
Associations of Microbiota and Nutrition with Cognitive Impairment in Diseases

 , , , , , ,
, , , , , , 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Cognitive Dysfunctions and Their Underlying Mechanisms in Various Diseases
2.1. Cognitive Function and Aging
2.2. Accelerated Cognitive Decline in Pathological Aging
2.3. Cognitive Function in Neurodegenerative Disorders
2.4. Cognitive Function in Psychiatric Disorders
2.5. Cognitive Function in Dietary Diseases
2.6. Cognitive Function in Obesity
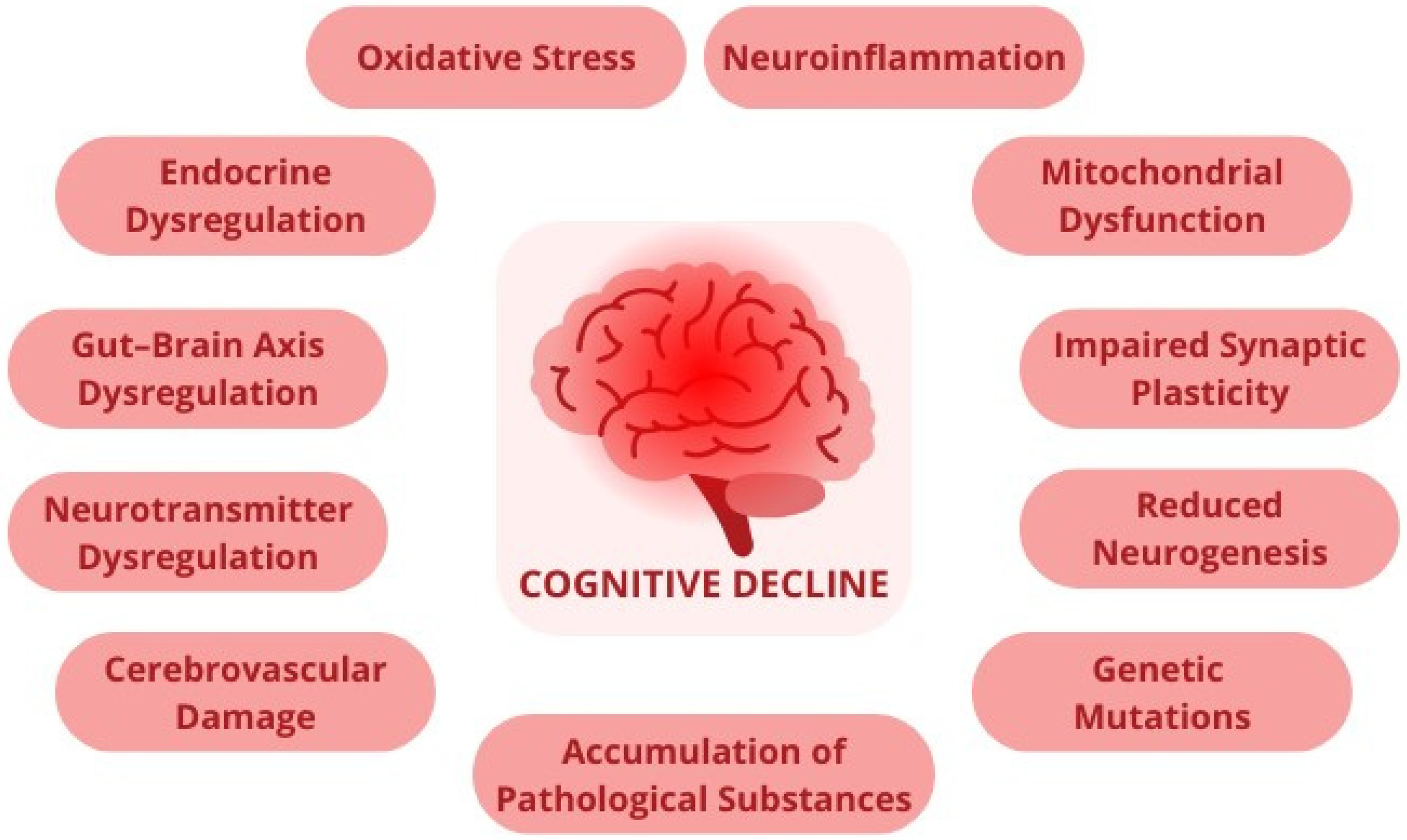
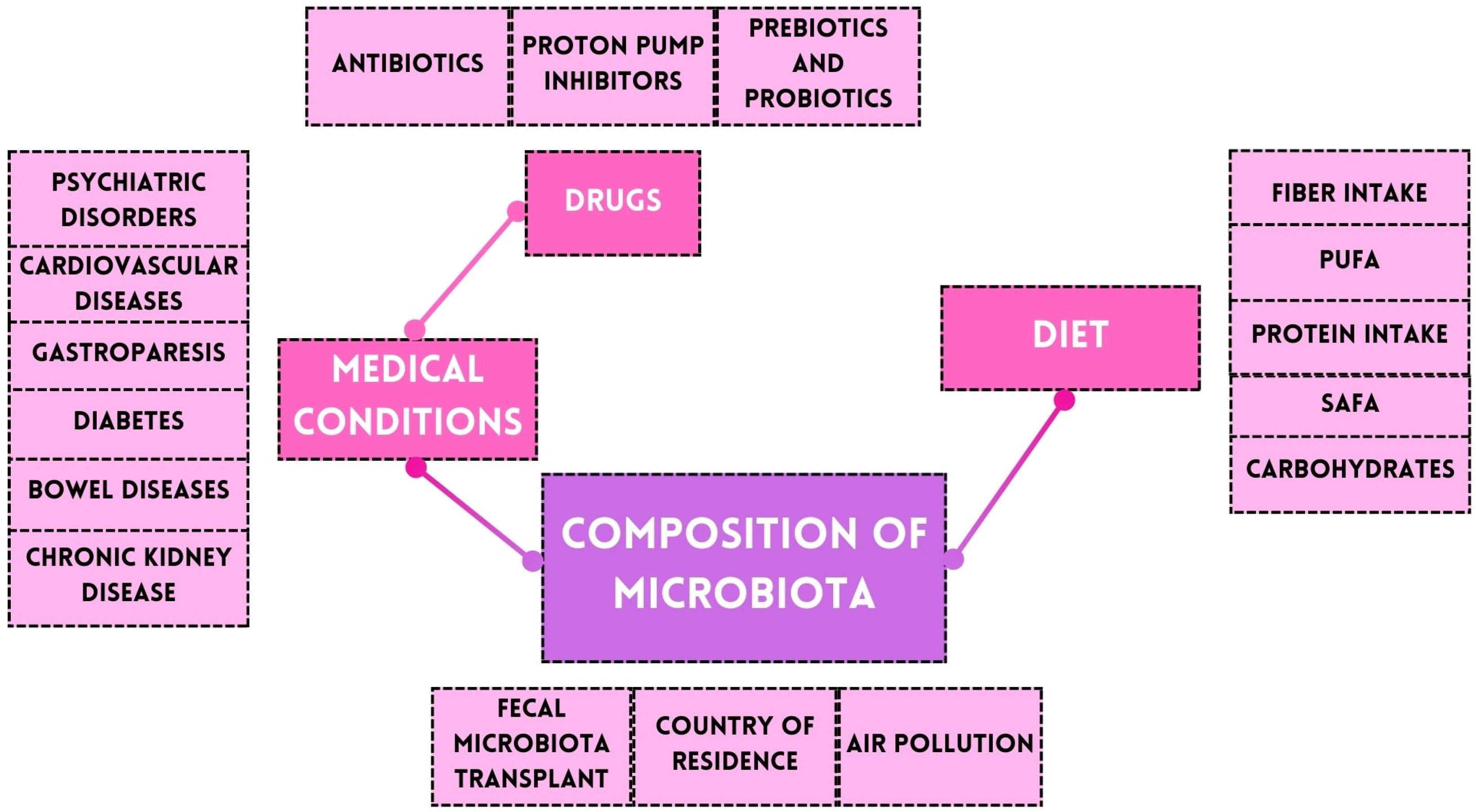
3. Microbiota and Gut–Brain Axis
Influence of Gut Microbiota Metabolites and Neurotransmitters
4. Diet
4.1. Mediterranean Diet
4.2. The Mediterranean-DASH Intervention for Neurodegenerative Delay (MIND) Diet
4.3. Vegetarian Diet
4.4. The Ketogenic Diet
4.5. Time Restriction in Eating (TRE) and Intermittent Fasting (IFA)
4.6. Ultra-Processed Foods (UPFs)
5. Role of Supplementation on Cognitive Functions
6. Potential Microbiota-Targeted Therapies for Cognitive Impairment Disorders
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bettio, L.E.B.; Rajendran, L.; Gil-Mohapel, J. The Effects of Aging in the Hippocampus and Cognitive Decline. Neurosci. Biobehav. Rev. 2017, 79, 66–86. [Google Scholar] [CrossRef] [PubMed]
- Kandlur, A.; Satyamoorthy, K.; Gangadharan, G. Oxidative Stress in Cognitive and Epigenetic Aging: A Retrospective Glance. Front. Mol. Neurosci. 2020, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Marzola, P.; Melzer, T.; Pavesi, E.; Gil-Mohapel, J.; Brocardo, P.S. Exploring the Role of Neuroplasticity in Development, Aging, and Neurodegeneration. Brain Sci. 2023, 13, 1610. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Cai, S.Q. Mechanisms Underlying Brain Aging Under Normal and Pathological Conditions. Neurosci. Bull. 2023, 39, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, B.; Rypma, B.; Gratton, G.; Fabiani, M. Age-Related Changes in Cerebrovascular Health and Their Effects on Neural Function and Cognition: A Comprehensive Review. Psychophysiology 2021, 58, e13796. [Google Scholar] [CrossRef]
- Mattson, M.P.; Arumugam, T.V. Hallmarks of Brain Aging: Adaptive and Pathological Modification by Metabolic States. Cell Metab. 2018, 27, 1176–1199. [Google Scholar] [CrossRef]
- Mormino, E.C.; Papp, K.V. Amyloid Accumulation and Cognitive Decline in Clinically Normal Older Individuals: Implications for Aging and Early Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 64, S633–S646. [Google Scholar] [CrossRef]
- Aarsland, D.; Creese, B.; Politis, M.; Chaudhuri, K.R.; Ffytche, D.H.; Weintraub, D.; Ballard, C. Cognitive Decline in Parkinson Disease. Nat. Rev. Neurol. 2017, 13, 217–231. [Google Scholar] [CrossRef]
- Brisch, R.; Saniotis, A.; Wolf, R.; Bielau, H.; Bernstein, H.G.; Steiner, J.; Bogerts, B.; Braun, K.; Kumaratilake, J.; Henneberg, M.; et al. The Role of Dopamine in Schizophrenia from a Neurobiological and Evolutionary Perspective: Old Fashioned, but Still in Vogue. Front. Psychiatry 2014, 5, 47. [Google Scholar]
- Li, H.; Hirano, S.; Furukawa, S.; Nakano, Y.; Kojima, K.; Ishikawa, A.; Tai, H.; Horikoshi, T.; Iimori, T.; Uno, T.; et al. The Relationship Between the Striatal Dopaminergic Neuronal and Cognitive Function with Aging. Front. Aging Neurosci. 2020, 12, 41. [Google Scholar] [CrossRef]
- Štrac, D.Š.; Pivac, N.; Mück-Šeler, D. The Serotonergic System and Cognitive Function. Transl. Neurosci. 2016, 7, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Reddy-Thootkur, M.; Kraguljac, N.V.; Lahti, A.C. The Role of Glutamate and GABA in Cognitive Dysfunction in Schizophrenia and Mood Disorders—A Systematic Review of Magnetic Resonance Spectroscopy Studies. Schizophr. Res. 2022, 249, 74–84. [Google Scholar] [CrossRef]
- Fourrier, C.; Singhal, G.; Baune, B.T. Neuroinflammation and Cognition across Psychiatric Conditions. CNS Spectrums 2019, 24, 4–15. [Google Scholar] [CrossRef]
- Yang, J.; Ran, M.; Li, H.; Lin, Y.; Ma, K.; Yang, Y.; Fu, X.; Yang, S. New Insight into Neurological Degeneration: Inflammatory Cytokines and Blood–Brain Barrier. Front. Mol. Neurosci. 2022, 15, 1013933. [Google Scholar] [CrossRef]
- Gąssowska-Dobrowolska, M.; Chlubek, M.; Kolasa, A.; Tomasiak, P.; Korbecki, J.; Skowrońska, K.; Tarnowski, M.; Masztalewicz, M.; Baranowska-Bosiacka, I. Microglia and Astroglia—The Potential Role in Neuroinflammation Induced by Pre- and Neonatal Exposure to Lead (Pb). Int. J. Mol. Sci. 2023, 24, 9903. [Google Scholar] [CrossRef] [PubMed]
- Franzoni, F.; Scarfò, G.; Guidotti, S.; Fusi, J.; Asomov, M.; Pruneti, C. Oxidative Stress and Cognitive Decline: The Neuroprotective Role of Natural Antioxidants. Front. Neurosci. 2021, 15, 729757. [Google Scholar] [CrossRef]
- Daniels, T.E.; Olsen, E.M.; Tyrka, A.R. Stress and Psychiatric Disorders: The Role of Mitochondria. Annu. Rev. Clin. Psychol. 2020, 16, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Teleanu, D.M.; Niculescu, A.G.; Lungu, I.I.; Radu, C.I.; Vladâcenco, O.; Roza, E.; Costăchescu, B.; Grumezescu, A.M.; Teleanu, R.I. An Overview of Oxidative Stress, Neuroinflammation and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef]
