

Antidepressant Effect of Heracleum moellendorffii Extract on Behavioral Changes in Astrocyte Ablation Mouse Model of Depression by Modulating Neuroinflammation through the Inhibition of Lipocalin-2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material Preparation
2.2. Animals
2.3. Cannula Manufacturing and Implantation
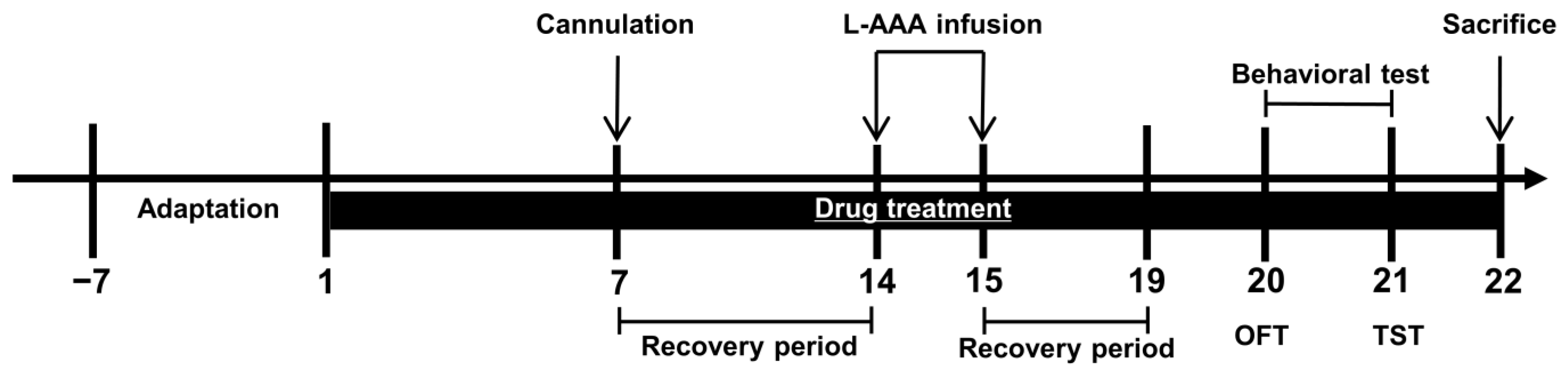
2.4. L-AAA Injection and Drug Administration
2.5. Behavioral Tests
2.6. Tissue Collection
2.7. Immunohistochemical Analysis
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Statistical Analysis
3. Results
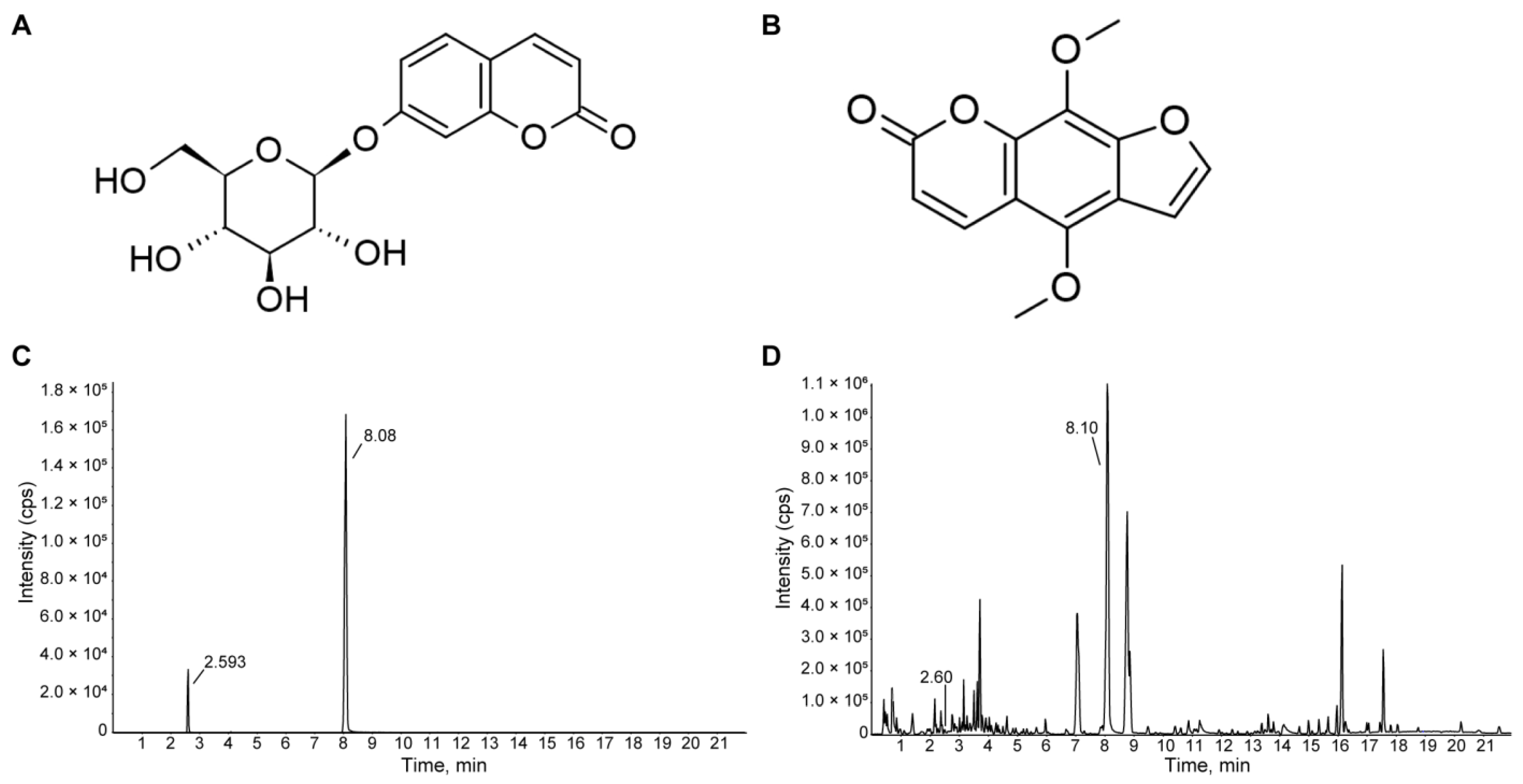
3.1. Identification of HME by Detecting Skimmin and Isopimpinellin in UHPLC Chromatograms
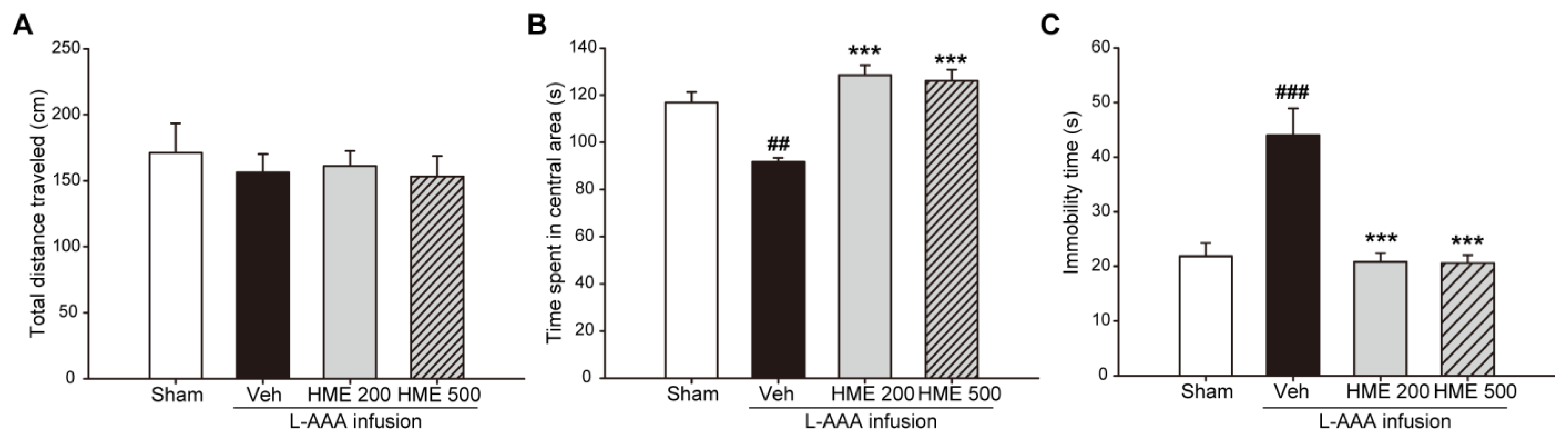
3.2. Effects of HME on Depression-like and Anxiety-like Behaviors in an L-AAA-Infused Mouse Model of Depression
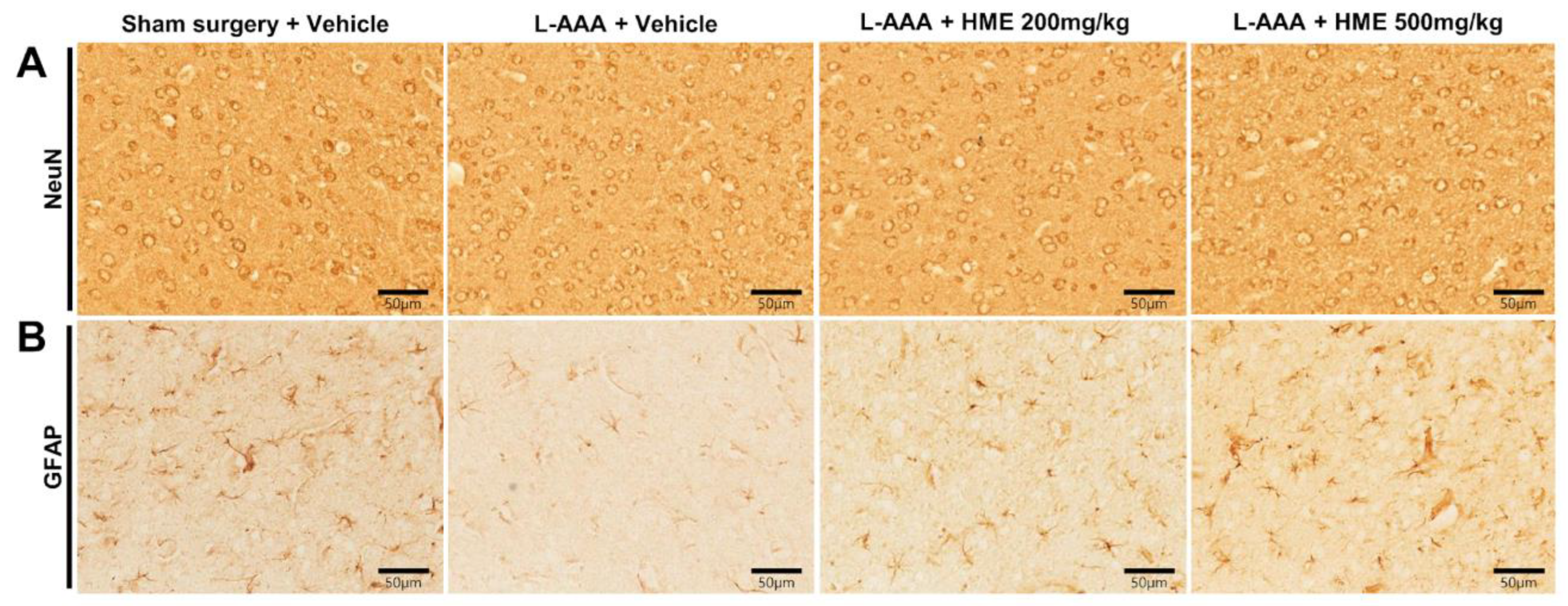
3.3. Effects of HME on NeuN and GFAP Expression in the PFC of an L-AAA-Infused Mouse Model of Depression
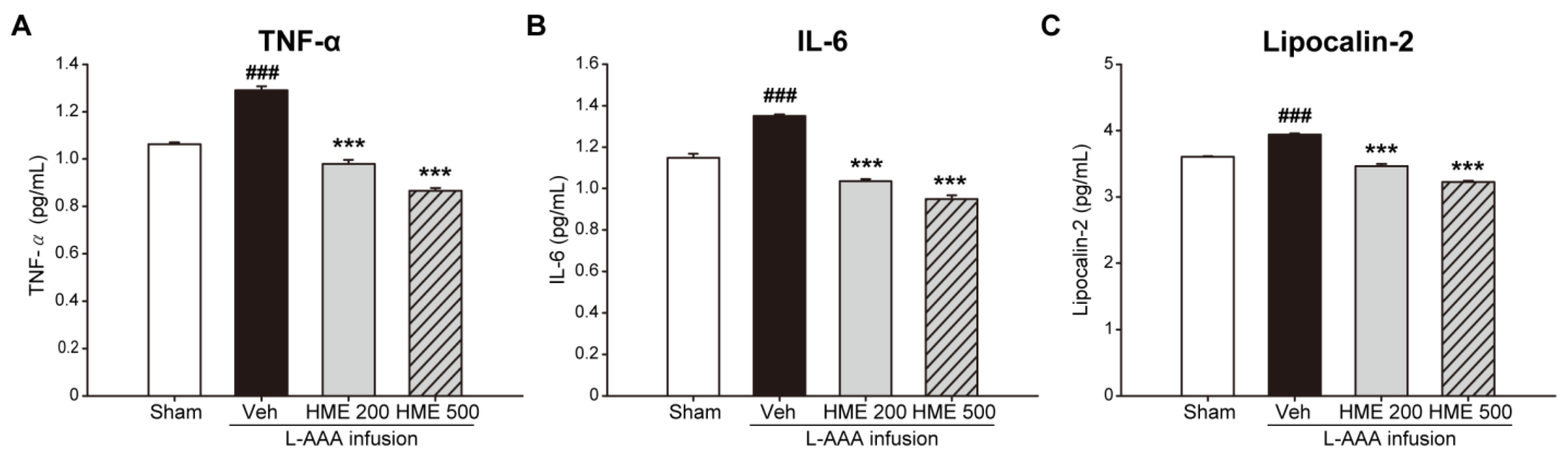
3.4. Anti-Inflammatory Effects of HME on Proinflammatory Cytokines and LCN2 in the PFC of an L-AAA-Infused Mouse Model of Depression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Su, W.-J.; Chen, Y.; Wu, T.-Y.; Gong, H.; Shen, X.-L.; Wang, Y.-X.; Sun, X.-J.; Jiang, C.-L. Effects of Hydrogen-Rich Water on Depressive-like Behavior in Mice. Sci. Rep. 2016, 6, 23742. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, A.; Santilli, C.; Furukawa, T.A.; Signoretti, A.; Nakagawa, A.; McGuire, H.; Churchill, R.; Barbui, C. Escitalopram versus other antidepressive agents for depression. Cochrane Database Syst. Rev. 2009, 2, CD006532. [Google Scholar] [CrossRef] [PubMed]
