

Pain Relief, Functional Recovery, and Chondroprotective Effects of Angelica gigas Nakai in Osteoarthritis Due to Its Anti-Inflammatory Property: An In Vitro and In Vivo Study

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of AG Extract
2.2. High-Performance Liquid Chromatography (HPLC)
2.3. Animal
2.4. Preparation of Monoiodo-Acetate (MIA) Injection and Diet
2.5. Weight Bearing on the Hind Limb
2.6. Cartilage Degradation
2.7. Serum Concention Analysis of MIA Rats
2.8. Writhing Test
2.9. RAW264.7 Cell Culture
2.10. Cell Toxicity Measurement and Nitric Oxide (NO) Generation
2.11. Quantitative Real-Time Polymerase Chain Reaction
2.12. Protein Expression Analysis
2.13. Statistical Analysis
3. Results
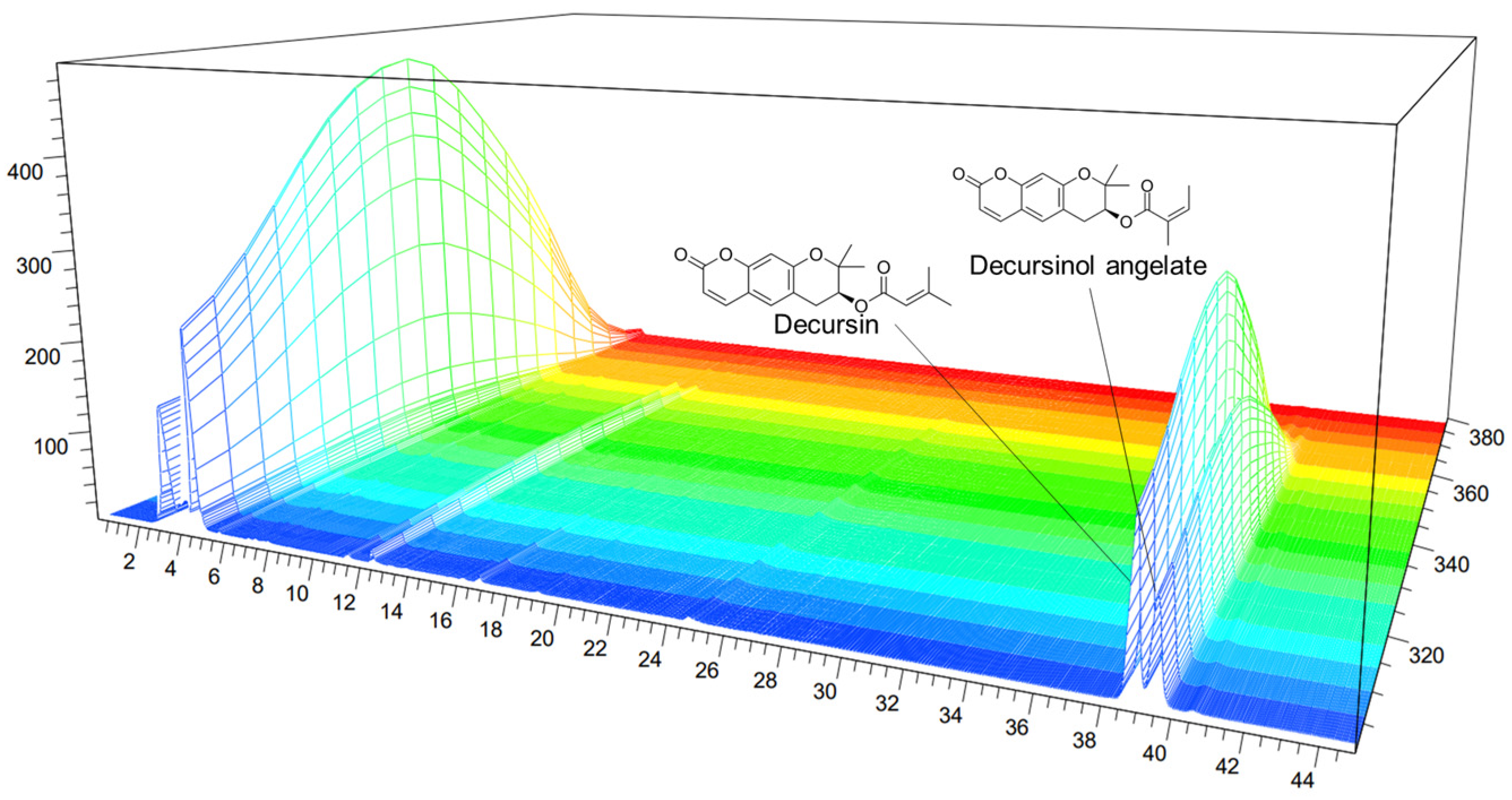
3.1. HPLC Analysis
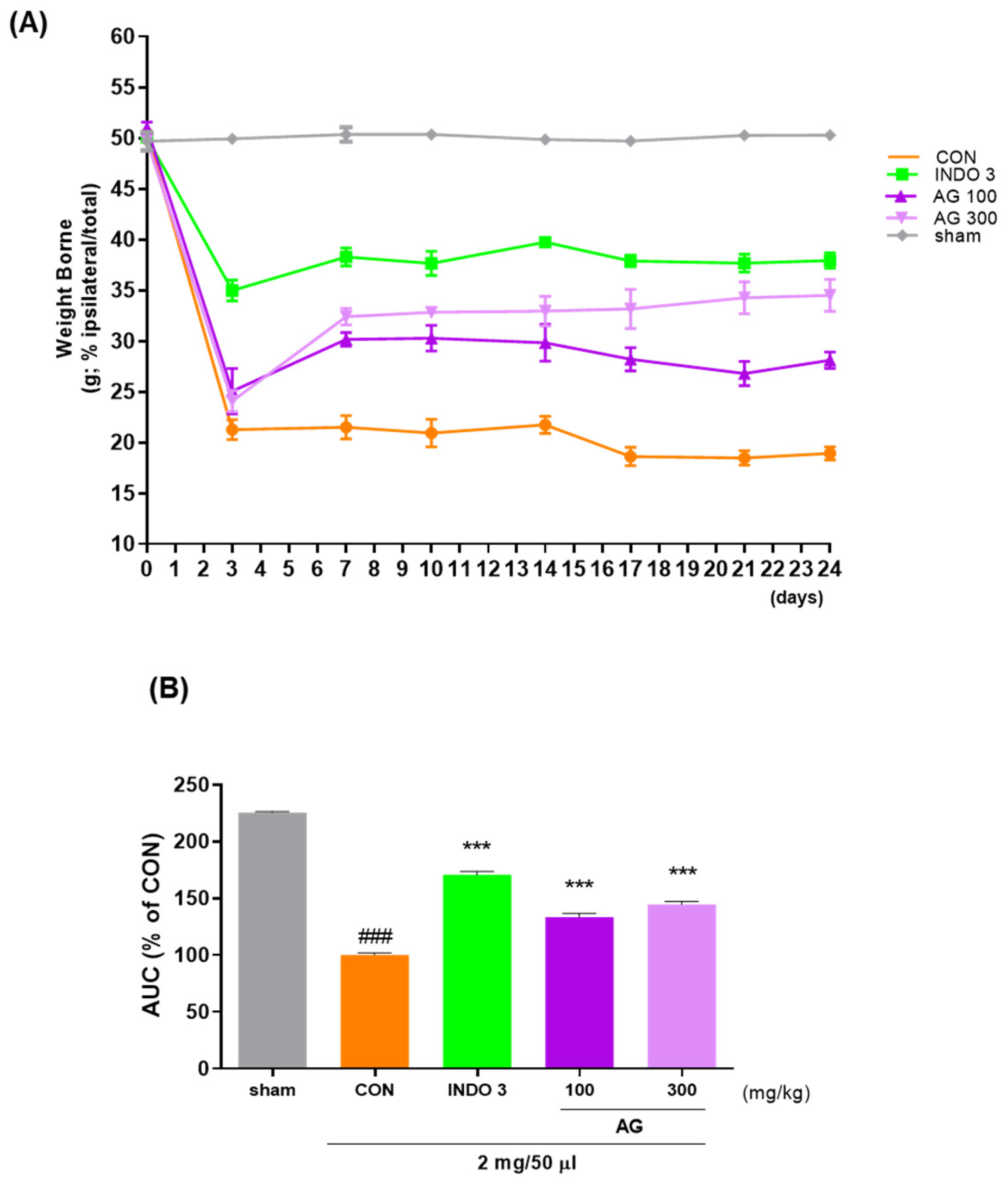
3.2. Evaluation of the Analgesic Effects Using MIA Animal Models
3.3. Cartilage Degradation in MIA Model
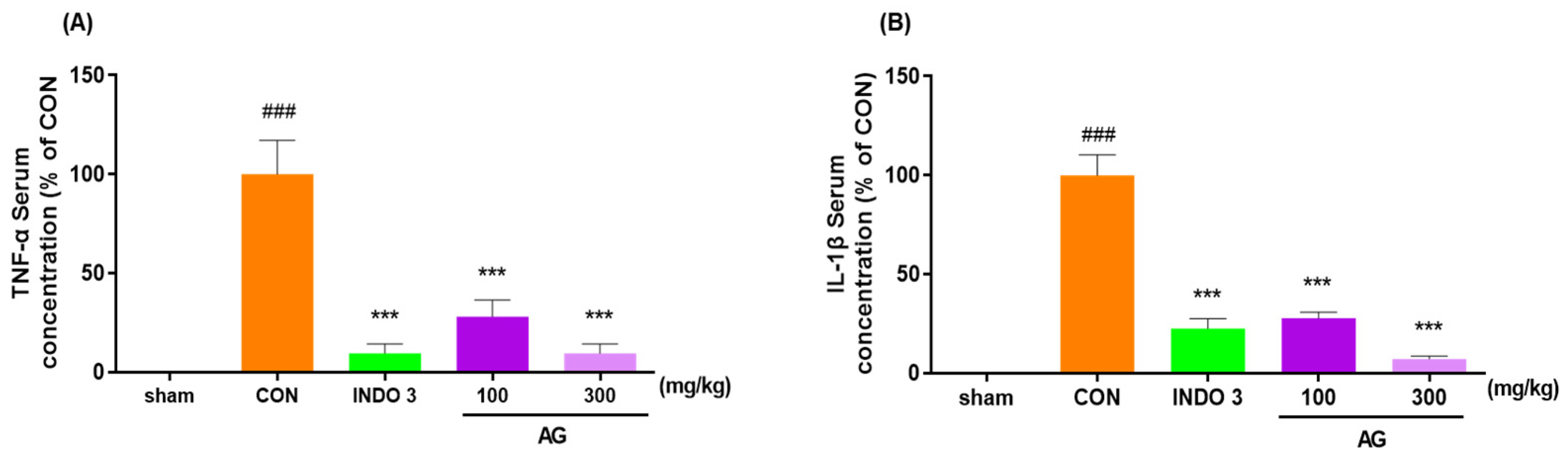
3.4. Serum Pro-Inflammatory Cytokine Levels in MIA Rats
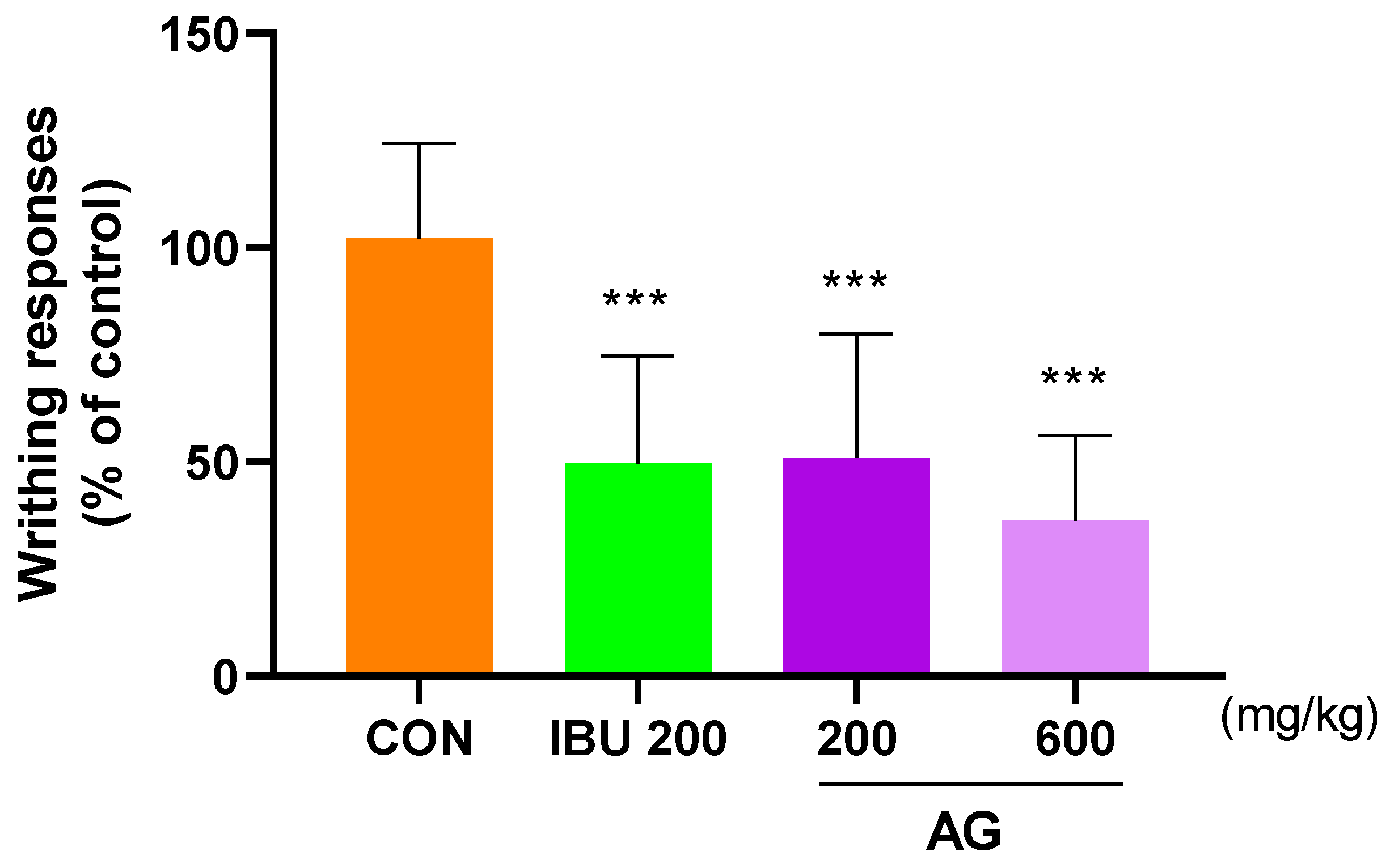
3.5. Analgesic Effects in the Acetic Acid-Induced Pain Animal Models
3.6. Cell Viability and NO Levels in RAW264.7 Cells
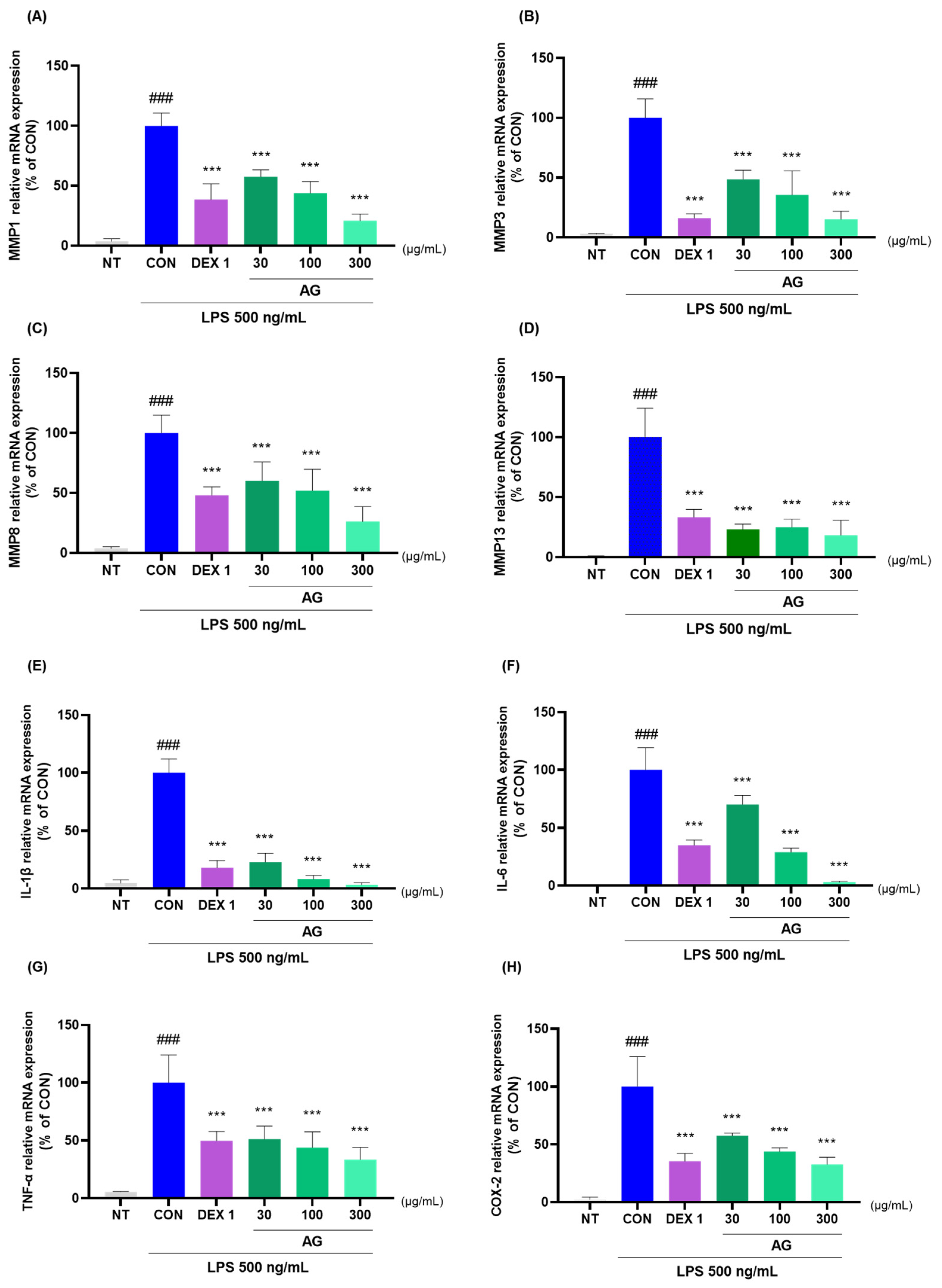
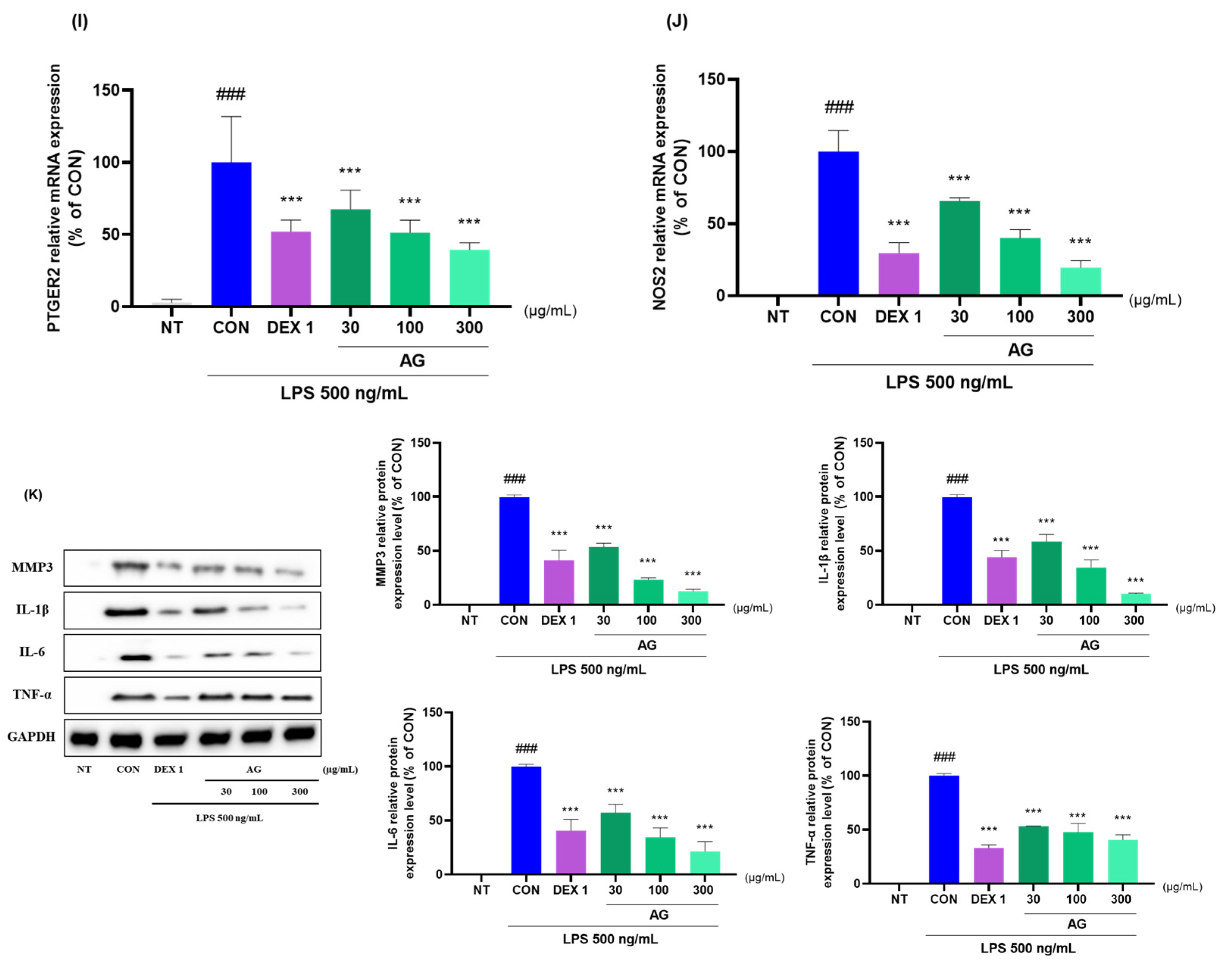
3.7. Anti-Inflammatory Effects of AG in LPS-Treated RAW264.7 Cells
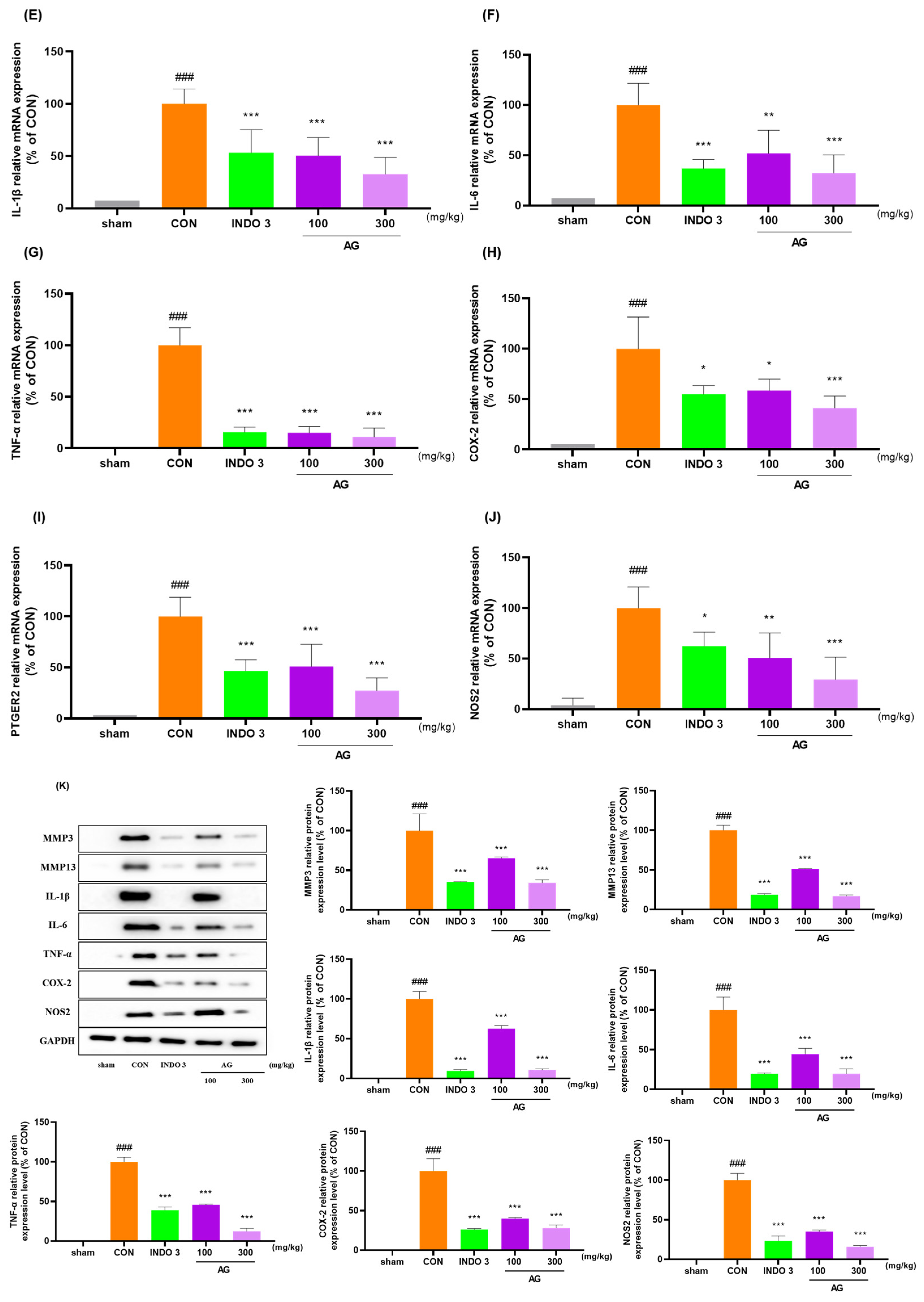
3.8. Effects on Cytokine Levels in Joint Cartilage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: Pathogenic Signaling Pathways and Therapeutic Targets. Signal Transduct. Target. Ther. 2023, 8, 56. [Google Scholar] [CrossRef]
- Muthu, S.; Korpershoek, J.V.; Novais, E.J.; Tawy, G.F.; Hollander, A.P.; Martin, I. Failure of Cartilage Regeneration: Emerging Hypotheses and Related Therapeutic Strategies. Nat. Rev. Rheumatol. 2023, 19, 403–416. [Google Scholar] [CrossRef]
