
Umbilical Cord Plasma Lysophospholipids and Triacylglycerols Associated with Birthweight Percentiles

 , , , , ,
, , , , ,
Abstract
1. Introduction
1.1. Maternal Glycemia and Offspring Birthweight
1.2. Placental Lipid Metabolism and Transplacental Lipid Supply
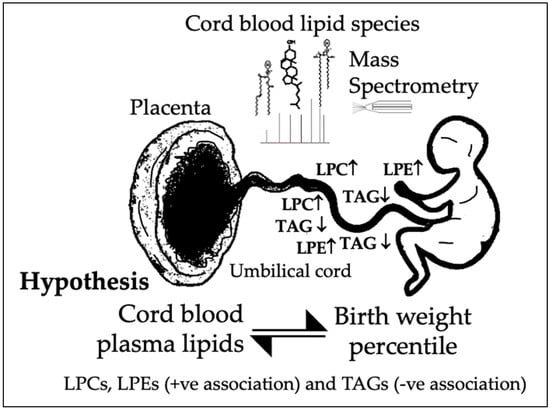
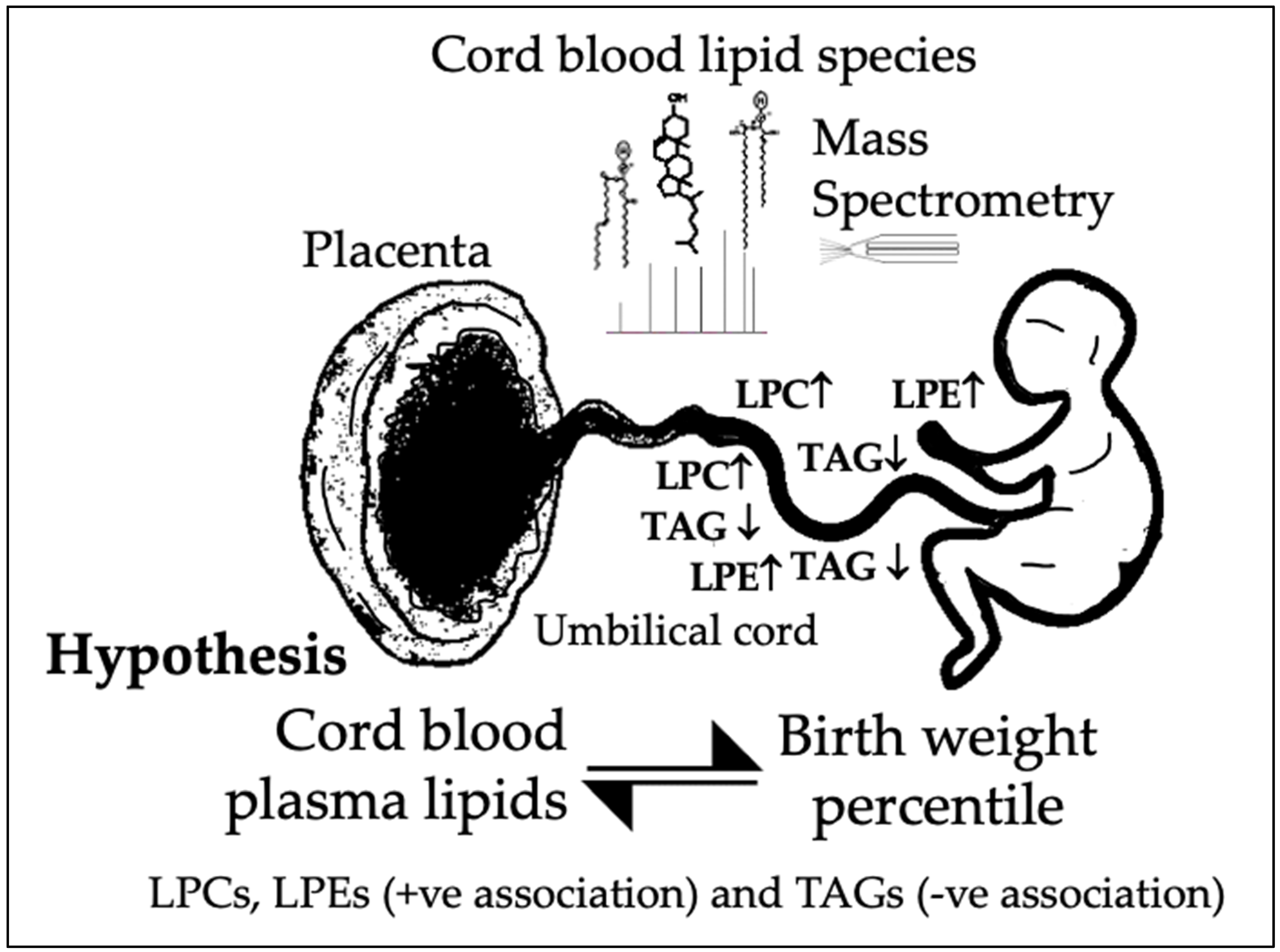
1.3. Hypothesis and Aims
2. Materials and Methods
2.1. The GUSTO Cohort and Sample Selection for This Sub-Study
2.2. Data and Cord Blood Collection
2.3. Cord Blood Plasma Lipidomic Analysis
2.4. Statistical Analysis
3. Results
3.1. Cohort Characteristics
3.2. Association of Cord Blood Lipid Species with Maternal Glucose
3.3. Association of Cord Blood Lipid Species with Birthweight
3.4. Sensitivity Analyses
4. Discussion
4.1. Cord Blood Lysophospholipids and Birthweight Percentile
4.2. Cord Blood Triacylglycerols and Birthweight Percentile
4.3. Cord Blood Lipids and Maternal Glycemia
4.4. Limitations and Strengths of Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Group, H.S.C.R.; Metzger, B.E.; Lowe, L.P.; Dyer, A.R.; Trimble, E.R.; Chaovarindr, U.; Coustan, D.R.; Hadden, D.R.; McCance, D.R.; Hod, M.; et al. Hyperglycemia and adverse pregnancy outcomes. N. Engl. J. Med. 2008, 358, 1991–2002. [Google Scholar] [CrossRef]
- Kc, K.; Shakya, S.; Zhang, H. Gestational diabetes mellitus and macrosomia: A literature review. Ann. Nutr. Metab. 2015, 66 (Suppl. S2), 14–20. [Google Scholar] [CrossRef]
- Ladfors, L.; Shaat, N.; Wiberg, N.; Katasarou, A.; Berntorp, K.; Kristensen, K. Fetal overgrowth in women with type 1 and type 2 diabetes mellitus. PLoS ONE 2017, 12, e0187917. [Google Scholar] [CrossRef]
- Schaefer-Graf, U.M.; Graf, K.; Kulbacka, I.; Kjos, S.L.; Dudenhausen, J.; Vetter, K.; Herrera, E. Maternal lipids as strong determinants of fetal environment and growth in pregnancies with gestational diabetes mellitus. Diabetes Care 2008, 31, 1858–1863. [Google Scholar] [CrossRef] [PubMed]
- Herrera, E.; Ortega-Senovilla, H. Implications of Lipids in Neonatal Body Weight and Fat Mass in Gestational Diabetic Mothers and Non-Diabetic Controls. Curr. Diab. Rep. 2018, 18, 7. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.; Ursell, T.; Wiggins, P.; Sens, P. Emerging roles for lipids in shaping membrane-protein function. Nature 2009, 459, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Ferchaud-Roucher, V.; Kramer, A.; Silva, E.; Pantham, P.; Weintraub, S.T.; Jansson, T.; Powell, T.L. A potential role for lysophosphatidylcholine in the delivery of long chain polyunsaturated fatty acids to the fetal circulation. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Perazzolo, S.; Hirschmugl, B.; Wadsack, C.; Desoye, G.; Lewis, R.M.; Sengers, B.G. The influence of placental metabolism on fatty acid transfer to the fetus. J. Lipid Res. 2017, 58, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Duttaroy, A.K.; Basak, S. Maternal Fatty Acid Metabolism in Pregnancy and Its Consequences in the Feto-Placental Development. Front Physiol 2021, 12, 787848. [Google Scholar] [CrossRef] [PubMed]
- Soh, S.E.; Tint, M.T.; Gluckman, P.D.; Godfrey, K.M.; Rifkin-Graboi, A.; Chan, Y.H.; Stunkel, W.; Holbrook, J.D.; Kwek, K.; Chong, Y.S.; et al. Cohort profile: Growing Up in Singapore Towards healthy Outcomes (GUSTO) birth cohort study. Int. J. Epidemiol. 2014, 43, 1401–1409. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.; Weir, J.M.; Mishra, P.; Huynh, K.; Nijagal, B.; Gupta, V.; Broekman, B.F.P.; Chong, M.F.; Chan, S.Y.; Tan, K.H.; et al. The placental lipidome of maternal antenatal depression predicts socio-emotional problems in the offspring. Transl. Psychiatry 2021, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Mikolajczyk, R.T.; Zhang, J.; Betran, A.P.; Souza, J.P.; Mori, R.; Gulmezoglu, A.M.; Merialdi, M. A global reference for fetal-weight and birthweight percentiles. Lancet 2011, 377, 1855–1861. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.H.Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- LaBarre, J.L.; Puttabyatappa, M.; Song, P.X.K.; Goodrich, J.M.; Zhou, L.; Rajendiran, T.M.; Soni, T.; Domino, S.E.; Treadwell, M.C.; Dolinoy, D.C.; et al. Maternal lipid levels across pregnancy impact the umbilical cord blood lipidome and infant birth weight. Sci Rep 2020, 10, 14209. [Google Scholar] [CrossRef]
- Kadakia, R.; Scholtens, D.M.; Rouleau, G.W.; Talbot, O.; Ilkayeva, O.R.; George, T.; Josefson, J.L. Cord Blood Metabolites Associated with Newborn Adiposity and Hyperinsulinemia. J Pediatr 2018, 203, 144–149.e141. [Google Scholar] [CrossRef]
- Hellmuth, C.; Uhl, O.; Standl, M.; Demmelmair, H.; Heinrich, J.; Koletzko, B.; Thiering, E. Cord Blood Metabolome Is Highly Associated with Birth Weight, but Less Predictive for Later Weight Development. Obes. Facts 2017, 10, 85–100. [Google Scholar] [CrossRef]
- Lu, Y.P.; Reichetzeder, C.; Prehn, C.; Yin, L.H.; Yun, C.; Zeng, S.; Chu, C.; Adamski, J.; Hocher, B. Cord Blood Lysophosphatidylcholine 16: 1 is Positively Associated with Birth Weight. Cell. Physiol. Biochem. 2018, 45, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Paton, C.M.; Ntambi, J.M. Biochemical and physiological function of stearoyl-CoA desaturase. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E28–E37. [Google Scholar] [CrossRef] [PubMed]