- Domin, H.; Szewczyk, B.; Pochwat, B.; Woźniak, M.; Śmiałowska, M. Antidepressant-like Activity of the Neuropeptide Y Y5 Receptor Antagonist Lu AA33810: Behavioral, Molecular, and Immunohistochemical Evidence. Psychopharmacology 2017, 234, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Sinyor, M.; Schaffer, A.; Levitt, A. The Sequenced Treatment Alternatives to Relieve Depression (STAR* D) Trial: A Review. Can. J. Psychiatry 2010, 55, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-L.; Deng, D.-P.; Pan, X.-H.; Wang, C.-Y.; Zhang, X.-L.; Chen, X.-M.; Wang, C.-H.; Liu, Y.-X.; Li, S.-C.; Bai, X.-Y.; et al. A Sub-Threshold Dose of Pilocarpine Increases Glutamine Synthetase in Reactive Astrocytes and Enhances the Progression of Amygdaloid-Kindling Epilepsy in Rats. NeuroReport 2016, 27, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Hirase, H.; Koizumi, S. Astrocytes as Therapeutic Targets in Brain Diseases. Neurosci. Res. 2018, 126, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Wilhelmsson, U.; Bogestål, Y.R.; Pekna, M. The Role of Astrocytes and Complement System in Neural Plasticity. Int. Rev. Neurobiol. 2007, 82, 95–111. [Google Scholar] [PubMed]
- Liu, F.; Wu, J.; Gong, Y.; Wang, P.; Zhu, L.; Tong, L.; Chen, X.; Ling, Y.; Huang, C. Harmine Produces Antidepressant-like Effects via Restoration of Astrocytic Functions. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Hidalgo, J.J.; Baucom, C.; Dilley, G.; Overholser, J.C.; Meltzer, H.Y.; Stockmeier, C.A.; Rajkowska, G. Glial Fibrillary Acidic Protein Immunoreactivity in the Prefrontal Cortex Distinguishes Younger from Older Adults in Major Depressive Disorder. Biol. Psychiatry 2000, 48, 861–873. [Google Scholar] [CrossRef]