- Scott, A.J.; Ellison, M.; Sinclair, D.A. The Economic Value of Targeting Aging. Nat. Aging 2021, 1, 616–623. [Google Scholar] [CrossRef]
- Tchkonia, T.; Palmer, A.K.; Kirkland, J.L. New Horizons: Novel Approaches to Enhance Healthspan Through Targeting Cellular Senescence and Related Aging Mechanisms. J. Clin. Endocrinol. Metab. 2021, 106, e1481–e1487. [Google Scholar] [CrossRef]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-Grade Inflammation as a Key Mediator of the Pathogenesis of Osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef]
- Chaib, S.; Tchkonia, T.; Kirkland, J.L. Cellular Senescence and Senolytics: The Path to the Clinic. Nat. Med. 2022, 28, 1556–1568. [Google Scholar] [CrossRef]
- Ansari, M.M.; Ghosh, M.; Lee, D.-S.; Son, Y.-O. Senolytic Therapeutics: An Emerging Treatment Modality for Osteoarthritis. Ageing Res. Rev. 2024, 96, 102275. [Google Scholar] [CrossRef]
- Motta, F.; Barone, E.; Sica, A.; Selmi, C. Inflammaging and Osteoarthritis. Clin. Rev. Allergy Immunol. 2023, 64, 222–238. [Google Scholar] [CrossRef]
- De Roover, A.; Escribano-Núñez, A.; Monteagudo, S.; Lories, R. Fundamentals of Osteoarthritis: Inflammatory Mediators in Osteoarthritis. Osteoarthr. Cartil. 2023, 31, 1303–1311. [Google Scholar] [CrossRef]
- Gibbs, A.J.; Gray, B.; Wallis, J.A.; Taylor, N.F.; Kemp, J.L.; Hunter, D.J.; Barton, C.J. Recommendations for the Management of Hip and Knee Osteoarthritis: A Systematic Review of Clinical Practice Guidelines. Osteoarthr. Cartil. 2023, 31, 1280–1292. [Google Scholar] [CrossRef]
- Richard, M.J.; Driban, J.B.; McAlindon, T.E. Pharmaceutical Treatment of Osteoarthritis. Osteoarthr. Cartil. 2023, 31, 458–466. [Google Scholar] [CrossRef]