- Biessels, G.; Despa, F. Cognitive Decline and Dementia in Diabetes Mellitus: Mechanisms and Clinical Implications. Nat. Rev. Endocrinol. 2018, 14, 591–604. [Google Scholar] [CrossRef]
- Croall, I.D.; Tooth, C.; Venneri, A.; Poyser, C.; Sanders, D.S.; Hoggard, N.; Hadjivassiliou, M. Cognitive Impairment in Coeliac Disease with Respect to Disease Duration and Gluten-Free Diet Adherence: A Pilot Study. Nutrients 2020, 12, 2028. [Google Scholar] [CrossRef]
- Saghafi-Asl, M.; Aliasgharzadeh, S.; Asghari-Jafarabadi, M. Factors Influencing Weight Management Behavior among College Students: An Application of the Health Belief Model. PLoS ONE 2020, 15, e0228058. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hu, Y.; Zhang, W.; Wang, J.; Ji, W.; Manza, P.; Volkow, N.D.; Zhang, Y.; Wang, G.J. Brain Functional and Structural Magnetic Resonance Imaging of Obesity and Weight Loss Interventions. Mol. Psychiatry 2023, 28, 1466–1479. [Google Scholar] [CrossRef]
- Anstey, K.J.; Cherbuin, N.; Budge, M.; Young, J. Body Mass Index in Midlife and Late-Life as a Risk Factor for Dementia: A Meta-Analysis of Prospective Studies. Obes. Rev. 2011, 12, e426–e437. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, J.C.D.; Killcross, A.S.; Jenkins, T.A. Obesity and Cognitive Decline: Role of Inflammation and Vascular Changes. Front. Neurosci. 2014, 8, 375. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.; Gasbarrini, A.; Mele, M. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Beam, A.; Clinger, E.; Hao, L. Effect of Diet and Dietary Components on the Composition of the Gut Microbiota. Nutrients 2021, 13, 2795. [Google Scholar] [CrossRef]
- McCallum, G.; Tropini, C. The Gut Microbiota and Its Biogeography. Nat. Rev. Microbiol. 2024, 22, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Pantazi, A.C.; Balasa, A.L.; Mihai, C.M.; Chisnoiu, T.; Lupu, V.V.; Kassim, M.A.K.; Mihai, L.; Frecus, C.E.; Chirila, S.I.; Lupu, A.; et al. Development of Gut Microbiota in the First 1000 Days after Birth and Potential Interventions. Nutrients 2023, 15, 3647. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- Campaniello, D.; Corbo, M.R.; Sinigaglia, M.; Speranza, B.; Racioppo, A.; Altieri, C.; Bevilacqua, A. How Diet and Physical Activity Modulate Gut Microbiota: Evidence, and Perspectives. Nutrients 2022, 14, 2456. [Google Scholar] [CrossRef] [PubMed]
- Młynarska, E.; Wasiak, J.; Gajewska, A.; Steć, G.; Jasińska, J.; Rysz, J.; Franczyk, B. Exploring the Significance of Gut Microbiota in Diabetes Pathogenesis and Management-A Narrative Review. Nutrients 2024, 16, 1938. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, Z.; Song, S.; Zhang, H.; Zhang, W.; Liu, L.; Chen, Y.; Sun, J. Gut Microbiota Modulates Stress-Induced Hypertension through the HPA Axis. Brain Res. Bull. 2020, 162, 49–58. [Google Scholar] [CrossRef]
- Geng, J.; Ni, Q.; Sun, W.; Li, L.; Feng, X. The Links between Gut Microbiota and Obesity and Obesity Related Diseases. Biomed. Pharmacother. 2022, 147, 112678. [Google Scholar] [CrossRef] [PubMed]
- Ohkusa, T.; Koido, S.; Nishikawa, Y.; Sato, N. Gut Microbiota and Chronic Constipation: A Review and Update. Front. Med. 2019, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, M.; Fasulo, E.; Ungaro, F.; Massimino, L.; Sinagra, E.; Danese, S.; Mandarino, F.V. Gut Dysbiosis in Irritable Bowel Syndrome: A Narrative Review on Correlation with Disease Subtypes and Novel Therapeutic Implications. Microorganisms 2023, 11, 2369. [Google Scholar] [CrossRef]
- Mandarino, F.V.; Sinagra, E.; Barchi, A.; Verga, M.C.; Brinch, D.; Raimondo, D.; Danese, S. Gastroparesis: The Complex Interplay with Microbiota and the Role of Exogenous Infections in the Pathogenesis of the Disease. Microorganisms 2023, 11, 1122. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Islam, F.; Or-Rashid, M.H.; Al Mamun, A.; Rahaman, M.S.; Islam, M.M.; Meem, A.F.K.; Sutradhar, P.R.; Mitra, S.; Mimi, A.A.; et al. The Gut Microbiota (Microbiome) in Cardiovascular Disease and Its Therapeutic Regulation. Front. Cell. Infect. Microbiol. 2022, 12, 903570. [Google Scholar] [CrossRef]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The Gut Microbiota and the Brain–Gut–Kidney Axis in Hypertension and Chronic Kidney Disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef]
- Zmora, N.; Suez, J.; Elinav, E. You Are What You Eat: Diet, Health and the Gut Microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Colella, M.; Charitos, I.A.; Ballini, A.; Cafiero, C.; Topi, S.; Palmirotta, R.; Santacroce, L. Microbiota Revolution: How Gut Microbes Regulate Our Lives. World J. Gastroenterol. 2023, 29, 4368–4383. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G. Nutritional Psychiatry: How Diet Affects Brain through Gut Microbiota. Nutrients 2021, 13, 1282. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A.; Nance, K.; Chen, S. The Gut–Brain Axis. Annu. Rev. Med. 2022, 73, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Baumann, P.; Tüscher, O.; Schick, S.; Endres, K. The Aging Enteric Nervous System. Int. J. Mol. Sci. 2023, 24, 9471. [Google Scholar] [CrossRef]