- Alam, M.B.; Seo, B.-J.; Zhao, P.; Lee, S.-H. Anti-Melanogenic Activities of Heracleum moellendorffii via ERK1/2-Mediated MITF Downregulation. Int. J. Mol. Sci. 2016, 17, 1844. [Google Scholar] [CrossRef]
- Bang, J.-E.; Choi, H.-Y.; Kim, S.-I. Anti-Oxidative Activity and Chemical Composition of Various Heracleum moellendorffii Hance Extracts. Korean J. Food Preserv. 2009, 16, 765–771. [Google Scholar]
- Park, H.-J.; Nugroho, A.; Jung, B.-R.; Won, Y.-H.; Jung, Y.-J.; Kim, W.-B.; Choi, J.-S. Isolation and Quantitative Analysis of Flavonoids with Peroxynitritescavenging Effect from the Young Leaves of Heracleum moellendorffii. Korean J. Plant Resour. 2010, 23, 393–398. [Google Scholar]
- Kwon, Y.S.; Cho, H.Y.; Kim, C.M. The Chemical Consituents from Heracleum moellendorffii Roots. Yakhak Hoeji 2000, 44, 521–527. [Google Scholar]
- Kokare, D.M.; Shelkar, G.P.; Borkar, C.D.; Nakhate, K.T.; Subhedar, N.K. A Simple and Inexpensive Method to Fabricate a Cannula System for Intracranial Injections in Rats and Mice. J. Pharmacol. Toxicol. Methods 2011, 64, 246–250. [Google Scholar] [CrossRef] [PubMed]
- McCall, J.G.; Kim, T.; Shin, G.; Huang, X.; Jung, Y.H.; Al-Hasani, R.; Omenetto, F.G.; Bruchas, M.R.; Rogers, J.A. Fabrication and Application of Flexible, Multimodal Light-Emitting Devices for Wireless Optogenetics. Nat. Protoc. 2013, 8, 2413–2428. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, P.J.; Whittle, N.; Flynn, S.M.; Graybeal, C.; Pinard, C.R.; Gunduz-Cinar, O.; Kravitz, A.V.; Singewald, N.; Holmes, A. Prefrontal Single-Unit Firing Associated with Deficient Extinction in Mice. Neurobiol. Learn. Mem. 2014, 113, 69–81. [Google Scholar] [CrossRef]
- Jung, E.S.; Lee, H.J.; Sim, H.R.; Baik, J.H. Cocaine-Induced Behavioral Sensitization in Mice: Effects of Microinjection of Dopamine D2 Receptor Antagonist into the Nucleus Accumbens. Exp. Neurobiol. 2013, 22, 224–231. [Google Scholar] [CrossRef]
