
Harnessing Prebiotics to Improve Type 2 Diabetes Outcomes


Abstract
1. Introduction
2. Effects of Specific Prebiotics on Microbial Composition and T2DM
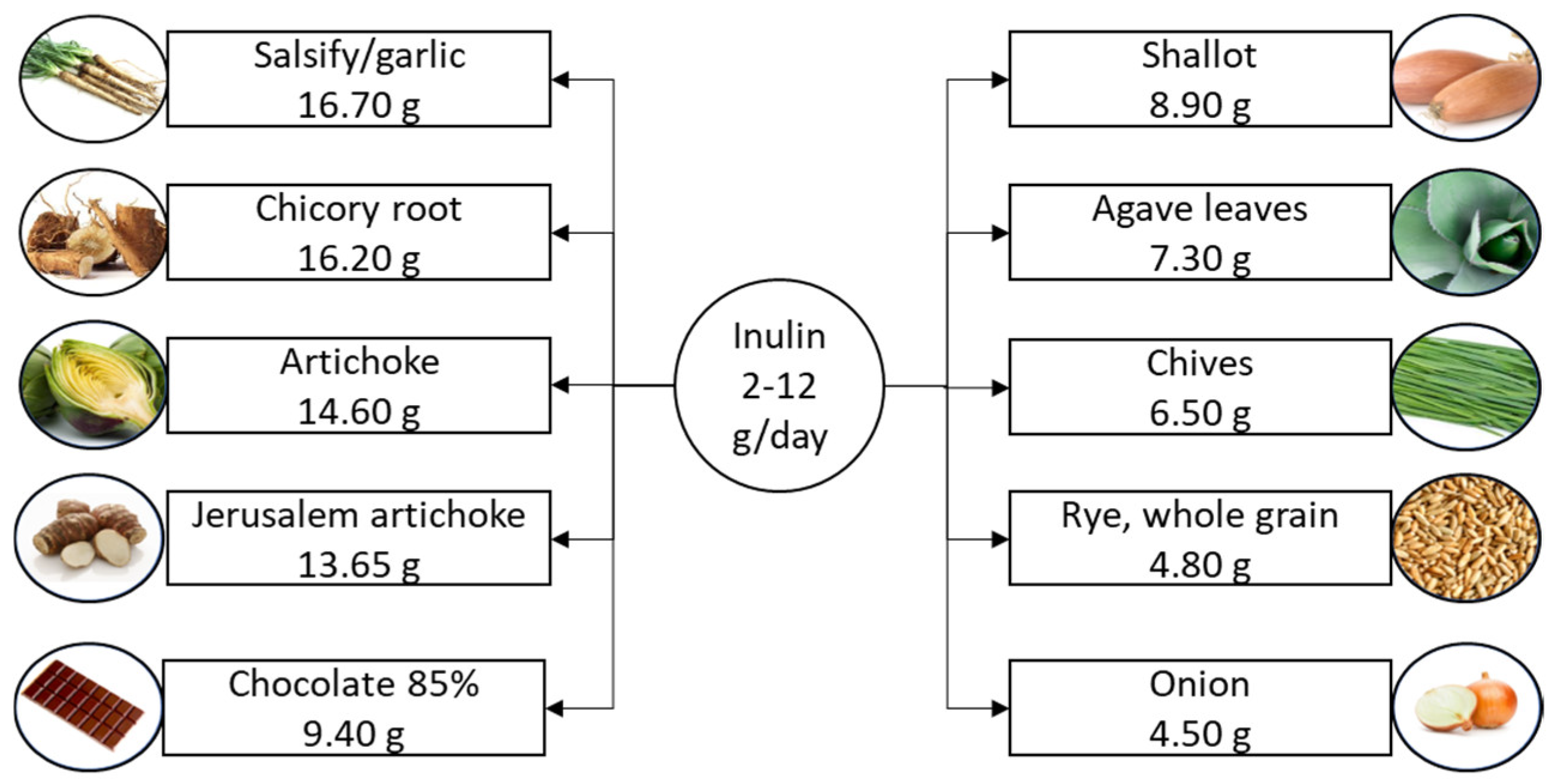
2.1. Inulin
2.2. Resistant Starch
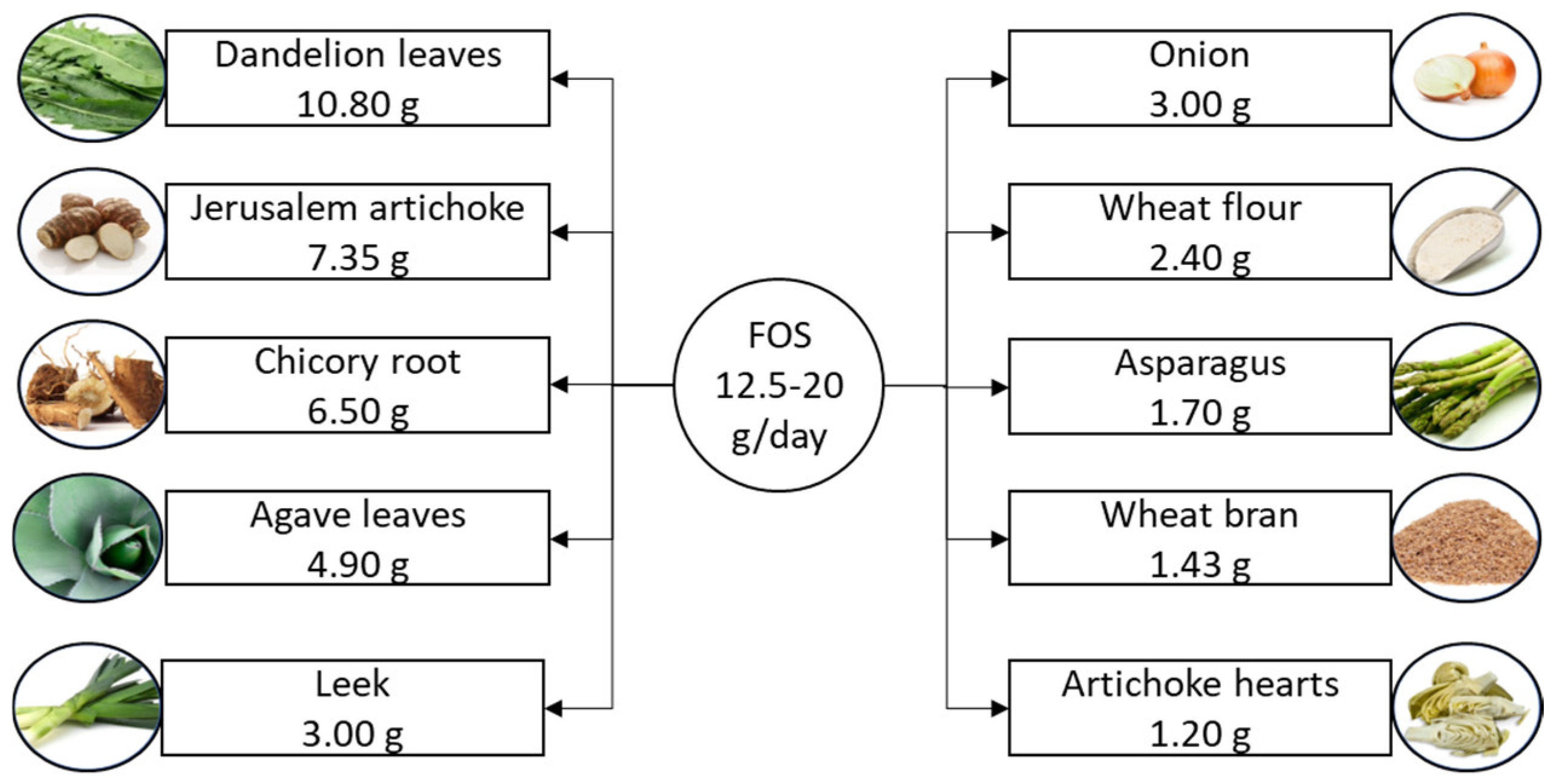
2.3. Fructooligosaccharides
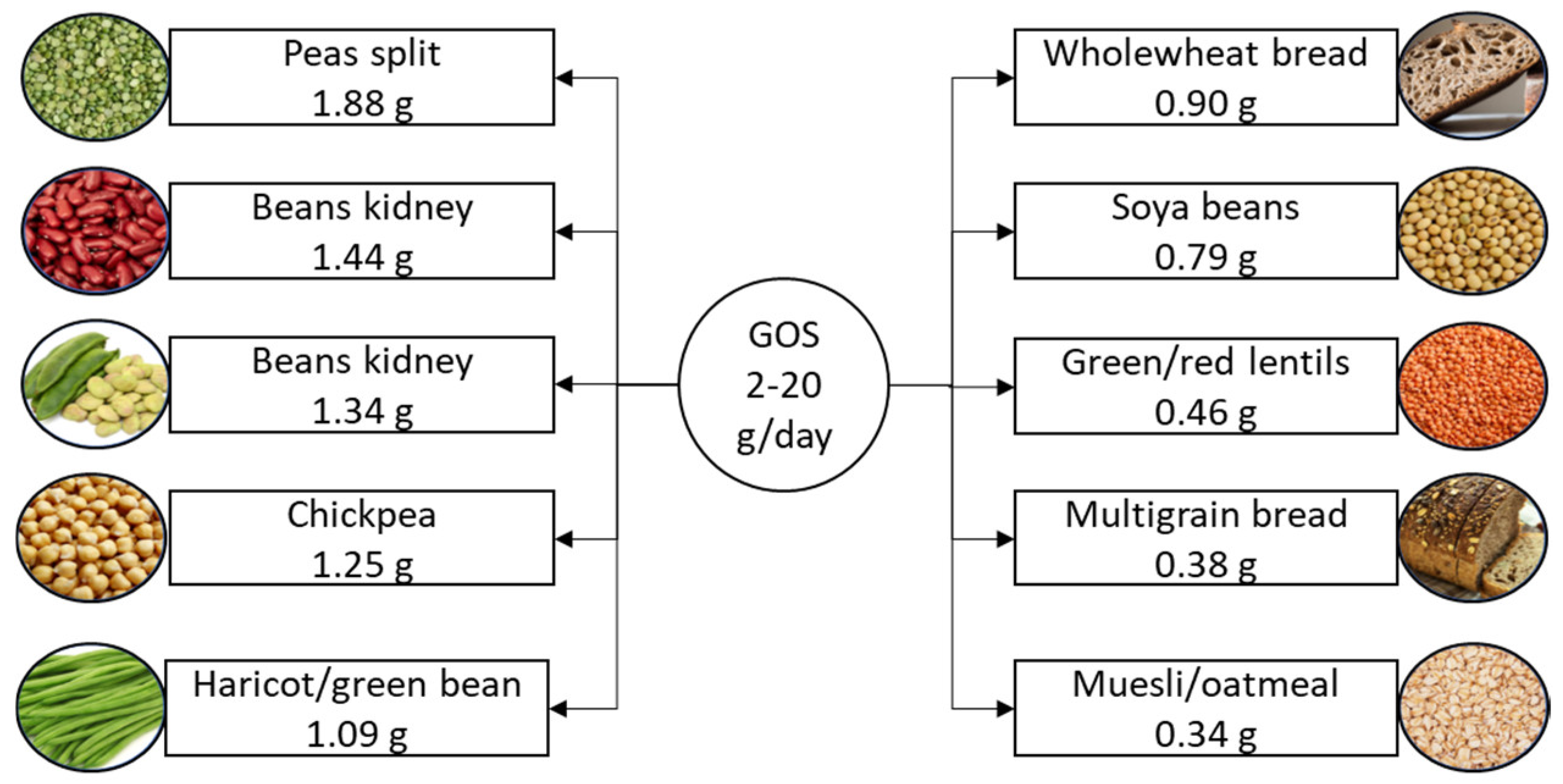
2.4. Galactooligosaccharides
2.5. Pectic Oligosaccharides
2.6. Polyphenols
2.7. β-Glucans
2.8. Dendrobium officinale
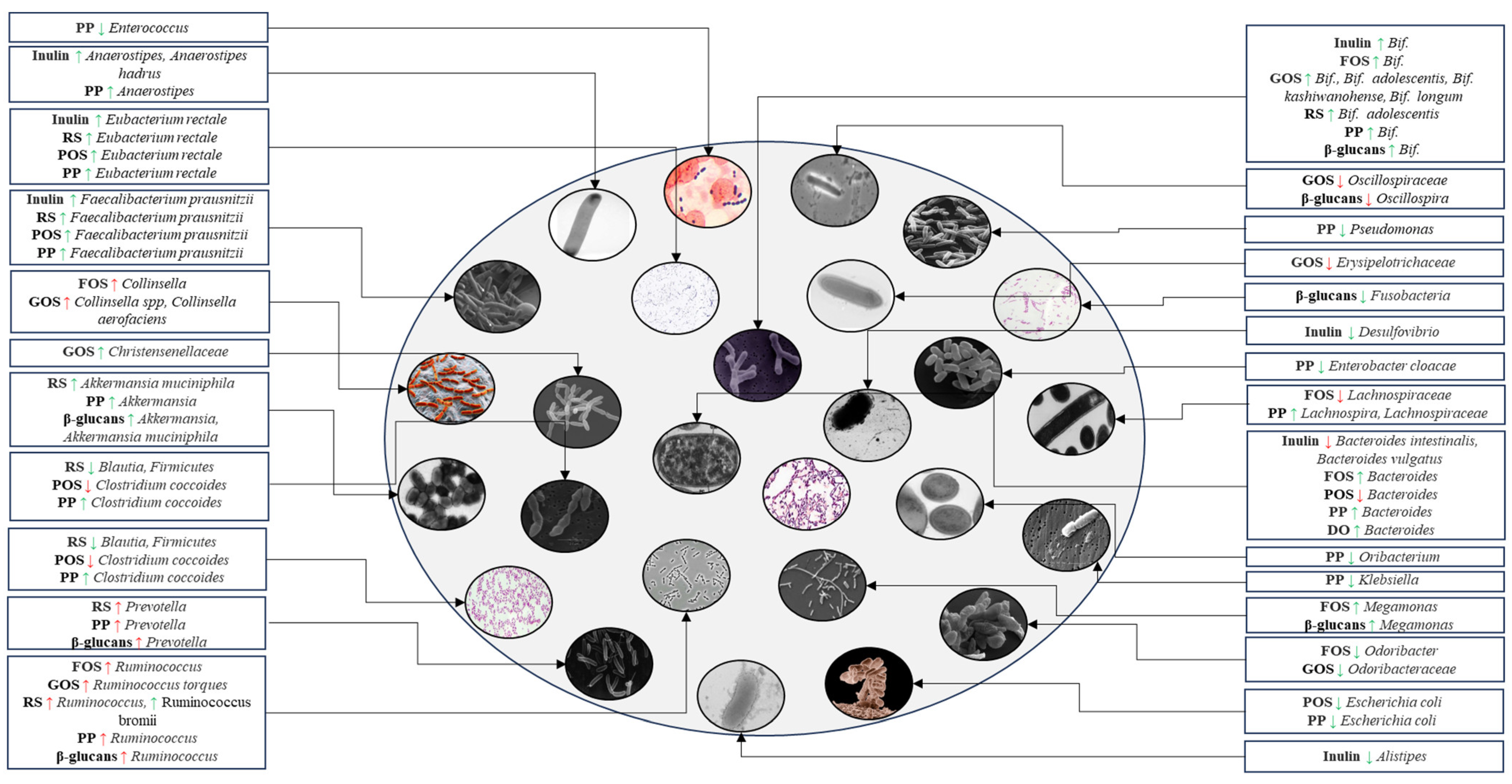
3. Changes in Gut Microbiota in T2DM
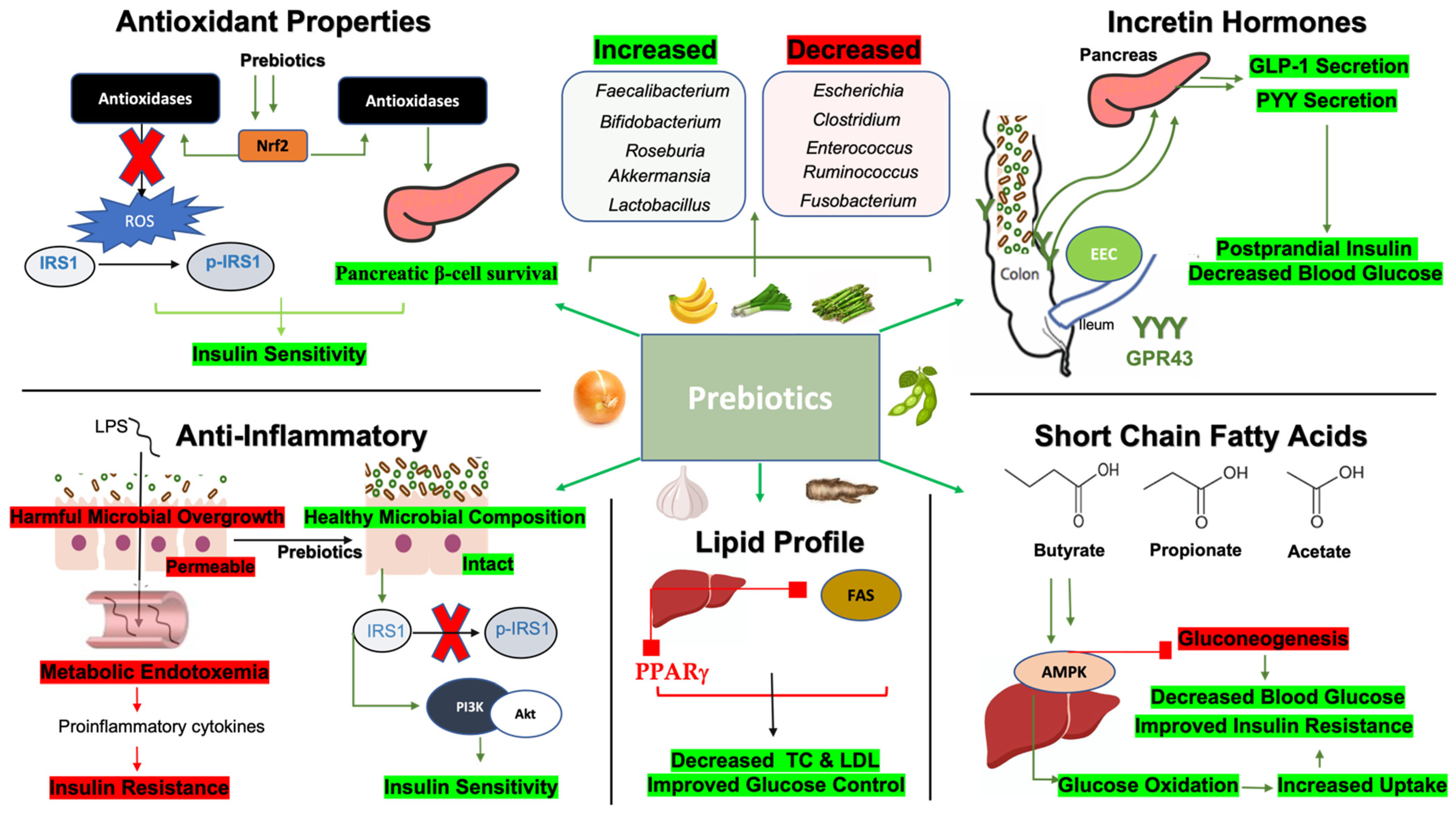
4. Mechanisms by Which Prebiotics Improve Glycemic Indices
4.1. Prebiotics, Short-Chain Fatty Acids (SCFAs), and Glycemic Indices
4.2. Prebiotics, Anti-Inflammatory Properties, and Glycemic Indices
4.3. Prebiotics and Incretin Hormones
4.4. Prebiotics, Lipid Metabolism, and Glycemic Indices
4.5. Prebiotics, Antioxidants, and Glycemic Indices
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lozupone, C.; Stombaugh, J.; Gordon, J.I.; Jansson, J.; Knight, R. Diversity, Stability and Resilience of the Human Gut Microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zhao, J. A Review on the Health Effects of Pesticides Based on Host Gut Microbiome and Metabolomics. Front. Mol. Biosci. 2021, 8, 632955. [Google Scholar] [CrossRef] [PubMed]
- Palmnäs-Bédard, M.; Costabile, G.; Vetrani, C.; Åberg, S.; Hjalmarsson, Y.; Dicksved, J.; Riccardi, G.; Landberg, R. The Human Gut Microbiota and Glucose Metabolism: A Scoping Review of Key Bacteria and the Potential Role of SCFAs. Am. J. Clin. Nutr. 2022, 116, 862–874. [Google Scholar] [CrossRef]
- Keivanlou, M.H.; Amini-Salehi, E.; Sattari, N.; Hashemi, M.; Saberian, P.; Prabhu, S.V.; Javid, M.; Mirdamadi, A.; Heidarzad, F.; Bakhshi, A.; et al. Gut microbiota interventions in type 2 diabetes mellitus: An umbrella review of glycemic indices. Diabetes Metab. Syndr. 2024, 18, 103110. [Google Scholar] [CrossRef] [PubMed]
- Krumbeck, J.A.; Maldonado-Gómez, M.X.; Martínez, I.; Frese, S.A.; Burkey, T.E.; Rasineni, K.; Ramer-Tait, A.E.; Harris, E.N.; Hutkins, R.W.; Walter, J. In Vivo Selection to Identify Bacterial Strains with Enhanced Ecological Performance in Synbiotic Applications. Appl. Environ. Microbiol. 2015, 81, 2455–2465. [Google Scholar] [CrossRef]
- Gibson, G.R.; Probert, H.M.; Loo, J.V.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Prebiotics and Probiotics in Digestive Health. Clin. Gastroenterol. Hepatol. 2019, 17, 333–344. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Ma, Y.; Yan, B.; Pei, W.; Wu, Q.; Ding, C.; Huang, C. The promotion mechanism of prebiotics for probiotics: A review. Front. Nutr. 2022, 9, 1000517. [Google Scholar] [CrossRef]
- Hamamah, S.; Iatcu, O.C.; Covasa, M. Nutrition at the Intersection between Gut Microbiota Eubiosis and Effective Management of Type 2 Diabetes. Nutrients 2024, 16, 269. [Google Scholar] [CrossRef]
- Akhter, N.; Wu, B.; Memon, A.M.; Mohsin, M. Probiotics and prebiotics associated with aquaculture: A review. Fish Shellfish. Immunol. 2015, 45, 733–741. [Google Scholar] [CrossRef]
- Megur, A.; Daliri, E.B.; Baltriukiene, D.; Burokas, A. Prebiotics as a Tool for the Prevention and Treatment of Obesity and Diabetes: Classification and Ability to Modulate the Gut Microbiota. Int. J. Mol. Sci. 2022, 23, 6097. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, K.; Fan, H.; Wei, M.; Xiong, Q. Targeting the gut microbiota and its metabolites for type 2 diabetes mellitus. Front. Endocrinol. 2023, 14, 1114424. [Google Scholar] [CrossRef] [PubMed]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef]
- Neyrinck, A.M.; Nazare, J.A.; Rodriguez, J.; Jottard, R.; Dib, S.; Sothier, M.; Berghe, L.V.D.; Alligier, M.; Alexiou, H.; Maquet, V.; et al. Development of a Repertoire and a Food Frequency Questionnaire for Estimating Dietary Fiber Intake Considering Prebiotics: Input from the FiberTAG Project. Nutrients 2020, 12, 2824. [Google Scholar] [CrossRef]
- Van de Wiele, T.; Boon, N.; Possemiers, S.; Jacobs, H.; Verstraete, W. Inulin-Type Fructans of Longer Degree of Polymerization Exert More Pronounced in vitro Prebiotic Effects. J. Appl. Microbiol. 2007, 102, 452–460. [Google Scholar] [CrossRef]
- Morris, C.; Morris, G.A. The Effect of Inulin and Fructo-Oligosaccharide Supplementation on the Textural, Rheological and Sensory Properties of Bread and Their Role in Weight Management: A Review. Food Chem. 2012, 133, 237–248. [Google Scholar] [CrossRef]
- Esmaeilnejad Moghadam, B.; Keivaninahr, F.; Fouladi, M.; Mokarram, R.R.; Nazemi, A. Inulin Addition to Yoghurt: Prebiotic Activity, Health Effects and Sensory Properties. Int. J. Dairy Technol. 2019, 72, 183–198. [Google Scholar] [CrossRef]
- Yousefi, M.; Khorshidian, N.; Hosseini, H. An Overview of the Functionality of Inulin in Meat and Poultry Products. Nutr. Food Sci. 2018, 48, 819–835. [Google Scholar] [CrossRef]
- Krivorotova, T.; Sereikaitė, J. Correlation between Fructan Exohydrolase Activity and the Quality of Helianthus tuberosus L. Tubers. Agronomy 2018, 8, 184. [Google Scholar] [CrossRef]
- Öztürk, B. A Rising Star Prebiotic Dietary Fiber: Inulin and Recent Applications in Meat Products. J. Food Health Sci. 2016, 3, 12–20. [Google Scholar] [CrossRef]
- Puchkova, T.S.; Pikhalo, D.M.; Karasyova, O.M. About the Universal Technology of Processing Jerusalem Artichoke and Chicory for Inulin. Food Syst. 2019, 2, 36–43. [Google Scholar] [CrossRef]
- Gargari, B.P.; Dehghan, P.; Aliasgharzadeh, A.; Jafar-Abadi, M.A. Effects of high performance inulin supplementation on glycemic control and antioxidant status in women with type 2 diabetes. Diabetes Metab. J. 2013, 37, 140–148. [Google Scholar] [CrossRef]
- Dehghan, P.; Gargari, B.P.; Jafar-Abadi, M.A. Oligofructose-enriched inulin improves some inflammatory markers and metabolic endotoxemia in women with type 2 diabetes mellitus: A randomized controlled clinical trial. Nutrition 2014, 30, 418–423. [Google Scholar] [CrossRef]
- Guess, N.D.; Dornhorst, A.; Oliver, N.; Frost, G.S. A Randomised Crossover Trial: The Effect of Inulin on Glucose Homeostasis in Subtypes of Prediabetes. Ann. Nutr. Metab. 2016, 68, 26–34. [Google Scholar] [CrossRef]
- Wang, X.; Wang, T.; Zhang, Q.; Xu, L.; Xiao, X. Dietary Supplementation with Inulin Modulates the Gut Microbiota and Improves Insulin Sensitivity in Prediabetes. Int. J. Endocrinol. 2021, 2021, 5579369. [Google Scholar] [CrossRef] [PubMed]
- Bonsu, N.K.A.; Johnson, S. Effects of inulin fibre Supplementation on Serum Glucose and Lipid Concentration in Patients with Type 2 Diabetes. Int. J. Diabetes Metab. 2019, 20, 80–86. [Google Scholar] [CrossRef]
- Holscher, H.D.; Bauer, L.L.; Gourineni, V.; Pelkman, C.L.; Fahey, G.C., Jr.; Swanson, K.S. Agave Inulin Supplementation Affects the Fecal Microbiota of Healthy Adults Participating in a Randomized, Double-Blind, Placebo-Controlled, Crossover Trial1, 2, 3. J. Nutr. 2015, 145, 2025–2032. [Google Scholar] [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of Human Gut Microbiota and Short-Chain Fatty Acids in Response to Dietary Interventions with Three Fermentable Fibers. mBio 2019, 10, e02566-18. [Google Scholar] [CrossRef]
- Dewulf, E.M.; Cani, P.D.; Claus, S.P.; Fuentes, S.; Puylaert, P.G.; Neyrinck, A.M.; Bindels, L.B.; de Vos, W.M.; Gibson, G.R.; Thissen, J.P.; et al. Insight into the prebiotic concept: Lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 2013, 62, 1112–1121. [Google Scholar] [CrossRef]
- Birkeland, E.; Gharagozlian, S.; Birkeland, K.I.; Valeur, J.; Mage, I.; Rud, I.; Aas, A.M. Prebiotic effect of inulin-type fructans on faecal microbiota and short-chain fatty acids in type 2 diabetes: A randomised controlled trial. Eur. J. Nutr. 2020, 59, 3325–3338. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Ji, G.; Zhang, L. Immunomodulatory effects of inulin and its intestinal metabolites. Front. Immunol. 2023, 14, 1224092. [Google Scholar] [CrossRef]
- Li, M.Y.; Duan, J.Q.; Wang, X.H.; Liu, M.; Yang, Q.Y.; Li, Y.; Cheng, K.; Liu, H.Q.; Wang, F. Inulin Inhibits the Inflammatory Response through Modulating Enteric Glial Cell Function in Type 2 Diabetic Mellitus Mice by Reshaping Intestinal Flora. ACS Omega 2023, 8, 36729–36743. [Google Scholar] [CrossRef]
- Li, K.; Zhang, L.; Xue, J.; Yang, X.; Dong, X.; Sha, L.; Lei, H.; Zhang, X.; Zhu, L.; Wang, Z.; et al. Dietary inulin alleviates diverse stages of type 2 diabetes mellitus via anti-inflammation and modulating gut microbiota in db/db mice. Food Funct. 2019, 10, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Roshanravan, N.; Alamdari, N.M.; Jafarabadi, M.A.; Mohammadi, A.; Shabestari, B.R.; Nasirzadeh, N.; Asghari, S.; Mansoori, B.; Akbarzadeh, M.; Ghavami, A.; et al. Effects of oral butyrate and inulin supplementation on inflammation-induced pyroptosis pathway in type 2 diabetes: A randomized, double-blind, placebo-controlled trial. Cytokine 2020, 131, 155101. [Google Scholar] [CrossRef]
- Martinez-Ramirez, O.C.; Salazar-Pina, A.; Ceron-Ramirez, X.; Rubio-Lightbourn, J.; Torres-Romero, F.; Casas-Avila, L.; Castro-Hernandez, C. Effect of Inulin Intervention on Metabolic Control and Methylation of INS and IRS1 Genes in Patients with Type 2 Diabetes Mellitus. Nutrients 2022, 14, 5195. [Google Scholar] [CrossRef]
