


Postbiotics as Adjuvant Therapy in Cancer Care

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
3. Prebiotics, Probiotics, and Postbiotics: An Overview
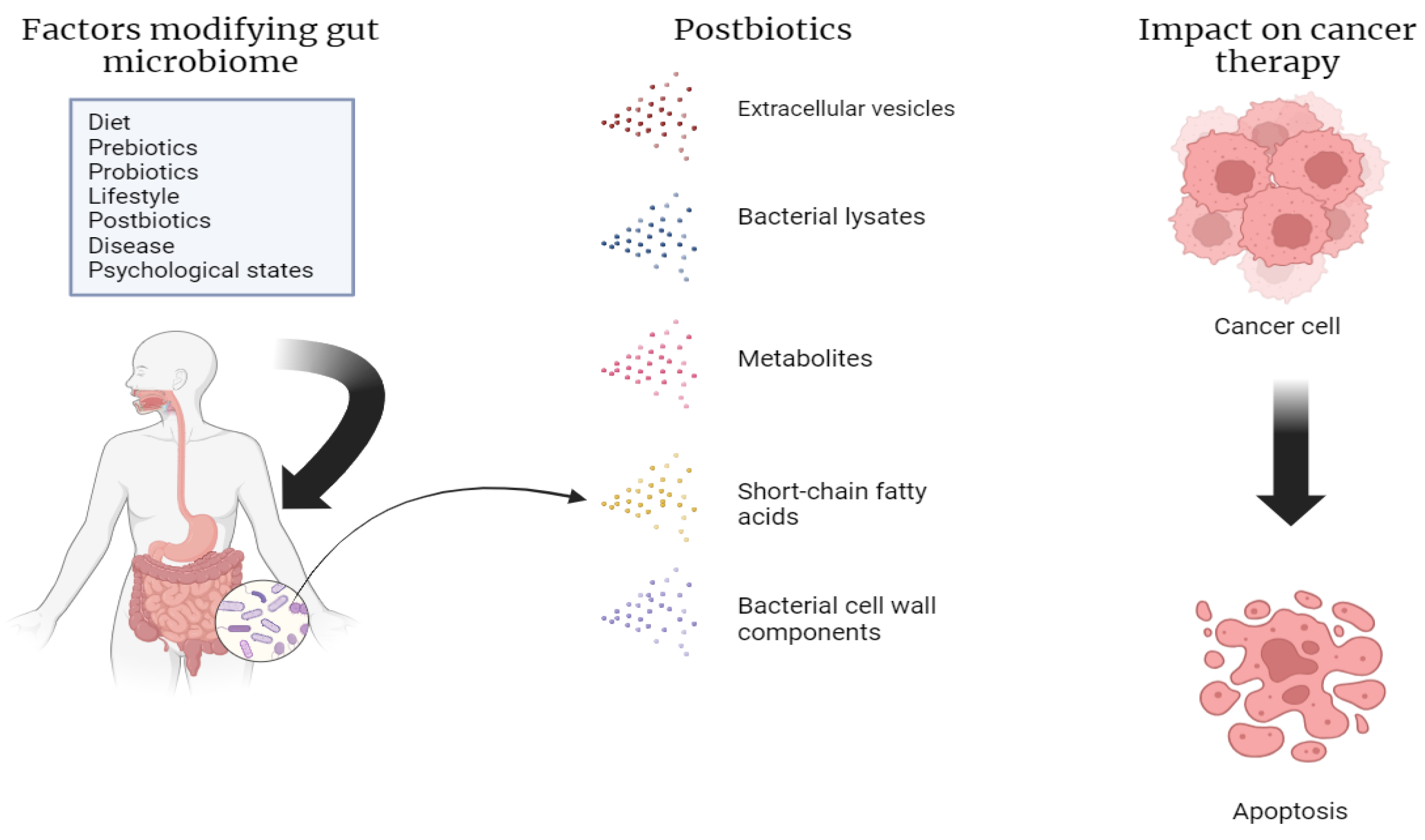
4. Types of Postbiotics
5. Role of Postbiotics in Gut Microbiotic Health and Cancer Microbiotic Health
5.1. Colorectal Cancer
5.2. Breast Cancer
5.3. Gastric Cancer
5.4. Cervical Cancer
5.5. Leukemia
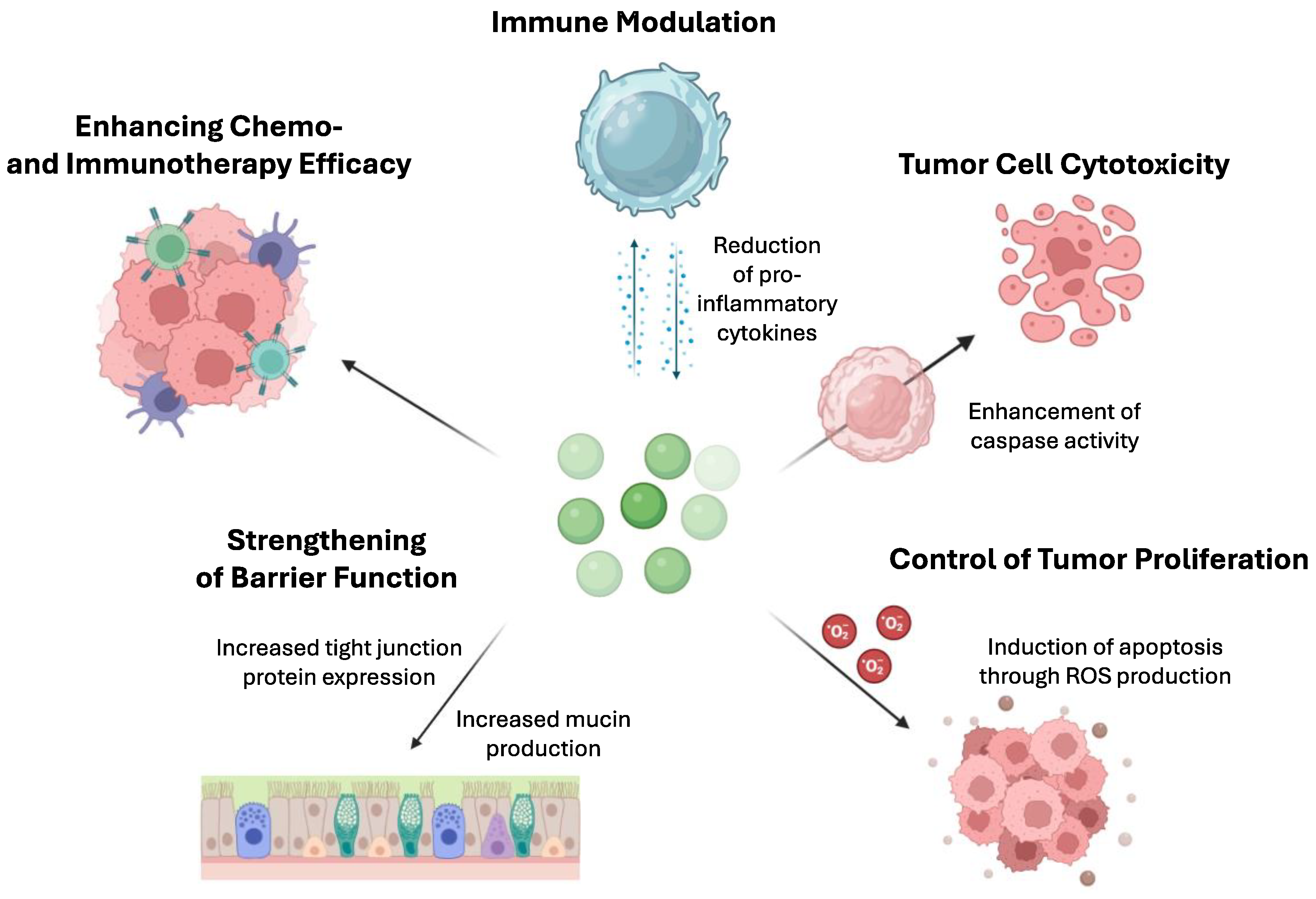
6. Mechanism of Action of Postbiotics in Cancer
7. Prebiotics, Postbiotics, and Purified Macromolecules for Cancer Care—Preclinical and Clinical Studies

{kind=link}
{kind=link}
| Intervention | Type of Study | Type of Cancer | Median Age (IQR) | Sample Size (n. Events) | Median Follow Up | Antitumoral Benefit | |
|---|---|---|---|---|---|---|---|
| Boutron-Ruault MC et al., 2005 [100] | Supplementation with s-FOS | Interventional prospective study | - Small adenomas - Large adenomas - Healthy controls | 61 (8) | 74 | - | Higher fecal butyrate concentration in the adenoma group after the 3-month administration of sc-FOS |
| Chen HM et al., 2013 [99] | None | Cross-sectional observational study | Patients with a resected AP vs. healthy controls | 58 (11) | 391 | - | Lower SCFA levels were found in the AP group Clostridium, Roseburia and Eubacterium spp were retrieved at higher levels in the healthy controls Enterococcus and Streptococcus spp. were more highly represented in the AP group |
| Motoori M et al., 2017 [107] | Synbiotic supplementation 10 days after chemotherapy | Phase II, randomized open-label study | Esophageal cancer | - | 61 | - | Decrease in toxicity incidence (nausea, diarrea and febrile neutropenia) |
| Hoyt M et al., 2019 [93] | Menokinones, phylloquinone | post hoc Observational study | Prostate cancer | 63 (6) | 28,356 (2978) | 11.3 months | No benefits as risk-reducing factor |
| Niccolai E et al., 2019 [96] | None | Cross-sectional, controlled, nonrandomized, observational study | Colorectal cancer vs. AP and healthy controls | CRC: 80 (13) AP: 46 (8) | 60 | - | CRC patients showed increased levels of butyric isobutyric, valeric and isovaleric acid, whereas the levels of acetic acid were reduced |
| Xie X et al., 2019 [106] | Supplementation with 30 mg/d of prebiotics (fiber) aiming to raise intestinal SCFAs | Interventional prospective study | Colon cancer development | 60 (9) | 135 | - | Preoperative period: higher levels of IgM, IgG and transferrin Postoperative period: higher levels of IgA, IgG, CD8+ Cells, B-cell lymphocytes |
| Ocvirk. S et al., 2020 [98] | None | Cross-sectional observation study | Apparently healthy patients | 51 (8) | 53 (AN: 32 RA: 21) | - | AN ate more fatty and caloric food than RA In AN, 16 out of 32 patients had colic adenomatous polypolsis, whereas none of RA developed polyposis. Stools from RA were more enriched by SCFAs than AN stools. |
| Haruna Y et al., 2021 [94] | Vitamin K + sorafenib vs. sorafenib alone | Phase 2, randomized, open-label study | Hepatocarcinoma | 72 (8) | 44 (44) | 70 months | Benefit for ORR (27.3% vs. 4.5%, p = 0.039) PFS (HR = 0.59, p = 0.12) No benefit for OS (HR = 0.59, p = 0.12) |
| Seethaler B et al., 2022 [95] | Increase in SCFA production through diet and physical activity | Phase 2, randomized, controlled, open label study | BRCA-mutated patients | 44 (2) | 260 women | - | Increase in level of fecal SCFAs produced Decrease in intestinal permeability mediated by SCFAs |
| Motoori M et al., 2017 [111] | Synbiotic supplementation in addition to enteral nutrition and prophylactic antibiotics during neoadjuvant chemotherapy | Phase II, randomized open-label multicenter study | Esophageal cancer | - | 81 | - | Signficiant decrease in grade 3 and grade 4 toxicity incidence (nausea, diarrea and neutropenia) but not febriel neutropenia (p = 0.088) |
| Liu L et al., 2023 [103] | Supplementation with either 50 or 100 mg/d of Riboflavin for 2 weeks | Interventional prospective study | Colon cancer development | 31 (11) | 105 | 28 days | Higher fecal butyrate level with riboflavin supplemantetion regardless of the given dose |
8. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Vemuri, R.; Shankar, E.M.; Chieppa, M.; Eri, R.; Kavanagh, K. Beyond Just Bacteria: Functional Biomes in the Gut Ecosystem Including Virome, Mycobiome, Archaeome and Helminths. Microorganisms 2020, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Temraz, S.; Nassar, F.; Nasr, R.; Charafeddine, M.; Mukherji, D.; Shamseddine, A. Gut Microbiome: A Promising Biomarker for Immunotherapy in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 4155. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Gieryńska, M.; Szulc-Dąbrowska, L.; Struzik, J.; Mielcarska, M.B.; Gregorczyk-Zboroch, K.P. Integrity of the Intestinal Barrier: The Involvement of Epithelial Cells and Microbiota-A Mutual Relationship. Animals 2022, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef] [PubMed]
- Durack, J.; Lynch, S.V. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [PubMed]
- Hasan, N.; Yang, H. Factors affecting the composition of the gut microbiota, and its modulation. PeerJ 2019, 7, e7502. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The concept of postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef]
- Mehta, J.P.; Ayakar, S.; Singhal, R.S. The potential of paraprobiotics and postbiotics to modulate the immune system: A Review. Microbiol. Res. 2023, 275, 127449. [Google Scholar] [CrossRef]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics-A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef]
- Liang, B.; Xing, D. The Current and Future Perspectives of Postbiotics. Probiotics Antimicrob. Proteins 2023, 15, 1626–1643. [Google Scholar] [CrossRef]
- Homayouni Rad, A.; Aghebati Maleki, L.; Samadi Kafil, H.; Fathi Zavoshti, H.; Abbasi, A. Postbiotics as Promising Tools for Cancer Adjuvant Therapy. Adv. Pharm. Bull. 2021, 11, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Verma, M.K.; Chauhan, N.S. A review of metabolic potential of human gut microbiome in human nutrition. Arch. Microbiol. 2018, 200, 203–217. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D. Expert consensus document: The international scientific association for probiotics and prebiotics (ISAPP) Consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Peredo-Lovillo, A.; Romero-Luna, H.E.; Jiménez-Fernández, M. Health Promoting microbial metabolites produced by gut microbiota after prebiotics metabolism. Food Res. Int. 2020, 136, 109473. [Google Scholar] [CrossRef]
- Manzoor, S.; Wani, S.M.; Rizwan, D. Role of probiotics and prebiotics in mitigation of different diseases. Nutrition 2022, 96, 111602. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the manipulation of bacteria–gut–brain signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Mansueto, P.; Seidita, A.; D’Alcamo, A.; Carroccio, A. Role of FODMAPs in Patients with Irritable Bowel Syndrome. Nutr. Clin. Pract. 2015, 30, 665–682. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Franz, C.M.A.P.; Huch, M.; Abriouel, H.; Holzapfel, W.; Gálvez, A. Enterococci as probiotics and their implications in food safety. Int. J. Food Microbiol. 2011, 151, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Jin, W.; Liu, S.J.; Jiao, Z.; Li, X. Probiotics, prebiotics, and postbiotics in health and disease. MedComm 2023, 4, e420. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Sheng, J.; Wang, M.; Luo, H.; Zhu, J.; Zhang, B.; Liu, Z.; Yang, X. Combination therapy of TGF-β blockade and commensalderived probiotics provides enhanced antitumor immune response and tumor suppression. Theranostics 2019, 9, 4115–4129. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A novel class of psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Oak, S.J.; Jha, R. The effects of probiotics in lactose intolerance: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1675–1683. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H. The international scientific association of probiotics and prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- George Kerry, R.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.-S.; Das, G. Benefaction of probiotics for human health: A review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, S.; Li, S.; Jiang, W.; Wang, J.; Xiao, J.; Chen, T.; Ma, J.; Khan, M.Z.; Wang, W. Unlocking the power of postbiotics: A revolutionary approach to nutrition for humans and animals. Cell Metab. 2024, 36, 725–744. [Google Scholar] [CrossRef]