- Kenny, R.; Dinan, T.; Cai, G.; Spencer, S.J. Effects of Mild Calorie Restriction on Anxiety and Hypothalamic-Pituitary-Adrenal Axis Responses to Stress in the Male Rat. Physiol. Rep. 2014, 2, e00265. [Google Scholar] [CrossRef]
- Nie, L.; Wei, G.; Peng, S.; Qu, Z.; Yang, Y.; Yang, Q.; Huang, X.; Liu, J.; Zhuang, Z.; Yang, X. Melatonin Ameliorates Anxiety and Depression-like Behaviors and Modulates Proteomic Changes in Triple Transgenic Mice of Alzheimer’s Disease. Biofactors 2017, 43, 593–611. [Google Scholar] [CrossRef]
- Rajkowska, G.; Stockmeier, C.A. Astrocyte Pathology in Major Depressive Disorder: Insights from Human Postmortem Brain Tissue. Curr. Drug Targets 2013, 14, 1225–1236. [Google Scholar] [CrossRef]
- Takada, M.; Hattori, T. Fine Structural Changes in the Rat Brain after Local Injections of Gliotoxin, Alpha-Aminoadipic Acid. Histol. Histopathol. 1986, 1, 271–275. [Google Scholar] [PubMed]
- Nishimura, R.; Santos, D.; Fu, S.; Dwyer, B. Induction of Cell Death by L-Alpha-Aminoadipic Acid Exposure in Cultured Rat Astrocytes: Relationship to Protein Synthesis. Neurotoxicology 2000, 21, 313–320. [Google Scholar] [PubMed]
- Banasr, M.; Duman, R.S. Glial Loss in the Prefrontal Cortex Is Sufficient to Induce Depressive-like Behaviors. Biol. Psychiatry 2008, 64, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Son, H.; Kim, G.; Kim, S.; Lee, D.H.; Roh, G.S.; Kang, S.S.; Cho, G.J.; Choi, W.S.; Kim, H.J. Glutamine Deficiency in the Prefrontal Cortex Increases Depressive-like Behaviours in Male Mice. J. Psychiatry Neurosci. 2013, 38, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ni, J.; Gao, T.; Gao, C.; Guo, L.; Yin, X. Activation of Astrocytic Sigma-1 Receptor Exerts Antidepressant-like Effect via Facilitating CD38-driven Mitochondria Transfer. Glia 2020, 68, 2415–2426. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Xiao, Q.; Xie, L.; Yang, F.; Wang, L.; Tu, J. Astrocyte, a Promising Target for Mood Disorder Interventions. Front. Mol. Neurosci. 2019, 12, 136. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Verkhratsky, A.; Gu, L.; Li, B. Targeting Astrocytes in Major Depression. Expert Rev. Neurother. 2015, 15, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Śmiałowska, M.; Szewczyk, B.; Woźniak, M.; Wawrzak-Wleciał, A.; Domin, H. Glial Degeneration as a Model of Depression. Pharmacol. Rep. 2013, 65, 1572–1579. [Google Scholar] [CrossRef]
- Fu, Y.-W.; Jin, S.-Y.; Li, J.-T.; Li, X.-W.; Gao, T.-M.; Yang, J.-M. Mature Astrocytes as Source for Astrocyte Repopulation after Deletion in the Medial Prefrontal Cortex: Implications for Depression. Glia 2024. [Google Scholar] [CrossRef]
- Müller, N.; Schwarz, M.J. The Immune-Mediated Alteration of Serotonin and Glutamate: Towards an Integrated View of Depression. Mol. Psychiatry 2007, 12, 988–1000. [Google Scholar] [CrossRef]
- Fan, N.; Luo, Y.; Ou, Y.; He, H. Altered Serum Levels of TNF-α, IL-6, and IL-18 in Depressive Disorder Patients. Hum. Psychopharmacol. Clin. Exp. 2017, 32, e2588. [Google Scholar] [CrossRef] [PubMed]
- García-García, M.L.; Tovilla-Zárate, C.A.; Villar-Soto, M.; Juárez-Rojop, I.E.; González-Castro, T.B.; Genis-Mendoza, A.D.; Ramos-Méndez, M.Á.; López-Nárvaez, M.L.; Saucedo-Osti, A.S.; Ruiz-Quiñones, J.A.; et al. Fluoxetine Modulates the Pro-Inflammatory Process of IL-6, IL-1β and TNF-α Levels in Individuals with Depression: A Systematic Review and Meta-Analysis. Psychiatry Res. 2022, 307, 114317. [Google Scholar] [CrossRef] [PubMed]