- Cavin, J.-B.; Cuddihey, H.; MacNaughton, W.K.; Sharkey, K.A. Acute Regulation of Intestinal Ion Transport and Permeability in Response to Luminal Nutrients: The Role of the Enteric Nervous System. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 318, G254–G264. [Google Scholar] [CrossRef]
- Furness, J.B. The Enteric Nervous System and Neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and Functional Importance in the Gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Komorniak, N.; Martynova-Van Kley, A.; Nalian, A.; Wroński, M.; Kaseja, K.; Kowalewski, B.; Kaźmierczak-Siedlecka, K.; Łoniewski, I.; Kaczmarczyk, M.; Podsiadło, K.; et al. Association between Fecal Microbiota, SCFA, Gut Integrity Markers and Depressive Symptoms in Patients Treated in the Past with Bariatric Surgery—The Cross-Sectional Study. Nutrients 2022, 14, 5372. [Google Scholar] [CrossRef]
- Cheung, S.G.; Goldenthal, A.R.; Uhlemann, A.-C.; Mann, J.J.; Miller, J.M.; Sublette, M.E. Systematic Review of Gut Microbiota and Major Depression. Front. Psychiatry 2019, 10, 34. [Google Scholar] [CrossRef]
- Obrenovich, M.E.M. Leaky Gut, Leaky Brain? Microorganisms 2018, 6, 107. [Google Scholar] [CrossRef] [PubMed]
- Dicks, L.M.T. Gut Bacteria and Neurotransmitters. Microorganisms 2022, 10, 1838. [Google Scholar] [CrossRef] [PubMed]
- Wasiak, J.; Gawlik-Kotelnicka, O. Intestinal Permeability and Its Significance in Psychiatric Disorders—A Narrative Review and Future Perspectives. Behav. Brain Res. 2023, 448, 114459. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Turner, J.R. Intestinal Permeability Defects: Is It Time to Treat? Clin. Gastroenterol. Hepatol. 2013, 11, 1075–1083. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Leaky Gut-Concept or Clinical Entity? Curr. Opin. Gastroenterol. 2016, 32, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky Gut as a Danger Signal for Autoimmune Diseases. Front. Immunol. 2017, 8, 598. [Google Scholar] [CrossRef]
- Vaure, C.; Liu, Y. A Comparative Review of Toll-Like Receptor 4 Expression and Functionality in Different Animal Species. Front. Immunol. 2014, 5, 316. [Google Scholar] [CrossRef]
- Brown, G.C. The Endotoxin Hypothesis of Neurodegeneration. J. Neuroinflamm. 2019, 16, 180. [Google Scholar] [CrossRef]
- Sandiego, C.M.; Gallezot, J.-D.; Pittman, B.; Nabulsi, N.; Lim, K.; Lin, S.-F.; Matuskey, D.; Lee, J.-Y.; O’Connor, K.C.; Huang, Y.; et al. Imaging Robust Microglial Activation after Lipopolysaccharide Administration in Humans with PET. Proc. Natl. Acad. Sci. USA 2015, 112, 12468–12473. [Google Scholar] [CrossRef]
- Bodea, L.-G.; Wang, Y.; Linnartz-Gerlach, B.; Kopatz, J.; Sinkkonen, L.; Musgrove, R.; Kaoma, T.; Muller, A.; Vallar, L.; Di Monte, D.A.; et al. Neurodegeneration by Activation of the Microglial Complement-Phagosome Pathway. J. Neurosci. 2014, 34, 8546–8556. [Google Scholar] [CrossRef]
- Misiak, B.; Łoniewski, I.; Marlicz, W.; Frydecka, D.; Szulc, A.; Rudzki, L.; Samochowiec, J. The HPA Axis Dysregulation in Severe Mental Illness: Can We Shift the Blame to Gut Microbiota? Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 102, 109951. [Google Scholar] [CrossRef] [PubMed]
- Kinashi, Y.; Hase, K. Partners in Leaky Gut Syndrome: Intestinal Dysbiosis and Autoimmunity. Front. Immunol. 2021, 12, 673708. [Google Scholar] [CrossRef]
- Kealy, J.; Greene, C.; Campbell, M. Blood-Brain Barrier Regulation in Psychiatric Disorders. Neurosci. Lett. 2020, 726, 133664. [Google Scholar] [CrossRef]
- Daneman, R.; Prat, A. The Blood–Brain Barrier. Cold Spring Harb. Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef]
- Rudzki, L.; Maes, M. The Microbiota-Gut-Immune-Glia (MGIG) Axis in Major Depression. Mol. Neurobiol. 2020, 57, 4269–4295. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Astrocyte Barriers to Neurotoxic Inflammation. Nat. Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Dienel, S.J.; Enwright, J.F.; Hoftman, G.D.; Lewis, D.A. Markers of Glutamate and GABA Neurotransmission in the Prefrontal Cortex of Schizophrenia Subjects: Disease Effects Differ across Anatomical Levels of Resolution. Schizophr. Res. 2020, 217, 86–94. [Google Scholar] [CrossRef]
- Brown, G.; Heneka, M. The Endotoxin Hypothesis of Alzheimer’s Disease. Mol. Neurodegener. 2024, 19, 30. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C.; Camacho, M.; Williams-Gray, C.H. The Endotoxin Hypothesis of Parkinson’s Disease. Mov. Disord. 2023, 38, 1143–1155. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, Y.F.; Miao, J.; Zheng, R.F.; Li, J.Y. Short-Chain Fatty Acids: Important Components of the Gut-Brain Axis against AD. Biomed. Pharmacother. 2024, 175, 116601. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Cheng, G.; Hardy, M. Gut Microbiome, Short-Chain Fatty Acids, Alpha-Synuclein, Neuroinflammation, and ROS/RNS: Relevance to Parkinson’s Disease and Therapeutic Implications. Redox Biol. 2024, 71, 103092. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, J.; Chen, Y. Regulation of Neurotransmitters by the Gut Microbiota and Effects on Cognition in Neurological Disorders. Nutrients 2021, 13, 2099. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The Short-Chain Fatty Acid Acetate Reduces Appetite via a Central Homeostatic Mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-Modulating Bacteria of the Human Gut Microbiota. Nat. Microbiol. 2018, 4, 396–403. [Google Scholar] [CrossRef]
