
Therapeutic Effects of Natural Products on Liver Cancer and Their Potential Mechanisms

 ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Therapeutic Drugs and Methods of Liver Cancer Treatment
2.1. Surgical Treatment
2.2. Local (Ablative) Treatment
2.3. Hepatic Artery Therapy
2.4. Systemic Therapy-Chemotherapy
2.5. Radiotherapy
2.6. Immunotherapy
2.7. TCM (Traditional Chinese Medicine) Therapy
3. NPs with Therapeutic Activity for Liver Cancer
3.1. Polysaccharides
3.2. Flavonoids
3.3. Terpenoids
3.4. Alkaloids
3.5. Polyphenol Compounds
3.6. Quinone
3.7. Volatile Oils
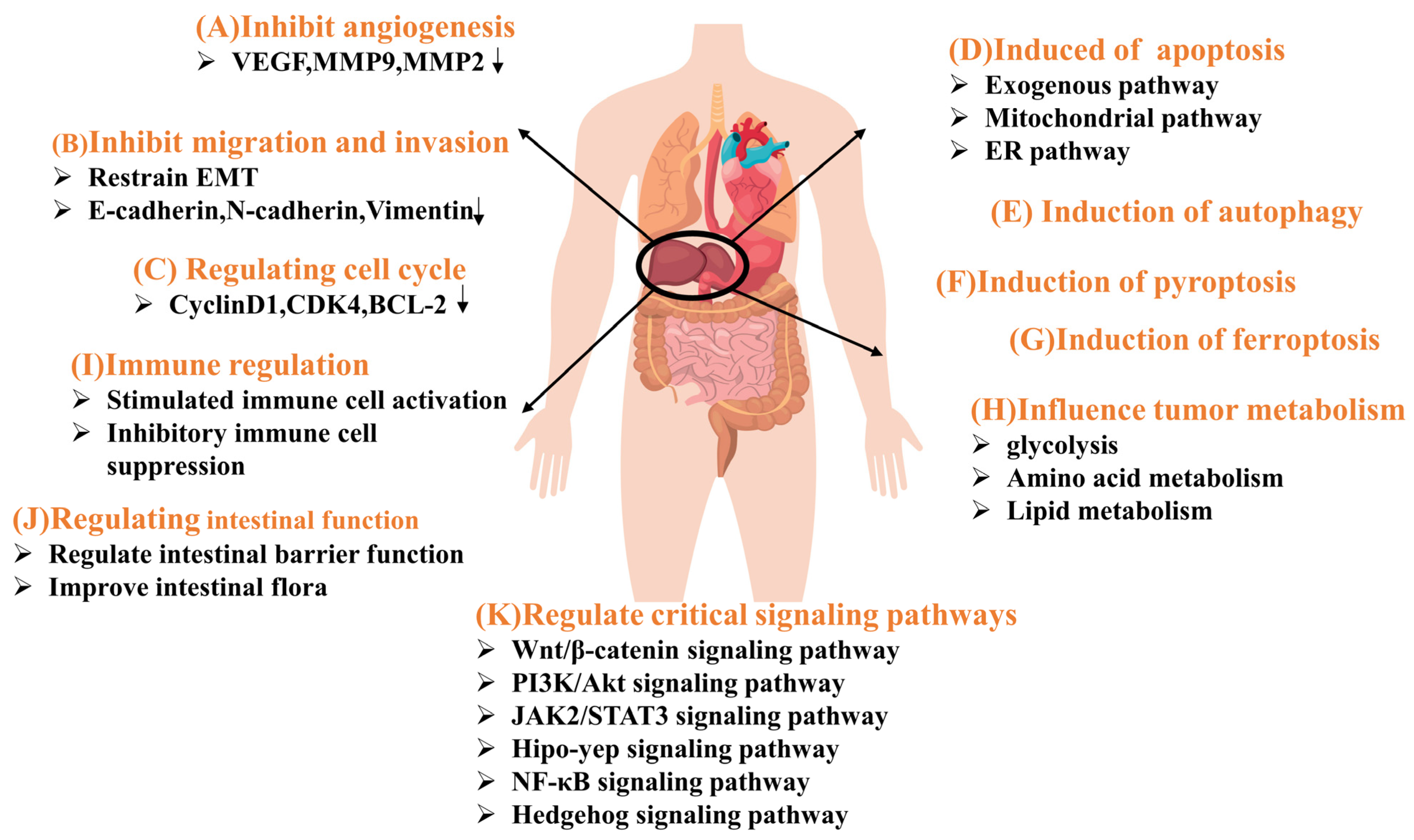
4. The Main Action Pathway and Potential Mechanism of Natural Compounds in the Treatment of Liver Cancer
4.1. Inhibition of Angiogenesis
4.2. Inhibition of Migration and Invasion
4.3. Regulating Cell Cycle
4.4. Induction of Apoptosis
4.5. Induction of Autophagy
4.6. Induction of Pyroptosis
4.7. Induction of Ferroptosis
4.8. Influence Tumor Metabolism
4.9. Immune Regulation
4.9.1. Activation of Stimulatory Immune Cells
4.9.2. Suppression of Inhibitory Immune Cells
4.10. Regulates Intestinal Function
4.11. Regulation of Key Signaling Pathways
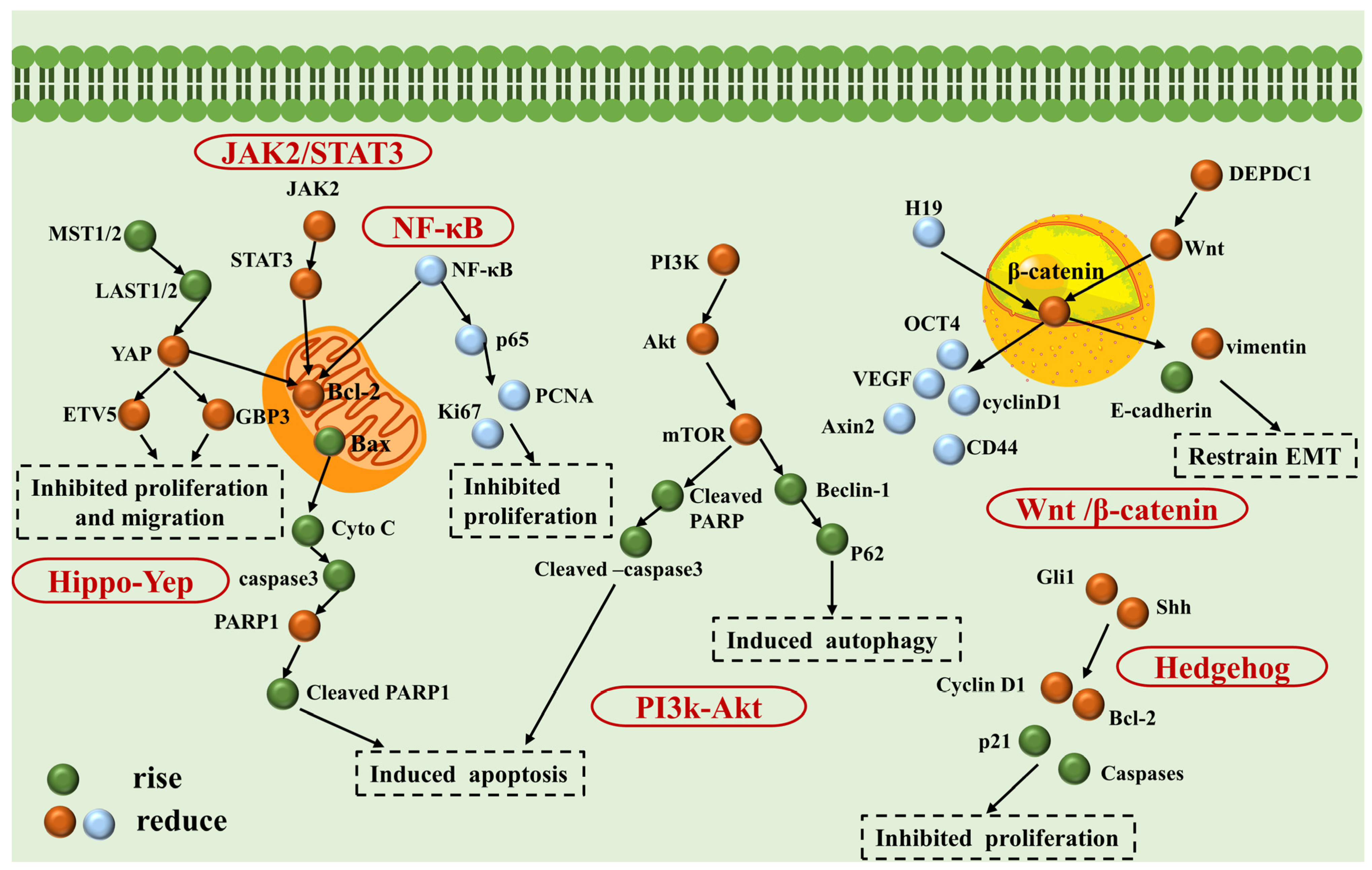
4.11.1. Wnt/β-Catenin Signaling Pathway
4.11.2. PI3K/Akt Signaling Pathway
4.11.3. JAK2/STAT3 Signaling Pathway
4.11.4. Hippo–YAP Signaling Pathway
4.11.5. NF-κB Signaling Pathway
4.11.6. Hedgehog Signaling Pathway
5. Summary and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sällberg, M.; Pasetto, A. Liver, Tumor and Viral Hepatitis: Key Players in the Complex Balance Between Tolerance and Immune Activation. Front. Immunol. 2020, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Balogh, J.; Victor Iii, D.; Asham, E.H.; Burroughs, S.G.; Boktour, M.; Saharia, A.; Li, X.; Ghobrial, R.M.; Monsour, H.P., Jr. Hepatocellular carcinoma: A review. J. Hepatocell. Carcinoma 2016, 3, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Man, S.; Luo, C.; Yan, M.; Zhao, G.; Ma, L.; Gao, W. Treatment for liver cancer: From sorafenib to natural products. Eur. J. Med. Chem. 2021, 224, 113690. [Google Scholar] [CrossRef]
- Pinter, M.; Sieghart, W.; Graziadei, I.; Vogel, W.; Maieron, A.; Königsberg, R.; Weissmann, A.; Kornek, G.; Plank, C.; Peck-Radosavljevic, M. Sorafenib in unresectable hepatocellular carcinoma from mild to advanced stage liver cirrhosis. Oncol. 2009, 14, 70–76. [Google Scholar] [CrossRef]
- Dika, I.E.; Abou-Alfa, G.K. Treatment options after sorafenib failure in patients with hepatocellular carcinoma. Clin. Mol. Hepatol. 2017, 23, 273–279. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, W.; Xu, L.; Zhou, H.; Yuan, M.; Xu, H. Recent Progress in Understanding the Action of Natural Compounds at Novel Therapeutic Drug Targets for the Treatment of Liver Cancer. Front. Oncol. 2022, 11, 795548. [Google Scholar] [CrossRef]
- Gao, S.; Jiang, X.; Wang, L.; Jiang, S.; Luo, H.; Chen, Y.; Peng, C. The pathogenesis of liver cancer and the therapeutic potential of bioactive substances. Front. Pharmacol. 2022, 13, 1029601. [Google Scholar] [CrossRef]
- Kamarajah, S.K.; Frankel, T.L.; Sonnenday, C.; Cho, C.S.; Nathan, H. Critical evaluation of the American Joint Commission on Cancer (AJCC) 8th edition staging system for patients with Hepatocellular Carcinoma (HCC): A Surveillance, Epidemiology, End Results (SEER) analysis. J. Surg. Oncol. 2018, 117, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Edge, S.B.; Compton, C.C. The American Joint Committee on Cancer: The 7th edition of the AJCC cancer staging manual and the future of TNM. Ann. Surg. Oncol. 2010, 17, 1471–1474. [Google Scholar] [CrossRef]
- Vauthey, J.N. Simplified Staging for Hepatocellular Carcinoma. J. Clin. Oncol. 2002, 20, 1527–1536. [Google Scholar] [CrossRef]
- Hemming, A.W.; Scudamore, C.H.; Shackleton, C.R.; Pudek, M.; Erb, S.R. Indocyanine green clearance as a predictor of successful hepatic resection in cirrhotic patients. Am. J. Surg. 1992, 163, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Kamo, N.; Kaido, T.; Yagi, S.; Okajima, H.; Uemoto, S. Liver transplantation for small hepatocellular carcinoma. Hepatobiliary Surg. Nutr. 2016, 5, 391–398. [Google Scholar] [CrossRef]