- Aiyar, N.; Disa, J.; Ao, Z.; Ju, H.; Nerurkar, S.; Willette, R.N.; Macphee, C.H.; Johns, D.G.; Douglas, S.A. Lysophosphatidylcholine induces inflammatory activation of human coronary artery smooth muscle cells. Mol. Cell Biochem. 2007, 295, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Sevastou, I.; Kaffe, E.; Mouratis, M.A.; Aidinis, V. Lysoglycerophospholipids in chronic inflammatory disorders: The PLA(2)/LPC and ATX/LPA axes. Biochim. Biophys. Acta 2013, 1831, 42–60. [Google Scholar] [CrossRef]
- Sugasini, D.; Yalagala, P.C.R.; Goggin, A.; Tai, L.M.; Subbaiah, P.V. Enrichment of brain docosahexaenoic acid (DHA) is highly dependent upon the molecular carrier of dietary DHA: Lysophosphatidylcholine is more efficient than either phosphatidylcholine or triacylglycerol. J. Nutr. Biochem. 2019, 74, 108231. [Google Scholar] [CrossRef]
- Yea, K.; Kim, J.; Yoon, J.H.; Kwon, T.; Kim, J.H.; Lee, B.D.; Lee, H.J.; Lee, S.J.; Kim, J.I.; Lee, T.G.; et al. Lysophosphatidylcholine activates adipocyte glucose uptake and lowers blood glucose levels in murine models of diabetes. J. Biol. Chem. 2009, 284, 33833–33840. [Google Scholar] [CrossRef] [PubMed]
- Geraghty, A.A.; Alberdi, G.; O’Sullivan, E.J.; O’Brien, E.C.; Crosbie, B.; Twomey, P.J.; McAuliffe, F.M. Maternal Blood Lipid Profile during Pregnancy and Associations with Child Adiposity: Findings from the ROLO Study. PLoS ONE 2016, 11, e0161206. [Google Scholar] [CrossRef]
- Jenkins, B.J.; Seyssel, K.; Chiu, S.; Pan, P.H.; Lin, S.Y.; Stanley, E.; Ament, Z.; West, J.A.; Summerhill, K.; Griffin, J.L.; et al. Odd Chain Fatty Acids; New Insights of the Relationship Between the Gut Microbiota, Dietary Intake, Biosynthesis and Glucose Intolerance. Sci. Rep. 2017, 7, 44845. [Google Scholar] [CrossRef] [PubMed]
- Bensadoun, A. Lipoprotein lipase. Annu. Rev. Nutr. 1991, 11, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Hide, W.A.; Chan, L.; Li, W.H. Structure and evolution of the lipase superfamily. J. Lipid Res. 1992, 33, 167–178. [Google Scholar] [CrossRef]
- Zechner, R.; Strauss, J.; Frank, S.; Wagner, E.; Hofmann, W.; Kratky, D.; Hiden, M.; Levak-Frank, S. The role of lipoprotein lipase in adipose tissue development and metabolism. Int. J. Obes. Relat. Metab. Disord. 2000, 24 (Suppl. S4), S53–S56. [Google Scholar] [CrossRef] [PubMed]