- Deyle, G.D.; Allen, C.S.; Allison, S.C.; Gill, N.W.; Hando, B.R.; Petersen, E.J.; Dusenberry, D.I.; Rhon, D.I. Physical Therapy versus Glucocorticoid Injection for Osteoarthritis of the Knee. N. Engl. J. Med. 2020, 382, 1420–1429. [Google Scholar] [CrossRef]
- Mobasheri, A.; Loeser, R. Clinical Phenotypes, Molecular Endotypes and Theratypes in OA Therapeutic Development. Nat. Rev. Rheumatol. [CrossRef]
- Su, J.; Yu, M.; Wang, H.; Wei, Y. Natural Anti-Inflammatory Products for Osteoarthritis: From Molecular Mechanism to Drug Delivery Systems and Clinical Trials. Phytother. Res. PTR 2023, 37, 4321–4352. [Google Scholar] [CrossRef]
- Panossian, A.; Lemerond, T.; Efferth, T. State-of-the-Art Review on Botanical Hybrid Preparations in Phytomedicine and Phytotherapy Research: Background and Perspectives. Pharmaceuticals 2024, 17, 483. [Google Scholar] [CrossRef]
- Ren, J.-L.; Yang, L.; Qiu, S.; Zhang, A.-H.; Wang, X.-J. Efficacy Evaluation, Active Ingredients, and Multitarget Exploration of Herbal Medicine. Trends Endocrinol. Metab. TEM 2023, 34, 146–157. [Google Scholar] [CrossRef]
- Li, W.; Yu, L.; Li, W.; Ge, G.; Ma, Y.; Xiao, L.; Qiao, Y.; Huang, W.; Huang, W.; Wei, M.; et al. Prevention and Treatment of Inflammatory Arthritis with Traditional Chinese Medicine: Underlying Mechanisms Based on Cell and Molecular Targets. Ageing Res. Rev. 2023, 89, 101981. [Google Scholar] [CrossRef]
- Fang, S.; Zhang, B.; Xiang, W.; Zheng, L.; Wang, X.; Li, S.; Zhang, T.; Feng, D.; Gong, Y.; Wu, J.; et al. Natural Products in Osteoarthritis Treatment: Bridging Basic Research to Clinical Applications. Chin. Med. 2024, 19, 25. [Google Scholar] [CrossRef]