- Tabrizian, P.; Holzner, M.; Halazun, K.; Agopian, V.G.; Busuttil, R.W.; Yao, F.; Roberts, J.; Florman, S.S.; Schwartz, M.E.; Emond, J.C. A US multicenter analysis of 2529 HCC patients undergoing liver transplantation: 10-year outcome assessing the role of down-staging to within Milan criteria. Hepatology 2019, 70, 10–11. [Google Scholar]
- Zhou, M.; Wang, H.; Zeng, X.; Yin, P.; Zhu, J.; Chen, W.; Li, X.; Wang, L.; Wang, L.; Liu, Y.; et al. Mortality, morbidity, and risk factors in China and its provinces, 1990-2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019, 394, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Takayama, T.; Hasegawa, K.; Izumi, N.; Kudo, M.; Shimada, M.; Yamanaka, N.; Inomata, M.; Kaneko, S.; Nakayama, H.; Kawaguchi, Y.; et al. Surgery versus Radiofrequency Ablation for Small Hepatocellular Carcinoma: A Randomized Controlled Trial (SURF Trial). Liver Cancer 2021, 11, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ni, C.-F. Current status of the combination therapy of transarterial chemoembolization and local ablation for hepatocellular carcinoma. Abdom. Radiol. 2019, 44, 2268–2275. [Google Scholar] [CrossRef] [PubMed]
- Heimbach, J.K.; Kulik, L.M.; Finn, R.S.; Sirlin, C.B.; Abecassis, M.M.; Roberts, L.R.; Zhu, A.X.; Murad, M.H.; Marrero, J.A. Aasld guidelines for the treatment of hepatocellular carcinoma. Hepatology 2018, 67, 358–380. [Google Scholar] [CrossRef]
- Vogel, A.; Cervantes, A.; Chau, I.; Daniele, B.; Llovet, J.M.; Meyer, T.; Nault, J.C.; Neumann, U.; Ricke, J.; Sangro, B.; et al. Hepatocellular carcinoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2019, 30, 871–873. [Google Scholar] [CrossRef]
- Kudo, M.; Kawamura, Y.; Hasegawa, K.; Tateishi, R.; Kariyama, K.; Shiina, S.; Toyoda, H.; Imai, Y.; Hiraoka, A.; Ikeda, M.; et al. Management of Hepatocellular Carcinoma in Japan: JSH Consensus Statements and Recommendations 2021 Update. Liver Cancer 2021, 10, 181–223. [Google Scholar] [CrossRef]
- Kudo, M.; Finn, R.S.; Qin, S.; Han, K.-H.; Ikeda, K.; Piscaglia, F.; Baron, A.; Park, J.-W.; Han, G.; Jassem, J.; et al. Lenvatinib versus sorafenib in first-line treatment of patients with unresectable hepatocellular carcinoma: A randomised phase 3 non-inferiority trial. Lancet 2018, 391, 1163–1173. [Google Scholar] [CrossRef]
- Huang, A.; Yang, X.-R.; Chung, W.-Y.; Dennison, A.R.; Zhou, J. Targeted therapy for hepatocellular carcinoma. Signal Transduct. Target. Ther. 2020, 5, 146. [Google Scholar] [CrossRef] [PubMed]
- Abou-Alfa, G.K.; Meyer, T.; Cheng, A.-L.; El-Khoueiry, A.B.; Rimassa, L.; Ryoo, B.-Y.; Cicin, I.; Merle, P.; Chen, Y.; Park, J.-W.; et al. Cabozantinib in Patients with Advanced and Progressing Hepatocellular Carcinoma. N. Engl. J. Med. 2018, 379, 54–63. [Google Scholar] [CrossRef]
- Bruix, J.; Qin, S.; Merle, P.; Granito, A.; Huang, Y.-H.; Bodoky, G.; Pracht, M.; Yokosuka, O.; Rosmorduc, O.; Breder, V.; et al. Regorafenib for patients with hepatocellular carcinoma who progressed on sorafenib treatment (RESORCE): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 389, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Bian, H.; Wang, L.; Sun, X.; Xu, X.; Yan, H.; Xia, M.; Chang, X.; Lu, Y.; Li, Y.; et al. Berberine attenuates nonalcoholic hepatic steatosis through the AMPK-SREBP-1c-SCD1 pathway. Free Radic. Biol. Med. 2019, 141, 192–204. [Google Scholar] [CrossRef]
- Chen, K.-W. Current systemic treatment of hepatocellular carcinoma: A review of the literature. World J. Hepatol. 2015, 7, 1412–1420. [Google Scholar] [CrossRef]
- Wang, A.; Wu, L.; Lin, J.; Han, L.; Bian, J.; Wu, Y.; Robson, S.C.; Xue, L.; Ge, Y.; Sang, X.; et al. Whole-exome sequencing reveals the origin and evolution of hepato-cholangiocarcinoma. Nat. Commun. 2018, 9, 894. [Google Scholar] [CrossRef]
- Chen, L.; Chen, S.; Zheng, C. Research progress in radiotherapy for liver cancers. J. Interv. Radiol. 2020, 29, 631–635. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, Y.; Feng, J.; Li, J.; Ji, J.; Wu, L.; Yu, Q.; Dai, W.; Wu, J.; Zhou, Y.; et al. Cellular based immunotherapy for primary liver cancer. J. Exp. Clin. Cancer Res. 2021, 40, 250. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, C.; Luo, J.; Gong, C.; Wang, L.; Zhu, S. Traditional Chinese Medicine for Cancer Treatment. Am. J. Chin. Med. 2024, 1–22. [Google Scholar] [CrossRef]