- Estrela, A.B.; Abraham, W.-R. Adenosine in the Inflamed Gut: A Janus Faced Compound. Curr. Med. Chem. 2011, 18, 2791–2815. [Google Scholar] [CrossRef]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The Critical Modulators Regulating Gut–Brain Axis. J. Cell. Physiol. 2017, 232, 2359–2372. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The Role of Short-Chain Fatty Acids in Microbiota–Gut–Brain Communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- González-Arancibia, C.; Urrutia-Piñones, J.; Illanes-González, J.; Martinez-Pinto, J.; Sotomayor-Zárate, R.; Julio-Pieper, M.; Bravo, J.A. Do Your Gut Microbes Affect Your Brain Dopamine? Psychopharmacology 2019, 236, 1611–1622. [Google Scholar] [CrossRef] [PubMed]
- Rudzki, L.; Maes, M. From “Leaky Gut” to Impaired Glia-Neuron Communication in Depression. In Major Depressive Disorder: Rethinking and Understanding Recent Discoveries; Springer: Singapore, 2021; pp. 129–155. [Google Scholar]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine Pathway Metabolism and the Microbiota-Gut-Brain Axis. Neuropharmacology 2017, 112, 399–412. [Google Scholar] [CrossRef]
- Puri, S.; Shaheen, M.; Grover, B. Nutrition and Cognitive Health: A Life Course Approach. Front. Public Health 2023, 11, 1023907. [Google Scholar] [CrossRef]
- Attuquayefio, T.; Stevenson, R.J. A Systematic Review of Longer-Term Dietary Interventions on Human Cognitive Function: Emerging Patterns and Future Directions. Appetite 2015, 95, 554–570. [Google Scholar] [CrossRef] [PubMed]
- Fortune, N.C.; Harville, E.W.; Guralnik, J.M.; Gustat, J.; Chen, W.; Qi, L.; Bazzano, L.A. Dietary Intake and Cognitive Function: Evidence from the Bogalusa Heart Study. Am. J. Clin. Nutr. 2019, 109, 1656–1663. [Google Scholar] [CrossRef] [PubMed]
- Petersson, S.D.; Philippou, E. Mediterranean Diet, Cognitive Function, and Dementia: A Systematic Review of the Evidence. Adv. Nutr. 2016, 7, 889–904. [Google Scholar] [CrossRef] [PubMed]
- Siervo, M.; Shannon, O.M.; Llewellyn, D.J.; Stephan, B.C.; Fontana, L. Mediterranean Diet and Cognitive Function: From Methodology to Mechanisms of Action. Free Radic. Biol. Med. 2021, 176, 105–117. [Google Scholar] [CrossRef]
- Valls-Pedret, C.; Sala-Vila, A.; Serra-Mir, M.; Corella, D.; de la Torre, R.; Martínez-González, M.Á.; Martínez-Lapiscina, E.H.; Fitó, M.; Pérez-Heras, A.; Salas-Salvadó, J.; et al. Mediterranean Diet and Age-Related Cognitive Decline. JAMA Intern Med. 2015, 175, 1094. [Google Scholar] [CrossRef]
- Cena, H.; Calder, P.C. Defining a Healthy Diet: Evidence for the Role of Contemporary Dietary Patterns in Health and Disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef]
- Gu, Y.; Luchsinger, J.A.; Stern, Y.; Scarmeas, N. Mediterranean Diet, Inflammatory and Metabolic Biomarkers, and Risk of Alzheimer’s Disease. J. Alzheimer’s Dis. 2010, 22, 483–492. [Google Scholar] [CrossRef]
- Féart, C. Adherence to a Mediterranean Diet, Cognitive Decline, and Risk of Dementia. JAMA 2009, 302, 638. [Google Scholar] [CrossRef]
- Pelletier, A.; Barul, C.; Féart, C.; Helmer, C.; Bernard, C.; Periot, O.; Dilharreguy, B.; Dartigues, J.; Allard, M.; Barberger-Gateau, P.; et al. Mediterranean Diet and Preserved Brain Structural Connectivity in Older Subjects. Alzheimer’s Dement. 2015, 11, 1023–1031. [Google Scholar] [CrossRef]
- World Health Organization. Risk Reduction of Cognitive Decline and Dementia: WHO Guidelines; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Kopf, J.C.; Suhr, M.J.; Clarke, J.; Eyun, S.; Riethoven, J.-J.M.; Ramer-Tait, A.E.; Rose, D.J. Role of Whole Grains versus Fruits and Vegetables in Reducing Subclinical Inflammation and Promoting Gastrointestinal Health in Individuals Affected by Overweight and Obesity: A Randomized Controlled Trial. Nutr. J. 2018, 17, 72. [Google Scholar] [CrossRef]
- Liu, X.; Morris, M.C.; Dhana, K.; Ventrelle, J.; Johnson, K.; Bishop, L.; Hollings, C.S.; Boulin, A.; Laranjo, N.; Stubbs, B.J.; et al. Mediterranean-DASH Intervention for Neurodegenerative Delay (MIND) Study: Rationale, Design and Baseline Characteristics of a Randomized Control Trial of the MIND Diet on Cognitive Decline. Contemp. Clin. Trials 2021, 102, 106270. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Tangney, C.C.; Wang, Y.; Sacks, F.M.; Barnes, L.L.; Bennett, D.A.; Aggarwal, N.T. MIND Diet Slows Cognitive Decline with Aging. Alzheimer’s Dement. 2015, 11, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Barnes, L.L.; Dhana, K.; Liu, X.; Carey, V.J.; Ventrelle, J.; Johnson, K.; Hollings, C.S.; Bishop, L.; Laranjo, N.; Stubbs, B.J.; et al. Trial of the MIND Diet for Prevention of Cognitive Decline in Older Persons. N. Engl. J. Med. 2023, 389, 602–611. [Google Scholar] [CrossRef]