- Jo, H.-G.; Baek, C.-Y.; Song, H.S.; Lee, D. Network Pharmacology and Experimental Verifications to Discover Scutellaria Baicalensis Georgi’s Effects on Joint Inflammation, Destruction, and Pain in Osteoarthritis. Int. J. Mol. Sci. 2024, 25, 2127. [Google Scholar] [CrossRef]
- Jo, H.-G.; Baek, C.Y.; Lee, J.; Hwang, Y.; Baek, E.; Hwang, J.H.; Lee, D. Anti-Inflammatory, Analgesic, Functional Improvement, and Chondroprotective Effects of Erigeron breviscapus (Vant.) Hand.-Mazz. Extract in Osteoarthritis: An In Vivo and In Vitro Study. Nutrients 2024, 16, 1035. [Google Scholar] [CrossRef]
- Jo, H.G.; Baek, C.Y.; Kim, D.; Kim, S.; Han, Y.; Park, C.; Song, H.S.; Lee, D. Network Analysis, in Vivo, and in Vitro Experiments Identified the Mechanisms by Which Piper longum L. [Piperaceae] Alleviates Cartilage Destruction, Joint Inflammation, and Arthritic Pain. Front. Pharmacol. 2023, 14, 1282943. [Google Scholar] [CrossRef]
- Jo, H.-G.; Baek, C.Y.; Kim, D.; Lee, D.; Song, H.S. Stem of Sorbus Commixta Hedl. Extract Inhibits Cartilage Degradation and Arthritic Pain in Experimental Model via Anti-Inflammatory Activity. Nutrients 2023, 15, 3774. [Google Scholar] [CrossRef]
- Jo, H.-G.; Lee, G.-Y.; Baek, C.Y.; Song, H.S.; Lee, D. Analgesic and Anti-Inflammatory Effects of Aucklandia Lappa Root Extracts on Acetic Acid-Induced Writhing in Mice and Monosodium Iodoacetate-Induced Osteoarthritis in Rats. Plants 2020, 10, 42. [Google Scholar] [CrossRef]
- Lü, J.; Jiang, C.; Schell, T.D.; Joshi, M.; Raman, J.D.; Xing, C. Angelica gigas: Signature Compounds, In Vivo Anticancer, Analgesic, Neuroprotective and Other Activities, and the Clinical Translation Challenges. Am. J. Chin. Med. 2022, 50, 1475–1527. [Google Scholar] [CrossRef]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE Guidelines 2.0: Updated Guidelines for Reporting Animal Research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- He, L.; Pan, Y.; Yu, J.; Wang, B.; Dai, G.; Ying, X. Decursin Alleviates the Aggravation of Osteoarthritis via Inhibiting PI3K-Akt and NF-kB Signal Pathway. Int. Immunopharmacol. 2021, 97, 107657. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, T.; Kang, J.-H.; Li, H.; Cho, H.; Jeon, R.; Ryu, J.-H.; Yim, M. Decursin from Angelica gigas Suppresses RANKL-Induced Osteoclast Formation and Bone Loss. Eur. J. Pharmacol. 2016, 774, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Shehzad, A.; Parveen, S.; Qureshi, M.; Subhan, F.; Lee, Y.S. Decursin and Decursinol Angelate: Molecular Mechanism and Therapeutic Potential in Inflammatory Diseases. Inflamm. Res. 2018, 67, 209–218. [Google Scholar] [CrossRef]
- Islam, S.U.; Lee, J.H.; Shehzad, A.; Ahn, E.-M.; Lee, Y.M.; Lee, Y.S. Decursinol Angelate Inhibits LPS-Induced Macrophage Polarization through Modulation of the NFκB and MAPK Signaling Pathways. Molecules 2018, 23, 1880. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Hale, T.W.; Chee, W.; Xing, C.; Jiang, C.; Lü, J. Single Oral Dose Pharmacokinetics of Decursin and Decursinol Angelate in Healthy Adult Men and Women. PLoS ONE 2015, 10, e0114992. [Google Scholar] [CrossRef] [PubMed]
- Jenei-Lanzl, Z.; Meurer, A.; Zaucke, F. Interleukin-1β Signaling in Osteoarthritis—Chondrocytes in Focus. Cell Signal. 2019, 53, 212–223. [Google Scholar] [CrossRef]
- Hosseinzadeh, A.; Kamrava, S.K.; Joghataei, M.T.; Darabi, R.; Shakeri-Zadeh, A.; Shahriari, M.; Reiter, R.J.; Ghaznavi, H.; Mehrzadi, S. Apoptosis Signaling Pathways in Osteoarthritis and Possible Protective Role of Melatonin. J. Pineal Res. 2016, 61, 411–425. [Google Scholar] [CrossRef]