- Riwes, M.; Reddy, P. Short chain fatty acids: Postbiotics/metabolites and graft versus host disease colitis. Semin. Hematol. 2020, 57, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mansuy-Aubert, V.; Ravussin, Y. Short chain fatty acids: The messengers from down below. Front. Neurosci. 2023, 17, 1197759. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.T.; Cresci, G.A.M. The Immunomodulatory Functions of Butyrate. J. Inflamm. Res. 2021, 14, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Chriett, S.; Dąbek, A.; Wojtala, M.; Vidal, H.; Balcerczyk, A.; Pirola, L. Prominent action of butyrate over β-hydroxybutyrate as histone deacetylase inhibitor, transcriptional modulator and anti-inflammatory molecule. Sci. Rep. 2019, 9, 742. [Google Scholar] [CrossRef]
- Bertani, B.; Ruiz, N. Function and Biogenesis of Lipopolysaccharides. EcoSal Plus 2018, 8, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Szentirmai, É.; Massie, A.R.; Kapás, L. Lipoteichoic acid, a cell wall component of Gram-positive bacteria, induces sleep and fever and suppresses feeding. Brain Behav. Immun. 2021, 92, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Shida, K.; Kiyoshima-Shibata, J.; Kaji, R.; Nagaoka, M.; Nanno, M. Peptidoglycan from lactobacilli inhibits interleukin-12 production by macrophages induced by Lactobacillus casei through Toll-like receptor 2-dependent and independent mechanisms. Immunology 2009, 128 (Suppl. S1), e858–e869. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, X.; Zhang, R.; Petitte, J.N.; Si, D.; Li, Z.; Cheng, J.; Du, M. Design and Development of a Novel Peptide for Treating Intestinal Inflammation. Front. Immunol. 2019, 10, 1841. [Google Scholar] [CrossRef] [PubMed]
- Vinderola, G.; Sanders, M.E.; Salminen, S.; Szajewska, H. Postbiotics: The concept and their use in healthy populations. Front. Nutr. 2022, 9, 1002213. [Google Scholar] [CrossRef]
- Martini, G.R.; Tikhonova, E.; Rosati, E.; DeCelie, M.B.; Sievers, L.K.; Tran, F.; Lessing, M.; Bergfeld, A.; Hinz, S.; Nikolaus, S. Selection of cross-reactive T cells by commensal and food-derived yeasts drives cytotoxic TH1 cell responses in Crohn’s disease. Nat. Med. 2023, 29, 2602–2614. [Google Scholar] [CrossRef]
- Triantafillou, V.; Workman, A.D.; Patel, N.N.; Maina, I.W.; Tong, C.C.L.; Kuan, E.C.; Kennedy, D.W.; Palmer, J.N.; Adappa, N.D.; Waizel-Haiat, S.; et al. Broncho-Vaxom® (OM-85 BV) soluble components stimulate sinonasal innate immunity. Int. Forum Allergy Rhinol. 2019, 9, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.S.; Thirumdas, R.; Mehwish, H.M.; Umair, M.; Khurshid, M.; Hayat, H.F.; Phimolsiripol, Y.; Pallarés, N.; Martí-Quijal, F.J.; Barba, F.J. Role of Food Antioxidants in Modulating Gut Microbial Communities: Novel Understandings in Intestinal Oxidative Stress Damage and Their Impact on Host Health. Antioxidants 2021, 10, 1563. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, I.H.R.; Abbasi, F.; Wang, L.; Abd El Hack, M.E.; Swelum, A.A.; Hao, R.; Yao, J.; Cao, Y. Folate promotes S-adenosyl methionine reactions and the microbial methylation cycle and boosts ruminants production and reproduction. AMB Express 2018, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Chiu, S.W.; Hong, K.S.; Saver, J.L.; Wu, Y.L.; Lee, J.D.; Lee, M.; Ovbiagele, B. Folic Acid in Stroke Prevention in Countries without Mandatory Folic Acid Food Fortification: A Meta-Analysis of Randomized Controlled Trials. J. Stroke 2018, 20, 99–109. [Google Scholar] [CrossRef]
