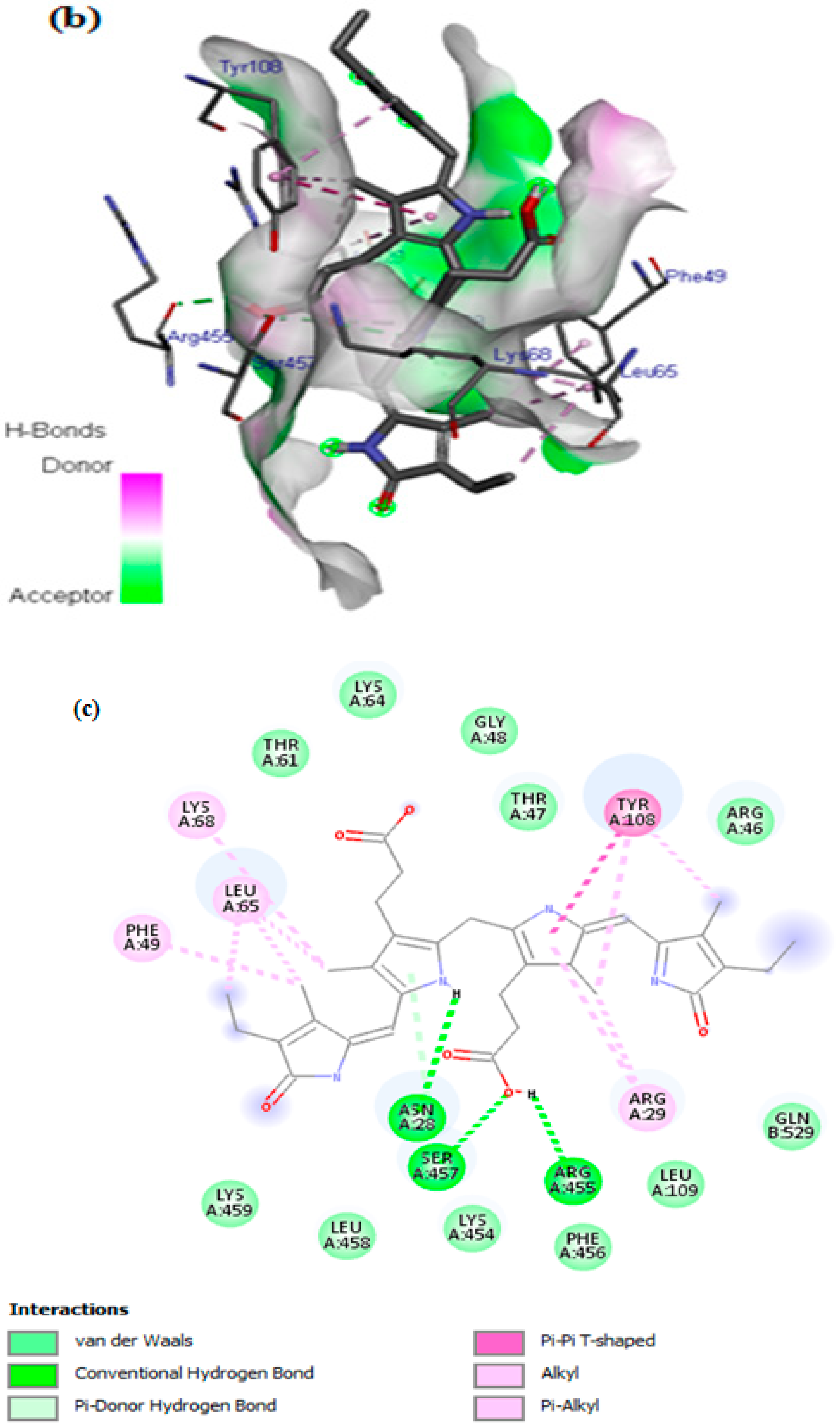
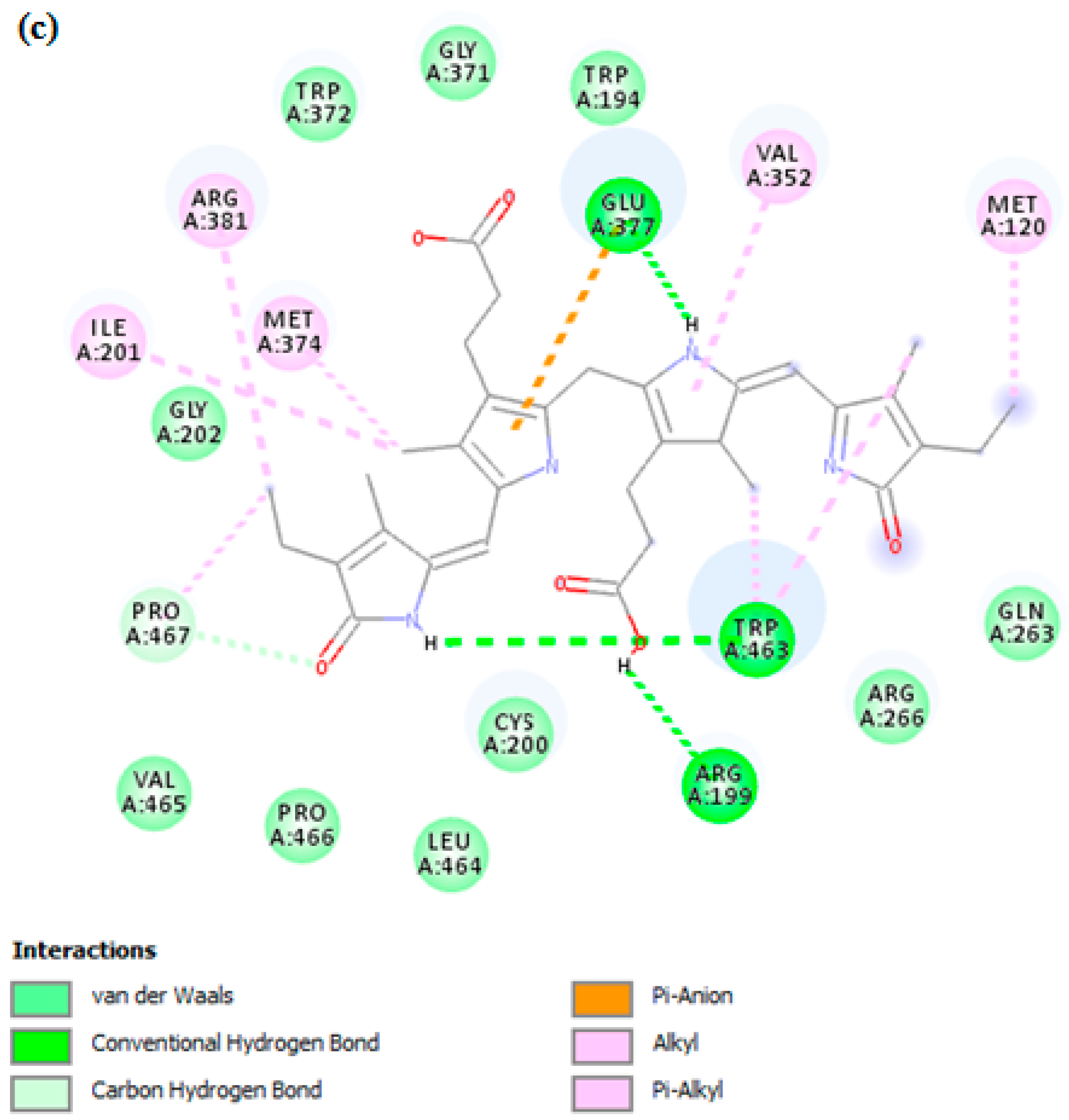
3.7. In Silico Docking Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- LeMieux, M.J.; Kalupahana, N.S.; Scoggin, S.; Moustaid-Moussa, N. Eicosapentaenoic Acid Reduces Adipocyte Hypertrophy and Inflammation in Diet-Induced Obese Mice in an Adiposity-Independent Manner. J. Nutr. 2015, 145, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B. Obesity and Mortality: Watch Your Waist, Not Just Your Weight. Arch. Intern. Med. 2007, 167, 875–876. [Google Scholar] [CrossRef]
- Moreno, L.A.; Rodriguez, G.; Fleta, J.; Bueno-Lozano, M.; Lazaro, A.; Bueno, G. Trends of Dietary Habits in Adolescents. Crit. Rev. Food Sci. Nutr. 2010, 50, 106–112. [Google Scholar] [CrossRef]
- Sommer, F.; Bäckhed, F. Know Your Neighbor: Microbiota and Host Epithelial Cells Interact Locally to Control Intestinal Function and Physiology. BioEssays News Rev. Mol. Cell. Dev. Biol. 2016, 38, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in Gut Microbiota Control Metabolic Endotoxemia-Induced Inflammation in High-Fat Diet-Induced Obesity and Diabetes in Mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Morales, P.; Fujio, S.; Navarrete, P.; Ugalde, J.A.; Magne, F.; Carrasco-Pozo, C.; Tralma, K.; Quezada, M.; Hurtado, C.; Covarrubias, N.; et al. Impact of Dietary Lipids on Colonic Function and Microbiota: An Experimental Approach Involving Orlistat-Induced Fat Malabsorption in Human Volunteers. Clin. Transl. Gastroenterol. 2016, 7, e161. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Liu, S.; Hui, Q.; Zhang, H.; Lahaye, L.; Bodin, J.-C.; Gong, J.; Nyachoti, M.; Yang, C. Thymol Improves Barrier Function and Attenuates Inflammatory Responses in Porcine Intestinal Epithelial Cells during Lipopolysaccharide (LPS)-Induced Inflammation. J. Agric. Food Chem. 2019, 67, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Siliciano, J.D.; Goodenough, D.A. Localization of the Tight Junction Protein, ZO-1, Is Modulated by Extracellular Calcium and Cell-Cell Contact in Madin-Darby Canine Kidney Epithelial Cells. J. Cell Biol. 1988, 107, 2389–2399. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liu, B.; Wang, X.; Yu, Q.; Fang, R. Epidermal Growth Factor, through Alleviating Oxidative Stress, Protect IPEC-J2 Cells from Lipopolysaccharides-Induced Apoptosis. Int. J. Mol. Sci. 2018, 19, 848. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic Endotoxemia Initiates Obesity and Insulin Resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Powell, D.N.; Kalman, D. Layered Defense: How Mucus and Tight Junctions Seal the Intestinal Barrier. J. Mol. Med. Berl. Ger. 2017, 95, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Bonnefont-Rousselot, D. Obésité et Stress Oxydant. Obésité 2014, 1, 8–13. [Google Scholar] [CrossRef]
- Rao, R. Oxidative Stress-Induced Disruption of Epithelial and Endothelial Tight Junctions. Front. Biosci. J. Virtual Libr. 2008, 13, 7210–7226. [Google Scholar] [CrossRef] [PubMed]