- Ning, C.; Wang, X.; Gao, S.; Mu, J.; Wang, Y.; Liu, S.; Zhu, J.; Meng, X. Chicory inulin ameliorates type 2 diabetes mellitus and suppresses JNK and MAPK pathways in vivo and in vitro. Mol. Nutr. Food Res. 2017, 61, 1600673. [Google Scholar] [CrossRef]
- Lewandowski, L.; Urbanowicz, I.; Kepinska, M.; Milnerowicz, H. Concentration/activity of superoxide dismutase isozymes and the pro-/antioxidative status, in context of type 2 diabetes and selected single nucleotide polymorphisms (genes: INS, SOD1, SOD2, SOD3)—Preliminary findings. Biomed. Pharmacother. 2021, 137, 111396. [Google Scholar] [CrossRef]
- Sidarala, V.; Kowluru, A. The Regulatory Roles of Mitogen-Activated Protein Kinase (MAPK) Pathways in Health and Diabetes: Lessons Learned from the Pancreatic beta-Cell. Recent Pat. Endocr. Metab. Immune Drug Discov. 2017, 10, 76–84. [Google Scholar] [CrossRef]
- Teichmann, J.; Cockburn, D.W. In Vitro Fermentation Reveals Changes in Butyrate Production Dependent on Resistant Starch Source and Microbiome Composition. Front. Microbiol. 2021, 12, 640253. [Google Scholar] [CrossRef]
- Fuentes-Zaragoza, E.; Riquelme-Navarrete, M.J.; Sánchez-Zapata, E.; Pérez-Álvarez, J.A. Resistant Starch as Functional Ingredient: A Review. Food Res. Int. 2010, 43, 931–942. [Google Scholar] [CrossRef]
- Patterson, M.A.; Maiya, M.; Stewart, M.L. Resistant Starch Content in Foods Commonly Consumed in the United States: A Narrative Review. J. Acad. Nutr. Diet. 2020, 120, 230–244. [Google Scholar] [CrossRef]
- Thuy, N.M.; Too, B.C.; Vuong, K.M.; Lan, P.T.; Tuyen, P.T.; Tram, N.B.; Van Tai, N. Resistant starch in various starchy vegetables and the relationship with its physical and chemical characteristics. J. Appl. Biol. Biotechnol. 2022, 10, 181–188. [Google Scholar] [CrossRef]
- Pratiwi, M.; Faridah, D.N.; Lioe, H.N. Structural Changes to Starch after Acid Hydrolysis, Debranching, Autoclaving-cooling Cycles, and Heat Moisture Treatment (HMT): A Review. Starch-Stärke 2017, 70, 1700028. [Google Scholar] [CrossRef]
- Wang, W.; Shen, M.; Kang, Z.; Wang, J.; Quan, Z.; Ji, X.; Zhao, S.; Liu, D.; Cao, L. Effect of Natural Fermentation of Sorghum on Resistant Starch Molecular Structure and Fermentation Property. J. Chem. 2020, 2020, 9835214. [Google Scholar] [CrossRef]
- Malik, M.K.; Kumar, V.; Singh, J.; Bhatt, P.; Dixit, R.; Kumar, S. Phosphorylation of Alkali Extracted Mandua Starch by STPP/STMP for Improving Digestion Resistibility. ACS Omega 2023, 8, 11750–11767. [Google Scholar] [CrossRef]
- Alvarenga, I.C.; Keller, L.; Waldy, C.; Aldrich, C.G. Extrusion Processing Modifications of a Dog Kibble at Large Scale Alter Levels of Starch Available to Animal Enzymatic Digestion. Foods 2021, 10, 2526. [Google Scholar] [CrossRef]
- Eroğlu, E.İ.; Büyüktuncer, Z. The Effect of Various Cooking Methods on Resistant Starch Content of Foods. Nutr. Food Sci. 2017, 47, 522–533. [Google Scholar] [CrossRef]
- Kuttigounder, D.; Lingamallu, J.R.; Bhattacharya, S. Turmeric Powder and Starch: Selected Physical, Physicochemical, and Microstructural Properties. J. Food Sci. 2011, 76, C1284–C1291. [Google Scholar] [CrossRef]
- Oyeyinka, S.A.; Kayitesi, E.; Adebo, O.A.; Oyedeji, A.B.; Ogundele, O.M.; Obilana, A.O.; Njobeh, P.B. A Review on the Physicochemical Properties and Potential Food Applications of Cowpea (Vigna unguiculata) Starch. Int. J. Food Sci. Technol. 2020, 56, 52–60. [Google Scholar] [CrossRef]
- Mishra, S.; Awasthi, M. Role of Resistant Starch in Food and Healthcare Industry: A Review. Asian J. Dairy Food Res. 2024. [Google Scholar] [CrossRef]
- Haub, M.D.; Hubach, K.L.; Al-Tamimi, E.; Ornelas, S.; Seib, P.A. Different Types of Resistant Starch Elicit Different Glucose Reponses in Humans. J. Nutr. Metab. 2010, 2010, 230501. [Google Scholar] [CrossRef]
- Pugh, J.E.; Cai, M.; Altieri, N.; Frost, G. A comparison of the effects of resistant starch types on glycemic response in individuals with type 2 diabetes or prediabetes: A systematic review and meta-analysis. Front. Nutr. 2023, 10, 1118229. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Song, Y.-H.; Zhao, R.; Xia, L.; Chen, Y.; Cui, Y.-P.; Rao, Z.-Y.; Zhou, Y.; Zhuang, W.; et al. Effects of the resistant starch on glucose, insulin, insulin resistance, and lipid parameters in overweight or obese adults: A systematic review and meta-analysis. Nutr. Diabetes 2019, 9, 19. [Google Scholar] [CrossRef]
- Xiong, K.; Wang, J.; Kang, T.; Xu, F.; Ma, A. Effects of resistant starch on glycaemic control: A systematic review and meta-analysis. Br. J. Nutr. 2021, 125, 1260–1269. [Google Scholar] [CrossRef]
- Lu, J.; Ma, B.; Qiu, X.; Sun, Z.; Xiong, K. Effects of resistant starch supplementation on oxidative stress and inflammation biomarkers: A systematic review and meta-analysis of randomized controlled trials. Asia Pac. J. Clin. Nutr. 2021, 30, 614–623. [Google Scholar] [CrossRef]
- Bodinham, C.L.; Smith, L.; Thomas, E.L.; Bell, J.D.; Swann, J.R.; Costabile, A.; Russell-Jones, D.; Umpleby, A.M.; Robertson, M.D. Efficacy of increased resistant starch consumption in human type 2 diabetes. Endocr. Connect. 2014, 3, 75–84. [Google Scholar] [CrossRef]
- Gargari, B.P.; Namazi, N.; Khalili, M.; Sarmadi, B.; Jafarabadi, M.A.; Dehghan, P. Is there any place for resistant starch, as alimentary prebiotic, for patients with type 2 diabetes? Complement. Ther. Med. 2015, 23, 810–815. [Google Scholar] [CrossRef]
- Maier, T.V.; Lucio, M.; Lee, L.H.; VerBerkmoes, N.C.; Brislawn, C.J.; Bernhardt, J.; Lamendella, R.; McDermott, J.E.; Bergeron, N.; Heinzmann, S.S.; et al. Impact of Dietary Resistant Starch on the Human Gut Microbiome, Metaproteome, and Metabolome. mBio 2017, 8, e01343-17. [Google Scholar] [CrossRef]
- Ordiz, M.I.; May, T.D.; Mihindukulasuriya, K.; Martin, J.; Crowley, J.; Tarr, P.I.; Ryan, K.; Mortimer, E.; Gopalsamy, G.; Maleta, K.; et al. The effect of dietary resistant starch type 2 on the microbiota and markers of gut inflammation in rural Malawi children. Microbiome 2015, 3, 37. [Google Scholar] [CrossRef]
- Upadhyaya, B.; McCormack, L.; Fardin-Kia, A.R.; Juenemann, R.; Nichenametla, S.; Clapper, J.; Specker, B.; Dey, M. Impact of dietary resistant starch type 4 on human gut microbiota and immunometabolic functions. Sci. Rep. 2016, 6, 28797. [Google Scholar] [CrossRef]
- Martínez, I.; Kim, J.; Duffy, P.R.; Schlegel, V.L.; Walter, J. Resistant starches types 2 and 4 have differential effects on the composition of the fecal microbiota in human subjects. PLoS ONE 2010, 5, e15046. [Google Scholar] [CrossRef]
- Sabater-Molina, M.; Larqué, E.; Torrella, F.; Zamora, S. Dietary Fructooligosaccharides and Potential Benefits on Health. J. Physiol. Biochem. 2009, 65, 315–328. [Google Scholar] [CrossRef]
- Lachman, J.; Fernández, E.C.; Orsák, M. Yacon [Smallanthus Sonchifolia (Poepp. Et Endl.) H. Robinson] Chemical Composition and Use—A Review. Plant Soil Environ. 2003, 49, 283–290. [Google Scholar] [CrossRef]
- Mao, S.; Liu, Y.; Yang, J.; Ma, X.; Zeng, F.; Zhang, Z.; Wang, S.; Han, H.C.; Qin, H.; Lu, F. Cloning, Expression and Characterization of a Novel Fructosyltransferase from Aspergillus niger and Its Application in the Synthesis of Fructooligosaccharides. RSC Adv. 2019, 9, 23856–23863. [Google Scholar] [CrossRef]
- Todero, L.M.; Vargas-Rechia, C.G.; Guimarães, L.H.S. Production of Short-chain Fructooligosaccharides (scFOS) Using Extracellular β-D-fructofuranosidase Produced by Aspergillus thermomutatus. J. Food Biochem. 2019, 43, e12937. [Google Scholar] [CrossRef]
- Garleb, K.A.; Snowden, M.K.; Wolf, B.W.; Chow, J.W. Application of Fructooligosaccharides to Medical Foods as a Fermentable Dietary Fiber. Biosci. Microflora 2002, 21, 43–54. [Google Scholar] [CrossRef]
- Yamashita, K.; Kawai, K.; Itakura, M. Effects of fructo-oligosaccharides on blood glucose and serum lipids in diabetic subjects. Nutr. Res. 1984, 4, 961–966. [Google Scholar] [CrossRef]
- Alles, M.S.; de Roos, N.M.; Bakx, J.C.; van de Lisdonk, E.; Zock, P.L.; Hautvast, J.G. Consumption of fructooligosaccharides does not favorably affect blood glucose and serum lipid concentrations in patients with type 2 diabetes. Am. J. Clin. Nutr. 1999, 69, 64–69. [Google Scholar] [CrossRef]
- Luo, J.; Van Yperselle, M.; Rizkalla, S.W.; Rossi, F.; Slama, G.; Bornet, F.R. Chronic consumption of short-chain fructooligosaccharides does not affect basal hepatic glucose production or insulin resistance in type 2 diabetics. J. Nutr. 2000, 130, 1572–1577. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, A.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Red Raspberry and Fructo-Oligosaccharide Supplementation, Metabolic Biomarkers, and the Gut Microbiota in Adults with Prediabetes: A Randomized Crossover Clinical Trial. J. Nutr. 2022, 152, 1438–1449. [Google Scholar] [CrossRef]
- Daou, C.; Zhang, H. Oat Beta-Glucan: Its Role in Health Promotion and Prevention of Diseases. Compr. Rev. Food Sci. Food Saf. 2012, 11, 355–365. [Google Scholar] [CrossRef]