- Song, D.; Wang, X.; Ma, Y.; Liu, N.N.; Wang, H. Beneficial insights into postbiotics against colorectal cancer. Front. Nutr. 2023, 10, 1111872. [Google Scholar] [CrossRef]
- Kullisaar, T.; Zilmer, M.; Mikelsaar, M.; Vihalemm, T.; Annuk, H.; Kairane, C.; Kilk, A. Two antioxidative lactobacilli strains as promising probiotics. Int. J. Food Microbiol. 2002, 72, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, M.A. Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology 2022, 11, 804. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Panda, M.; Baral, B.; Varshney, N.R.S.; Bhandari, V.; Parmar, H.S.; Prasad, A.; Jha, H.C. Outer Membrane Vesicles: An Emerging Vaccine Platform. Vaccines 2022, 10, 1578. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, Q.; Wang, S.; Weng, W.; Jing, Y.; Su, J. Bacterial extracellular vesicles as bioactive nanocarriers for drug delivery: Advances and perspectives. Bioact. Mater. 2021, 14, 169–181. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. The Role of Probiotics in Colorectal Cancer Management. Evid.-Based Complement. Altern. Med. 2020, 2020, 3535982. [Google Scholar] [CrossRef]
- Pasquereau-Kotula, E.; Martins, M.; Aymeric, L.; Dramsi, S. Significance of Streptococcus gallolyticus subsp. gallolyticus Association with Colorectal Cancer. Front. Microbiol. 2018, 9, 614. [Google Scholar]
- Jastrząb, R.; Graczyk, D.; Siedlecki, P. Molecular and Cellular Mechanisms Influenced by Postbiotics. Int. J. Mol. Sci. 2021, 22, 13475. [Google Scholar] [CrossRef] [PubMed]
- Bourebaba, Y.; Marycz, K.; Mularczyk, M.; Bourebaba, L. Postbiotics as potential new therapeutic agents for metabolic disorders management. Biomed. Pharmacother. 2022, 153, 113–138. [Google Scholar] [CrossRef]
- Lou, X.; Xue, J.; Shao, R.; Mo, C.; Wang, F.; Chen, G. Postbiotics as potential new therapeutic agents for sepsis. Burn. Trauma 2023, 11, tkad022. [Google Scholar] [CrossRef] [PubMed]
- Smet, A.; Kupcinskas, J.; Link, A.; Hold, G.L.; Bornschein, J. The Role of Microbiota in Gastrointestinal Cancer and Cancer Treatment: Chance or Curse? Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 857–874. [Google Scholar] [CrossRef]
- Al-Hilu, S.A.; Al-Shujairi, W.H. Dual Role of Bacteria in Carcinoma: Stimulation and Inhibition. Int. J. Microbiol. 2020, 2020, 4639761. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Lai, C.H.; Tsai, W.H.; Chen, K.W.; Peng, S.L.; Lin, J.H.; Lin, Y.H. Nanoparticle-enhanced postbiotics: Revolutionizing cancer therapy through effective delivery. Life Sci. 2024, 337, 122379. [Google Scholar] [CrossRef]
- Xie, W.; Zhong, Y.S.; Li, X.J.; Kang, Y.K.; Peng, Q.Y.; Ying, H.Z. Postbiotics in colorectal cancer: Intervention mechanisms and perspectives. Front. Microbiol. 2024, 15, 1360225. [Google Scholar] [CrossRef]
- German, R.; Marino, N.; Hemmerich, C.; Podicheti, R.; Rusch, D.B.; Stiemsma, L.T.; Gao, H.; Xuei, X.; Rockey, P.; Storniolo, A.M. Exploring breast tissue microbial composition and the association with breast cancer risk factors. Breast Cancer Res. BCR 2023, 25, 82. [Google Scholar] [CrossRef]
- Wasiak, J.; Głowacka, P.; Pudlarz, A.; Pieczonka, A.M.; Dzitko, K.; Szemraj, J.; Witusik-Perkowska, M. Lactic Acid Bacteria-Derived Postbiotics as Adjunctive Agents in Breast Cancer Treatment to Boost the Antineoplastic Effect of a Conventional Therapeutic Comprising Tamoxifen and a New Drug Candidate: An Aziridine-Hydrazide Hydrazone Derivative. Molecules 2024, 29, 2292. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, C.; Cao, W.; Li, L.; Liu, K.; Zhu, H.; Balcha, F.; Fang, Y. Extracellular vesicles from Lacticaseibacillus paracasei PC-H1 inhibit HIF-1α-mediated glycolysis of colon cancer. Future Microbiol. 2024, 19, 227–239. [Google Scholar] [CrossRef]
- Matsuya-Ogawa, M.; Shibata, T.; Itoh, H.; Murakami, H.; Yaguchi, C.; Sugihara, K.; Kanayama, N. Oncoprotective Effects of Short-Chain Fatty Acids on Uterine Cervical Neoplasia. Nutr. Cancer 2019, 71, 312–319. [Google Scholar] [CrossRef]