- Marciano, F.; Vajro, P. Oxidative Stress and Gut Microbiota. In Gastrointestinal Tissue; Elsevier: Amsterdam, The Netherlands, 2017; pp. 113–123. [Google Scholar]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ Effector T Cells Contribute to Macrophage Recruitment and Adipose Tissue Inflammation in Obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Perrard, X.D.; Wang, Q.; Perrard, J.L.; Polsani, V.R.; Jones, P.H.; Smith, C.W.; Ballantyne, C.M. CD11c Expression in Adipose Tissue and Blood and Its Role in Diet-Induced Obesity. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 186–192. [Google Scholar] [CrossRef]
- Akkari, I.; Hassine, A.; Soumaya, M.; Harbi, R.; Jazia, E.B. Surpoids et Maladie de Crohn: Cause Ou Conséquence? Rev. Médecine Interne 2022, 43, A204–A205. [Google Scholar] [CrossRef]
- Shen, L. Tight Junctions on the Move: Molecular Mechanisms for Epithelial Barrier Regulation. Ann. N. Y. Acad. Sci. 2012, 1258, 9–18. [Google Scholar] [CrossRef]
- Ivashkin, V.; Poluektov, Y.; Kogan, E.; Shifrin, O.; Sheptulin, A.; Kovaleva, A.; Kurbatova, A.; Krasnov, G.; Poluektova, E. Disruption of the Pro-Inflammatory, Anti-Inflammatory Cytokines and Tight Junction Proteins Expression, Associated with Changes of the Composition of the Gut Microbiota in Patients with Irritable Bowel Syndrome. PLoS ONE 2021, 16, e0252930. [Google Scholar] [CrossRef]
- Kong, J.-M.; Goh, N.-K.; Chia, L.-S.; Chia, T.-F. Recent Advances in Traditional Plant Drugs and Orchids. Acta Pharmacol. Sin. 2003, 24, 7–21. [Google Scholar]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The Promising Future of Microalgae: Current Status, Challenges, and Optimization of a Sustainable and Renewable Industry for Biofuels, Feed, and Other Products. Microb. Cell Factories 2018, 17, 36. [Google Scholar] [CrossRef]
- Yousefi, R.; Mottaghi, A.; Saidpour, A. Spirulina Platensis Effectively Ameliorates Anthropometric Measurements and Obesity-Related Metabolic Disorders in Obese or Overweight Healthy Individuals: A Randomized Controlled Trial. Complement. Ther. Med. 2018, 40, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Kulshreshtha, A.; Zacharia, A.J.; Jarouliya, U.; Bhadauriya, P.; Prasad, G.B.K.S.; Bisen, P.S. Spirulina in Health Care Management. Curr. Pharm. Biotechnol. 2008, 9, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Bortolini, D.G.; Maciel, G.M.; Fernandes, I.d.A.A.; Pedro, A.C.; Rubio, F.T.V.; Branco, I.G.; Haminiuk, C.W.I. Functional Properties of Bioactive Compounds from Spirulina Spp.: Current Status and Future Trends. Food Chem. Mol. Sci. 2022, 5, 100134. [Google Scholar] [CrossRef]
- Zheng, J.; Inoguchi, T.; Sasaki, S.; Maeda, Y.; McCarty, M.F.; Fujii, M.; Ikeda, N.; Kobayashi, K.; Sonoda, N.; Takayanagi, R. Phycocyanin and Phycocyanobilin from Spirulina Platensis Protect against Diabetic Nephropathy by Inhibiting Oxidative Stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R110–R120. [Google Scholar] [CrossRef] [PubMed]
- Citi, V.; Torre, S.; Flori, L.; Usai, L.; Aktay, N.; Dunford, N.T.; Lutzu, G.A.; Nieri, P. Nutraceutical Features of the Phycobiliprotein C-Phycocyanin: Evidence from Arthrospira Platensis (Spirulina). Nutrients 2024, 16, 1752. [Google Scholar] [CrossRef]