- Peng, Y.; Wu, X.; Zhang, Y.; Yin, Y.; Chen, X.; Zheng, D.; Wang, J. An Overview of Traditional Chinese Medicine in the Treatment After Radical Resection of Hepatocellular Carcinoma. J. Hepatocell. Carcinoma 2023, 10, 2305–2321. [Google Scholar] [CrossRef]
- Pu, Y.; Zhu, J.; Xu, J.; Zhang, S.; Bao, Y. Antitumor effect of a polysaccharide from Pseudostellaria heterophylla through reversing tumor-associated macrophages phenotype. Int. J. Biol. Macromol. 2022, 220, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xing, J.; Huang, Y.; Bo, R.; Zheng, S.; Luo, L.; Niu, Y.; Zhang, Y.; Hu, Y.; Liu, J.; et al. Activation effect of Ganoderma lucidum polysaccharides liposomes on murine peritoneal macrophages. Int. J. Biol. Macromol. 2015, 82, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Liu, S.; Li, X.; He, J.; He, L.; Li, Y.; Yang, C.; Li, Y.; Hua, Y.; Guo, J. The Large Molecular Weight Polysaccharide from Wild Cordyceps and Its Antitumor Activity on H22 Tumor-Bearing Mice. Molecules 2023, 28, 3351. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Xu, J.; Liao, W.; Wen, Y.; Jiang, S.; Wen, J.; Zhao, C. Suppression of hepatocellular carcinoma by Ulva lactuca ulvan via gut microbiota and metabolite interactions. J. Adv. Res. 2023, 52, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Zhou, S.; Wang, Y.; Xu, X.; Zhang, L.; Cai, Z. New insights into the anti- hepatoma mechanism of triple-helix β- glucan by metabolomics profiling. Carbohydr. Polym. 2021, 269, 118289. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.; Hao, W. Immunomodulatory function and anti-tumor mechanism of natural polysaccharides: A review. Front. Immunol. 2023, 14, 1147641. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ji, H.-Y.; Liu, C.; Liu, A.-J. The structural characteristics of an acid-soluble polysaccharide from Grifola frondosa and its antitumor effects on H22-bearing mice. Int. J. Biol. Macromol. 2020, 158, 1288–1298. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Niu, L.-L.; Liu, H.-P.; Wu, Y.-R.; Li, M.-Y.; Jia, Q. Structural characterization of a novel polysaccharide from Pleurotus citrinopileatus and its antitumor activity on H22 tumor-bearing mice. Int. J. Biol. Macromol. 2020, 168, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-L.; Tang, J.-F.; Tan, W.-L.; Zhang, T.; Zeng, D.; Zhao, S.; Ran, J.-H.; Li, J.; Wang, Y.-P.; Chen, D.-L. The anti-hepatocellular carcinoma effects of polysaccharides from Ganoderma lucidum by regulating macrophage polarization via the MAPK/NF-κB signaling pathway. Food Funct. 2023, 14, 3155–3168. [Google Scholar] [CrossRef]
- Liu, C.; Dai, K.-Y.; Ji, H.-Y.; Jia, X.-Y.; Liu, A.-J. Structural characterization of a low molecular weight Bletilla striata polysaccharide and antitumor activity on H22 tumor-bearing mice. Int. J. Biol. Macromol. 2022, 205, 553–562. [Google Scholar] [CrossRef]
- Zhang, J.; Dai, K.; Li, M. Preparation of Water-Soluble Acetylaminoglucan with Low Molecular Weight and Its Anti-Tumor Activity on H22 Tumor-Bearing Mice. Molecules 2022, 27, 7273. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Chang, M.; Liu, H.; Ding, S.; Yan, Z.; Si, K.; Gong, T. The Structural Characteristics of an Acidic Water-Soluble Polysaccharide from Bupleurum chinense DC and Its In Vivo Anti-Tumor Activity on H22 Tumor-Bearing Mice. Polymers 2022, 14, 1119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, M.; Liu, H.; Zhang, S.; Wang, B.; Liu, Y.; Zhang, X.; Zhang, M. Polysaccharide of Asparagus cochinchinensis (Lour.) Merr: Preliminary characterization and antitumor activity in vivo. Food Biosci. 2023, 56, 103387. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Q.; Liu, H.; Niu, L.; Li, M.; Jia, Q. A heteropolysaccharide from Rhodiola rosea L.: Preparation, purification and anti-tumor activities in H22-bearing mice. Food Sci. Hum. Wellness 2023, 12, 536–545. [Google Scholar] [CrossRef]
- Ren, F.; Li, J.; Yuan, X.; Wang, Y.; Wu, K.; Kang, L.; Luo, Y.; Zhang, H.; Yuan, Z. Dandelion polysaccharides exert anticancer effect on Hepatocellular carcinoma by inhibiting PI3K/AKT/mTOR pathway and enhancing immune response. J. Funct. Foods 2019, 55, 263–274. [Google Scholar] [CrossRef]