- Holtrop, P.; Swails, T.; Riggs, T. Hypertriglyceridemia in extremely low birth weight infants receiving lipid emulsions. J. Neonatal. Perinatal. Med. 2015, 8, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Lowe, W.L., Jr.; Bain, J.R.; Nodzenski, M.; Reisetter, A.C.; Muehlbauer, M.J.; Stevens, R.D.; Ilkayeva, O.R.; Lowe, L.P.; Metzger, B.E.; Newgard, C.B.; et al. Maternal BMI and Glycemia Impact the Fetal Metabolome. Diabetes Care 2017, 40, 902–910. [Google Scholar] [CrossRef]
- Patel, N.; Hellmuth, C.; Uhl, O.; Godfrey, K.; Briley, A.; Welsh, P.; Pasupathy, D.; Seed, P.T.; Koletzko, B.; Poston, L.; et al. Cord Metabolic Profiles in Obese Pregnant Women: Insights Into Offspring Growth and Body Composition. J. Clin. Endocrinol. Metab. 2018, 103, 346–355. [Google Scholar] [CrossRef]
- Rico, J.E.; Saed Samii, S.; Mathews, A.T.; Lovett, J.; Haughey, N.J.; McFadden, J.W. Temporal changes in sphingolipids and systemic insulin sensitivity during the transition from gestation to lactation. PLoS ONE 2017, 12, e0176787. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulou, M.; Gordillo, R.; Koliaki, C.; Gancheva, S.; Jelenik, T.; De Filippo, E.; Herder, C.; Markgraf, D.; Jankowiak, F.; Esposito, I.; et al. Specific Hepatic Sphingolipids Relate to Insulin Resistance, Oxidative Stress, and Inflammation in Nonalcoholic Steatohepatitis. Diabetes Care 2018, 41, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.C.; Shulman, G.I. Roles of Diacylglycerols and Ceramides in Hepatic Insulin Resistance. Trends Pharmacol. Sci. 2017, 38, 649–665. [Google Scholar] [CrossRef] [PubMed]
- de Seymour, J.; Chia, A.; Colega, M.; Jones, B.; McKenzie, E.; Shirong, C.; Godfrey, K.; Kwek, K.; Saw, S.M.; Conlon, C.; et al. Maternal Dietary Patterns and Gestational Diabetes Mellitus in a Multi-Ethnic Asian Cohort: The GUSTO Study. Nutrients 2016, 8, 574. [Google Scholar] [CrossRef]
- Wang, W.; Xie, X.; Yuan, T.; Wang, Y.; Zhao, F.; Zhou, Z.; Zhang, H. Epidemiological trends of maternal hypertensive disorders of pregnancy at the global, regional, and national levels: A population-based study. BMC Pregnancy Childbirth 2021, 21, 364. [Google Scholar] [CrossRef]

{kind=link}
| Mean (SD) or | Range | |
|---|---|---|
| n (%) | (Lowest, Highest) | |
| Maternal age (years) | 31.91 (4.91) | (19.22, 41.10) |
| Pre-pregnancy BMI (kg/m2) | 21.78 (4.01) | (15.30, 36.90) |
| Spontaneous conception | 69 (92.0%) | |
| Parity | ||
| Nulliparous | 43 (57.3%) | |
| Multiparous | 32 (42.7%) | |
| Plasma glucose in 75 g OGTT Ϯ | ||
| Fasting glucose (mmol/L) | 4.42 (0.62) | (3.60, 7.90) |
| 2 h post-load glucose (mmol/L) | 6.60 (1.65) | (3.30, 12.20) |
| Gestational diabetes § | 15 (20.0%) | |
| Hypertensive disorders | ||
| Chronic hypertension Pre-eclampsia | 1 (1.3%) 2 (2.7%) | |