- Arrari, F.; Jabri, M.-A.; Ayari, A.; Dakhli, N.; Ben Fayala, C.; Boubaker, S.; Sebai, H. Chromatographic Analyses of Spirulina (Arthrospira platensis) and Mechanism of Its Protective Effects against Experimental Obesity and Hepatic Steatosis in Rats. Med. Kaunas Lith. 2023, 59, 1823. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The Antioxidant, Immunomodulatory, and Anti-Inflammatory Activities of Spirulina: An Overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef]
- Arrari, F.; Jabri, M.-A.; Ayari, A.; Dakhli, N.; Ben Fayala, C.; Boubaker, S.; Sebai, H. Amino Acid HPLC-FLD Analysis of Spirulina and Its Protective Mechanism against the Combination of Obesity and Colitis in Wistar Rats. Heliyon 2024, 10, e30103. [Google Scholar] [CrossRef]
- Karczewski, J.; Śledzińska, E.; Baturo, A.; Jończyk, I.; Maleszko, A.; Samborski, P.; Begier-Krasińska, B.; Dobrowolska, A. Obesity and Inflammation. Eur. Cytokine Netw. 2018, 29, 83–94. [Google Scholar] [CrossRef]
- Finamore, A.; Palmery, M.; Bensehaila, S.; Peluso, I. Antioxidant, Immunomodulating, and Microbial-Modulating Activities of the Sustainable and Ecofriendly Spirulina. Oxid. Med. Cell. Longev. 2017, 2017, 3247528. [Google Scholar] [CrossRef]
- Ghaem Far, Z.; Babajafari, S.; Kojuri, J.; Mohammadi, S.; Nouri, M.; Rostamizadeh, P.; Rahmani, M.H.; Azadian, M.; Ashrafi-Dehkordi, E.; Zareifard, A.; et al. Antihypertensive and Antihyperlipemic Effects of Spirulina (Arthrospira platensis) Sauce on Patients with Hypertension: A Randomized Triple-Blind Placebo-Controlled Clinical Trial. Phytother. Res. 2021, 35, 6181–6190. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I.; Bursal, E.; Sehitoğlu, M.H.; Bilsel, M.; Gören, A.C. Polyphenol Contents and Antioxidant Activity of Lyophilized Aqueous Extract of Propolis from Erzurum, Turkey. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2010, 48, 2227–2238. [Google Scholar] [CrossRef] [PubMed]
- Chantret, I.; Rodolosse, A.; Barbat, A.; Dussaulx, E.; Brot-Laroche, E.; Zweibaum, A.; Rousset, M. Differential Expression of Sucrase-Isomaltase in Clones Isolated from Early and Late Passages of the Cell Line Caco-2: Evidence for Glucose-Dependent Negative Regulation. J. Cell Sci. 1994, 107 Pt 1, 213–225. [Google Scholar] [CrossRef]
- Sowmya, S.; Suba Sri, M.; Dineshkumar, R. In Vitro Therapeutic Effect of Spirulina Extract. Asian J. Biol. Life Sci. 2022, 10, 582–589. [Google Scholar] [CrossRef]
- Béaslas, O.; Cueille, C.; Delers, F.; Chateau, D.; Chambaz, J.; Rousset, M.; Carrière, V. Sensing of Dietary Lipids by Enterocytes: A New Role for SR-BI/CLA-1. PLoS ONE 2009, 4, e4278. [Google Scholar] [CrossRef]
- Chateau, D.; Pauquai, T.; Delers, F.; Rousset, M.; Chambaz, J.; Demignot, S. Lipid Micelles Stimulate the Secretion of Triglyceride-Enriched Apolipoprotein B48-Containing Lipoproteins by Caco-2 Cells. J. Cell. Physiol. 2005, 202, 767–776. [Google Scholar] [CrossRef]