- Chourbaji, S.; Urani, A.; Inta, I.; Sanchis-Segura, C.; Brandwein, C.; Zink, M.; Schwaninger, M.; Gass, P. IL-6 Knockout Mice Exhibit Resistance to Stress-Induced Development of Depression-like Behaviors. Neurobiol. Dis. 2006, 23, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Monje, F.J.; Cabatic, M.; Divisch, I.; Kim, E.-J.; Herkner, K.R.; Binder, B.R.; Pollak, D.D. Constant Darkness Induces IL-6-Dependent Depression-Like Behavior through the NF-κB Signaling Pathway. J. Neurosci. 2011, 31, 9075–9083. [Google Scholar] [CrossRef] [PubMed]
- Kaster, M.P.; Gadotti, V.M.; Calixto, J.B.; Santos, A.R.S.; Rodrigues, A.L.S. Depressive-like Behavior Induced by Tumor Necrosis Factor-α in Mice. Neuropharmacology 2012, 62, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, L.; Yi, S.; Jiang, X.; Qiao, Y.; Zhang, Y.; Xiao, C.; Zhou, T. Mouse Astrocytes Promote Microglial Ramification by Releasing TGF-β and Forming Glial Fibers. Front. Cell. Neurosci. 2020, 14, 195. [Google Scholar] [CrossRef] [PubMed]
- Moschen, A.R.; Adolph, T.E.; Gerner, R.R.; Wieser, V.; Tilg, H. Lipocalin-2: A Master Mediator of Intestinal and Metabolic Inflammation. Trends Endocrinol. Metab. 2017, 28, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Jha, M.K.; Lee, S.; Park, D.H.; Kook, H.; Park, K.-G.; Lee, I.-K.; Suk, K. Diverse Functional Roles of Lipocalin-2 in the Central Nervous System. Neurosci. Biobehav. Rev. 2015, 49, 135–156. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Zhang, C.; Rao, X.; Wan, W.; Lin, W.; Huang, S.; Ying, J.; Lin, Y.; Hua, F. The Interaction of Lipocalin-2 and Astrocytes in Neuroinflammation: Mechanisms and Therapeutic Application. Front. Immunol. 2024, 15, 1358719. [Google Scholar] [CrossRef]