- Ren, F.; Yang, Y.; Wu, K.; Zhao, T.; Shi, Y.; Song, M.; Li, J. The Effects of Dandelion Polysaccharides on Iron Metabolism by Regulating Hepcidin via JAK/STAT Signaling Pathway. Oxidative Med. Cell. Longev. 2021, 2021, 7184760. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.-Q.; Ding, S.-Y.; Chen, P.; Liu, H.-P.; Chang, M.-L.; Shi, S.-Y. A water-soluble polysaccharide from Eucommia folium: The structural characterization and anti-tumor activity in vivo. Glycoconj. J. 2022, 39, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Wei, J.; Chen, S.; Wang, H.; Ning, L.; Luo, S.-H.; Liu, C.-L.; Song, G.; Yao, Q. Fucoidan Inhibits the Progression of Hepatocellular Carcinoma via Causing lncRNA LINC00261 Overexpression. Front. Oncol. 2021, 11, 653902. [Google Scholar] [CrossRef]
- Su, J.; Liao, D.; Su, Y.; Liu, S.; Jiang, L.; Wu, J.; Liu, Z.; Wu, Y. Novel polysaccharide extracted from Sipunculus nudus inhibits HepG2 tumour growth in vivo by enhancing immune function and inducing tumour cell apoptosis. J. Cell. Mol. Med. 2021, 25, 8338–8351. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, L.; Chen, X.; Zhao, Y.; Yang, A.; Huang, H.; Ouyang, L.; Pang, D.; Xie, J.; Liu, D.; et al. DHMMF, a natural flavonoid from Resina Draconis, inhibits hepatocellular carcinoma progression via inducing apoptosis and G2/M phase arrest mediated by DNA damage-driven upregulation of p21. Biochem. Pharmacol. 2023, 211, 115518. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Wang, Y.; Liu, F.; Liu, Y.; Liang, J.; Zhan, R.; Wu, Y.; Ren, H.; Zhang, X.; et al. Sinensetin suppresses angiogenesis in liver cancer by targeting the VEGF/VEGFR2/AKT signaling pathway. Exp. Ther. Med. 2022, 23, 360. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Liu, M.; Wang, Y.; Luo, S.; Xu, Y.; Ye, B.; Zheng, L.; Meng, K.; Li, L. Icaritin Induces Anti-tumor Immune Responses in Hepatocellular Carcinoma by Inhibiting Splenic Myeloid-Derived Suppressor Cell Generation. Front. Immunol. 2021, 12, 609295. [Google Scholar] [CrossRef] [PubMed]
- Baby, J.; Devan, A.R.; Kumar, A.R.; Gorantla, J.N.; Nair, B.; Aishwarya, T.S.; Nath, L.R. Cogent role of flavonoids as key orchestrators of chemoprevention of hepatocellular carcinoma: A review. J. Food Biochem. 2021, 45, e13761. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, R.; Xiu, Z.; Yang, X.; Zhu, Y.; Han, J.; Li, S.; Li, Y.; Sun, L.; Li, X.; et al. Neobavaisoflavone induces pyroptosis of liver cancer cells via Tom20 sensing the activated ROS signal. Phytomedicine 2023, 116, 154869. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Das, A.K.; Bhattacharya, S.; Gachhui, R.; Sil, P.C. Isorhamnetin exerts anti-tumor activity in DEN + CCl4-induced HCC mice. Med. Oncol. 2023, 40, 188. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.-f.; Zheng, Y.-B.; Shi, C.-J.; Zhang, F.-w.; Zhang, J.-f.; Fu, W.-m. H19-Wnt/β-catenin regulatory axis mediates the suppressive effects of apigenin on tumor growth in hepatocellular carcinoma. Eur. J. Pharmacol. 2021, 893, 173810. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Zhang, P.; Sun, Z.; Liu, X.; Zhang, X.; Liu, X.; Wang, D.; Meng, Z. Lysionotin induces apoptosis of hepatocellular carcinoma cells via caspase-3 mediated mitochondrial pathway. Chem. -Biol. Interact. 2021, 344, 109500. [Google Scholar] [CrossRef] [PubMed]
- Chandrababu, G.; Varkey, M.; Devan, A.R.; Anjaly, M.V.; Unni, A.R.; Nath, L.R. Kaempferide exhibits an anticancer effect against hepatocellular carcinoma in vitro and in vivo. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2023, 396, 2461–2467. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Almutairi, M.M.; Li, S.; Li, J. Wogonin inhibits cell cycle progression by activating the glycogen synthase kinase-3 beta in hepatocellular carcinoma. Phytomedicine 2020, 68, 153174. [Google Scholar] [CrossRef]
- Chen, H.; Shang, X.; Yuan, H.; Niu, Q.; Chen, J.; Luo, S.; Li, W.; Li, X. Total flavonoids of Oldenlandia diffusa (Willd.) Roxb. suppresses the growth of hepatocellular carcinoma through endoplasmic reticulum stress-mediated autophagy and apoptosis. Front. Pharmacol. 2022, 13, 1019670. [Google Scholar] [CrossRef]
- Song, L.; Luo, Y.; Li, S.; Hong, M.; Wang, Q.; Chi, X.; Yang, C. ISL Induces Apoptosis and Autophagy in Hepatocellular Carcinoma via Downregulation of PI3K/AKT/mTOR Pathway in vivo and in vitro. Drug Des. Dev. Ther. 2020, 14, 4363–4376. [Google Scholar] [CrossRef]