- Tran, T.T.T.; Postal, B.G.; Demignot, S.; Ribeiro, A.; Osinski, C.; Pais de Barros, J.-P.; Blachnio-Zabielska, A.; Leturque, A.; Rousset, M.; Ferré, P.; et al. Short Term Palmitate Supply Impairs Intestinal Insulin Signaling via Ceramide Production. J. Biol. Chem. 2016, 291, 16328–16338. [Google Scholar] [CrossRef]
- Kowapradit, J.; Opanasopit, P.; Ngawhiranpat, T.; Apirakaramwong, A.; Rojanarata, T.; Ruktanonchai, U.; Sajomsang, W. Methylated N-(4-N,N-Dimethylaminobenzyl) Chitosan, a Novel Chitosan Derivative, Enhances Paracellular Permeability across Intestinal Epithelial Cells (Caco-2). AAPS PharmSciTech 2008, 9, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Błaszczak-Świątkiewicz, K.; Mikiciuk-Olasik, E. Some Characteristics of Activity of Potential Chemotherapeutics–Benzimidazole Derivatives. Adv. Med. Sci. 2015, 60, 125–132. [Google Scholar] [CrossRef]
- Hartree, E.F. Determination of Protein: A Modification of the Lowry Method That Gives a Linear Photometric Response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde Determination as Index of Lipid Peroxidation. Methods Enzymol. 1990, 186, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, J.; Lindsay, R.H. Estimation of Total, Protein-Bound, and Nonprotein Sulfhydryl Groups in Tissue with Ellman’s Reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Misra, H.P.; Fridovich, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef] [PubMed]
- Flohé, L.; Günzler, W.A. Assays of Glutathione Peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Dingeon, B.; Ferry, J.P.; Roullet, A. Automatic Assay of Blood Sugar by Trinder’s Method. Ann. Biol. Clin. 1975, 33, 3–13. [Google Scholar]
- Payá, M.; Halliwell, B.; Hoult, J.R. Interactions of a Series of Coumarins with Reactive Oxygen Species. Scavenging of Superoxide, Hypochlorous Acid and Hydroxyl Radicals. Biochem. Pharmacol. 1992, 44, 205–214. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the Superoxide Anion Radical in the Autoxidation of Pyrogallol and a Convenient Assay for Superoxide Dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Hirata, T.; Tanaka, M.; Ooike, M.; Tsunomura, T.; Sakaguchi, M. Antioxidant Activities of Phycocyanobilin Prepared from Spirulina Platensis. J. Appl. Phycol. 2000, 12, 435–439. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Visualizer, D.S. Biovia; Dassault Systèmes: San Diego, CA, USA, 2020. [Google Scholar]
- Francescangeli, F.; De Angelis, M.L.; Zeuner, A. Dietary Factors in the Control of Gut Homeostasis, Intestinal Stem Cells, and Colorectal Cancer. Nutrients 2019, 11, 2936. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.P.B.; Texeira, T.F.S.; Ferreira, A.B.; Peluzio, M.d.C.G.; Alfenas, R.d.C.G. Influence of a High-Fat Diet on Gut Microbiota, Intestinal Permeability and Metabolic Endotoxaemia. Br. J. Nutr. 2012, 108, 801–809. [Google Scholar] [CrossRef]
- Ghezzal, S.; Postal, B.G.; Quevrain, E.; Brot, L.; Seksik, P.; Leturque, A.; Thenet, S.; Carrière, V. Palmitic Acid Damages Gut Epithelium Integrity and Initiates Inflammatory Cytokine Production. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158530. [Google Scholar] [CrossRef]