- Naudé, P.J.W.; Eisel, U.L.M.; Comijs, H.C.; Groenewold, N.A.; De Deyn, P.P.; Bosker, F.J.; Luiten, P.G.M.; den Boer, J.A.; Oude Voshaar, R.C. Neutrophil Gelatinase-Associated Lipocalin: A Novel Inflammatory Marker Associated with Late-Life Depression. J. Psychosom. Res. 2013, 75, 444–450. [Google Scholar] [CrossRef]
- Sağlam, C.; Turan, İ.; Özaçmak, H.S. The Effect of Glucagon like Peptide-1 Receptor Agonist on Behavioral Despair and Anxiety-like Behavior in Ovariectomized Rats: Modulation of BDNF/CREB, Nrf2 and Lipocalin 2. Behav. Brain Res. 2022, 435, 114053. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.N.; Kim, J.D.; Yeo, J.H.; Son, H.-J.; Park, S.B.; Park, G.H.; Eo, H.J.; Jeong, J.B. Heracleum moellendorffii Roots Inhibit the Production of Pro-Inflammatory Mediators through the Inhibition of NF-κB and MAPK Signaling, and Activation of ROS/Nrf2/HO-1 Signaling in LPS-Stimulated RAW264. 7 Cells. BMC Complement. Altern. Med. 2019, 19, 310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, J.; Li, H.; Li, Y.; Liu, Y.; Zhang, D.; Zhang, F.; Zhou, W.; Chen, X. Skimmin, a Coumarin, Suppresses the Streptozotocin-Induced Diabetic Nephropathy in Wistar Rats. Eur. J. Pharmacol. 2012, 692, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xin, H.; Li, Y.; Zhang, D.; Shi, J.; Yang, J.; Chen, X. Skimmin, a Coumarin from Hydrangea Paniculata, Slows down the Progression of Membranous Glomerulonephritis by Anti-Inflammatory Effects and Inhibiting Immune Complex Deposition. Evid.-Based Complement. Altern. Med. 2013, 2013, 819296. [Google Scholar]
- Souri, E.; Farsam, H.; Sarkheil, P.; Ebadi, F. Antioxidant Activity of Some Furanocoumarins Isolated from Heracleum persicum. Pharm. Biol. 2004, 42, 396–399. [Google Scholar] [CrossRef]