- Li, G.; Qi, L.; Chen, H.; Tian, G. Involvement of NF-κB/PI3K/AKT signaling pathway in the protective effect of prunetin against a diethylnitrosamine induced hepatocellular carcinogenesis in rats. J. Biochem. Mol. Toxicol. 2022, 36, e23016. [Google Scholar] [CrossRef] [PubMed]
- Ke, M.; Zhang, Z.; Xu, B.; Zhao, S.; Ding, Y.; Wu, X.; Wu, R.; Lv, Y.; Dong, J. Baicalein and baicalin promote antitumor immunity by suppressing PD-L1 expression in hepatocellular carcinoma cells. Int. Immunopharmacol. 2019, 75, 105824. [Google Scholar] [CrossRef] [PubMed]
- Mo’men, Y.S.; Hussein, R.M.; Kandeil, M.A. Involvement of PI3K/Akt pathway in the protective effect of hesperidin against a chemically induced liver cancer in rats. J. Biochem. Mol. Toxicol. 2019, 33, e22305. [Google Scholar] [CrossRef] [PubMed]
- Badroon, N.; Abdul Majid, N.; Al-Suede, F.; Nazari, V.M.; Giribabu, N.; Abdul Majid, A.; Eid, E.; Alshawsh, M. Cardamonin Exerts Antitumor Effect on Human Hepatocellular Carcinoma Xenografts in Athymic Nude Mice through Inhibiting NF-κβ Pathway. Biomedicines 2020, 8, 586. [Google Scholar] [CrossRef]
- Fu, H.; Liu, X.; Jin, L.; Lang, J.; Hu, Z.; Mao, W.; Cheng, C.; Shou, Q. Safflower yellow reduces DEN-induced hepatocellular carcinoma by enhancing liver immune infiltration through promotion of collagen degradation and modulation of gut microbiota. Food Funct. 2021, 12, 10632–10643. [Google Scholar] [CrossRef]
- Li, L.; Xu, H.; Qu, L.; Xu, K.; Liu, X. Daidzin inhibits hepatocellular carcinoma survival by interfering with the glycolytic/gluconeogenic pathway through downregulation of TPI1. Biofactors 2022, 48, 883–896. [Google Scholar] [CrossRef]
- Shao, H.; Chen, J.; Li, A.; Ma, L.; Tang, Y.; Chen, H.; Chen, Y.; Liu, J. Salvigenin Suppresses Hepatocellular Carcinoma Glycolysis and Chemoresistance Through Inactivating the PI3K/AKT/GSK-3β Pathway. Appl. Biochem. Biotechnol. 2023, 195, 5217–5237. [Google Scholar] [CrossRef]
- Yang, W.; Chen, X.; Li, Y.; Guo, S.; Wang, Z.; Yu, X. Advances in Pharmacological Activities of Terpenoids. Nat. Prod. Commun. 2020, 15, 1934578X20903555. [Google Scholar] [CrossRef]
- Wróblewska-Łuczka, P.; Cabaj, J.; Bargieł, J.; Łuszczki, J.J. Anticancer effect of terpenes: Focus on malignant melanoma. Pharmacol. Rep. 2023, 75, 1115–1125. [Google Scholar] [CrossRef]
- de Vasconcelos Cerqueira Braz, J.; Carvalho Nascimento Júnior, J.A.; Serafini, M.R. Terpenes with Antitumor Activity: A Patent Review. Recent Pat. Anti-Cancer Drug Discov. 2020, 15, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Huo, R.; Su, W.; Xu, W.; Qiu, Z.; Wang, W.; Qiu, Y. The anticancer mechanism of Terpenoid-based drug candi-dates: Focus on tumor microenvironment. Anti-Cancer Agents Med. Chem. 2023. ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H. Pharmacological and medical applications of Panax ginseng and ginsenosides: A review for use in cardiovascular diseases. J. Ginseng Res. 2018, 42, 264–269. [Google Scholar] [CrossRef]
- Jiang, R.; Luo, S.; Zhang, M.; Wang, W.; Zhuo, S.; Wu, Y.; Qiu, Q.; Yuan, Y.; Jiang, X. Ginsenoside Rh4 inhibits inflammation-related hepatocellular carcinoma progression by targeting HDAC4/IL-6/STAT3 signaling. Mol. Genet. Genom. 2023, 298, 1479–1492. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, M.; Chen, J.; Zhao, J.; Su, J.; Zhang, X. Ginsenoside Compound K Regulates HIF-1α-Mediated Glycolysis Through Bclaf1 to Inhibit the Proliferation of Human Liver Cancer Cells. Front. Pharmacol. 2020, 11, 583334. [Google Scholar] [CrossRef]
- Qu, L.; Liu, Y.; Deng, J.; Ma, X.; Fan, D. Ginsenoside Rk3 is a novel PI3K/AKT-targeting therapeutics agent that regulates autophagy and apoptosis in hepatocellular carcinoma. J. Pharm. Anal. 2023, 13, 463–482. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.-R.; Huang, Q.-X.; Hong, H.; Pan, Y.; Luo, T.; Li, J.; Deng, Z.-Y.; Chen, F. Ginsenoside Rh2 and its octyl ester derivative inhibited invasion and metastasis of hepatocellular carcinoma via the c-Jun/COX2/PGE2 pathway. Phytomedicine 2023, 121, 155131. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-C.; Hu, Q.-R.; Luo, T.; Wei, W.-C.; Wu, H.-J.; Li, J.; Zheng, L.-F.; Xu, Q.-Y.; Deng, Z.-Y.; Chen, F. The immunomodulatory effects of ginsenoside derivative Rh2-O on splenic lymphocytes in H22 tumor-bearing mice is partially mediated by TLR4. Int. Immunopharmacol. 2021, 101, 108316. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Z.; Zhu, Y.; Li, S.; Li, Y.; Yang, X.; Li, Y.; Song, G.; Jin, N.; Fang, J.; Han, J.; et al. Betulinic acid inhibits growth of hepatoma cells through activating the NCOA4-mediated ferritinophagy pathway. J. Funct. Foods 2023, 102, 105441. [Google Scholar] [CrossRef]