- Chen, J.-C.; Liu, K.S.; Yang, T.-J.; Hwang, J.-H.; Chan, Y.-C.; Lee, I.-T. Spirulina and C-Phycocyanin Reduce Cytotoxicity and Inflammation-Related Genes Expression of Microglial Cells. Nutr. Neurosci. 2012, 15, 252–256. [Google Scholar] [CrossRef]
- Yu, T.; Wang, Y.; Chen, X.; Xiong, W.; Tang, Y.; Lin, L. Spirulina Platensis Alleviates Chronic Inflammation with Modulation of Gut Microbiota and Intestinal Permeability in Rats Fed a High-Fat Diet. J. Cell. Mol. Med. 2020, 24, 8603–8613. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Deng, J.; Hu, X.; Zhou, S.; Wu, J.; Xiao, D.; Darko, K.O.; Huang, Y.; Tao, T.; Peng, M.; et al. Vitamin A Inhibits the Action of LPS on the Intestinal Epithelial Barrier Function and Tight Junction Proteins. Food Funct. 2019, 10, 1235–1242. [Google Scholar] [CrossRef]
- Benham, A.M. Protein Secretion and the Endoplasmic Reticulum. Cold Spring Harb. Perspect. Biol. 2012, 4, a012872. [Google Scholar] [CrossRef]
- Deguil, J.; Pineau, L.; Rowland Snyder, E.C.; Dupont, S.; Beney, L.; Gil, A.; Frapper, G.; Ferreira, T. Modulation of Lipid-Induced ER Stress by Fatty Acid Shape. Traffic Cph. Den. 2011, 12, 349–362. [Google Scholar] [CrossRef]
- Pardo, V.; González-Rodríguez, Á.; Muntané, J.; Kozma, S.C.; Valverde, Á.M. Role of Hepatocyte S6K1 in Palmitic Acid-Induced Endoplasmic Reticulum Stress, Lipotoxicity, Insulin Resistance and in Oleic Acid-Induced Protection. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2015, 80, 298–309. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, Z.; Zhu, L.; Ma, S.; Luo, Y.; Liang, H.; Liu, Q.; Chen, J.; Guli, S.; Chen, X. Orchestration of MUC2—The Key Regulatory Target of Gut Barrier and Homeostasis: A Review. Int. J. Biol. Macromol. 2023, 236, 123862. [Google Scholar] [CrossRef] [PubMed]
- Camuesco, D.; Comalada, M.; Rodríguez-Cabezas, M.E.; Nieto, A.; Lorente, M.D.; Concha, A.; Zarzuelo, A.; Gálvez, J. The Intestinal Anti-Inflammatory Effect of Quercitrin Is Associated with an Inhibition in iNOS Expression. Br. J. Pharmacol. 2004, 143, 908–918. [Google Scholar] [CrossRef] [PubMed]
- Layam, A.; Reddy, C.L.K. Antidiabetic Property of Spirulina. Diabetol. Croat. 2006, 35, 29–33. [Google Scholar]
- Wu, L.; Ho, J.A.; Shieh, M.-C.; Lu, I.-W. Antioxidant and Antiproliferative Activities of Spirulina and Chlorella Water Extracts. J. Agric. Food Chem. 2005, 53, 4207–4212. [Google Scholar] [CrossRef]
- González de Rivera, C.; Miranda-Zamora, R.; Díaz-Zagoya, J.C.; Juárez-Oropeza, M.A. Preventive Effect of Spirulina Maxima on the Fatty Liver Induced by a Fructose-Rich Diet in the Rat, a Preliminary Report. Life Sci. 1993, 53, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Jin, Y.; Hofseth, A.B.; Pena, E.; Habiger, J.; Chumanevich, A.; Poudyal, D.; Nagarkatti, M.; Nagarkatti, P.S.; Singh, U.P.; et al. Resveratrol Suppresses Colitis and Colon Cancer Associated with Colitis. Cancer Prev. Res. 2010, 3, 549–559. [Google Scholar] [CrossRef]
- Piovan, A.; Filippini, R.; Argentini, C.; Moro, S.; Giusti, P.; Zusso, M. The Effect of C-Phycocyanin on Microglia Activation Is Mediated by Toll-like Receptor 4. Int. J. Mol. Sci. 2022, 23, 1440. [Google Scholar] [CrossRef] [PubMed]