- Mahalak, K.K.; Firrman, J.; Narrowe, A.B.; Hu, W.; Jones, S.M.; Bittinger, K.; Moustafa, A.M.; Liu, L. Fructooligosaccharides (FOS) differentially modifies the in vitro gut microbiota in an age-dependent manner. Front. Nutr. 2023, 9, 1058910. [Google Scholar] [CrossRef]
- Wongkrasant, P.; Pongkorpsakol, P.; Chitwattananont, S.; Satianrapapong, W.; Tuangkijkul, N.; Muanprasat, C. Fructo-oligosaccharides alleviate inflammation-associated apoptosis of GLP-1 secreting L cells via inhibition of iNOS and cleaved caspase-3 expression. J. Pharmacol. Sci. 2020, 143, 65–73. [Google Scholar] [CrossRef]
- Martinez-Saez, N.; Hochkogler, C.M.; Somoza, V.; Del Castillo, M.D. Biscuits with No Added Sugar Containing Stevia, Coffee Fibre and Fructooligosaccharides Modifies alpha-Glucosidase Activity and the Release of GLP-1 from HuTu-80 Cells and Serotonin from Caco-2 Cells after In Vitro Digestion. Nutrients 2017, 9, 694. [Google Scholar] [CrossRef]
- da Silva, M.V.T.; Nunes, S.S.; Costa, W.C.; Sanches, S.M.D.; Silveira, A.L.M.; Ferreira, A.R.S.; Filha, R.S.; Correia, M.; Mota, L.D.G.; Generoso, S.V. Acute intake of fructooligosaccharide and partially hydrolyzed guar gum on gastrointestinal transit: A randomized crossover clinical trial. Nutrition 2022, 102, 111737. [Google Scholar] [CrossRef]
- Ikeda, H.; Shibao, K.; Yamaguchi, R.; Yoh, M.; Shimamatsu, M.; Hiyamuta, K.; Itaya, K.; Ohkita, Y.; Sugi, K.; Koga, Y.; et al. Quantitative evaluation of regional myocardial blood flow by digital subtraction angiography: Correlations with exercise electrocardiography and Tl-201 myocardial scintigraphy. J. Cardiography Suppl. 1987, 12, 81–89. [Google Scholar]
- Tian, S.; Wang, J.; Yu, H.; Wang, J.; Zhu, W. Changes in Ileal Microbial Composition and Microbial Metabolism by an Early-Life Galacto-Oligosaccharides Intervention in a Neonatal Porcine Model. Nutrients 2019, 11, 1753. [Google Scholar] [CrossRef]
- Tanabe, K.; Nakamura, S.; Omagari, K.; Oku, T. Determination Trial of Nondigestible Oligosaccharide in Processed Foods by Improved AOAC Method 2009.01 Using Porcine Small Intestinal Enzyme. J. Agric. Food Chem. 2015, 63, 5747–5752. [Google Scholar] [CrossRef]
- Satar, R.; Jafri, M.A.; Rasool, M.; Ansari, S.A. Role of Glutaraldehyde in Imparting Stability to Immobilized Β-Galactosidase Systems. Braz. Arch. Biol. Technol. 2018, 60. [Google Scholar] [CrossRef]
- Albrecht, S.; Schols, H.A.; Klarenbeek, B.; Voragen, A.G.J.; Gruppen, H. Introducing Capillary Electrophoresis With Laser-Induced Fluorescence (CE−LIF) as a Potential Analysis and Quantification Tool for Galactooligosaccharides Extracted From Complex Food Matrices. J. Agric. Food Chem. 2010, 58, 2787–2794. [Google Scholar] [CrossRef]
- Vulevic, J.; Juric, A.; Tzortzis, G.; Gibson, G.R. A mixture of trans-galactooligosaccharides reduces markers of metabolic syndrome and modulates the fecal microbiota and immune function of overweight adults. J. Nutr. 2013, 143, 324–331. [Google Scholar] [CrossRef]
- Pedersen, C.; Gallagher, E.; Horton, F.; Ellis, R.J.; Ijaz, U.Z.; Wu, H.; Jaiyeola, E.; Diribe, O.; Duparc, T.; Cani, P.D.; et al. Host-microbiome interactions in human type 2 diabetes following prebiotic fibre (galacto-oligosaccharide) intake. Br. J. Nutr. 2016, 116, 1869–1877. [Google Scholar] [CrossRef]
- Canfora, E.E.; van der Beek, C.M.; Hermes, G.D.A.; Goossens, G.H.; Jocken, J.W.E.; Holst, J.J.; van Eijk, H.M.; Venema, K.; Smidt, H.; Zoetendal, E.G.; et al. Supplementation of Diet with Galacto-oligosaccharides Increases Bifidobacteria, but Not Insulin Sensitivity, in Obese Prediabetic Individuals. Gastroenterology 2017, 153, 87–97.e83. [Google Scholar] [CrossRef]
- Gopalakrishnan, A.; Clinthorne, J.F.; Rondini, E.A.; McCaskey, S.J.; Gurzell, E.A.; Langohr, I.M.; Gardner, E.M.; Fenton, J.I. Supplementation with galacto-oligosaccharides increases the percentage of NK cells and reduces colitis severity in Smad3-deficient mice. J. Nutr. 2012, 142, 1336–1342. [Google Scholar] [CrossRef]
- Gonai, M.; Shigehisa, A.; Kigawa, I.; Kurasaki, K.; Chonan, O.; Matsuki, T.; Yoshida, Y.; Aida, M.; Hamano, K.; Terauchi, Y. Galacto-oligosaccharides ameliorate dysbiotic Bifidobacteriaceae decline in Japanese patients with type 2 diabetes. Benef. Microbes 2017, 8, 705–716. [Google Scholar] [CrossRef]
- Beteri, B.; Barone, M.; Turroni, S.; Brigidi, P.; Tzortzis, G.; Vulevic, J.; Sekulic, K.; Motei, D.E.; Costabile, A. Impact of Combined Prebiotic Galacto-Oligosaccharides and Bifidobacterium breve-Derived Postbiotic on Gut Microbiota and HbA1c in Prediabetic Adults: A Double-Blind, Randomized, Placebo-Controlled Study. Nutrients 2024, 16, 2205. [Google Scholar] [CrossRef]
- Wilson, B.; Eyice, Ö.; Koumoutsos, I.; Lomer, M.C.; Irving, P.M.; Lindsay, J.O.; Whelan, K. Prebiotic Galactooligosaccharide Supplementation in Adults with Ulcerative Colitis: Exploring the Impact on Peripheral Blood Gene Expression, Gut Microbiota, and Clinical Symptoms. Nutrients 2021, 13, 3598. [Google Scholar] [CrossRef]
- Marzorati, M.; Ghyselinck, J.; Van den Abbeele, P.; Maruszak, A.; Harthoorn, L. Galactooligosaccharide (GOS) Reduces Branched Short-Chain Fatty Acids, Ammonium, and pH in a Short-Term Colonic Fermentation Model. Appl. Microbiol. 2023, 3, 90–103. [Google Scholar] [CrossRef]
- Tingirikari, J.M.R. Microbiota-accessible pectic poly- and oligosaccharides in gut health. Food Funct. 2018, 9, 5059–5073. [Google Scholar] [CrossRef]
- Bai, Y.; Gilbert, R.G. Mechanistic Understanding of the Effects of Pectin on In Vivo Starch Digestion: A Review. Nutrients 2022, 14, 5107. [Google Scholar] [CrossRef]
- Babbar, N.; Dejonghe, W.; Gatti, M.; Sforza, S.; Elst, K. Pectic oligosaccharides from agricultural by-products: Production, characterization and health benefits. Crit. Rev. Biotechnol. 2016, 36, 594–606. [Google Scholar] [CrossRef]
- Gullón, B.; Gómez, B.; Martínez-Sabajanes, M.; Yáñez, R.; Parajó, J.; Alonso, J. Pectic oligosaccharides: Manufacture and functional properties. Trends Food Sci. Technol. 2013, 30, 153–161. [Google Scholar] [CrossRef]
- Brouns, F.; Theuwissen, E.; Adam, A.; Bell, M.; Berger, A.; Mensink, R.P. Cholesterol-lowering properties of different pectin types in mildly hyper-cholesterolemic men and women. Eur. J. Clin. Nutr. 2012, 66, 591–599. [Google Scholar] [CrossRef]
- Orqueda, M.E.; Zampini, I.C.; Torres, S.; Isla, M.I. Functional Characterization and Toxicity of Pectin from Red Chilto Fruit Waste (Peels). Plants 2023, 12, 2603. [Google Scholar] [CrossRef]
- Jenkins, D.A.; Leeds, A.; Wolever, T.S.; Goff, D.; George, K.; Alberti, M.M.; Gassull, M.; Derek, T.; Hockaday, R. Unabsorbable carbohydrates and diabetes: Decreased post-prandial hyperglycaemia. Lancet 1976, 2, 172–174. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Leeds, A.R.; Gassull, M.A.; Cochet, B.; Alberti, G.M. Decrease in postprandial insulin and glucose concentrations by guar and pectin. Ann. Intern. Med. 1977, 86, 20–23. [Google Scholar] [CrossRef]
- do Socoro Ramos de Queiroz, M.; Idalino Janebro, D.; Auxiliadora Lins da Cunha, M.; dos Santos Medeiros, J.; Sabaa-Srur, A.U.; de Fatima FM Diniz, M.; dos Santos, S.C. Effect of the yellow passion fruit peel flour (Passiflora edulis f. flavicarpa deg.) in insulin sensitivity in type 2 diabetes mellitus patients. Nutr. J. 2012, 11, 89. [Google Scholar] [CrossRef]
- Jones, M.; Gu, X.; Stebbins, N.B.; Crandall, P.G.; Ricke, S.C.; Lee, S.O. Effects of Soybean Pectin on Blood Glucose and Insulin Responses in Healthy Men. FASEB J. 2015, 29. [Google Scholar] [CrossRef]
- Pollini, L.; Blasi, F.; Ianni, F.; Grispoldi, L.; Moretti, S.; Veroli, A.D.; Cossignani, L.; Cenci-Goga, B.T. Ultrasound-Assisted Extraction and Characterization of Polyphenols From Apple Pomace, Functional Ingredients for Beef Burger Fortification. Molecules 2022, 27, 1933. [Google Scholar] [CrossRef]
- Li, T.; Li, S.; Du, L.; Wang, N.; Guo, M.; Zhang, J.; Yan, F.; Zhang, H. Effects of haw pectic oligosaccharide on lipid metabolism and oxidative stress in experimental hyperlipidemia mice induced by high-fat diet. Food Chem. 2010, 121, 1010–1013. [Google Scholar] [CrossRef]
- Segar, H.M.; Gani, S.A.; Khayat, M.E.; Rahim, M.B.H.A. Antioxidant and Antidiabetic Properties of Pectin Extracted from Pomegranate (Punica granatum) Peel. J. Biochem. Microbiol. Biotechnol. 2023, 11, 35–40. [Google Scholar] [CrossRef]
- Nie, Q.; Hu, J.; He, G.; Li, M.; Sun, Y.; Chen, H.; Zuo, S.; Fang, Q.; Huang, X.J.; Yin, J.; et al. Bioactive Dietary Fibers Selectively Promote Gut Microbiota to Exert Antidiabetic Effects. J. Agric. Food Chem. 2021, 69, 7000–7015. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, P.; Zhang, H. Pectin in cancer therapy: A review. Trends Food Sci. Technol. 2015, 44, 258–271. [Google Scholar] [CrossRef]
- Gómez, B.; Gullón, B.; Yáñez, R.; Schols, H.; Alonso, J.L. Prebiotic potential of pectins and pectic oligosaccharides derived from lemon peel wastes and sugar beet pulp: A comparative evaluation. J. Funct. Foods 2016, 20, 108–121. [Google Scholar] [CrossRef]
- Leijdekkers, A.G.; Aguirre, M.; Venema, K.; Bosch, G.; Gruppen, H.; Schols, H.A. In Vitro fermentability of sugar beet pulp derived oligosaccharides using human and pig fecal inocula. J. Agric. Food Chem. 2014, 62, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Olano-Martin, E.; Rimbach, G.H.; Gibson, G.R.; Rastall, R.A. Pectin and pectic-oligosaccharides induce apoptosis in in vitro human colonic adenocarcinoma cells. Anticancer Res. 2003, 23, 341–346. [Google Scholar]