- Medawar, E.; Huhn, S.; Villringer, A.; Veronica Witte, A. The Effects of Plant-Based Diets on the Body and the Brain: A Systematic Review. Transl. Psychiatry 2019, 9, 226. [Google Scholar] [CrossRef]
- Rajaram, S.; Jones, J.; Lee, G.J. Plant-Based Dietary Patterns, Plant Foods, and Age-Related Cognitive Decline. Adv. Nutr. 2019, 10, S422–S436. [Google Scholar] [CrossRef]
- Katonova, A.; Sheardova, K.; Amlerova, J.; Angelucci, F.; Hort, J. Effect of a Vegan Diet on Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 14924. [Google Scholar] [CrossRef] [PubMed]
- Devranis, P.; Vassilopoulou, Ε.; Tsironis, V.; Sotiriadis, P.M.; Chourdakis, M.; Aivaliotis, M.; Tsolaki, M. Mediterranean Diet, Ketogenic Diet or MIND Diet for Aging Populations with Cognitive Decline: A Systematic Review. Life 2023, 13, 173. [Google Scholar] [CrossRef]
- Rong, L.; Peng, Y.; Shen, Q.; Chen, K.; Fang, B.; Li, W. Effects of Ketogenic Diet on Cognitive Function of Patients with Alzheimer’s Disease: A Systematic Review and Meta-Analysis. J. Nutr. Health Aging 2024, 28, 100306. [Google Scholar] [CrossRef]
- Price, S.; Ruppar, T.M. Ketogenic Therapies in Parkinson’s Disease, Alzheimer’s Disease, and Mild Cognitive Impairment: An Integrative Review. Appl. Nurs. Res. 2023, 74, 151745. [Google Scholar] [CrossRef]
- Grochowska, K.; Przeliorz, A. The Effect of the Ketogenic Diet on the Therapy of Neurodegenerative Diseases and Its Impact on Improving Cognitive Functions. Dement Geriatr. Cogn. Dis. Extra 2022, 12, 100–106. [Google Scholar] [CrossRef]
- Sharifi, S.; Rostami, F.; Babaei Khorzoughi, K.; Rahmati, M. Effect of Time-Restricted Eating and Intermittent Fasting on Cognitive Function and Mental Health in Older Adults: A Systematic Review. Prev. Med. Rep. 2024, 42, 102757. [Google Scholar] [CrossRef] [PubMed]
- Elias, A.; Padinjakara, N.; Lautenschlager, N.T. Effects of Intermittent Fasting on Cognitive Health and Alzheimer’s Disease. Nutr. Rev. 2023, 81, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wang, Y.; Zhang, X.; Ma, M.; Xie, Z.; Pan, Q.; Ma, Z.; Peppelenbosch, M.P. Remodeling of the Gut Microbiome during Ramadan-Associated Intermittent Fasting. Am. J. Clin. Nutr. 2021, 113, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth, L.; Machado, P.; Zinöcker, M.; Baker, P.; Lawrence, M. Ultra-Processed Foods and Health Outcomes: A Narrative Review. Nutrients 2020, 12, 1955. [Google Scholar] [CrossRef]
- Martínez Leo, E.E.; Segura Campos, M.R. Effect of ultra-processed diet on gut microbiota and thus its role in neurodegenerative diseases. Nutrition 2020, 71, 110609. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Song, R.; Liu, Y.; Wu, Z.; Zhang, X. Effects of Ultra-Processed Foods on the Microbiota-Gut-Brain Axis: The Bread-and-Butter Issue. Food Res. Int. 2023, 167, 112730. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, D.; Zhang, Y.; Wang, J.; Dong, L.; Hu, Y.; Wang, S. Long-Term Exposure to Advanced Lipid Peroxidation End Products Impairs Cognitive Function through Microbiota-Gut-Brain Axis. Food Chem. 2024, 461, 140864. [Google Scholar] [CrossRef]
- R Cardoso, B.; Machado, P.; Steele, E.M. Association between Ultra-Processed Food Consumption and Cognitive Performance in US Older Adults: A Cross-Sectional Analysis of the NHANES 2011–2014. Eur. J. Nutr. 2022, 61, 3975–3985. [Google Scholar] [CrossRef]
- Pourmotabbed, A.; Talebi, S.; Mehrabani, S.; Babaei, A.; Khosroshahi, R.A.; Bagheri, R.; Wong, A.; Ghoreishy, S.M.; Amirian, P.; Zarpoosh, M.; et al. The Association of Ultra-Processed Food Intake with Neurodegenerative Disorders: A Systematic Review and Dose-Response Meta-Analysis of Large-Scale Cohorts. Nutr. Neurosci. 2024, 1–14. [Google Scholar] [CrossRef]
- Claudino, P.A.; Bueno, N.B.; Piloneto, S.; Halaiko, D.; Azevedo de Sousa, L.P.; Barroso Jara Maia, C.H.; Netto, B.D.M. Consumption of Ultra-Processed Foods and Risk for Alzheimer’s Disease: A Systematic Review. Front. Nutr. 2023, 10, 1288749. [Google Scholar] [CrossRef]
- Fan, H.; Yuan, M.; Wang, S.; Yang, X.; Shu, L.; Pu, Y.; Zou, Q.; Zhang, X.; Wang, C.; Cai, Z. Dietary Salt Promotes Cognitive Impairment through Repression of SIRT3/PINK1-Mediated Mitophagy and Fission. Mol. Cell. Biochem. 2024. [Google Scholar] [CrossRef] [PubMed]
- D’Cunha, N.M.; Sergi, D.; Lane, M.M.; Naumovski, N.; Gamage, E.; Rajendran, A.; Kouvari, M.; Gauci, S.; Dissanayka, T.; Marx, W.; et al. The Effects of Dietary Advanced Glycation End-Products on Neurocognitive and Mental Disorders. Nutrients 2022, 14, 2421. [Google Scholar] [CrossRef] [PubMed]
- Bhave, V.M.; Oladele, C.R.; Ament, Z.; Kijpaisalratana, N.; Jones, A.C.; Couch, C.A.; Patki, A.; Garcia Guarniz, A.-L.; Bennett, A.; Crowe, M.; et al. Associations Between Ultra-Processed Food Consumption and Adverse Brain Health Outcomes. Neurology 2024, 102, e209432. [Google Scholar] [CrossRef]