- Yan, H.; Zhang, S.; Yang, L.; Jiang, M.; Xin, Y.; Liao, X.; Li, Y.; Lu, J. The Antitumor Effects of α-Linolenic Acid. J. Pers. Med. 2024, 14, 260. [Google Scholar] [CrossRef]
- Han, M.; Wang, N.; Han, W.; Liu, X.; Sun, T.; Xu, J. Specific vaginal and gut microbiome and the anti-tumor effect of butyrate in cervical cancer women. Transl. Oncol. 2024, 44, 1011902. [Google Scholar] [CrossRef]
- Martyniak, A.; Zakrzewska, Z.; Schab, M.; Zawartka, A.; Wędrychowicz, A.; Skoczeń, S.; Tomasik, P.J. Prevention and Health Benefits of Prebiotics, Probiotics and Postbiotics in Acute Lymphoblastic Leukemia. Microorganisms 2023, 11, 1775. [Google Scholar] [CrossRef]
- Park, K.B.; Oh, S.H.; Kim, N.S.; Oh, C.H.; Jeon, J.I. Kimchi Fermented in a Kimchi Refrigerator Showed Enhanced Anti-cancer Effects on Human Leukemia and Gastric Cancer Cells (LB405). FASEB J. 2014, 28, LB405. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism Characteristics of Lactic Acid Bacteria and the Expanding Applications in Food Industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Chuah, L.-O.; Foo, H.L.; Loh, T.C.; Mohammed, N.B.; Yeap, S.K.; Abdul, N.E.; Abdul, R.; Yusoff, K. Postbiotic Metabolites Produced by Lactobacillus Plantarum Strains Exert Selective Cytotoxicity Effects on Cancer Cells. BMC Complement. Altern. Med. 2019, 19, 114. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zeng, X.; Shen, J.; Wu, Z.; Guo, Y.; Du, Q.; Tu, M.; Pan, D. New clues for postbiotics to improve host health: A review from the perspective of function and mechanisms. J. Sci. Food Agric. 2024, 104, 6376–6387. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Arai, S.; Sato, S.; Iwabuchi, N.; Takara, T.; Tanaka, M. Effects of Heat-Killed Lacticaseibacillus paracasei MCC1849 on Immune Parameters in Healthy Adults-A Randomized, Double-Blind, Placebo-Controlled, Parallel-Group Study. Nutrients 2024, 16, 216. [Google Scholar] [CrossRef] [PubMed]
- Maghsood, F.; Mirshafiey, A.; Farahani, M.M.; Modarressi, M.H.; Jafari, P.; Motevaseli, E. Dual Effects of Cell Free Supernatants from Lactobacillus acidophilus and Lactobacillus rhamnosus GG in Regulation of MMP-9 by Up-Regulating TIMP-1 and Down-Regulating CD147 in PMADifferentiated THP-1 Cells. Cell J. 2018, 19, 559–568. [Google Scholar]
- Allouche, R.; Hafeez, Z.; Dary-Mourot, A.; Genay, M.; Miclo, L. Streptococcus thermophilus: A Source of Postbiotics Displaying Anti-Inflammatory Effects in THP 1 Macrophages. Molecules 2023, 29, 1552. [Google Scholar] [CrossRef] [PubMed]
- Kavita, H.; Chand, U.; Kushawaha, P.K. Postbiotics: An alternative and innovative intervention for the therapy of inflammatory bowel disease. Microbiol. Res. 2024, 279, 127550. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.T.; Han, Y.; Shapiro, D.; West, G.; Fiocchi, C.; Cresci, G.A.M. The Postbiotic Butyrate Mitigates Gut Mucosal Disruption Caused by Acute Ethanol Exposure. Int. J. Mol. Sci. 2024, 25, 1665. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Chen, C.; Patil, S.; Dong, S. Unveiling the therapeutic symphony of probiotics, prebiotics, and postbiotics in gut-immune harmony. Front. Nutr. 2024, 11, 1355542. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef] [PubMed]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 42. [Google Scholar] [CrossRef] [PubMed]
- Larraufie, P.; Martin-Gallausiaux, C.; Lapaque, N.; Dore, J.; Gribble, F.M.; Reimann, F.; Blottiere, H.M. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 2018, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Nowak, A.; Zakłos-Szyda, M.; Rosicka-Kaczmarek, J.; Motyl, I. Anticancer Potential of Post-Fermentation Media and Cell Extracts of Probiotic Strains: An In Vitro Study. Cancers 2022, 14, 1853. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, H.J.; Ji, K. The Proliferation Inhibitory Effect of Postbiotics Prepared from Probiotics with Antioxidant Activity against HT-29 Cells. Appl. Sci. 2022, 12, 12519. [Google Scholar] [CrossRef]