- Kastner, U.; Glasl, S.; Follrich, B.; Guggenbichler, J.P.; Jurenitsch, J. Acid oligosaccharides as the active principle of aqueous carrot extracts for prevention and therapy of gastrointestinal infections. Wien. Med. Wochenschr. 2002, 152, 379–381. [Google Scholar] [CrossRef]
- Olano-Martin, E.; Gibson, G.R.; Rastell, R.A. Comparison of the in vitro bifidogenic properties of pectins and pectic-oligosaccharides. J. Appl. Microbiol. 2002, 93, 505–511. [Google Scholar] [CrossRef]
- Mandalari, G.; Nueno Palop, C.; Tuohy, K.; Gibson, G.R.; Bennett, R.N.; Waldron, K.W.; Bisignano, G.; Narbad, A.; Faulds, C.B. In vitro evaluation of the prebiotic activity of a pectic oligosaccharide-rich extract enzymatically derived from bergamot peel. Appl. Microbiol. Biotechnol. 2007, 73, 1173–1179. [Google Scholar] [CrossRef]
- Gullón, B.; Gullón, P.; Sanz, Y.; Alonso, J.; Parajó, J.C. Prebiotic potential of a refined product containing pectic oligosaccharides. LWT—Food Sci. Technol. 2011, 44, 1687–1696. [Google Scholar] [CrossRef]
- Chen, J.; Liang, R.H.; Liu, W.; Li, T.; Liu, C.M.; Wu, S.S.; Wang, Z.J. Pectic-oligosaccharides prepared by dynamic high-pressure microfluidization and their in vitro fermentation properties. Carbohydr. Polym. 2013, 91, 175–182. [Google Scholar] [CrossRef]
- Manderson, K.; Pinart, M.; Tuohy, K.M.; Grace, W.E.; Hotchkiss, A.T.; Widmer, W.; Yadhav, M.P.; Gibson, G.R.; Rastall, R.A. In vitro determination of prebiotic properties of oligosaccharides derived from an orange juice manufacturing by-product stream. Appl. Environ. Microbiol. 2005, 71, 8383–8389. [Google Scholar] [CrossRef]
- Fanaro, S.; Jelinek, J.; Stahl, B.; Boehm, G.; Kock, R.; Vigi, V. Acidic oligosaccharides from pectin hydrolysate as new component for infant formulae: Effect on intestinal flora, stool characteristics, and pH. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 186–190. [Google Scholar] [CrossRef]
- Magne, F.; Hachelaf, W.; Suau, A.; Boudraa, G.; Bouziane-Nedjadi, K.; Rigottier-Gois, L.; Touhami, M.; Desjeux, J.F.; Pochart, P. Effects on faecal microbiota of dietary and acidic oligosaccharides in children during partial formula feeding. J. Pediatr. Gastroenterol. Nutr. 2008, 46, 580–588. [Google Scholar] [CrossRef]
- Jiang, L.-L.; Gong, X.; Ji, M.-Y.; Wang, C.-C.; Wang, J.-H.; Li, M.-H. Bioactive compounds from plant-based functional foods: A promising choice for the prevention and management of hyperuricemia. Foods 2020, 9, 973. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Sarkar, D.; Christopher, A.; Shetty, K. Phenolic Bioactives From Plant-Based Foods for Glycemic Control. Front. Endocrinol. 2022, 12, 727503. [Google Scholar] [CrossRef] [PubMed]
- Wisnuwardani, R.W.; De Henauw, S.; Androutsos, O.; Forsner, M.; Gottrand, F.; Huybrechts, I.; Knaze, V.; Kersting, M.; Le Donne, C.; Marcos, A.; et al. Estimated dietary intake of polyphenols in European adolescents: The HELENA study. Eur. J. Nutr. 2019, 58, 2345–2363. [Google Scholar] [CrossRef]
- Costa, C.; Tsatsakis, A.; Mamoulakis, C.; Teodoro, M.; Briguglio, G.; Caruso, E.; Tsoukalas, D.; Margina, D.; Dardiotis, E.; Kouretas, D.; et al. Current evidence on the effect of dietary polyphenols intake on chronic diseases. Food Chem. Toxicol. 2017, 110, 286–299. [Google Scholar] [CrossRef]
- Couto, A.F.; Estevinho, B.N. Valorization of Agricultural By-Products (Fragaria vesca) through the Production of Value-Added Micro/Nanostructures Using Electrohydrodynamic Techniques. Foods 2024, 13, 1162. [Google Scholar] [CrossRef] [PubMed]
- Kooti, W.; Farokhipour, M.; Asadzadeh, Z.; Ashtary-Larky, D.; Asadi-Samani, M. The role of medicinal plants in the treatment of diabetes: A systematic review. Electron. Physician 2016, 8, 1832. [Google Scholar] [CrossRef] [PubMed]
- Gowd, V.; Xie, L.; Sun, C.; Chen, W. Phenolic profile of bayberry followed by simulated gastrointestinal digestion and gut microbiota fermentation and its antioxidant potential in HepG2 cells. J. Funct. Foods 2020, 70, 103987. [Google Scholar] [CrossRef]
- Li, D.; Yang, Y.; Sun, L.; Chen, L.; Zhao, P.; Wang, Z.-C.; Guo, Y. Effect of young apple (Malus domestica Borkh. cv. Red Fuji) polyphenols on alleviating insulin resistance. Food Biosci. 2020, 36, 100637. [Google Scholar] [CrossRef]
- Gutiérrez-Grijalva, E.P.; Antunes-Ricardo, M.; Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Heredia, J.B. Cellular Antioxidant Activity and in vitro inhibition of α-glucosidase, α-amylase and pancreatic lipase of oregano polyphenols under simulated gastrointestinal digestion. Food Res. Int. 2019, 116, 676–686. [Google Scholar] [CrossRef]
- Gowd, V.; Bao, T.; Wang, L.; Huang, Y.; Chen, S.; Zheng, X.; Cui, S.; Chen, W. Antioxidant and antidiabetic activity of blackberry after gastrointestinal digestion and human gut microbiota fermentation. Food Chem. 2018, 269, 618–627. [Google Scholar] [CrossRef]
- Li, F.; Zhang, B.; Chen, G.; Fu, X. The novel contributors of anti-diabetic potential in mulberry polyphenols revealed by UHPLC-HR-ESI-TOF-MS/MS. Food Res. Int. 2017, 100, 873–884. [Google Scholar] [CrossRef]
- Obaroakpo, J.U.; Liu, L.; Zhang, S.; Lu, J.; Liu, L.; Pang, X.; Lv, J. In Vitro modulation of glucagon-like peptide release by DPP-IV inhibitory polyphenol-polysaccharide conjugates of sprouted quinoa yoghurt. Food Chem. 2020, 324, 126857. [Google Scholar] [CrossRef]
- Elsayed, R.H.; Kamel, E.M.; Mahmoud, A.M.; El-Bassuony, A.A.; Bin-Jumah, M.; Lamsabhi, A.M.; Ahmed, S.A. Rumex dentatus L. phenolics ameliorate hyperglycemia by modulating hepatic key enzymes of carbohydrate metabolism, oxidative stress and PPARγ in diabetic rats. Food Chem. Toxicol. 2020, 138, 111202. [Google Scholar] [CrossRef]
- Li, H.; Park, H.-M.; Ji, H.-S.; Han, J.; Kim, S.-K.; Park, H.-Y.; Jeong, T.-S. Phenolic-enriched blueberry-leaf extract attenuates glucose homeostasis, pancreatic β-cell function, and insulin sensitivity in high-fat diet–induced diabetic mice. Nutr. Res. 2020, 73, 83–96. [Google Scholar] [CrossRef]
- de Miranda Pedroso, T.F.; Bonamigo, T.R.; da Silva, J.; Vasconcelos, P.; Felix, J.M.; Cardoso, C.A.L.; Souza, R.I.C.; Dos Santos, A.C.; Volobuff, C.R.F.; Formagio, A.S.N.; et al. Chemical constituents of Cochlospermum regium (Schrank) Pilg. root and its antioxidant, antidiabetic, antiglycation, and anticholinesterase effects in Wistar rats. Biomed. Pharmacother. 2019, 111, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.W.; Hsu, A.; Tan, B.K.H. Chlorogenic acid stimulates glucose transport in skeletal muscle via AMPK activation: A contributor to the beneficial effects of coffee on diabetes. PLoS ONE 2012, 7, e32718. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.W.; Hsu, A.; Song, L.; Huang, D.; Tan, B.K.H. Polyphenols-rich Vernonia amygdalina shows anti-diabetic effects in streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2011, 133, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, G.R.; Ignacimuthu, S.; Paulraj, M.G. Solanum torvum Swartz. fruit containing phenolic compounds shows antidiabetic and antioxidant effects in streptozotocin induced diabetic rats. Food Chem. Toxicol. 2011, 49, 2725–2733. [Google Scholar] [CrossRef]
- Abubakar, S.M.; Ukeyima, M.T.; Spencer, J.P.; Lovegrove, J.A. Acute effects of Hibiscus sabdariffa calyces on postprandial blood pressure, vascular function, blood lipids, biomarkers of insulin resistance and inflammation in humans. Nutrients 2019, 11, 341. [Google Scholar] [CrossRef]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic patients. J. Nutr. 2015, 145, 742–748. [Google Scholar] [CrossRef]
- Kianbakht, S.; Abasi, B.; Dabaghian, F.H. Anti-hyperglycemic effect of Vaccinium arctostaphylos in type 2 diabetic patients: A randomized controlled trial. Forsch. Komplementärmedizin/Res. Complement. Med. 2013, 20, 17–22. [Google Scholar] [CrossRef]
- Anhê, F.F.; Desjardins, Y.; Pilon, G.; Dudonné, S.; Genovese, M.I.; Lajolo, F.M.; Marette, A. Polyphenols and type 2 diabetes: A prospective review. PharmaNutrition 2013, 1, 105–114. [Google Scholar] [CrossRef]
- Calderón-Pérez, L.; Llauradó, E.; Companys, J.; Pla-Pagà, L.; Pedret, A.; Rubió, L.; Gosalbes, M.J.; Yuste, S.; Solà, R.; Valls, R.M. Interplay between dietary phenolic compound intake and the human gut microbiome in hypertension: A cross-sectional study. Food Chem. 2021, 344, 128567. [Google Scholar] [CrossRef]
- Wiese, M.; Bashmakov, Y.; Chalyk, N.; Nielsen, D.S.; Krych, Ł.; Kot, W.; Klochkov, V.; Pristensky, D.; Bandaletova, T.; Chernyshova, M.; et al. Prebiotic Effect of Lycopene and Dark Chocolate on Gut Microbiome with Systemic Changes in Liver Metabolism, Skeletal Muscles and Skin in Moderately Obese Persons. BioMed Res. Int 2019, 2019, 4625279. [Google Scholar] [CrossRef]
- Cuervo, A.; Valdés, L.; Salazar, N.; de los Reyes-Gavilán, C.G.; Ruas-Madiedo, P.; Gueimonde, M.; González, S. Pilot study of diet and microbiota: Interactive associations of fibers and polyphenols with human intestinal bacteria. J. Agric. Food Chem. 2014, 62, 5330–5336. [Google Scholar] [CrossRef]