- Li, J.-M.; Yu, R.; Zhang, L.-P.; Wen, S.-Y.; Wang, S.-J.; Zhang, X.-Y.; Xu, Q.; Kong, L.-D. Dietary Fructose-Induced Gut Dysbiosis Promotes Mouse Hippocampal Neuroinflammation: A Benefit of Short-Chain Fatty Acids. Microbiome 2019, 7, 98. [Google Scholar] [CrossRef]
- Ayten, Ş.; Bilici, S. Modulation of Gut Microbiota Through Dietary Intervention in Neuroinflammation and Alzheimer’s and Parkinson’s Diseases. Curr. Nutr. Rep. 2024, 13, 82–96. [Google Scholar] [CrossRef]
- Magnusson, K.R.; Hauck, L.; Jeffrey, B.M.; Elias, V.; Humphrey, A.; Nath, R.; Perrone, A.; Bermudez, L.E. Relationships between Diet-Related Changes in the Gut Microbiome and Cognitive Flexibility. Neuroscience 2015, 300, 128–140. [Google Scholar] [CrossRef]
- Dhana, K.; Franco, O.H.; Ritz, E.M.; Ford, C.N.; Desai, P.; Krueger, K.R.; Holland, T.M.; Dhana, A.; Liu, X.; Aggarwal, N.T.; et al. Healthy Lifestyle and Life Expectancy with and without Alzheimer’s Dementia: Population Based Cohort Study. BMJ 2022, 377, e068390. [Google Scholar] [CrossRef]
- Dhana, K.; Evans, D.A.; Rajan, K.B.; Bennett, D.A.; Morris, M.C. Healthy Lifestyle and the Risk of Alzheimer Dementia: Findings from 2 Longitudinal Studies. Neurology 2020, 95, e374–e383. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Wu, T.; Zhao, J.; Han, F.; Marseglia, A.; Liu, H.; Huang, G. Effects of 6-Month Folic Acid Supplementation on Cognitive Function and Blood Biomarkers in Mild Cognitive Impairment: A Randomized Controlled Trial in China. J. Gerontol.—Ser. A Biol. Sci. Med. Sci. 2016, 71, 1376–1383. [Google Scholar] [CrossRef]
- Ma, F.; Li, Q.; Zhou, X.; Zhao, J.; Song, A.; Li, W.; Liu, H.; Xu, W.; Huang, G. Effects of Folic Acid Supplementation on Cognitive Function and Aβ-Related Biomarkers in Mild Cognitive Impairment: A Randomized Controlled Trial. Eur. J. Nutr. 2019, 58, 345–356. [Google Scholar] [CrossRef]
- Malouf, R.; Evans, J.G. Folic Acid with or without Vitamin B12 for the Prevention and Treatment of Healthy Elderly and Demented People. Cochrane Database Syst. Rev. 2008, 2008, CD004514. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhou, X.; Li, Q.; Zhao, J.; Song, A.; An, P.; Du, Y.; Xu, W.; Huang, G. Effects of Folic Acid and Vitamin B12, Alone and in Combination on Cognitive Function and Inflammatory Factors in the Elderly with Mild Cognitive Impairment: A Single-Blind Experimental Design. Curr. Alzheimer Res. 2019, 16, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, S.; Ge, B.; Zhou, D.; Li, M.; Li, W.; Ma, F.; Liu, Z.; Ji, Y.; Huang, G. Effects of Folic Acid and Vitamin B12 Supplementation on Cognitive Impairment and Inflammation in Patients with Alzheimer’s Disease: A Randomized, Single-Blinded, Placebo-Controlled Trial. J. Prev. Alzheimer’s Dis. 2021, 8, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lu, C.J.; Chien, K.L.; Chen, S.T.; Chen, R.C. Efficacy of Multivitamin Supplementation Containing Vitamins B6 and B12 and Folic Acid as Adjunctive Treatment with a Cholinesterase Inhibitor in Alzheimer’s Disease: A 26-Week, Randomized, Double-Blind, Placebo-Controlled Study in Taiwanese Patients. Clin. Ther. 2007, 29, 2204–2214. [Google Scholar] [CrossRef]
- Li, M.; Li, W.; Gao, Y.; Chen, Y.; Bai, D.; Weng, J.; Du, Y.; Ma, F.; Wang, X.; Liu, H.; et al. Effect of Folic Acid Combined with Docosahexaenoic Acid Intervention on Mild Cognitive Impairment in Elderly: A Randomized Double-Blind, Placebo-Controlled Trial. Eur. J. Nutr. 2021, 60, 1795–1808. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Miao, R.; Li, Q.; Wu, T.; Ma, F. Effects of DHA Supplementation on Hippocampal Volume and Cognitive Function in Older Adults with Mild Cognitive Impairment: A 12-Month Randomized, Double-Blind, Placebo-Controlled Trial. J. Alzheimer’s Dis. 2017, 55, 497–507. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Lou, Y.; Hu, J.; Miao, R.; Ma, F. DHA Supplementation Improves Cognitive Function via Enhancing Aβ-Mediated Autophagy in Chinese Elderly with Mild Cognitive Impairment: A Randomised Placebo-Controlled Trial. J. Neurol. Neurosurg. Psychiatry 2018, 89, 382–388. [Google Scholar] [CrossRef]
- Atmadja, T.; Kusharto, C.; Sinaga, T. Supplementation of Catfish (Clarias Gariepinus) Oil Enriched with Omega-3 Soft Capsule Improves Oxidative Stress and Cognitive Function in Elderly. J. Nutr. Sci. Vitaminol. 2020, 66, S47–S50. [Google Scholar] [CrossRef]
- Nolan, J.M.; Power, R.; Howard, A.N.; Bergin, P.; Roche, W.; Prado-Cabrero, A.; Pope, G.; Cooke, J.; Power, T.; Mulcahy, R. Supplementation with Carotenoids, Omega-3 Fatty Acids, and Vitamin E Has a Positive Effect on the Symptoms and Progression of Alzheimer’s Disease. J. Alzheimer’s Dis. 2022, 90, 233–249. [Google Scholar] [CrossRef]
- Patan, M.J.; Kennedy, D.O.; Husberg, C.; Hustvedt, S.O.; Calder, P.C.; Khan, J.; Forster, J.; Jackson, P.A. Supplementation with Oil Rich in Eicosapentaenoic Acid, but Not in Docosahexaenoic Acid, Improves Global Cognitive Function in Healthy, Young Adults: Results from Randomized Controlled Trials. Am. J. Clin. Nutr. 2021, 114, 914–924. [Google Scholar] [CrossRef]