- Khafaga, A.F.; El-Sayed, Y.S. Spirulina Ameliorates Methotrexate Hepatotoxicity via Antioxidant, Immune Stimulation, and Proinflammatory Cytokines and Apoptotic Proteins Modulation. Life Sci. 2018, 196, 9–17. [Google Scholar] [CrossRef]
- Lee, J.; Park, A.; Kim, M.J.; Lim, H.-J.; Rha, Y.-A.; Kang, H.-G. Spirulina Extract Enhanced a Protective Effect in Type 1 Diabetes by Anti-Apoptosis and Anti-ROS Production. Nutrients 2017, 9, 1363. [Google Scholar] [CrossRef]
- Ngu, E.-L.; Tan, C.-Y.; Lai, N.J.-Y.; Wong, K.-H.; Lim, S.-H.; Ming, L.C.; Tan, K.-O.; Phang, S.-M.; Yow, Y.-Y. Spirulina Platensis Suppressed iNOS and Proinflammatory Cytokines in Lipopolysaccharide-Induced BV2 Microglia. Metabolites 2022, 12, 1147. [Google Scholar] [CrossRef]
- Omar, A.E.; Al-Khalaifah, H.S.; Osman, A.; Gouda, A.; Shalaby, S.I.; Roushdy, E.M.; Abdo, S.A.; Ali, S.A.; Hassan, A.M.; Amer, S.A. Modulating the Growth, Antioxidant Activity, and Immunoexpression of Proinflammatory Cytokines and Apoptotic Proteins in Broiler Chickens by Adding Dietary Spirulina Platensis Phycocyanin. Antioxidants 2022, 11, 991. [Google Scholar] [CrossRef] [PubMed]
- Tounsi, L.; Ben Hlima, H.; Hentati, F.; Hentati, O.; Derbel, H.; Michaud, P.; Abdelkafi, S. Microalgae: A Promising Source of Bioactive Phycobiliproteins. Mar. Drugs 2023, 21, 440. [Google Scholar] [CrossRef]
- Li, Y. The Bioactivities of Phycocyanobilin from Spirulina. J. Immunol. Res. 2022, 2022, 4008991. [Google Scholar] [CrossRef]
- McCarty, M.F. Clinical Potential of Spirulina as a Source of Phycocyanobilin. J. Med. Food 2007, 10, 566–570. [Google Scholar] [CrossRef]
- Müller, K.; Engesser, R.; Timmer, J.; Nagy, F.; Zurbriggen, M.D.; Weber, W. Synthesis of Phycocyanobilin in Mammalian Cells. Chem. Commun. 2013, 49, 8970–8972. [Google Scholar] [CrossRef]
- Marín-Prida, J.; Pavón-Fuentes, N.; Llópiz-Arzuaga, A.; Fernández-Massó, J.R.; Delgado-Roche, L.; Mendoza-Marí, Y.; Santana, S.P.; Cruz-Ramírez, A.; Valenzuela-Silva, C.; Nazábal-Gálvez, M.; et al. Phycocyanobilin Promotes PC12 Cell Survival and Modulates Immune and Inflammatory Genes and Oxidative Stress Markers in Acute Cerebral Hypoperfusion in Rats. Toxicol. Appl. Pharmacol. 2013, 272, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, Q.-Y.; Yu, L.-M.; Liu, B.; Li, M.-Y.; Zhu, R.-Z. Phycocyanobilin Accelerates Liver Regeneration and Reduces Mortality Rate in Carbon Tetrachloride-Induced Liver Injury Mice. World J. Gastroenterol. WJG 2015, 21, 5465. [Google Scholar] [CrossRef]
- McCarty, M.F. Supplementation with Phycocyanobilin, Citrulline, Taurine, and Supranutritional Doses of Folic Acid and Biotin—Potential for Preventing or Slowing the Progression of Diabetic Complications. Healthcare 2017, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Bedard, K.; Krause, K.-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.-F.; Teixeira, M.; Valentine, J.S. Superoxide Dismutases and Superoxide Reductases. Chem. Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef]