- Song, M.Y.; Wang, J.H.; Eom, T.; Kim, H. Schisandra chinensis fruit modulates the gut microbiota composition in association with metabolic markers in obese women: A randomized, double-blind placebo-controlled study. Nutr. Res. 2015, 35, 655–663. [Google Scholar] [CrossRef]
- Moreno-Indias, I.; Sánchez-Alcoholado, L.; Pérez-Martínez, P.; Andrés-Lacueva, C.; Cardona, F.; Tinahones, F.; Queipo-Ortuño, M.I. Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food Funct. 2016, 7, 1775–1787. [Google Scholar] [CrossRef]
- Wijayabahu, A.T.; Waugh, S.G.; Ukhanova, M.; Mai, V. Dietary raisin intake has limited effect on gut microbiota composition in adult volunteers. Nutr. J. 2019, 18, 14. [Google Scholar] [CrossRef]
- Rodríguez-Morató, J.; Matthan, N.R.; Liu, J.; de la Torre, R.; Chen, C.O. Cranberries attenuate animal-based diet-induced changes in microbiota composition and functionality: A randomized crossover controlled feeding trial. J. Nutr. Biochem. 2018, 62, 76–86. [Google Scholar] [CrossRef]
- Yuan, X.; Long, Y.; Ji, Z.; Gao, J.; Fu, T.; Yan, M.; Zhang, L.; Su, H.; Zhang, W.; Wen, X.; et al. Green Tea Liquid Consumption Alters the Human Intestinal and Oral Microbiome. Mol. Nutr. Food Res. 2018, 62, e1800178. [Google Scholar] [CrossRef]
- Tessari, P.; Lante, A. A Multifunctional Bread Rich in Beta Glucans and Low in Starch Improves Metabolic Control in Type 2 Diabetes: A Controlled Trial. Nutrients 2017, 9, 297. [Google Scholar] [CrossRef]
- El Khoury, D.; Cuda, C.; Luhovyy, B.L.; Anderson, G.H. Beta glucan: Health benefits in obesity and metabolic syndrome. J. Nutr. Metab. 2012, 2012, 851362. [Google Scholar] [CrossRef]
- Chen, J.; Raymond, K. Beta-glucans in the treatment of diabetes and associated cardiovascular risks. Vasc. Health Risk Manag. 2008, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Andrade, E.F.; Lima, A.R.V.; Nunes, I.E.; Orlando, D.R.; Gondim, P.N.; Zangerônimo, M.G.; Alves, F.H.F.; Pereira, L.J. Exercise and Beta-Glucan Consumption (Saccharomyces cerevisiae) Improve the Metabolic Profile and Reduce the Atherogenic Index in Type 2 Diabetic Rats (HFD/STZ). Nutrients 2016, 8, 792. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Shen, N.; Wang, S.; Zhao, G. Oat Beta-Glucan Ameliorates Insulin Resistance in Mice Fed on High-Fat and High-Fructose Diet. Food Nutr. Res. 2013, 57, 22754. [Google Scholar] [CrossRef]
- Dong, J.; Cai, F.; Shen, R.; Liu, Y. Hypoglycaemic effects and inhibitory effect on intestinal disaccharidases of oat beta-glucan in streptozotocin-induced diabetic mice. Food Chem. 2011, 129, 1066–1071. [Google Scholar] [CrossRef]
- Sima, P.; Vannucci, L.; Vetvicka, V. β-glucans and cholesterol. Int. J. Mol. Med. 2018, 41, 1799–1808. [Google Scholar]
- Zaremba, S.M.; Gow, I.F.; Drummond, S.; McCluskey, J.T.; Steinert, R.E. Effects of oat β-glucan consumption at breakfast on ad libitum eating, appetite, glycemia, insulinemia and GLP-1 concentrations in healthy subjects. Appetite 2018, 128, 197–204. [Google Scholar] [CrossRef]
- Arena, M.P.; Caggianiello, G.; Fiocco, D.; Russo, P.; Torelli, M.; Spano, G.; Capozzi, V. Barley β-glucans-containing food enhances probiotic performances of beneficial bacteria. Int. J. Mol. Sci. 2014, 15, 3025–3039. [Google Scholar] [CrossRef]
- Cao, Y.; Sun, Y.; Zou, S.; Duan, B.; Sun, M.; Xu, X. Yeast β-glucan suppresses the chronic inflammation and improves the microenvironment in adipose tissues of ob/ob mice. J. Agric. Food Chem. 2018, 66, 621–629. [Google Scholar] [CrossRef]
- Cao, Y.; Zou, S.; Xu, H.; Li, M.; Tong, Z.; Xu, M.; Xu, X. Hypoglycemic activity of the Baker’s yeast β-glucan in obese/type 2 diabetic mice and the underlying mechanism. Mol. Nutr. Food Res. 2016, 60, 2678–2690. [Google Scholar] [CrossRef]
- Zhu, Y.; Dong, L.; Huang, L.; Shi, Z.; Dong, J.; Yao, Y.; Shen, R. Effects of oat β-glucan, oat resistant starch, and the whole oat flour on insulin resistance, inflammation, and gut microbiota in high-fat-diet-induced type 2 diabetic rats. J. Funct. Foods 2020, 69, 103939. [Google Scholar] [CrossRef]
- Miyamoto, J.; Watanabe, K.; Taira, S.; Kasubuchi, M.; Li, X.; Irie, J.; Itoh, H.; Kimura, I. Barley β-glucan improves metabolic condition via short-chain fatty acids produced by gut microbial fermentation in high fat diet fed mice. PLoS ONE 2018, 13, e0196579. [Google Scholar] [CrossRef]
- Mio, K.; Otake, N.; Nakashima, S.; Matsuoka, T.; Aoe, S. Ingestion of high β-glucan barley flour enhances the intestinal immune system of diet-induced obese mice by prebiotic effects. Nutrients 2021, 13, 907. [Google Scholar] [CrossRef]
- Nie, C.; Yan, X.; Xie, X.; Zhang, Z.; Zhu, J.; Wang, Y.; Wang, X.; Xu, N.; Luo, Y.; Sa, Z. Structure of β-glucan from Tibetan hull-less barley and its in vitro fermentation by human gut microbiota. Chem. Biol. Technol. Agric. 2021, 8, 12. [Google Scholar] [CrossRef]
- Velikonja, A.; Lipoglavšek, L.; Zorec, M.; Orel, R.; Avguštin, G. Alterations in gut microbiota composition and metabolic parameters after dietary intervention with barley beta glucans in patients with high risk for metabolic syndrome development. Anaerobe 2019, 55, 67–77. [Google Scholar] [CrossRef]
- Aoe, S.; Yamanaka, C.; Fuwa, M.; Tamiya, T.; Nakayama, Y.; Miyoshi, T.; Kitazono, E. Effects of BARLEYmax and high-β-glucan barley line on short-chain fatty acids production and microbiota from the cecum to the distal colon in rats. PLoS ONE 2019, 14, e0218118. [Google Scholar] [CrossRef]
- De Angelis, M.; Montemurno, E.; Vannini, L.; Cosola, C.; Cavallo, N.; Gozzi, G.; Maranzano, V.; Di Cagno, R.; Gobbetti, M.; Gesualdo, L. Effect of Whole-Grain Barley on the Human Fecal Microbiota and Metabolome. Appl. Environ. Microbiol. 2015, 81, 7945–7956. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Kamil, A.; Fleige, L.; Chung, Y.; De Chavez, P.; Marzorati, M. Different Oat Ingredients Stimulate Specific Microbial Metabolites in the Gut Microbiome of Three Human Individuals In Vitro. ACS Omega 2018, 3, 12446–12456. [Google Scholar] [CrossRef]
- Li, M.; Trapika, I.; Tang, S.Y.S.; Cho, J.L.; Qi, Y.; Li, C.G.; Li, Y.; Yao, M.; Yang, D.; Liu, B.; et al. Mechanisms and Active Compounds Polysaccharides and Bibenzyls of Medicinal Dendrobiums for Diabetes Management. Front. Nutr. 2021, 8, 811870. [Google Scholar] [CrossRef]
- Peng, D.; Tian, W.; An, M.; Chen, Y.; Zeng, W.; Zhu, S.; Li, P.; Du, B. Characterization of antidiabetic effects of Dendrobium officinale derivatives in a mouse model of type 2 diabetes mellitus. Food Chem. 2023, 399, 133974. [Google Scholar] [CrossRef]
- Liu, H.; Xing, Y.; Wang, Y.; Ren, X.; Zhang, D.; Dai, J.; Xiu, Z.; Yu, S.; Dong, Y. Dendrobium officinale Polysaccharide Prevents Diabetes via the Regulation of Gut Microbiota in Prediabetic Mice. Foods 2023, 12, 2310. [Google Scholar] [CrossRef]
- Chen, X.; Chen, C.; Fu, X. Dendrobium officinale Polysaccharide Alleviates Type 2 Diabetes Mellitus by Restoring Gut Microbiota and Repairing Intestinal Barrier via the LPS/TLR4/TRIF/NF-kB Axis. J. Agric. Food Chem. 2023, 71, 11929–11940. [Google Scholar] [CrossRef]
- Zou, J.; Song, Q.; Shaw, P.C.; Zuo, Z. Dendrobium officinale regulate lipid metabolism in diabetic mouse liver via PPAR-RXR signaling pathway: Evidence from an integrated multi-omics analysis. Biomed. Pharmacother. 2024, 173, 116395. [Google Scholar] [CrossRef]
- Carrizales-Sanchez, A.K.; Tamez-Rivera, O.; Rodriguez-Gutierrez, N.A.; Elizondo-Montemayor, L.; Gradilla-Hernandez, M.S.; Garcia-Rivas, G.; Pacheco, A.; Senes-Guerrero, C. Characterization of gut microbiota associated with metabolic syndrome and type-2 diabetes mellitus in Mexican pediatric subjects. BMC Pediatr. 2023, 23, 210. [Google Scholar] [CrossRef]
- Ionescu, R.F.; Enache, R.M.; Cretoiu, S.M.; Gaspar, B.S. Gut Microbiome Changes in Gestational Diabetes. Int. J. Mol. Sci. 2022, 23, 12839. [Google Scholar] [CrossRef]
- Polidori, I.; Marullo, L.; Ialongo, C.; Tomassetti, F.; Colombo, R.; di Gaudio, F.; Calugi, G.; Marrone, G.; Noce, A.; Bernardini, S.; et al. Characterization of Gut Microbiota Composition in Type 2 Diabetes Patients: A Population-Based Study. Int. J. Environ. Res. Public Health 2022, 19, 15913. [Google Scholar] [CrossRef]
- Zhang, L.; Chu, J.; Hao, W.; Zhang, J.; Li, H.; Yang, C.; Yang, J.; Chen, X.; Wang, H. Gut Microbiota and Type 2 Diabetes Mellitus: Association, Mechanism, and Translational Applications. Mediat. Inflamm. 2021, 2021, 5110276. [Google Scholar] [CrossRef]
- Ejtahed, H.S.; Hoseini-Tavassol, Z.; Khatami, S.; Zangeneh, M.; Behrouzi, A.; Ahmadi Badi, S.; Moshiri, A.; Hasani-Ranjbar, S.; Soroush, A.R.; Vaziri, F.; et al. Main gut bacterial composition differs between patients with type 1 and type 2 diabetes and non-diabetic adults. J. Diabetes Metab. Disord. 2020, 19, 265–271. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, E.; Liu, C.; Wicks, H.; Yildiz, S.; Razack, F.; Ying, Z.; Kooijman, S.; Koonen, D.P.Y.; Heijink, M.; et al. Dietary butyrate ameliorates metabolic health associated with selective proliferation of gut Lachnospiraceae bacterium 28-4. JCI Insight 2023, 8, e166655. [Google Scholar] [CrossRef]