- van Helvoort, E.M.; Welsing, P.M.J.; Jansen, M.P.; Gielis, W.P.; Loef, M.; Kloppenburg, M.; Blanco, F.; Haugen, I.K.; Berenbaum, F.; Bay-Jensen, A.-C.; et al. Neuropathic Pain in the IMI-APPROACH Knee Osteoarthritis Cohort: Prevalence and Phenotyping. RMD Open 2021, 7, e002025. [Google Scholar] [CrossRef]
- Bjurström, M.F.; Blennow, K.; Zetterberg, H.; Bodelsson, M.; Waldén, M.; Dietz, N.; Hall, S.; Hansson, O.; Irwin, M.R.; Mattsson-Carlgren, N. Central Nervous System Monoaminergic Activity in Hip Osteoarthritis Patients with Disabling Pain: Associations with Pain Severity and Central Sensitization. Pain Rep. 2022, 7, e988. [Google Scholar] [CrossRef] [PubMed]
- Grillet, B.; Pereira, R.V.S.; Van Damme, J.; Abu El-Asrar, A.; Proost, P.; Opdenakker, G. Matrix Metalloproteinases in Arthritis: Towards Precision Medicine. Nat. Rev. Rheumatol. 2023, 19, 363–377. [Google Scholar] [CrossRef]
- Diekman, B.O.; Loeser, R.F. Aging and the Emerging Role of Cellular Senescence in Osteoarthritis. Osteoarthr. Cartil. 2024, 32, 365–371. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | |
|---|---|
| Colum | Luna C18 column (5 μm, 250 mm × 4.6 mm; Phenomenex, Torrance, CA, USA) |
| Mobile phase | (A) DW, (B) acetonitrile |
| Flow rate | 0–5 min, 35–35%; 5–10 min, 35–50%; 10–30 min, 50–55%; 30–40 min, 55–70%; 40–45 min, 70–35% (B) |
| Injection volume | 1.0 mL/min |
| Detection wavelength | 330 nm |
| Temperature | 30 °C |
| Group Name | OA Model (50 μL, Intra-Articular; mg/mL) | Diet (AIN-93G) | Final Concentration (mg/kg) |
|---|---|---|---|
| sham | Saline | - | - |
| CON | MIA 40 | - | - |
| INDO 3 | MIA 40 | +0.003% indomethacin | indomethacin 3 |
| AG 100 | MIA 40 | +0.11% AG | AG 100 |