- Wang, J.; Pu, J.; Zhang, Z.; Feng, Z.; Han, J.; Su, X.; Shi, L. Triterpenoids of Ganoderma lucidum inhibited S180 sarcoma and H22 hepatoma in mice by regulating gut microbiota. Heliyon 2023, 9, e16682. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, X.; Fan, D. Ginsenoside CK Inhibits Hypoxia-Induced Epithelial–Mesenchymal Transformation through the HIF-1α/NF-κB Feedback Pathway in Hepatocellular Carcinoma. Foods 2021, 10, 1195. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Fu, J.; Lv, X.; Wang, L.; Wang, H.; Li, R. Saikosaponin b2 inhibits tumor angiogenesis in liver cancer via down-regulation of VEGF/ERK/HIF-1α signaling. Oncol. Rep. 2023, 50, 8573. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Gao, Z.; Lv, X.; Zhu, Y.; Li, R.; Li, S. Saikosaponin-b2 Inhibits Primary Liver Cancer by Regulating the STK4/IRAK1/NF-κB Pathway. Biomedicines 2023, 11, 2859. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.F.; Hou, S.; Xu, J.-C.; Chen, Y.; Zhu, L.-L.; Xu, Y.-Y.; Chen, Y.-Q.; Li, M.-M.; Li, L.-L.; Yang, J.-J.; et al. Amelioratory effects of astragaloside IV on hepatocarcinogenesis via Nrf2-mediated pSmad3C/3L transformation. Phytomedicine 2023, 117, 154903. [Google Scholar] [CrossRef]
- Hu, X.; Jiao, F.; Zhang, L.; Jiang, Y. Dihydrotanshinone inhibits hepatocellular carcinoma by suppressing the JAK2/STAT3 pathway. Front. Pharmacol. 2021, 12, 654986. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, N.; Meng, Z.; Lu, S.; Peng, G. Pseudolaric acid B triggers cell apoptosis by activating AMPK/JNK/DRP1/mitochondrial fission pathway in hepatocellular carcinoma. Toxicology 2023, 493, 153556. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Sun, F.; Deng, K.; Lin, G.; Yin, W.; Chen, H.; Yang, D.; Liu, K.; Zhang, Y.; Huang, L. Mallotucin D, a Clerodane Diterpenoid from Croton crassifolius, Suppresses HepG2 Cell Growth via Inducing Autophagic Cell Death and Pyroptosis. Int. J. Mol. Sci. 2022, 23, 14217. [Google Scholar] [CrossRef]
- Chang, W.-T.; Bow, Y.-D.; Fu, P.-J.; Li, C.-Y.; Wu, C.-Y.; Chang, Y.-H.; Teng, Y.-N.; Li, R.-N.; Lu, M.-C.; Liu, Y.-C.; et al. A Marine Terpenoid, Heteronemin, Induces Both the Apoptosis and Ferroptosis of Hepatocellular Carcinoma Cells and Involves the ROS and MAPK Pathways. Oxidative Med. Cell. Longev. 2021, 2021, 7689045. [Google Scholar] [CrossRef]
- Liu, C.; Yang, S.; Wang, K.; Bao, X.; Liu, Y.; Zhou, S.; Liu, H.; Qiu, Y.; Wang, T.; Yu, H. Alkaloids from Traditional Chinese Medicine against hepatocellular carcinoma. Biomed. Pharmacother. 2019, 120, 109543. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, S.; Liu, J.; Fang, B.; Yao, J.; Cheng, B. Matrine inhibits the invasive and migratory properties of human hepatocellular carcinoma by regulating epithelial-mesenchymal transition. Mol. Med. Rep. 2018, 18, 911–919. [Google Scholar] [CrossRef]
- Li, P.; Liu, Y.; He, Q. Anisodamine Suppressed the Growth of Hepatocellular Carcinoma Cells, Induced Apoptosis and Regulated the Levels of Inflammatory Factors by Inhibiting NLRP3 Inflammasome Activation. Drug Des. Dev. Ther. 2020, 14, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Liu, Y.; Huang, J.; Yin, S.; Sheng, H.; Han, X.; Chen, Q.; Wang, T.; Chen, S.; Qiu, Y.; et al. Stachydrine hydrochloride inhibits hepatocellular carcinoma progression via LIF/AMPK axis. Phytomedicine 2022, 100, 154066. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Pan, L.; Wu, J.; Li, L.; Xu, H.; Yang, L.; Xu, K.; Wang, C. Cepharanthine inhibits hepatocellular carcinoma cell growth and proliferation by regulating amino acid metabolism and suppresses tumorigenesis in vivo. Int. J. Biol. Sci. 2021, 17, 4340–4352. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Cao, C.; Deng, Z.; Li, J.; Zhou, X.; Huang, Z.; Cen, C. Effects of matrine in combination with cisplatin on liver cancer. Oncol. Lett. 2020, 21, 12327. [Google Scholar] [CrossRef] [PubMed]