- Zhang, H.L.; Wu, X.Y.; Mi, J.; Peng, Y.J.; Wang, Z.G.; Liu, Y.; Wu, X.L.; Gao, Y. A New Anti-Inflammatory Alkaloid from Roots of Heracleum dissectum. Chem. Biodivers. 2017, 14, e1700184. [Google Scholar] [CrossRef]
- Heinrich, M.; Jalil, B.; Abdel-Tawab, M.; Echeverria, J.; Kulić, Ž.; McGaw, L.J.; Pezzuto, J.M.; Potterat, O.; Wang, J.-B. Best Practice in the Chemical Characterisation of Extracts Used in Pharmacological and Toxicological Research—The ConPhyMP—Guidelines12. Front. Pharmacol. 2022, 13, 953205. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.; Kim, Y.; Kwon, Y.; Cho, S.-H. Antidepressant Effect of Heracleum moellendorffii Extract on Behavioral Changes in Astrocyte Ablation Mouse Model of Depression by Modulating Neuroinflammation through the Inhibition of Lipocalin-2. Nutrients 2024, 16, 2049. https://doi.org/10.3390/nu16132049
Hong S, Kim Y, Kwon Y, Cho S-H. Antidepressant Effect of Heracleum moellendorffii Extract on Behavioral Changes in Astrocyte Ablation Mouse Model of Depression by Modulating Neuroinflammation through the Inhibition of Lipocalin-2. Nutrients. 2024; 16(13):2049. https://doi.org/10.3390/nu16132049
Chicago/Turabian StyleHong, Soonsang, Yunna Kim, YongJu Kwon, and Seung-Hun Cho. 2024. "Antidepressant Effect of Heracleum moellendorffii Extract on Behavioral Changes in Astrocyte Ablation Mouse Model of Depression by Modulating Neuroinflammation through the Inhibition of Lipocalin-2" Nutrients 16, no. 13: 2049. https://doi.org/10.3390/nu16132049
APA StyleHong, S., Kim, Y., Kwon, Y., & Cho, S.-H. (2024). Antidepressant Effect of Heracleum moellendorffii Extract on Behavioral Changes in Astrocyte Ablation Mouse Model of Depression by Modulating Neuroinflammation through the Inhibition of Lipocalin-2. Nutrients, 16(13), 2049. https://doi.org/10.3390/nu16132049