- Pentón-Rol, G.; Marín-Prida, J.; McCarty, M.F. C-Phycocyanin-Derived Phycocyanobilin as a Potential Nutraceutical Approach for Major Neurodegenerative Disorders and COVID-19-Induced Damage to the Nervous System. Curr. Neuropharmacol. 2021, 19, 2250. [Google Scholar] [CrossRef] [PubMed]
- Farouk, A.; Mohsen, M.; Ali, H.; Shaaban, H.; Albaridi, N. Antioxidant Activity and Molecular Docking Study of Volatile Constituents from Different Aromatic Lamiaceous Plants Cultivated in Madinah Monawara, Saudi Arabia. Molecules 2021, 26, 4145. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Calderon, O.; Chacaltana-Ramos, L.J.; Huayanca-Gutiérrez, I.C.; Algarni, M.A.; Alqarni, M.; Batiha, G.E.-S. Chemical Constituents, In Vitro Antioxidant Activity and In Silico Study on NADPH Oxidase of Allium sativum L. (Garlic) Essential Oil. Antioxidants 2021, 10, 1844. [Google Scholar] [CrossRef]
- Khan, M.S.; Khan, M.K.A.; Siddiqui, M.H.; Arif, J.M. An in Vivo and in Silico Approach to Elucidate the Tocotrienol-Mediated Fortification against Infection and Inflammation Induced Alterations in Antioxidant Defense System. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 916–930. [Google Scholar] [PubMed]
- Hejazi, I.I.; Khanam, R.; Mehdi, S.H.; Bhat, A.R.; Moshahid Alam Rizvi, M.; Islam, A.; Thakur, S.C.; Athar, F. New Insights into the Antioxidant and Apoptotic Potential of Glycyrrhiza glabra L. during Hydrogen Peroxide Mediated Oxidative Stress: An in Vitro and in Silico Evaluation. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 94, 265–279. [Google Scholar] [CrossRef]
- da Silva Pantoja, L.V.P.; Trindade, S.S.A.; da Silva Carneiro, A.; Silva, J.P.B.; da Paixão, T.P.; Romeiro, C.F.R.; de Moraes, C.S.P.; Pinto, A.C.G.; Raposo, N.R.B.; de Andrade, M.A. Computational Study of the Main Flavonoids from Chrysobalanus icaco L. against NADPH-Oxidase and in Vitro Antioxidant Activity. Res. Soc. Dev. 2022, 11, e5011628542. [Google Scholar] [CrossRef]
- Bolli, R.; Shinmura, K.; Tang, X.-L.; Kodani, E.; Xuan, Y.-T.; Guo, Y.; Dawn, B. Discovery of a New Function of Cyclooxygenase (COX)-2: COX-2 Is a Cardioprotective Protein That Alleviates Ischemia/Reperfusion Injury and Mediates the Late Phase of Preconditioning. Cardiovasc. Res. 2002, 55, 506–519. [Google Scholar] [CrossRef]
- Shreshtha, S.; Sharma, P.; Kumar, P.; Sharma, R.; Singh, S.P. Nitric Oxide: It’s Role in Immunity. J. Clin. Diagn. Res. 2018, 12, BE01–BE05. [Google Scholar] [CrossRef]
- Guo, W.; Zeng, M.; Zhu, S.; Li, S.; Qian, Y.; Wu, H. Phycocyanin Ameliorates Mouse Colitis via Phycocyanobilin-Dependent Antioxidant and Anti-Inflammatory Protection of the Intestinal Epithelial Barrier. Food Funct. 2022, 13, 3294–3307. [Google Scholar] [CrossRef]
- Prasanth, S.; Kumar Arun, G.; Haridas, M.; Sabu, A. Phycocyanin of Marine Oscillatoria Sp. Inhibits Lipoxygenase by Protein-Protein Interaction-Induced Change of Active Site Entry Apace: A Model for Non-Specific Biofunctions of Phycocyanins. Int. J. Biol. Macromol. 2020, 165, 1111–1118. [Google Scholar] [CrossRef]
- Espinoza-Culupú, A.; Del Santos, N.; Farfán-López, M.; Mendes, E.; da Silva Junior, P.I.; Marques Borges, M. In Silico and In Vitro Approach for Evaluation of the Anti-Inflammatory and Antioxidant Potential of Mygalin. Int. J. Mol. Sci. 2023, 24, 17019. [Google Scholar] [CrossRef] [PubMed]