- Grigorescu, I.; Dumitrascu, D.L. Implication of gut microbiota in diabetes mellitus and obesity. Acta Endocrinol. 2016, 12, 206–214. [Google Scholar] [CrossRef]
- Jiang, L.; Fu, Q.; Wang, S.; Chen, Y.; Li, J.; Xiao, Y.; Sun, R.; Shen, E.A.; Wang, J.; Zhang, Y.; et al. Effect of RG (Coptis root and ginseng) formula in patients with type 2 diabetes mellitus: A study protocol for a randomized controlled and double-blinding trial. Trials 2022, 23, 305. [Google Scholar] [CrossRef]
- Gradisteanu Pircalabioru, G.; Chifiriuc, M.-C.; Picu, A.; Petcu, L.M.; Trandafir, M.; Savu, O. Snapshot into the Type-2-Diabetes-Associated Microbiome of a Romanian Cohort. Int. J. Mol. Sci. 2022, 23, 15023. [Google Scholar] [CrossRef]
- Yin, X.Q.; An, Y.X.; Yu, C.G.; Ke, J.; Zhao, D.; Yu, K. The Association between Fecal Short-Chain Fatty Acids, Gut Microbiota, and Visceral Fat in Monozygotic Twin Pairs. Diabetes Metab. Syndr. Obes. 2022, 15, 359–368. [Google Scholar] [CrossRef]
- Sugawara, Y.; Kanazawa, A.; Aida, M.; Yoshida, Y.; Yamashiro, Y.; Watada, H. Association of gut microbiota and inflammatory markers in obese patients with type 2 diabetes mellitus: Post hoc analysis of a synbiotic interventional study. Biosci. Microbiota Food Health 2022, 41, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Madhulika, A.; Deepika, G.; Rao, G.V.; Reddy, D.N.; Subramanyam, C.; Sasikala, M.; Talukdar, R. Altered intestinal microbiota in patients with chronic pancreatitis: Implications in diabetes and metabolic abnormalities. Sci. Rep. 2017, 7, 43640. [Google Scholar] [CrossRef] [PubMed]
- Aljuraiban, G.S.; Alfhili, M.A.; Aldhwayan, M.M.; Aljazairy, E.A.; Al-Musharaf, S. Metagenomic Shotgun Sequencing Reveals Specific Human Gut Microbiota Associated with Insulin Resistance and Body Fat Distribution in Saudi Women. Biomolecules 2023, 13, 640. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, S.A.; Vella, C.A.; New, D.D.; Aunjum, A.; Antush, M.; Geidl, R.; Andrews, K.R.; Balemba, O.B. High-Resolution Taxonomic Characterization Reveals Novel Human Microbial Strains with Potential as Risk Factors and Probiotics for Prediabetes and Type 2 Diabetes. Microorganisms 2023, 11, 758. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhong, M.; Shen, Q.; Wu, Y.; Cao, M.; Ju, S.; Chen, L. Gut microbiota in early pregnancy among women with Hyperglycaemia vs. Normal blood glucose. BMC Pregnancy Childbirth 2020, 20, 284. [Google Scholar] [CrossRef]
- Allin, K.H.; Tremaroli, V.; Caesar, R.; Jensen, B.A.H.; Damgaard, M.T.F.; Bahl, M.I.; Licht, T.R.; Hansen, T.H.; Nielsen, T.; Dantoft, T.M.; et al. Aberrant intestinal microbiota in individuals with prediabetes. Diabetologia 2018, 61, 810–820. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergstrom, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Backhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef]
- Yan, J.; Wang, Z.; Bao, G.; Xue, C.; Zheng, W.; Fu, R.; Zhang, M.; Ding, J.; Yang, F.; Sun, B. Causal effect between gut microbiota and metabolic syndrome in European population: A bidirectional mendelian randomization study. Cell Biosci. 2024, 14, 67. [Google Scholar] [CrossRef]
- Chen, X.-L.; Cai, K.; Zhang, W.; Su, S.-L.; Zhao, L.-H.; Qiu, L.-P.; Duan, J.-A. Bear bile powder ameliorates type 2 diabetes via modulation of metabolic profiles, gut microbiota, and metabolites. Front. Pharmacol. 2023, 13, 1090955. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.C.; Chien, Y.W.; Yang, S.C. The alteration of gut microbiota in newly diagnosed type 2 diabetic patients. Nutrition 2019, 63–64, 51–56. [Google Scholar] [CrossRef]
- Kallapura, G.; Prakash, A.S.; Sankaran, K.; Manjappa, P.; Chaudhary, P.; Ambhore, S.; Dhar, D. Microbiota based personalized nutrition improves hyperglycaemia and hypertension parameters and reduces inflammation: A prospective, open label, controlled, randomized, comparative, proof of concept study. PeerJ 2024, 12, e17583. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Medina-Vera, I.; Tood, M.; Aguilar Lopez, M.; Guevara, M.; Flores, A.; Tovar, A.; Torres, N. Effect of a combination of functional foods (nopal, soy protein, chia seed and inulin) on the gut microbiota of subjects with type 2 diabetes. FASEB J. 2017, 31. [Google Scholar] [CrossRef]
- Deledda, A.; Palmas, V.; Heidrich, V.; Fosci, M.; Lombardo, M.; Cambarau, G.; Lai, A.; Melis, M.; Loi, E.; Loviselli, A.; et al. Dynamics of Gut Microbiota and Clinical Variables after Ketogenic and Mediterranean Diets in Drug-Naïve Patients with Type 2 Diabetes Mellitus and Obesity. Metabolites 2022, 12, 1092. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhao, L.-H.; Tian, X.; Kong, M.-W.; He, J.-Q.; Ge, X.-C.; Liu, X.-Y.; Feng, Z.-B.; Gao, Y. Characteristics of Gut Microbiota in Female Patients with Diabetic Microvascular Complications. J. Diabetes Res. 2022, 2022, 2980228. [Google Scholar] [CrossRef]
- Huang, W.; Lin, Z.; Sun, A.; Deng, J.; Manyande, A.; Xiang, H.; Zhao, G.F.; Hong, Q. The role of gut microbiota in diabetic peripheral neuropathy rats with cognitive dysfunction. Front. Microbiol. 2023, 14, 1156591. [Google Scholar] [CrossRef]
- Li, H.; Fang, Q.; Nie, Q.; Hu, J.; Yang, C.; Huang, T.; Li, H.; Nie, S. Hypoglycemic and Hypolipidemic Mechanism of Tea Polysaccharides on Type 2 Diabetic Rats via Gut Microbiota and Metabolism Alteration. J. Agric. Food Chem. 2020, 68, 10015–10028. [Google Scholar] [CrossRef]
- Koshida, T.; Gohda, T.; Sugimoto, T.; Asahara, T.; Asao, R.; Ohsawa, I.; Gotoh, H.; Murakoshi, M.; Suzuki, Y.; Yamashiro, Y. Gut Microbiome and Microbiome-Derived Metabolites in Patients with End-Stage Kidney Disease. Int. J. Mol. Sci. 2023, 24, 11456. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.-S.; Zhang, Y.; Ren, J.-N.; Yang, S.-Z.; Li, X.; Fan, G.; Pan, S.-Y. Effect of different ways of ingesting orange essential oil on blood immune index and intestinal microflora in mice. J. Sci. Food Agric. 2023, 103, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.-S.; Song, E.-J.; Lee, S.-Y.; Park, S.-L.; Kim, D.; Kim, D.; Kim, J.-H.; Lim, S.-I.; Nam, Y.-D. Effects of bentonite Bgp35b-p on the gut microbiota of mice fed a high-fat diet. J. Sci. Food Agric. 2018, 98, 4369–4373. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, T.P.M.; Herrema, H.; Yu, H.; Bakker, G.J.; Winkelmeijer, M.; Soukhatcheva, G.; Dai, D.; Ma, C.; Havik, S.R.; Balvers, M.; et al. Gut-derived bacterial flagellin induces beta-cell inflammation and dysfunction. Gut Microbes 2022, 14, 2111951. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Di, J.; Jiang, S.; Li, X.; Hua, F. Machine learning models for prediction of invasion Klebsiella pneumoniae liver abscess syndrome in diabetes mellitus: A singled centered retrospective study. BMC Infect. Dis. 2023, 23, 284. [Google Scholar] [CrossRef]
- Uema, T.; Millman, J.F.; Okamoto, S.; Nakamura, T.; Yamashiro, K.; Uehara, M.; Honma, K.-I.; Miyazato, M.; Ashikari, A.; Saito, S.; et al. Profile of gut microbiota and serum metabolites associated with metabolic syndrome in a remote island most afflicted by obesity in Japan. Sci. Rep. 2022, 12, 17292. [Google Scholar] [CrossRef]
- Kurasawa, M.; Nishikido, T.; Koike, J.; Tominaga, S.; Tamemoto, H. Gas-forming liver abscess associated with rapid hemolysis in a diabetic patient. World J. Diabetes 2014, 5, 224–229. [Google Scholar] [CrossRef]
- Ke, J.; An, Y.; Cao, B.; Lang, J.; Wu, N.; Zhao, D. Orlistat-Induced Gut Microbiota Modification in Obese Mice. Evid. Based Complement. Altern. Med 2020, 2020, 9818349. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- Moffa, S.; Mezza, T.; Cefalo, C.; Cinti, F.; Impronta, F.; Sorice, G.P.; Santoro, A.; Di Giuseppe, G.; Pontecorvi, A.; Giaccari, A. The Interplay Between Immune System and Microbiota in Diabetes. Mediat. Inflamm. 2019, 2019, 9367404. [Google Scholar] [CrossRef]
- Roshanravan, N.; Mahdavi, R.; Alizadeh, E.; Jafarabadi, M.A.; Hedayati, M.; Ghavami, A.; Alipour, S.; Alamdari, N.M.; Barati, M.; Ostadrahimi, A. Effect of Butyrate and Inulin Supplementation on Glycemic Status, Lipid Profile and Glucagon-Like Peptide 1 Level in Patients with Type 2 Diabetes: A Randomized Double-Blind, Placebo-Controlled Trial. Horm. Metab. Res. 2017, 49, 886–891. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, R.F.; Berthon, B.S.; Jensen, M.E.; Baines, K.J.; Wood, L.G. Short-chain fatty acids, prebiotics, synbiotics, and systemic inflammation: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2017, 106, 930–945. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef]
- Holmes, Z.C.; Villa, M.M.; Durand, H.K.; Jiang, S.; Dallow, E.P.; Petrone, B.L.; Silverman, J.D.; Lin, P.H.; David, L.A. Microbiota responses to different prebiotics are conserved within individuals and associated with habitual fiber intake. Microbiome 2022, 10, 114. [Google Scholar] [CrossRef]
- Kaye, D.M.; Shihata, W.A.; Jama, H.A.; Tsyganov, K.; Ziemann, M.; Kiriazis, H.; Horlock, D.; Vijay, A.; Giam, B.; Vinh, A.; et al. Deficiency of Prebiotic Fiber and Insufficient Signaling through Gut Metabolite-Sensing Receptors Leads to Cardiovascular Disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef]