- Su, G.-F.; Huang, Z.-X.; Huang, D.-L.; Chen, P.-X.; Wang, Y.; Wang, Y.-F. Cepharanthine hydrochloride inhibits the Wnt/β-catenin/Hedgehog signaling axis in liver cancer. Oncol. Rep. 2022, 47, 8294. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Xia, Y.; Xu, P.; Zheng, W.; Gao, Y.; Xie, F.; Ji, Z. Veratramine suppresses human HepG2 liver cancer cell growth in vitro and in vivo by inducing autophagic cell death. Oncol. Rep. 2020, 44, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Hu, L.; Chen, Y.; Wang, F.; Xiao, Y.; Tong, Z.; Li, H.; Xiang, M.; Tong, Q.; Zhang, Y. Chaetocochin J exhibits anti-hepatocellular carcinoma effect independent of hypoxia. Bioorganic Chem. 2023, 139, 106701. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, D.; Wu, F.; Tu, J.; Song, J.; Xu, M.; Ji, J. Sophoridine suppresses lenvatinib-resistant hepatocellular carcinoma growth by inhibiting RAS/MEK/ERK axis via decreasing VEGFR2 expression. J. Cell. Mol. Med. 2020, 25, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Y.; Lin, J.; Jia, R.; An, T.; Dong, T.; Zhang, Y.; Yang, X. Cyclovirobuxine D Exerts Anticancer Effects by Suppressing the EGFR-FAK-AKT/ERK1/2-Slug Signaling Pathway in Human Hepatocellular Carcinoma. DNA Cell Biol. 2020, 39, 355–367. [Google Scholar] [CrossRef]
- Zheng, L.; Fang, S.; Chen, A.; Chen, W.; Qiao, E.; Chen, M.; Shu, G.; Zhang, D.; Kong, C.; Weng, Q.; et al. Piperlongumine synergistically enhances the antitumour activity of sorafenib by mediating ROS-AMPK activation and targeting CPSF7 in liver cancer. Pharmacol. Res. 2022, 177, 106140. [Google Scholar] [CrossRef]
- Kong, W.-S.; Shen, F.-X.; Xie, R.-F.; Zhou, G.; Feng, Y.-M.; Zhou, X. Bufothionine Induces Autophagy in H22 Hepatoma-bearing Mice by Inhibiting JAK2/STAT3 Pathway, a Possible Anti-cancer Mechanism of Cinobufacini. J. Ethnopharmacol. 2021, 270, 113848. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yang, Y.; Hu, Y.; Yang, R.; Huang, J.; Liu, Y.; Wu, Y.; Li, S.; Ma, C.; Humphries, F.; et al. Gasdermin D restricts anti-tumor immunity during PD-L1 checkpoint blockade. Cell Rep. 2022, 41, 111553. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, D.-H.; Lai, K.-D.; Su, H.; Lu, G.-S.; Wang, L.; Lv, J.-H. Ventilagolin Suppresses Migration, Invasion and Epithelial-Mesenchymal Transition of Hepatocellular Carcinoma Cells by Downregulating Pim-1. Drug Des. Dev. Ther. 2021, 15, 4885–4899. [Google Scholar] [CrossRef] [PubMed]
- Sufianova, G.; Gareev, I.; Beylerli, O.; Wu, J.; Shumadalova, A.; Sufianov, A.; Chen, X.; Zhao, S. Modern aspects of the use of natural polyphenols in tumor prevention and therapy. Front. Cell Dev. Biol. 2022, 10, 1011435. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, Y.; Yang, W.; Dong, J.; Li, L. Regulation of dietary polyphenols on cancer cell pyroptosis and the tumor immune microenvironment. Front. Nutr. 2022, 9, 974896. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Jian, P.; Zhang, Q.; Liang, W.; Zhou, K.; Hu, Q.; Liu, Y.; Liu, R.; Zhang, L. Tannins in Terminalia bellirica inhibit hepatocellular carcinoma growth by regulating EGFR-signaling and tumor immunity. Food Funct. 2021, 12, 3720–3739. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Nan, H.; Shi, N.; Hao, W.; Dong, J.; Chen, H. Chlorogenic acid inhibits proliferation in human hepatoma cells by suppressing noncanonical NF-κB signaling pathway and triggering mitochondrial apoptosis. Mol. Biol. Rep. 2021, 48, 2351–2364. [Google Scholar] [CrossRef]
- Han, L.; Wang, Y.; Sun, S. Curcumin inhibits proliferation of hepatocellular carcinoma cells through down regulation of DJ-1. Cancer Biomark. 2020, 29, 1–8. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Li, X. Curcumae Ameliorates Diethylnitrosamine-Induced Hepatocellular Carcinoma via Alteration of Oxidative Stress, Inflammation and Gut Microbiota. J. Inflamm. Res. 2021, 14, 5551–5566. [Google Scholar] [CrossRef]
- Dai, H.; Li, M.; Yang, W.; Sun, X.; Wang, P.; Wang, X.; Su, J.; Wang, X.; Hu, X.; Zhao, M. Resveratrol inhibits the malignant progression of hepatocellular carcinoma via MARCH1-induced regulation of PTEN/AKT signaling. Aging 2020, 12, 11717–11731. [Google Scholar] [CrossRef]
- Liu, G.; Shi, A.; Wang, N.; Li, M.; He, X.; Yin, C.; Tu, Q.; Shen, X.; Tao, Y.; Wang, Q.; et al. Polyphenolic Proanthocyanidin-B2 suppresses proliferation of liver cancer cells and hepatocellular carcinogenesis through directly binding and inhibiting AKT activity. Redox Biol. 2020, 37, 101701. [Google Scholar] [CrossRef]