| AG 300 | MIA 40 | +0.33% AG | AG 300 |
| Grade | Cartilage Appearance |
|---|---|
| 0 | A typical representation of the cartilage surface |
| 1 | Modest yellowish discoloration or modest fibrillation |
| 2 | The middle or superficial layers of cartilage are affected by erosion |
| 3 | Severe deterioration that reaches the subchondral bone |
| 4 | Large-scale erosion and extensive exposure of subchondral bone |
| MMP-1 | F | AAC TTG GGT GAA GAC GTC CA |
| R | TCC TGT CAC TTT CAG CCC AA | |
| MMP-3 | F | GTA CGG CTG TGT GCT CAT CC |
| R | TCA GCC CAA GGA ACT TCT GC | |
| MMP-8 | F | TCT GTT CTT CTT CCA CAC ACA G |
| R | GCA ATC ATA GTG GCA TTC CT | |
| MMP-13 | F | ACC TTC TTC TTG TTG AG TTG GA |
| R | CTG CAT TTC TCG GAG TCT A | |
| IL-1β | F | AAC TCA ACT GTG AAA TAG CAG C |
| R | TCC ACA GCC ACA ATG AGT G | |
| IL-6 | F | TCC GCA AGA GAC TTC CAG C |
| R | CCT CCG ACT TGT GAA GTG G | |
| TNF-α | F | GCA TGA TCC GAG ATG TGG AA |
| R | GAT GAG AGG GAG CCC ATT TG | |
| COX-2 | F | GTT CCA ACC CAT GTC AAA AC |
| R | TGT CAG GAA TCT CGG CGT AG | |
| Ptger2 | F | TGT GTG TAC TGT CCG TCT GC |
| R | CAG GGA TCC AGT CTC GGT GT | |
| NOS2 | F | AGT CAA CTA CAA GCC CCA CG |
| R | GCA GCT TGT CCA GGG ATT CT | |
| GAPDH | F | CTT GTG ACA AAG TGG ACA TTG TT |
| R | TGA CCA GCT TCC CAT TCT C |
| MMP-1 | F | ATG CCT AGC CTT CCT TTG CT |
| R | TTC CAG GTA TTT CCA GAC TG | |
| MMP-3 | F | AAG TTC CTC GGG TTG GAG AT |
| R | ACC AAC ATC AGG AAC ACC AC | |
| MMP-8 | F | CAA TCA ATT CCG GTC TTC GA |
| R | GGT TAG CAA GAA ATC ACC AGA | |
| MMP-13 | F | AAC CAA GAT GTG GAG TGC CT |
| R | GAC CAG ACC TTG AAG GCT TT | |
| IL-1β | F | CCA GCT TCA AAT CTC GCA GC |
| R | GTG CTC ATG TCC TCA TCC TGG | |
| IL-6 | F | CAC TTC ACA AGT CGG AGG CT |