- Ben Attia, T.; Nahdi, A.; Horchani, M.; Elmay, M.V.; Ksentini, M.; Ben Jannet, H.; Mhamdi, A. Olea europaea L. Leaf Extract Mitigates Pulmonary Inflammation and Tissue Destruction in Wistar Rats Induced by Concurrent Exposure to Noise and Toluene. Drug Chem. Toxicol. 2024, 47, 1072–1086. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) |
|---|---|
| XBP1Fow XBP1 Rev | TGGCCGGGTCTGCTGAGTCCG ATCCATGGGGAGATGTTCTGG |
| CHOP Fow CHOP Rev | AGAACCAGGAAACGGAAACAGA TCTCCTTCATGCGCTGCTTT |
| ZO-1 Fow ZO-1 Rev | GACCTTGAGCAGCCGTCATA CCGTAGGCGATGGTCATAGTT |
| OCL-1 Fow OCL-1 Rev | CTTTGGCTACGGAGGTGGCTAT CTTTGGCTGCTCTTGGGTCTG |
| iNOSFow iNOSRev | TGCAGACACGTGCGTACTC GGTAGCCAGCATAGCGGATG |
| GAPDH Fow GAPDH Rev | CATGGCCTTCCGTGTTCCTA CCTGCTTCACCACCTTCTTGAT |
| Intermolecular Interactions | |||
|---|---|---|---|
| Protein | Docking Energy (kcal/mol) | Conventional Hydrogen Bonds | Interacting Amino Acid Residues |
| NADPH oxidase | −8 | 6 | SER293, THR291, ASN350, TYR288, LEU346, SER326, ILE297, LYS187, PRO298, TYR188, TYR296, LEU259, ALA295 |
| COX-2 | −8.3 | 3 | LYS68, LEU65, PHE49, ASN28, SER454, ARG455, TYR108, ARG28 |
| iNOS | −9.1 | 3 | GLU377, MET374, ARG381, ILE201, PRO465, ARG199, TRP463, VAL352, MET120 |
| SOD | −6.4 | 4 | VAL29, GLY27, GLN22, LEU67, HIS110, VAL103, ARG69, GLU77, ASP109, GLY108, SER105, ASN26, SER107, PRO28, SER105, ASP101, ILE104, SER102 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arrari, F.; Ortiz-Flores, R.-M.; Lhamyani, S.; Garcia-Fuentes, E.; Jabri, M.-A.; Sebai, H.; Bermudez-Silva, F.-J. Protective Effects of Spirulina Against Lipid Micelles and Lipopolysaccharide-Induced Intestinal Epithelium Disruption in Caco-2 Cells: In Silico Molecular Docking Analysis of Phycocyanobilin. Nutrients 2024, 16, 4074. https://doi.org/10.3390/nu16234074
Arrari F, Ortiz-Flores R-M, Lhamyani S, Garcia-Fuentes E, Jabri M-A, Sebai H, Bermudez-Silva F-J. Protective Effects of Spirulina Against Lipid Micelles and Lipopolysaccharide-Induced Intestinal Epithelium Disruption in Caco-2 Cells: In Silico Molecular Docking Analysis of Phycocyanobilin. Nutrients. 2024; 16(23):4074. https://doi.org/10.3390/nu16234074
Chicago/Turabian StyleArrari, Fatma, Rodolfo-Matias Ortiz-Flores, Said Lhamyani, Eduardo Garcia-Fuentes, Mohamed-Amine Jabri, Hichem Sebai, and Francisco-Javier Bermudez-Silva. 2024. "Protective Effects of Spirulina Against Lipid Micelles and Lipopolysaccharide-Induced Intestinal Epithelium Disruption in Caco-2 Cells: In Silico Molecular Docking Analysis of Phycocyanobilin" Nutrients 16, no. 23: 4074. https://doi.org/10.3390/nu16234074
APA StyleArrari, F., Ortiz-Flores, R.-M., Lhamyani, S., Garcia-Fuentes, E., Jabri, M.-A., Sebai, H., & Bermudez-Silva, F.-J. (2024). Protective Effects of Spirulina Against Lipid Micelles and Lipopolysaccharide-Induced Intestinal Epithelium Disruption in Caco-2 Cells: In Silico Molecular Docking Analysis of Phycocyanobilin. Nutrients, 16(23), 4074. https://doi.org/10.3390/nu16234074