- Chen, M.; Ma, L.; Yu, H.; Huang, S.; Zhang, J.; Gong, J.; Yang, L.; Chen, L.; Luo, H.; Tian, L. JK5G postbiotics attenuate immune-related adverse events in NSCLC patients by regulating gut microbiota: A randomized controlled trial in China. Front. Oncol. 2023, 13, 1155592. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Xu, J.; Zhao, Z.; Guo, Y.; Zhang, H.; Jurutka, P.W.; Huang, D.; Cao, C.; Cheng, S. Dietary Lactobacillus rhamnosus GG extracellular vesicles enhance antiprogrammed cell death 1 (anti-PD-1) immunotherapy efficacy against colorectal cancer. Food Funct. 2023, 14, 10314–10328. [Google Scholar] [CrossRef] [PubMed]
- Fong, W.; Li, Q.; Yu, J. Gut microbiota modulation: A novel strategy for prevention and treatment of colorectal cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef] [PubMed]
- Kvakova, M.; Kamlarova, A.; Stofilova, J.; Benetinova, V.; Bertkova, I. Probiotics and postbiotics in colorectal cancer: Prevention and complementary therapy. World J. Gastroenterol. 2022, 28, 3370–3382. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xu, H.; Ou, Y.; Feng, Z.; Zhang, Q.; Zhu, Q.; Cai, Z. LPS-induced CXCR7 expression promotes gastric Cancer proliferation and migration via the TLR4/MD-2 pathway. Diagn. Pathol. 2019, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Arabzadeh, S.; Hossein, G.; Salehi-Dulabi, Z.; Zarnani, A.H. WNT5A-ROR2 is induced by inflammatory mediators and is involved in the migration of human ovarian cancer cell line SKOV-3. Cell Mol. Biol. Lett. 2016, 21, 9. [Google Scholar] [CrossRef] [PubMed]
- Deepak, V.; Ram Kumar Pandian, S.; Sivasubramaniam, S.D.; Nellaiah, H.; Sundar, K. Optimization of anticancer exopolysaccharide production from probiotic Lactobacillus acidophilus by response surface methodology. Prep. Biochem. Biotechnol. 2016, 46, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Hattar, K.; Reinert, C.P.; Sibelius, U.; Gokyildirim, M.Y.; Subtil, F.S.B.; Wilhelm, J.; Eul, B.; Dahlem, G.; Grimminger, F.; Seeger, W.; et al. Lipoteichoic acids from Staphylococcus aureus stimulate proliferation of human non-small-cell lung cancer cells in vitro. Cancer Immunol. Immunother. 2017, 66, 799–809. [Google Scholar] [CrossRef]
- Xie, N.; Wang, Y.; Wang, Q.; Li, F.R.; Guo, B. Lipoteichoic acid of Bifidobacterium in combination with 5-fluorouracil inhibit tumor growth and relieve the immunosuppression. Bull. Cancer 2012, 99, E55–E63. [Google Scholar] [CrossRef]
- Sadeghi, F.; Peymaeei, F.; Falahati, M.; Safari, E.; Farahyar, S.; Roudbar Mohammadi, S.; Roudbary, M. The effect of Candida cell wall beta-glucan on treatment-resistant LL/2 cancer cell line: In vitro evaluation. Mol. Biol. Rep. 2020, 47, 3653–3661. [Google Scholar] [CrossRef]
- Luo, S.; Li, Z.; Mao, L.; Chen, S.; Sun, S. Sodium butyrate induces autophagy in colorectal cancer cells through LKB1/AMPK signaling. J. Physiol. Biochem. 2019, 75, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Watkins, S.M.; Carter, L.C.; Mak, J.; Tsau, J.; Yamamoto, S.; German, J.B. Butyric acid and tributyrin induce apoptosis in human hepatic tumour cells. J. Dairy Res. 1999, 66, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, M.; Reger, M.; Marley, A.; Fan, H.; Liu, Z.; Zhang, J. Vitamin K intake and prostate cancer risk in the Prostate, Lung, Colorectal, and Ovarian Cancer (PLCO) Screening Trial. Am. J. Clin. Nutr. 2019, 109, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Haruna, Y.; Yakushijin, T.; Kawamoto, S. Efficacy and safety of sorafenib plus vitamin K treatment for hepatocellular carcinoma: A phase II, randomized study. Cancer Med. 2021, 10, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Seethaler, B.; Nguyen, N.K.; Basrai, M. Short-chain fatty acids are key mediators of the favorable effects of the Mediterranean diet on intestinal barrier integrity: Data from the randomized controlled LIBRE trial. Am. J. Clin. Nutr. 2022, 116, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Niccolai, E.; Baldi, S.; Ricci, F.; Russo, E.; Nannini, G.; Menicatti, M. Evaluation and comparison of short chain fatty acids composition in gut diseases. World J. Gastroenterol. 