- Djuricic, I.; Calder, P.C. Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef] [PubMed]
- Glück, T.; Alter, P. Marine Omega-3 Highly Unsaturated Fatty Acids: From Mechanisms to Clinical Implications in Heart Failure and Arrhythmias. Vasc. Pharmacol. 2016, 82, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Poddar, J.; Pradhan, M.; Ganguly, G.; Chakrabarti, S. Biochemical Deficits and Cognitive Decline in Brain Aging: Intervention by Dietary Supplements. J. Chem. Neuroanat. 2019, 95, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Power, R.; Nolan, J.M.; Prado-Cabrero, A.; Roche, W.; Coen, R.; Power, T.; Mulcahy, R. Omega-3 Fatty Acid, Carotenoid and Vitamin E Supplementation Improves Working Memory in Older Adults: A Randomised Clinical Trial. Clin. Nutr. 2022, 41, 405–414. [Google Scholar] [CrossRef]
- Danthiir, V.; Burns, N.R.; Nettelbeck, T.; Wilson, C.; Wittert, G. The Older People, Omega-3, and Cognitive Health (EPOCH) Trial Design and Methodology: A Randomised, Double-Blind, Controlled Trial Investigating the Effect of Long-Chain Omega-3 Fatty Acids on Cognitive Ageing and Wellbeing in Cognitively Healthy Older Adults. Nutr. J. 2011, 10, 117. [Google Scholar] [CrossRef]
- Chew, E.Y.; Clemons, T.E.; Agrón, E.; Launer, L.J.; Grodstein, F.; Bernstein, P.S. Effect of Omega-3 Fatty Acids, Lutein/Zeaxanthin, or Other Nutrient Supplementation on Cognitive Function: The AREDS2 Randomized Clinical Trial. JAMA 2015, 314, 791–801. [Google Scholar] [CrossRef]
- Giudici, K.V.; de Souto Barreto, P.; Beard, J.; Cantet, C.; Araujo de Carvalho, I.; Rolland, Y.; Vellas, B. Effect of Long-Term Omega-3 Supplementation and a Lifestyle Multidomain Intervention on Intrinsic Capacity among Community-Dwelling Older Adults: Secondary Analysis of a Randomized, Placebo-Controlled Trial (MAPT Study). Maturitas 2020, 141, 39–45. [Google Scholar] [CrossRef]
- Lin, P.Y.; Cheng, C.; Satyanarayanan, S.K.; Chiu, L.T.; Chien, Y.C.; Chuu, C.P.; Lan, T.H.; Su, K.P. Omega-3 Fatty Acids and Blood-Based Biomarkers in Alzheimer’s Disease and Mild Cognitive Impairment: A Randomized Placebo-Controlled Trial. Brain Behav. Immun. 2022, 99, 289–298. [Google Scholar] [CrossRef]
- Mengelberg, A.; Leathem, J.; Podd, J.; Hill, S.; Conlon, C. The Effects of Docosahexaenoic Acid Supplementation on Cognition and Well-Being in Mild Cognitive Impairment: A 12-Month Randomised Controlled Trial. Int. J. Geriatr. Psychiatry 2022, 37, 1–12. [Google Scholar] [CrossRef]
- Jia, J.; Hu, J.; Huo, X.; Miao, R.; Zhang, Y.; Ma, F. Effects of Vitamin D Supplementation on Cognitive Function and Blood Aβ-Related Biomarkers in Older Adults with Alzheimer’s Disease: A Randomised, Double-Blind, Placebo-Controlled Trial. J. Neurol. Neurosurg. Psychiatry 2019, 90, 1347–1352. [Google Scholar] [CrossRef]
- Ghaderi, A.; Rasouli-Azad, M.; Farhadi, M.H.; Mirhosseini, N.; Motmaen, M.; Pishyareh, E.; Omidi, A.; Asemi, Z. Exploring the Effects of Vitamin D Supplementation on Cognitive Functions and Mental Health Status in Subjects under Methadone Maintenance Treatment. J. Addict. Med. 2020, 14, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, Y.; Meng, X. Vitamin D and Neurodegenerative Diseases. Heliyon 2023, 9, e12877. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, R.; Diaz Bustamante, L.; Ghattas, K.N.; Ilyas, S.; Al-Refai, R.; Khan, S. Role of Lifestyle in Neuroplasticity and Neurogenesis in an Aging Brain. Cureus 2020, 12, e10639. [Google Scholar] [CrossRef]
- Deluca, G.C.; Kimball, S.M.; Kolasinski, J.; Ramagopalan, S.V.; Ebers, G.C. Review: The Role of Vitamin D in Nervous System Health and Disease. Neuropathol. Appl. Neurobiol. 2013, 39, 458–484. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, H.; Xiong, Y.; Chen, C.; Duan, K.; Jia, J.; Ma, F. Vitamin D Supplementation Improves Cognitive Function through Reducing Oxidative Stress Regulated by Telomere Length in Older Adults with Mild Cognitive Impairment: A 12-Month Randomized Controlled Trial. J. Alzheimer’s Dis. 2020, 78, 1509–1518. [Google Scholar] [CrossRef]
- Klimova, B.; Dziuba, S.; Cierniak-Emerych, A. The Effect of Healthy Diet on Cognitive Performance Among Healthy Seniors—A Mini Review. Front. Hum. Neurosci. 2020, 14, 325. [Google Scholar] [CrossRef]
- Mccleery, J.; Abraham, R.P.; Denton, D.A.; Rutjes, A.W.S.; Chong, L.Y.; Al-Assaf, A.S.; Griffith, D.J.; Rafeeq, S.; Yaman, H.; Malik, M.A.; et al. Vitamin and Mineral Supplementation for Preventing Dementia or Delaying Cognitive Decline in People with Mild Cognitive Impairment. Cochrane Database Syst. Rev. 2018, 2018, CD011905. [Google Scholar] [CrossRef]
- Naomi, R.; Embong, H.; Othman, F.; Ghazi, H.F.; Maruthey, N.; Bahari, H. Probiotics for Alzheimer’s Disease: A Systematic Review. Nutrients 2022, 14, 20. [Google Scholar] [CrossRef]
- Tan, A.H.; Hor, J.W.; Chong, C.W.; Lim, S.Y. Probiotics for Parkinson’s Disease: Current Evidence and Future Directions. JGH Open 2021, 5, 414–419. [Google Scholar] [CrossRef]