- Wongkrasant, P.; Pongkorpsakol, P.; Ariyadamrongkwan, J.; Meesomboon, R.; Satitsri, S.; Pichyangkura, R.; Barrett, K.E.; Muanprasat, C. A prebiotic fructo-oligosaccharide promotes tight junction assembly in intestinal epithelial cells via an AMPK-dependent pathway. Biomed. Pharmacother. 2020, 129, 110415. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Kang, S.; Hua, C.S.; Ting, Z.; Park, S. Synbiotic effects of beta-glucans from cauliflower mushroom and Lactobacillus fermentum on metabolic changes and gut microbiome in estrogen-deficient rats. Genes Nutr. 2017, 12, 31. [Google Scholar] [CrossRef]
- Richter, E.A.; Ruderman, N.B. AMPK and the biochemistry of exercise: Implications for human health and disease. Biochem. J. 2009, 418, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Ishii, M.; Akagawa, M. Propionate suppresses hepatic gluconeogenesis via GPR43/AMPK signaling pathway. Arch. Biochem. Biophys. 2019, 672, 108057. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, Y.; Chen, S.; Gu, J.; Ni, Y. Insoluble and Soluble Dietary Fibers from Kiwifruit (Actinidia deliciosa) Modify Gut Microbiota to Alleviate High-Fat Diet and Streptozotocin-Induced TYPE 2 Diabetes in Rats. Nutrients 2022, 14, 3369. [Google Scholar] [CrossRef]
- Pesaro, A.E.; Bittencourt, M.S.; Franken, M.; Carvalho, J.A.M.; Bernardes, D.; Tuomilehto, J.; Santos, R.D. The Finnish Diabetes Risk Score (FINDRISC), incident diabetes and low-grade inflammation. Diabetes Res. Clin. Pract. 2021, 171, 108558. [Google Scholar] [CrossRef] [PubMed]
- Naseri, K.; Saadati, S.; Ghaemi, F.; Ashtary-Larky, D.; Asbaghi, O.; Sadeghi, A.; Afrisham, R.; de Courten, B. The effects of probiotic and synbiotic supplementation on inflammation, oxidative stress, and circulating adiponectin and leptin concentration in subjects with prediabetes and type 2 diabetes mellitus: A GRADE-assessed systematic review, meta-analysis, and meta-regression of randomized clinical trials. Eur. J. Nutr. 2023, 62, 543–561. [Google Scholar] [CrossRef]
- Colantonio, A.G.; Werner, S.L.; Brown, M. The Effects of Prebiotics and Substances with Prebiotic Properties on Metabolic and Inflammatory Biomarkers in Individuals with Type 2 Diabetes Mellitus: A Systematic Review. J. Acad. Nutr. Diet. 2020, 120, 587–607.e2. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Jayashree, B.; Bibin, Y.S.; Prabhu, D.; Shanthirani, C.S.; Gokulakrishnan, K.; Lakshmi, B.S.; Mohan, V.; Balasubramanyam, M. Increased circulatory levels of lipopolysaccharide (LPS) and zonulin signify novel biomarkers of proinflammation in patients with type 2 diabetes. Mol. Cell. Biochem. 2014, 388, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef]
- Martinez Baez, A.; Ayala, G.; Pedroza-Saavedra, A.; Gonzalez-Sanchez, H.M.; Chihu Amparan, L. Phosphorylation Codes in IRS-1 and IRS-2 Are Associated with the Activation/Inhibition of Insulin Canonical Signaling Pathways. Curr. Issues Mol. Biol. 2024, 46, 634–649. [Google Scholar] [CrossRef]
- Emamgholipour, S.; Esmaeili, F.; Shabani, M.; Hasanpour, S.Z.; Pilehvari, M.; Zabihi-Mahmoudabadi, H.; Motevasseli, M.; Shanaki, M. Alterations of SOCS1 and SOCS3 transcript levels, but not promoter methylation levels in subcutaneous adipose tissues in obese women. BMC Endocr. Disord. 2023, 23, 7. [Google Scholar] [CrossRef]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-kappaB, inflammation, and metabolic disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef]
- Su, M.; Tang, T.; Tang, W.; Long, Y.; Wang, L.; Liu, M. Astragalus improves intestinal barrier function and immunity by acting on intestinal microbiota to treat T2DM: A research review. Front. Immunol. 2023, 14, 1243834. [Google Scholar] [CrossRef]
- Wibowo, H.; Harbuwono, D.S.; Tahapary, D.L.; Kartika, R.; Pradipta, S.; Larasati, R.A. Impact of Sodium Butyrate Treatment in LPS-Stimulated Peripheral Blood Mononuclear Cells of Poorly Controlled Type 2 DM. Front. Endocrinol. 2021, 12, 652942. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zeng, Y.; Liu, Y.; Wang, X.; Chen, Y.; Lepp, D.; Tsao, R.; Sadakiyo, T.; Zhang, H.; Mine, Y. Regulatory Effect of Isomaltodextrin on a High-Fat Diet Mouse Model with LPS-Induced Low-Grade Chronic Inflammation. J. Agric. Food Chem. 2022, 70, 11258–11273. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Dong, H.; Zang, Z.; Wu, W.; Zhu, W.; Zhang, H.; Guan, Y. Kudzu Resistant Starch: An Effective Regulator of Type 2 Diabetes Mellitus. Oxidative Med. Cell. Longev. 2021, 2021, 4448048. [Google Scholar] [CrossRef]
- Parnell, J.A.; Reimer, R.A. Prebiotic fibres dose-dependently increase satiety hormones and alter Bacteroidetes and Firmicutes in lean and obese JCR:LA-cp rats. Br. J. Nutr. 2012, 107, 601–613. [Google Scholar] [CrossRef]
- Cani, P.D.; Lecourt, E.; Dewulf, E.M.; Sohet, F.M.; Pachikian, B.D.; Naslain, D.; De Backer, F.; Neyrinck, A.M.; Delzenne, N.M. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am. J. Clin. Nutr. 2009, 90, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wu, Y.; Zhang, Q.; Xiao, X. Crosstalk between glucagon-like peptide 1 and gut microbiota in metabolic diseases. mBio 2024, 15, e0203223. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef]
- Brooks, L.; Viardot, A.; Tsakmaki, A.; Stolarczyk, E.; Howard, J.K.; Cani, P.D.; Everard, A.; Sleeth, M.L.; Psichas, A.; Anastasovskaj, J.; et al. Fermentable carbohydrate stimulates FFAR2-dependent colonic PYY cell expansion to increase satiety. Mol. Metab. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Cani, P.D.; Hoste, S.; Guiot, Y.; Delzenne, N.M. Dietary non-digestible carbohydrates promote L-cell differentiation in the proximal colon of rats. Br. J. Nutr. 2007, 98, 32–37. [Google Scholar] [CrossRef]
- Kaji, I.; Karaki, S.; Tanaka, R.; Kuwahara, A. Density distribution of free fatty acid receptor 2 (FFA2)-expressing and GLP-1-producing enteroendocrine L cells in human and rat lower intestine, and increased cell numbers after ingestion of fructo-oligosaccharide. J. Mol. Histol. 2011, 42, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, Y.; Wang, J.; Geng, W. Kombucha Reduces Hyperglycemia in Type 2 Diabetes of Mice by Regulating Gut Microbiota and Its Metabolites. Foods 2022, 11, 754. [Google Scholar] [CrossRef] [PubMed]
- Maselli, D.B.; Camilleri, M. Effects of GLP-1 and Its Analogs on Gastric Physiology in Diabetes Mellitus and Obesity. In Diabetes: From Research to Clinical Practice; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2021; Volume 1307, pp. 171–192. [Google Scholar] [CrossRef]
- Flaig, B.; Garza, R.; Singh, B.; Hamamah, S.; Covasa, M. Treatment of Dyslipidemia through Targeted Therapy of Gut Microbiota. Nutrients 2023, 15, 228. [Google Scholar] [CrossRef] [PubMed]
- Jiao, A.R.; Diao, H.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Huang, Z.Q.; Luo, Y.H.; Luo, J.Q.; Mao, X.B.; et al. Oral administration of short chain fatty acids could attenuate fat deposition of pigs. PLoS ONE 2018, 13, e0196867. [Google Scholar] [CrossRef]
- Prins, G.H.; Rios-Morales, M.; Gerding, A.; Reijngoud, D.J.; Olinga, P.; Bakker, B.M. The Effects of Butyrate on Induced Metabolic-Associated Fatty Liver Disease in Precision-Cut Liver Slices. Nutrients 2021, 13, 4203. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Nehmi, V.A.; Murata, G.M.; Moraes, R.C.M.; Lima, G.C.A.; De Miranda, D.A.; Radloff, K.; Costa, R.G.F.; Jesus, J.C.R.; De Freitas, J.A.; Viana, N.I.; et al. A novel supplement with yeast beta-glucan, prebiotic, minerals and Silybum marianum synergistically modulates metabolic and inflammatory pathways and improves steatosis in obese mice. J. Integr. Med. 2021, 19, 439–450. [Google Scholar] [CrossRef]
- Farhangi, M.A.; Dehghan, P.; Namazi, N. Prebiotic supplementation modulates advanced glycation end-products (AGEs), soluble receptor for AGEs (sRAGE), and cardiometabolic risk factors through improving metabolic endotoxemia: A randomized-controlled clinical trial. Eur. J. Nutr. 2020, 59, 3009–3021. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular Mechanisms Linking Oxidative Stress and Diabetes Mellitus. Oxidative Med. Cell. Longev. 2020, 2020, 8609213. [Google Scholar] [CrossRef]
- Gerber, P.A.; Rutter, G.A. The Role of Oxidative Stress and Hypoxia in Pancreatic Beta-Cell Dysfunction in Diabetes Mellitus. Antioxid. Redox Signal. 2017, 26, 501–518. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Zhu, Y.; Deng, H.; Huang, X.; Jayavanth, P.; Xiao, Y.; Wu, J.; Jiao, R. Theabrownin from Dark Tea Ameliorates Insulin Resistance via Attenuating Oxidative Stress and Modulating IRS-1/PI3K/Akt Pathway in HepG2 Cells. Nutrients 2023, 15, 3862. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Bobryshev, Y.V.; Kozarov, E.; Sobenin, I.A.; Orekhov, A.N. Role of gut microbiota in the modulation of atherosclerosis-associated immune response. Front. Microbiol. 2015, 6, 671. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, H.; Wang, G. Correlations between inflammatory response, oxidative stress, intestinal pathological damage and intestinal flora variation in rats with type 2 diabetes mellitus. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10162–10168. [Google Scholar] [CrossRef] [PubMed]