| Pregnancy-induced hypertension | 6 (8.0%) | |
| Female neonate | 38 (50.7%) | |
| Gestational age at delivery (weeks) | 39.15 (0.96) | (37.43, 41.14) |
| Mode of delivery | ||
| Vaginal | 51 (68.0%) | |
| Intrapartum cesarean section | 10 (13.3%) | |
| Non-labor cesarean section | 14 (18.7%) | |
| Standardized birthweight percentile # | 53.55 (34.79) | (1.25, 99.98) |
| Size at birth § | ||
| SGA (<10th percentile) | 14 (18.7%) | |
| LGA (>90th percentile) | 16 (21.3%) |
| Lipid | Beta Coefficient (SD log10 Lipid per mmol/L Glucose) Ϯ | LCL | UCL | p-Value | FDR- Corrected p-Value |
|---|---|---|---|---|---|
| DAG38:3 | 0.64 | 0.26 | 1.02 | 1.48 × 10−3 | 2.13 × 10−1 |
| PC(O-36:2) | 0.59 | 0.23 | 0.95 | 2.17 × 10−3 | 2.13 × 10−1 |
| PC(P-36:1) | 0.59 | 0.23 | 0.95 | 2.17 × 10−3 | 2.13 × 10−1 |
| oddPC 35:2 | 0.56 | 0.2 | 0.93 | 3.49 × 10−3 | 2.13 × 10−1 |
| DAG37:4 | 0.55 | 0.19 | 0.91 | 3.72 × 10−3 | 2.13 × 10−1 |
| DAG38:2 | 0.55 | 0.19 | 0.92 | 3.99 × 10−3 | 2.13 × 10−1 |
| SM d18:1/18:0 | 0.58 | 0.2 | 0.96 | 4.27 × 10−3 | 2.13 × 10−1 |
| DAG43:4 | 0.56 | 0.18 | 0.93 | 4.83 × 10−3 | 2.13 × 10−1 |
| PC(P-38:1) | 0.58 | 0.19 | 0.97 | 4.93 × 10−3 | 2.13 × 10−1 |
| DAG42:4 | 0.52 | 0.16 | 0.88 | 5.98 × 10−3 | 2.13 × 10−1 |
| PC 32:2 | 0.55 | 0.17 | 0.93 | 6.16 × 10−3 | 2.13 × 10−1 |
| DAG46:5 | 0.51 | 0.15 | 0.87 | 6.73 × 10−3 | 2.13 × 10−1 |
| DAG37:3 | 0.53 | 0.15 | 0.91 | 7.37 × 10−3 | 2.13 × 10−1 |
| SM d18:1/20:1 | 0.53 | 0.16 | 0.91 | 7.37 × 10−3 | 2.13 × 10−1 |
| SM d18:1/18:1 | 0.53 | 0.15 | 0.91 | 8.15 × 10−3 | 2.19 × 10−1 |
| PS 38:3 | 0.53 | 0.14 | 0.93 | 9.63 × 10−3 | 2.22 × 10−1 |
| DAG39:1 | 0.5 | 0.12 | 0.88 | 1.19 × 10−2 | 2.22 × 10−1 |
| DAG42:3 | 0.47 | 0.11 | 0.83 | 1.21 × 10−2 | 2.22 × 10−1 |
| DAG43:5 | 0.48 | 0.11 | 0.85 | 1.42 × 10−2 | 2.22 × 10−1 |
| MHCer 18:2/C22:0 | 0.48 | 0.11 | 0.86 | 1.46 × 10−2 | 2.22 × 10−1 |
| MHCer 18:1/C20:0 | 0.51 | 0.11 | 0.91 | 1.47 × 10−2 | 2.22 × 10−1 |
| DAG38:4 | 0.51 | 0.11 | 0.91 | 1.47 × 10−2 | 2.22 × 10−1 |
| Cer d17:0/C23:1 | 0.51 | 0.11 | 0.91 | 1.48 × 10−2 | 2.22 × 10−1 |
| Cer d18:1/C26:1 | 0.51 | 0.11 | 0.91 | 1.50 × 10−2 | 2.22 × 10−1 |
| SM d18:1/22:0 | 0.48 | 0.1 | 0.86 | 1.53 × 10−2 | 2.22 × 10−1 |
| Lipid | Beta Coefficient (SD log10 Lipid per mmol/L Glucose) Ϯ | LCL | UCL | p-Value | FDR- Corrected p-Value |
|---|---|---|---|---|---|
| Cer d18:1/C33:1 | 0.36 | 0.21 | 0.51 | 1.22 × 10−5 | 4.93 × 10−3 |
| Cer d18:2/C16:0 | 0.3 | 0.14 | 0.45 | 3.27 × 10−4 | 6.61 × 10−2 |
| Cer d19:0/C15:2 | 0.28 | 0.13 | 0.44 | 7.25 × 10−4 | 7.26 × 10−2 |