2019, 25, 5543–5558. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, F.; Adewiah, S.; Fatchiyah, F. The level short chain fatty acids and HSP 70 in colorectal cancer and non-colorectal cancer. Acta Inform. Med. 2018, 26, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Ocvirk, S.; Posma, J.M.; Li, J.V.; Koller, K.R.; Day, G.M.; Flanagan, C.A. A prospective cohort analysis of gut microbial co-metabolism in Alaska Native and rural African people at high and low risk of colorectal cancer. Am. J. Clin. Nutr. 2020, 111, 406–419. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Yu, Y.N.; Wang, J.L.; Lin, Y.W.; Kong, X.; Yang, C.Q. Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 2013, 97, 1044–1052. [Google Scholar] [CrossRef]
- Boutron-Ruault, M.C.; Marteau, P.; Lavergne-Slove, A.; Myara, A.; Gerhardt, M.F.; Franchisseur, C. Effects of a 3-mo consumption of short-chain fructo-oligosaccharides on parameters of colorectal carcinogenesis in patients with or without small or large colorectal adenomas. Nutr. Cancer 2005, 53, 160–168. [Google Scholar] [CrossRef]
- Kashtan, H.; Stern, H.S.; Jenkins, D.J.A.; Jenkins, A.L.; Thompson, L.U.; Hay, K. Colonic fermentation and markers of colorectal-cancer risk. Am. J. Clin. Nutr. 1992, 55, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Alvandi, E.; Wong, W.K.M.; Joglekar, M.V.; Spring, K.J.; Hardikar, A.A. Short-chain fatty acid concentrations in the incidence and risk-stratification of colorectal cancer: A systematic review and meta-analysis. BMC Med. 2022, 20, 323. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sadaghian, S.M.; Gabarrini, G.; Lisotto, P.; von Martels, J.Z.H.; Wardill, H.R.; Dijkstra, G.; Steinert, R.E.; Harmsen, H.J.M. Riboflavin Supplementation Promotes Butyrate Production in the Absence of Gross Compositional Changes in the Gut Microbiota. Antioxid. Redox Signal. 2023, 38, 282–297. [Google Scholar] [CrossRef] [PubMed]
- Fechner, A.; Fenske, K.; Jahreis, G. Effects of legume kernel fibres and citrus fibre on putative risk factors for colorectal cancer: A randomised, double-blind, crossover human intervention trial. Nutr. J. 2013, 12, 101. [Google Scholar] [CrossRef]
- Ohigashi, S.; Sudo, K.; Kobayashi, D.; Takahashi, T.; Nomoto, K.; Onodera, H. Significant changes in the intestinal environment after surgery in patients with colorectal cancer. J. Gastrointest. Surg. 2013, 17, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; He, Y.; Li, H.; Yu, D.; Na, L.; Sun, T.; Zhang, D.; Shi, X.; Xia, Y.; Jiang, T.; et al. Effects of prebiotics on immunologic indicators and intestinal microbiota structure in perioperative colorectal cancer patients. Nutrition 2019, 61, 132–142. [Google Scholar] [CrossRef]
- Motoori, M.; Yano, M.; Miyata, H.; Sugimura, K.; Saito, T.; Omori, T.; Fujiwara, Y.; Miyoshi, N.; Akita, H.; Gotoh, K.; et al. Randomized study of the effect of synbiotics during neoadjuvant chemotherapy on adverse events in esophageal cancer patients. Clin. Nutr. 2017, 36, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Hijová, E.; Szabadosova, V.; Štofilová, J.; Hrčková, G. Chemopreventive and metabolic effects of inulin on colon cancer development. J. Vet. Sci. 2013, 14, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Gao, C.; Hou, J.; Gu, J.; Law, B.Y.K.; Xu, Y. Non-Digestible Carbohydrate and the Risk of Colorectal Neoplasia: A Systematic Review. Nutr. Cancer 2021, 73, 31–44. [Google Scholar] [CrossRef]
- van Vorstenbosch, R.; Cheng, H.R.; Jonkers, D.; Penders, J.; Schoon, E.; Masclee, A.; van Schooten, F.J.; Smolinska, A.; Mujagic, Z. Systematic Review: Contribution of the Gut Microbiome to the Volatile Metabolic Fingerprint of Colorectal Neoplasia. Metabolites 2022, 13, 55. [Google Scholar] [CrossRef]