- Semenova, N.; Garashchenko, N.; Kolesnikov, S.; Darenskaya, M.; Kolesnikova, L. Gut Microbiome Interactions with Oxidative Stress: Mechanisms and Consequences for Health. Pathophysiology 2024, 31, 309–330. [Google Scholar] [CrossRef]
- Aryal, D.; Joshi, S.; Thapa, N.K.; Chaudhary, P.; Basaula, S.; Joshi, U.; Bhandari, D.; Rogers, H.M.; Bhattarai, S.; Sharma, K.R.; et al. Dietary phenolic compounds as promising therapeutic agents for diabetes and its complications: A comprehensive review. Food Sci. Nutr. 2024, 12, 3025–3045. [Google Scholar] [CrossRef] [PubMed]
- Polce, S.A.; Burke, C.; Franca, L.M.; Kramer, B.; Paes, A.M.d.A.; Carrillo-Sepulveda, M.A. Ellagic Acid Alleviates Hepatic Oxidative Stress and Insulin Resistance in Diabetic Female Rats. Nutrients 2018, 10, 531. [Google Scholar] [CrossRef]
- Duan, J.; Yang, M.; Liu, Y.; Xiao, S.; Zhang, X. Curcumin protects islet beta cells from streptozotocin-induced type 2 diabetes mellitus injury via its antioxidative effects. Endokrynol. Pol. 2022, 73, 942–946. [Google Scholar] [CrossRef]
- Basu, A.; Newman, E.D.; Bryant, A.L.; Lyons, T.J.; Betts, N.M. Pomegranate polyphenols lower lipid peroxidation in adults with type 2 diabetes but have no effects in healthy volunteers: A pilot study. J. Nutr. Metab. 2013, 2013, 708381. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prebiotic | Study Type/Study Duration/Prebiotic Dosage | Results/Implications | Reference |
|---|---|---|---|
| Inulin | RCT/2 months/10 g | 9% decrease in fasting blood glucose 10.5% decrease in HgbA1c levels ~19% increase in total antioxidant capacity Insulin resistance markers unchanged in this study | [22] |
| RCT/8 weeks/10 g | 9.5% decrease in fasting blood glucose 8.4% decrease in HgbA1c 8% decrease in IL-6 and ~20% decrease in TNFα 31.7% decrease in CRP | [23] | |
| Randomized Crossover Trial/2 weeks/30 g | Significant increase in incremental postprandial insulin release at 30 min and 60 min Significant reduction in insulin resistance as measured by HOMA-IR score | [24] | |
| Prospective Single-Arm Study/6 months/15 g | Decreased fasting glucose, 2 h post-OGTT insulin Improved HOMA-IR score Increased relative abundance of Bifidobacterium and Lactobacillus Decreased Alistipes | [25] | |
| RCT/12 weeks/10 g | No significant effects on cholesterol, blood sugar, or HgbA1c | [26] | |
| RCT/6 weeks/16 g | Significant increase in Bifidobacterium in T2DM patients Significantly higher fecal concentrations of total SCFA | [31] | |
| RCT/45 days/10 g | Decreased relative expression of TLR4, NF-κB1, Caspase-1, NLRP3, IL-1β, and IL-18 Improved total antioxidant capacity Increased superoxide dismutase and catalase enzymatic activity | [35] | |
| Longitudinal/2 months/10 g | INS gene unmethylation allows for improved insulin sensitivity and metabolic parameters IRS1 gene methylation observed through findings is not significant | [36] | |
| RCT/45 days | Improved glycemic indices, lipid profile, and GLP-1 secretion | [211] | |
| Resistant Starches | Meta-Analysis of 36 RCTs | Resistant starch type 2 improved acute postprandial insulin response Resistant starch types 1 and 2 improved postprandial glucose Resistant starch type 2 improved fasting glucose | [53] |
| Meta-Analysis of 13 Case–Control Studies | Resistant starch reduced fasting insulin and fasting glucose while increasing insulin sensitivity Metabolic parameters including LDL concentration and HgbA1c were improved | [54] | |
| Meta-Analysis of 19 RCTs | Effects of fasting insulin and glucose tolerance test were not significant Effect size on improving fasting glucose was larger when resistant starch dose was greater than 28 g/day and intervention period was greater than 8 weeks | [55] | |
| Meta-Analysis of 16 RCTs | Improved total antioxidant capacity Reduced CRP concentration in T2DM patients Reduced IL-6 and TNF concentrations | [56] | |
| RCT/8 weeks/10 g | Resistant starch type 2 decreased Hgb A1c by 3%, TNF by 19%, and TG by 15% Increased HFL by 25% Changes in fasting blood glucose, CRP not significant in this study | [58] | |
| RCT/12 weeks/40 g | Resistant starch type 2 significantly lowered postprandial glucose Postprandial GLP1 was higher indicating beneficial effects on meal handling | [57] | |
| Fructo- oligosaccharides | Randomized Crossover Study/20 days/15 g | FOSs did not significantly affect fasting blood glucose concentrations, serum total cholesterol, serum TG, serum free fatty acids, or serum acetate | [69] |
| Double-Blind Crossover Study/4 weeks/20 g | FOSs had no effect on plasma glucose, insulin concentrations, or basal hepatic glucose production No effects were observed on glycated hemoglobin | [70] | |
| Randomized Crossover trial/8 weeks/Polyphenol + 8 g FOS | FOSs reduced hepatic insulin resistance Adding FOSs to polyphenols improved β-cell function Increased Eubacterium and Bifidobacterium Decreased Ruminococcus gnavus, a species correlated with increased hepatic insulin resistance in this study No effects were observed on plasma cholesterol or LDL | [71] | |
| Crossover RCT/Short-Term Intake (2 h)/20 g | Increased gastric emptying in the short term Reduction in small intestinal transit No changes in incretin hormones or subjective feelings of hunger or satiety | [76] | |
| RCT/180 min | FOS-containing yacon syrup had no effect on GLP-1 levels or subjective appetite sensation | [77] | |
| Galacto- oligosaccharides | RCT/12 weeks | Increased concentrations of fecal Bifidobacterium spp. Decreased fecal calprotectin, plasma CRP, and serum total cholesterol Decreased serum insulin was noted | [82] |
| RCT/12 weeks/5.5 g | No significant effects on clinical outcomes including glucose tolerance, intestinal permeability, and gut microbiota Changes in Veillonellaceae, however, correlated inversely with IL-6 and glucose response | [83] | |
| RCT/12 months/15 g | Increased abundance of Bifidobacterium spp. No differences in fecal SCFA concentrations No significant changes in incretins, LPSs, or other markers of inflammation No significant changes in insulin sensitivity | [84] | |
| RCT/4 weeks/10 g | No improvement in glucose tolerance during study period Marked restoration of Bifidobacterium spp. No significant effects on LPS-binding protein | [86] | |
| Pectic Oligosaccharides | Clinical Trial/120 min/10 g | Markedly decreased postprandial blood glucose and significantly lowered insulin levels in non-insulin-dependent diabetes In insulin-dependent diabetics, similar results were shown in postprandial glucose | [96] |
| Clinical Trial/45 min/10 g | Decreased postprandial glucose and insulin levels | [97] | |
| RCT/2 weeks/30 g | Significantly reduced fasting blood glucose values, and HgbA1c and HOMA-IR values were observed | [98] | |
| Randomized Crossover/180 min | Significant reduction in postprandial blood glucose and insulin responses throughout 180 min Glucose was lowered by 13.2% | [99] | |
| Polyphenols | N/A: In Vitro Analysis | Polyphenolic extracts and digests from oregano exhibited cellular antioxidant capacity These extracts promoted hypoglycemic and hypolipidemic properties | [125] |
| N/A: In Vitro Analysis | Significantly increased glucose consumption and glycogen content in hepatic cells Attenuated ROS overproduction and glutathione depletion in hepatic cells | [126] | |
| N/A: In Vitro Analysis | Upregulated GLP-1 release in dose-dependent manner Proglucagon, its precursor, and mRNA expression was increased 2.68-fold | [128] | |
| RCT/1–2 h/250 milliliters | Reduction in serum glucose, plasma insulin, serum TG, and CRP levels Significant improvement in antioxidant response | [135] | |
| RCT/24 weeks | Lowered fasting plasma glucose by 8.5% and improved HOMA-IR score by 13% β-hydroxybutyrate was elevated by 42.4% Significantly decreased serum LDL by 8% and TG by 23%, while increasing HDL by 19% | [136] | |
| RCT/2 months/350 mg every 8 h | Lowered fasting blood glucose, 2 h postprandial glucose and HgbA1c No significant effect on liver or kidney function | [137] | |
| β-glucans | Controlled Trial/6 months/7 g | Reduction in HgbA1c by 0.5 points Postprandial and plasma glucose was decreased No significant change in body weight or plasma lipids | [147] |
| RCT/90 min/4 g | GLP-1 was significantly reduced at 90 min Blood glucose was reduced at 30 min Plasma insulin was reduced at 30 and 60 min | [154] | |
| RCT/4 weeks/6 g | Increased SCFA concentrations with 43% increase in propionic acid Higher abundances of Bifidobacterium and Akkermansia in metabolic-responsive individuals | [162] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iatcu, O.C.; Hamamah, S.; Covasa, M. Harnessing Prebiotics to Improve Type 2 Diabetes Outcomes. Nutrients 2024, 16, 3447. https://doi.org/10.3390/nu16203447
Iatcu OC, Hamamah S, Covasa M. Harnessing Prebiotics to Improve Type 2 Diabetes Outcomes. Nutrients. 2024; 16(20):3447. https://doi.org/10.3390/nu16203447
Chicago/Turabian StyleIatcu, Oana C., Sevag Hamamah, and Mihai Covasa. 2024. "Harnessing Prebiotics to Improve Type 2 Diabetes Outcomes" Nutrients 16, no. 20: 3447. https://doi.org/10.3390/nu16203447
APA StyleIatcu, O. C., Hamamah, S., & Covasa, M. (2024). Harnessing Prebiotics to Improve Type 2 Diabetes Outcomes. Nutrients, 16(20), 3447. https://doi.org/10.3390/nu16203447