| PC 36:3 | 0.26 | 0.11 | 0.41 | 1.24 × 10−3 | 7.26 × 10−2 |
| PC(O-36:2) | 0.25 | 0.1 | 0.39 | 1.48 × 10−3 | 7.26 × 10−2 |
| PC(P-36:1) | 0.25 | 0.1 | 0.39 | 1.48 × 10−3 | 7.26 × 10−2 |
| LPC 14:0 | 0.25 | 0.1 | 0.4 | 1.57 × 10−3 | 7.26 × 10−2 |
| DAG47:6 | 0.24 | 0.1 | 0.38 | 1.60 × 10−3 | 7.26 × 10−2 |
| Cer d18:1/C33:0 | 0.27 | 0.11 | 0.43 | 1.76 × 10−3 | 7.26 × 10−2 |
| LPC 16:1 | 0.24 | 0.1 | 0.39 | 1.80 × 10−3 | 7.26 × 10−2 |
| PC(P-38:1) | 0.25 | 0.1 | 0.41 | 2.17 × 10−3 | 7.96 × 10−2 |
| LPC 20:2 | 0.23 | 0.09 | 0.37 | 2.45 × 10−3 | 8.07 × 10−2 |
| PE(P-18:0/22:5) | 0.24 | 0.09 | 0.39 | 2.71 × 10−3 | 8.07 × 10−2 |
| DAG43:4 | 0.24 | 0.09 | 0.39 | 2.80 × 10−3 | 8.07 × 10−2 |
| oddPC 35:2 | 0.22 | 0.07 | 0.37 | 4.54 × 10−3 | 1.22 × 10−1 |
| PC 38:3 | 0.23 | 0.07 | 0.38 | 5.33 × 10−3 | 1.35 × 10−1 |
| LPC 16:0 | 0.22 | 0.07 | 0.38 | 6.20 × 10−3 | 1.44 × 10−1 |
| PE(O-18:2/20:3) | 0.22 | 0.07 | 0.37 | 6.44 × 10−3 | 1.44 × 10−1 |
| PE(O-40:6) | 0.21 | 0.06 | 0.37 | 7.76 × 10−3 | 1.51 × 10−1 |
| PE(P-16:0/20:4) | 0.22 | 0.06 | 0.38 | 8.21 × 10−3 | 1.51 × 10−1 |
| PE(P-36:4) | 0.22 | 0.06 | 0.38 | 8.21 × 10−3 | 1.51 × 10−1 |
| Cer d18:1/C26:0 | 0.23 | 0.06 | 0.39 | 8.23 × 10−3 | 1.51 × 10−1 |
| DAG46:6 | 0.2 | 0.05 | 0.35 | 9.16 × 10−3 | 1.61 × 10−1 |
| DAG47:5 | 0.2 | 0.05 | 0.34 | 1.01 × 10−2 | 1.71 × 10−1 |
| Cer d18:1/C28:0 | 0.21 | 0.05 | 0.37 | 1.16 × 10−2 | 1.78 × 10−1 |
| Association between Lipid and Standardized Birthweight Percentile † | Fasting Glucose Influence on Standardized Birthweight Percentile † | 2 h Glucose Influence on Standardized Birthweight Percentile † | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lipid Species (Predictor) | Beta Coefficient (BW%/SD log10 Lipid) †† | Lipid LCL | Lipid UCL | FDR- Corrected p-Value | Beta Coefficient (BW%/mmol/L) ††† | Lipid LCL | Lipid UCL | FDR- Corrected p-Value | Beta Coefficient (BW%/mmol/L) ††† | Lipid LCL | Lipid UCL | FDR- Corrected p-Value |
| LPC 20:2 | 21.28 | 12.7 | 29.87 | 2.25 × 10−3 | 11.19 | −6.87 | 29.25 | 4.51 × 10−1 | −3.86 | −11.42 | 3.7 | 9.79 × 10−1 |
| LPC 18:1 | 18.99 | 11 | 26.99 | 2.25 × 10−3 | 9.55 | −8.71 | 27.81 | 4.51 × 10−1 | −1.84 | −9.33 | 5.64 | 9.79 × 10−1 |
| LPC 16:1 | 18.38 | 9.38 | 27.38 | 7.07 × 10−3 | 11.95 | −6.96 | 30.85 | 4.51 × 10−1 | −3.66 | −11.62 | 4.31 | 9.79 × 10−1 |
| LPC 18:2 | 16.42 | 8.29 | 24.55 | 7.07 × 10−3 | 11.38 | −7.56 | 30.32 | 4.51 × 10−1 | −2.49 | −10.34 | 5.36 | 9.79 × 10−1 |
| LPE 18:1 | 16.15 | 8.15 | 24.15 | 7.07 × 10−3 | 4.34 | −14.85 | 23.54 | 6.61 × 10−1 | 3.18 | −4.58 | 10.94 | 9.79 × 10−1 |