- Liu, Q.; Xi, Y.; Wang, Q.; Liu, J.; Li, P.; Meng, X.; Liu, K.; Chen, W.; Liu, X.; Liu, Z. Mannan Oligosaccharide Attenuates Cognitive and Behavioral Disorders in the 5xFAD Alzheimer’s Disease Mouse Model via Regulating the Gut Microbiota-Brain Axis. Brain Behav. Immun. 2021, 95, 330–343. [Google Scholar] [CrossRef]
- Castelli, V.; d’Angelo, M.; Lombardi, F.; Alfonsetti, M.; Antonosante, A.; Catanesi, M.; Benedetti, E.; Palumbo, P.; Cifone, M.G.; Giordano, A.; et al. Effects of the Probiotic Formulation SLAB51 in in Vitro and in Vivo Parkinson’s Disease Models. Aging 2020, 12, 4641–4659. [Google Scholar] [CrossRef] [PubMed]
- Nimgampalle, M. Anti-Alzheimer Properties of Probiotic, Lactobacillus Plantarum MTCC 1325 in Alzheimer’s Disease Induced Albino Rats. J. Clin. Diagn. Res. 2017, 11, KC01–KC05. [Google Scholar] [CrossRef] [PubMed]
- Imdad, A.; Minkoff, N.Z.; Tanner-Smith, E.E.; Zackular, J.P.; Acra, S.; Nicholson, M.R. Fecal Microbiota Transplantation for the Treatment of Recurrent Clostridioides difficile (Clostridium difficile). Cochrane Database Syst. Rev. 2021, 2021, CD013871. [Google Scholar] [CrossRef]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The Role of Microbiota-Gut-Brain Axis in Neuropsychiatric and Neurological Disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef]
- Warner, B.B. The Contribution of the Gut Microbiome to Neurodevelopment and Neuropsychiatric Disorders. Pediatr. Res. 2019, 85, 216–224. [Google Scholar] [CrossRef]
- Kesika, P.; Suganthy, N.; Sivamaruthi, B.S.; Chaiyasut, C. Role of Gut-Brain Axis, Gut Microbial Composition, and Probiotic Intervention in Alzheimer’s Disease. Life Sci. 2021, 264, 118627. [Google Scholar] [CrossRef]
- Bhatt, S.; Kanoujia, J.; Mohana Lakshmi, S.; Patil, C.R.; Gupta, G.; Chellappan, D.K.; Dua, K. Role of Brain-Gut-Microbiota Axis in Depression: Emerging Therapeutic Avenues. CNS Neurol. Disord. Drug Targets 2023, 22, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Wang, Y.-F.; Lei, L.; Zhang, Y. Impacts of Microbiota and Its Metabolites through Gut-Brain Axis on Pathophysiology of Major Depressive Disorder. Life Sci. 2024, 351, 122815. [Google Scholar] [CrossRef] [PubMed]
- Mossad, O.; Nent, E.; Woltemate, S.; Folschweiller, S.; Buescher, J.M.; Schnepf, D.; Erny, D.; Staeheli, P.; Bartos, M.; Szalay, A.; et al. Microbiota-Dependent Increase in δ-Valerobetaine Alters Neuronal Function and Is Responsible for Age-Related Cognitive Decline. Nat. Aging 2021, 1, 1127–1136. [Google Scholar] [CrossRef]
- Xiong, Y.; Pu, Y.; Li, L.; Su, Y.; Niu, J.; Xiao, Z. Gut Microbiota-derived Metabolite Trimethylamine N -oxide Aggravates Cognitive Dysfunction Induced by Femoral Fracture Operation in Mice. Kaohsiung J. Med. Sci. 2024, 40, 732–743. [Google Scholar] [CrossRef]
- Parletta, N.; Milte, C.M.; Meyer, B.J. Nutritional Modulation of Cognitive Function and Mental Health. J. Nutr. Biochem. 2013, 24, 725–743. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, V.E.; Herrera, P.F.; Laura, R. Effect of Nutrition on Neurodegenerative Diseases. A Systematic Review. Nutr. Neurosci. 2021, 24, 810–834. [Google Scholar] [CrossRef] [PubMed]
- Freeman, L.R.; Haley-Zitlin, V.; Rosenberger, D.S.; Granholm, A.C. Damaging Effects of a High-Fat Diet to the Brain and Cognition: A Review of Proposed Mechanisms. Nutr. Neurosci. 2014, 17, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Beilharz, J.E.; Maniam, J.; Morris, M.J. Diet-Induced Cognitive Deficits: The Role of Fat and Sugar, Potential Mechanisms and Nutritional Interventions. Nutrients 2015, 7, 6719–6738. [Google Scholar] [CrossRef]
- Van Den Brink, A.C.; Brouwer-Brolsma, E.M.; Berendsen, A.A.M.; Van De Rest, O. The Mediterranean, Dietary Approaches to Stop Hypertension (DASH), and Mediterranean-DASH Intervention for Neurodegenerative Delay (MIND) Diets Are Associated with Less Cognitive Decline and a Lower Risk of Alzheimer’s Disease—A Review. Adv. Nutr. 2019, 10, 1040–1065. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Młynarska, E.; Jakubowska, P.; Frąk, W.; Gajewska, A.; Sornowska, J.; Skwira, S.; Wasiak, J.; Rysz, J.; Franczyk, B. Associations of Microbiota and Nutrition with Cognitive Impairment in Diseases. Nutrients 2024, 16, 3570. https://doi.org/10.3390/nu16203570
Młynarska E, Jakubowska P, Frąk W, Gajewska A, Sornowska J, Skwira S, Wasiak J, Rysz J, Franczyk B. Associations of Microbiota and Nutrition with Cognitive Impairment in Diseases. Nutrients. 2024; 16(20):3570. https://doi.org/10.3390/nu16203570
Chicago/Turabian StyleMłynarska, Ewelina, Paulina Jakubowska, Weronika Frąk, Agata Gajewska, Joanna Sornowska, Sylwia Skwira, Jakub Wasiak, Jacek Rysz, and Beata Franczyk. 2024. "Associations of Microbiota and Nutrition with Cognitive Impairment in Diseases" Nutrients 16, no. 20: 3570. https://doi.org/10.3390/nu16203570
APA StyleMłynarska, E., Jakubowska, P., Frąk, W., Gajewska, A., Sornowska, J., Skwira, S., Wasiak, J., Rysz, J., & Franczyk, B. (2024). Associations of Microbiota and Nutrition with Cognitive Impairment in Diseases. Nutrients, 16(20), 3570. https://doi.org/10.3390/nu16203570