| R | CAA GTG CAT CAT CGT TGT TC | |
| TNF-α | F | GAG AAG TTC CCA AAT GGC CT |
| R | AGC CAC TCC AGC TGC TCC T | |
| COX-2 | F | ATC CAT GTC AAA ACC GTG GG |
| R | TTG GGG TGG GCT TCA GCA G | |
| Ptger2 | F | CTG GTA ACG GAA TTG GTG C |
| R | TGG CCA GAC TAA AGA AGG TC | |
| NOS2 | F | ACC AAG ATG GCC TGG AGG AA |
| R | CCG ACC TGA TGT TGC CAT TG | |
| GAPDH | F | ATG GTG AAG GTC GGT GTG |
| R | GCC GTG AGT GGA GTC ATA C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, H.-G.; Baek, C.Y.; Hwang, Y.; Baek, E.; Song, H.S.; Lee, D. Pain Relief, Functional Recovery, and Chondroprotective Effects of Angelica gigas Nakai in Osteoarthritis Due to Its Anti-Inflammatory Property: An In Vitro and In Vivo Study. Nutrients 2024, 16, 2435. https://doi.org/10.3390/nu16152435
Jo H-G, Baek CY, Hwang Y, Baek E, Song HS, Lee D. Pain Relief, Functional Recovery, and Chondroprotective Effects of Angelica gigas Nakai in Osteoarthritis Due to Its Anti-Inflammatory Property: An In Vitro and In Vivo Study. Nutrients. 2024; 16(15):2435. https://doi.org/10.3390/nu16152435
Chicago/Turabian StyleJo, Hee-Geun, Chae Yun Baek, Yeseul Hwang, Eunhye Baek, Ho Sueb Song, and Donghun Lee. 2024. "Pain Relief, Functional Recovery, and Chondroprotective Effects of Angelica gigas Nakai in Osteoarthritis Due to Its Anti-Inflammatory Property: An In Vitro and In Vivo Study" Nutrients 16, no. 15: 2435. https://doi.org/10.3390/nu16152435
APA StyleJo, H.-G., Baek, C. Y., Hwang, Y., Baek, E., Song, H. S., & Lee, D. (2024). Pain Relief, Functional Recovery, and Chondroprotective Effects of Angelica gigas Nakai in Osteoarthritis Due to Its Anti-Inflammatory Property: An In Vitro and In Vivo Study. Nutrients, 16(15), 2435. https://doi.org/10.3390/nu16152435