| LPC 20:3 | 15.87 | 7.14 | 24.6 | 1.51 × 10−2 | 10.32 | −8.98 | 29.62 | 4.51 × 10−1 | −1.94 | −9.9 | 6.03 | 9.79 × 10−1 |
| LPC 18:3 | 15.61 | 6.32 | 24.9 | 2.43 × 10−2 | 17.93 | −2.08 | 37.93 | 4.51 × 10−1 | −4 | −12.43 | 4.42 | 9.79 × 10−1 |
| LPC 22:5 | 14.71 | 6.14 | 23.27 | 2.23 × 10−2 | 10.66 | −8.81 | 30.14 | 4.51 × 10−1 | −1.24 | −9.23 | 6.74 | 9.79 × 10−1 |
| LPC 20:1 | 13.35 | 5.03 | 21.67 | 3.28 × 10−2 | 5.52 | −14.4 | 25.44 | 5.92 × 10−1 | 0.39 | −7.58 | 8.36 | 9.79 × 10−1 |
| LPE 18:2 | 13.35 | 5.3 | 21.4 | 2.66 × 10−2 | 7.78 | −11.87 | 27.43 | 4.64 × 10−1 | 0.41 | −7.53 | 8.34 | 9.79 × 10−1 |
| LPC 20:0p/20:1e | 13.32 | 5.49 | 21.15 | 2.31 × 10−2 | 12.88 | −6.67 | 32.42 | 4.51 × 10−1 | −1.58 | −9.61 | 6.45 | 9.79 × 10−1 |
| LPC 20:4 | 13.19 | 4.53 | 21.85 | 4.61 × 10−2 | 12.45 | −7.38 | 32.27 | 4.51 × 10−1 | −1.37 | −9.52 | 6.78 | 9.79 × 10−1 |
| TAG50:2 | −27.67 | −41.81 | −13.52 | 8.57 × 10−3 | 9.16 | −9.91 | 28.22 | 4.51 × 10−1 | −0.05 | −7.78 | 7.68 | 9.97 × 10−1 |
| TAG50:4 | −25.47 | −36.7 | −14.24 | 2.39 × 10−3 | 10.39 | −8.08 | 28.85 | 4.51 × 10−1 | 0.67 | −6.81 | 8.15 | 9.79 × 10−1 |
| TAG54:3 | −20.27 | −31.31 | −9.24 | 1.47 × 10−2 | 12.73 | −6.57 | 32.02 | 4.51 × 10−1 | −0.2 | −8.02 | 7.63 | 9.87 × 10−1 |
| TAG50:3 | −19.96 | −30.18 | −9.74 | 8.57 × 10−3 | 11.29 | −7.77 | 30.35 | 4.51 × 10−1 | 0.01 | −7.73 | 7.74 | 9.99 × 10−1 |
| TAG54:1 | −19.93 | −29.07 | −10.79 | 3.76 × 10−3 | 13.77 | −4.91 | 32.46 | 4.51 × 10−1 | −1.04 | −8.64 | 6.56 | 9.79 × 10−1 |
| TAG52:3 | −18.81 | −27 | −10.61 | 2.38 × 10−3 | 13.8 | −4.66 | 32.27 | 4.51 × 10−1 | −0.51 | −7.99 | 6.97 | 9.79 × 10−1 |
| TAG58:1 | −18.6 | −28.21 | −8.99 | 8.93 × 10−3 | 8.91 | −10.2 | 28.02 | 4.51 × 10−1 | 0.54 | −7.2 | 8.28 | 9.79 × 10−1 |
| TAG52:2 | −18.48 | −26.14 | −10.83 | 2.25 × 10−3 | 10.66 | −7.52 | 28.85 | 4.51 × 10−1 | −0.64 | −8.03 | 6.76 | 9.79 × 10−1 |
| TAG56:0 | −17.37 | −24.81 | −9.93 | 2.28 × 10−3 | 12.84 | −5.52 | 31.21 | 4.51 × 10−1 | −0.02 | −7.46 | 7.42 | 9.98 × 10−1 |
| TAG54:2 | −16.76 | −24.76 | −8.76 | 5.90 × 10−3 | 14.93 | −3.98 | 33.83 | 4.51 × 10−1 | −1.2 | −8.87 | 6.48 | 9.79 × 10−1 |
| TAG58:2 | −16.31 | −25.29 | −7.33 | 1.51 × 10−2 | 9 | −10.32 | 28.33 | 4.51 × 10−1 | 0.79 | −7.03 | 8.62 | 9.79 × 10−1 |
| TAG52:1 | −16.31 | −24.87 | −7.74 | 1.03 × 10−2 | 11.94 | −7.21 | 31.1 | 4.51 × 10−1 | −0.8 | −8.6 | 7.01 | 9.79 × 10−1 |
| TAG52:4 | −16.03 | −24.22 | −7.85 | 8.57 × 10−3 | 12.01 | −7.04 | 31.07 | 4.51 × 10−1 | 0.33 | −7.39 | 8.05 | 9.80 × 10−1 |
| TAG53:0 | −16.02 | −25.53 | −6.5 | 2.43 × 10−2 | 10.61 | −8.92 | 30.14 | 4.51 × 10−1 | −0.67 | −8.63 | 7.3 | 9.79 × 10−1 |