- Motoori, M.; Sugimura, K.; Tanaka, K.; Shiraishi, O.; Kimura, Y.; Miyata, H.; Yamasaki, M.; Makino, T.; Miyazaki, Y.; Iwama, M.; et al. Comparison of synbiotics combined with enteral nutrition and prophylactic antibiotics as supportive care in patients with esophageal cancer undergoing neoadjuvant chemotherapy: A multicenter randomized study. Clin. Nutr. 2022, 41, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, R.; Afaghi, A.; Babakhani, S.; Sohrabi, M.R.; Hosseini-Fard, S.R.; Babolhavaeji, K.; Khani Ali Akbari, S.; Yousefimashouf, R.; Karampoor, S. Role of microbiota-derived short-chain fatty acids in cancer development and prevention. Biomed. Pharmacother. 2021, 139, 111619. [Google Scholar] [CrossRef] [PubMed]
- Wegh, C.A.; Geerlings, S.Y.; Knol, J.; Roeselers, G.; Belzer, C. Postbiotics and their potential applications in early life nutrition and beyond. Int. J. Mol. Sci. 2019, 20, 4673. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Sun, Z.; Yang, Z.; Qiao, X. Microbiota-derived short-chain fatty acids and modulation of host-derived peptides formation: Focused on host defense peptides. Biomed. Pharmacother. 2023, 162, 114586. [Google Scholar] [CrossRef]
| Characteristics | Prebiotics | Probiotics | Postbiotics |
|---|---|---|---|
| Origin | Nondigestible fibers | Live microorganisms | Inanimate, dead or inactivated microorganisms |
| Sources | Vegetables and fruits | Yogurt, kefir, kimchi, miso, tempeh | Kefir, kombucha, yogurt, miso, tempeh, kimchi, butter |
| Primary Role | Provide nourishment to probiotics | Improve digestion, strengthen the immune system and positively influence mental health | Offer similar benefits of probiotic without the introduction of live microorganisms |
| Postbiotic Used | Type of Study | Bacterial Source | Type of Cancer | Up/Down | Antitumor Activity | |
|---|---|---|---|---|---|---|
| Li et al., 2019 [85] | LPS | In vitro | Helicobacter pylori | Gastric cancer cell lines (SGC7901, BGC823, others) |  | Proliferation |
| Migration | |||||
| Arabzadeh et al., 2016 [86] | LTA LPS | In vitro | - | Ovarian cancer cell (SKOV-3 cell line) |  | Cell viability |
| Inflammation | |||||
| Cell invasion | |||||
| Wnt5A–ROR2 complex | |||||
| Deepak et al., 2016 [87] | EPS | In vitro | Lactobacillus acidophilus | Colon cancer cells | | TIMP-3 |
| HO-1 | |||||
| HIF-2α | |||||
| PAI-1 | |||||
| VEGF | |||||
| HIF-1α | |||||
| Hattar et al., 2017 [88] | LTA | In vitro | Staphylococcus aureus | Colorectal cancer (HCT-116 cell line) | | Apoptosis |
| Adhesion | |||||
| Migration | |||||
| Xie et al., 2012 [89] | LTA + 5-fluorouracil | In vivo | Bifidobacterium | Hepatoma-22 cells inoculated in mice | | Tumor growth |
| T lymphocyte proliferation | |||||
| IFN-gamma regulatory | |||||
| T-cells | |||||
| TIM-3 | |||||
| FOXP3 | |||||
| Sadeghi et al., 2020 [90] | Beta-Glucan | In vitro | Candida albicans | Lung cancer cells | | SOX2 |
| OCT4 | |||||
| Luo et al., 2019 [91] | Sodium Butyrate | In vitro | - | Colorectal cancer cells (HCT-116 cell line) | | Autophagy |
| Autolysosomes | |||||
| AMP kinase | |||||
| LKB1 | |||||
| Watkins et al., 1999 [92] | Sodium Butyrate | In vitro | - | Hep G2 cells | | Histone H4 Acetylation |
| DNA Fragmentation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balendra, V.; Rosenfeld, R.; Amoroso, C.; Castagnone, C.; Rossino, M.G.; Garrone, O.; Ghidini, M. Postbiotics as Adjuvant Therapy in Cancer Care. Nutrients 2024, 16, 2400. https://doi.org/10.3390/nu16152400
Balendra V, Rosenfeld R, Amoroso C, Castagnone C, Rossino MG, Garrone O, Ghidini M. Postbiotics as Adjuvant Therapy in Cancer Care. Nutrients. 2024; 16(15):2400. https://doi.org/10.3390/nu16152400
Chicago/Turabian StyleBalendra, Vyshnavy, Roberto Rosenfeld, Chiara Amoroso, Cecilia Castagnone, Maria Grazia Rossino, Ornella Garrone, and Michele Ghidini. 2024. "Postbiotics as Adjuvant Therapy in Cancer Care" Nutrients 16, no. 15: 2400. https://doi.org/10.3390/nu16152400
APA StyleBalendra, V., Rosenfeld, R., Amoroso, C., Castagnone, C., Rossino, M. G., Garrone, O., & Ghidini, M. (2024). Postbiotics as Adjuvant Therapy in Cancer Care. Nutrients, 16(15), 2400. https://doi.org/10.3390/nu16152400