| TAG54:0 | −14.87 | −23.12 | −6.62 | 1.55 × 10−2 | 8.63 | −10.73 | 27.99 | 4.51 × 10−1 | 0.66 | −7.17 | 8.49 | 9.79 × 10−1 |
| TAG53:1 | −14.76 | −23.34 | −6.19 | 2.23 × 10−2 | 10.41 | −9.06 | 29.87 | 4.51 × 10−1 | 0.17 | −7.73 | 8.07 | 9.87 × 10−1 |
| TAG55:3 | −14.73 | −23.32 | −6.14 | 2.23 × 10−2 | 7.46 | −12.1 | 27.03 | 4.73 × 10−1 | 0.99 | −6.9 | 8.89 | 9.79 × 10−1 |
| TAG57:3 | −14.66 | −23.71 | −5.61 | 3.10 × 10−2 | 9.83 | −9.81 | 29.48 | 4.51 × 10−1 | 0.17 | −7.79 | 8.14 | 9.87 × 10−1 |
| TAG53:3 | −14.3 | −23.07 | −5.54 | 3.00 × 10−2 | 8.02 | −11.67 | 27.7 | 4.63 × 10−1 | 0.77 | −7.18 | 8.71 | 9.79 × 10−1 |
| TAG48:3 | −14.27 | −22.59 | −5.94 | 2.23 × 10−2 | 11.15 | −8.33 | 30.63 | 4.51 × 10−1 | 0.82 | −7.08 | 8.71 | 9.79 × 10−1 |
| TAG48:2 | −13.24 | −21.86 | −4.62 | 4.61 × 10−2 | 9.99 | −9.79 | 29.77 | 4.51 × 10−1 | 0.9 | −7.11 | 8.91 | 9.79 × 10−1 |
| TAG52:0 | −12.62 | −20.93 | −4.32 | 4.61 × 10−2 | 7.93 | −11.95 | 27.81 | 4.64 × 10−1 | 0.92 | −7.1 | 8.94 | 9.79 × 10−1 |
| TAG58:3 | −12.4 | −20.57 | −4.24 | 4.61 × 10−2 | 8.18 | −11.69 | 28.05 | 4.62 × 10−1 | 0.58 | −7.45 | 8.6 | 9.79 × 10−1 |
| TAG56:1 | −12.11 | −20.08 | −4.15 | 4.61 × 10−2 | 8.47 | −11.38 | 28.32 | 4.53 × 10−1 | 0.56 | −7.46 | 8.58 | 9.79 × 10−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, G.; Narasimhan, K.; Cheong, W.F.; Ng, S.; Aris, I.M.; Loy, S.L.; Bendt, A.K.; Tan, K.H.; Yap, F.K.P.; Shek, L.P.; et al. Umbilical Cord Plasma Lysophospholipids and Triacylglycerols Associated with Birthweight Percentiles. Nutrients 2024, 16, 274. https://doi.org/10.3390/nu16020274
Wong G, Narasimhan K, Cheong WF, Ng S, Aris IM, Loy SL, Bendt AK, Tan KH, Yap FKP, Shek LP, et al. Umbilical Cord Plasma Lysophospholipids and Triacylglycerols Associated with Birthweight Percentiles. Nutrients. 2024; 16(2):274. https://doi.org/10.3390/nu16020274
Chicago/Turabian StyleWong, Gerard, Kothandaraman Narasimhan, Wei Fun Cheong, Sharon Ng, Izzuddin M. Aris, See Ling Loy, Anne K. Bendt, Kok Hian Tan, Fabian K. P. Yap, Lynette P. Shek, and et al. 2024. "Umbilical Cord Plasma Lysophospholipids and Triacylglycerols Associated with Birthweight Percentiles" Nutrients 16, no. 2: 274. https://doi.org/10.3390/nu16020274
APA StyleWong, G., Narasimhan, K., Cheong, W. F., Ng, S., Aris, I. M., Loy, S. L., Bendt, A. K., Tan, K. H., Yap, F. K. P., Shek, L. P., Chong, Y. S., Gluckman, P. D., Godfrey, K. M., Lee, Y. S., Wenk, M. R., Karnani, N., & Chan, S.-Y. (2024). Umbilical Cord Plasma Lysophospholipids and Triacylglycerols Associated with Birthweight Percentiles. Nutrients, 16(2), 274. https://doi.org/10.3390/nu16020274