- Na, W.; Ma, B.; Shi, S.; Chen, Y.; Zhang, H.; Zhan, Y.; An, H. Procyanidin B1, a novel and specific inhibitor of Kv10.1 channel, suppresses the evolution of hepatoma. Biochem. Pharmacol. 2020, 178, 114089. [Google Scholar] [CrossRef]
- Gong, H.; He, Z.; Peng, A.; Zhang, X.; Cheng, B.; Sun, Y.; Zheng, L.; Huang, K. Effects of several quinones on insulin aggregation. Sci. Rep. 2014, 4, srep05648. [Google Scholar] [CrossRef]
- Qin, B.; Zeng, Z.; Xu, J.; Shangwen, J.; Ye, Z.J.; Wang, S.; Wu, Y.; Peng, G.; Wang, Q.; Gu, W.; et al. Emodin inhibits invasion and migration of hepatocellular carcinoma cells via regulating autophagy-mediated degradation of snail and β-catenin. BMC Cancer 2022, 22, 671. [Google Scholar] [CrossRef]
- Khan, H.; Jia, W.; Yu, Z.; Zaib, T.; Feng, J.; Jiang, Y.; Song, H.; Bai, Y.; Yang, B.; Feng, H. Emodin succinyl ester inhibits malignant proliferation and migration of hepatocellular carcinoma by suppressing the interaction of AR and EZH2. Biomed. Pharmacother. 2020, 128, 110244. [Google Scholar] [CrossRef]
- Tadros, S.A.; Attia, Y.M.; Maurice, N.W.; Fahim, S.A.; Abdelwahed, F.M.; Ibrahim, S.; Badary, O.A. Thymoquinone Suppresses Angiogenesis in DEN-Induced Hepatocellular Carcinoma by Targeting miR-1-3p. Int. J. Mol. Sci. 2022, 23, 15904. [Google Scholar] [CrossRef]
- Yao, L.; Yan, D.; Jiang, B.; Xue, Q.; Chen, X.; Huang, Q.; Qi, L.; Tang, D.; Chen, X.; Liu, J. Plumbagin is a novel GPX4 protein degrader that induces apoptosis in hepatocellular carcinoma cells. Free Radic. Biol. Med. 2023, 203, 1–10. [Google Scholar] [CrossRef]
- Li, Y.; Tian, Y.; Zhong, W.; Wang, N.; Wang, Y.; Zhang, Y.; Zhang, Z.; Li, J.; Ma, F.; Zhao, Z.; et al. Artemisia argyi Essential Oil Inhibits Hepatocellular Carcinoma Metastasis via Suppression of DEPDC1 Dependent Wnt/β-catenin Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 664791. [Google Scholar] [CrossRef]
- Costa, E.V.; de Souza, C.A.S.; Galvão, A.F.C.; Silva, V.R.; Santos, L.d.S.; Dias, R.B.; Rocha, C.A.G.; Soares, M.B.P.; da Silva, F.M.A.; Koolen, H.H.F.; et al. Duguetia pycnastera Sandwith (Annonaceae) Leaf Essential Oil Inhibits HepG2 Cell Growth In Vitro and In Vivo. Molecules 2022, 27, 5664. [Google Scholar] [CrossRef]
- Lima, E.J.S.P.d.; Fontes, S.S.; Nogueira, M.L.; Silva, V.R.; Santos, L.d.S.; D’Elia, G.M.A.; Dias, R.B.; Sales, C.B.S.; Rocha, C.A.G.; Vannier-Santos, M.A.; et al. Essential oil from leaves of Conobea scoparioides (Cham. & Schltdl.) Benth. (Plantaginaceae) causes cell death in HepG2 cells and inhibits tumor development in a xenograft model. Biomed. Pharmacother. 2020, 129, 110402. [Google Scholar] [CrossRef]
- de Oliveira, F.P.; da C Rodrigues, A.C.B.; de Lima, E.J.S.P.; Silva, V.R.; de S Santos, L.; da Anunciação, T.A.; Nogueira, M.L.; Soares, M.B.P.; Dias, R.B.; Gurgel Rocha, C.A.; et al. Essential Oil from Bark of Aniba parviflora (Meisn.) Mez (Lauraceae) Reduces HepG2 Cell Proliferation and Inhibits Tumor Development in a Xenograft Model. Chem. Biodivers. 2021, 18, e2000938. [Google Scholar] [CrossRef]
- Nogueira, M.L.; Lima, E.J.S.P.d.; Adrião, A.A.X.; Fontes, S.S.; Silva, V.R.; Santos, L.d.S.; Soares, M.B.P.; Dias, R.B.; Rocha, C.A.G.; Costa, E.V.; et al. Cyperus articulatus L. (Cyperaceae) Rhizome Essential Oil Causes Cell Cycle Arrest in the G2/M Phase and Cell Death in HepG2 Cells and Inhibits the Development of Tumors in a Xenograft Model. Molecules 2020, 25, 2687. [Google Scholar] [CrossRef]
- Wang, Y.-F.; Zheng, Y.; Cha, Y.-Y.; Feng, Y.; Dai, S.-X.; Zhao, S.; Chen, H.; Xu, M. Essential oil of lemon myrtle (Backhousia citriodora) induces S-phase cell cycle arrest and apoptosis in HepG2 cells. J. Ethnopharmacol. 2023, 312, 116493. [Google Scholar] [CrossRef]
- Huang, X.-F.; Sheu, G.-T.; Chang, K.-F.; Huang, Y.-C.; Hung, P.-H.; Tsai, N.-M. Pogostemon cablin Triggered ROS-Induced DNA Damage to Arrest Cell Cycle Progression and Induce Apoptosis on Human Hepatocellular Carcinoma In Vitro and In Vivo. Molecules 2020, 25, 5639. [Google Scholar] [CrossRef]
- Wen, Y.; Zhou, X.; Lu, M.; He, M.; Tian, Y.; Liu, L.; Wang, M.; Tan, W.; Deng, Y.; Yang, X.; et al. Bclaf1 promotes angiogenesis by regulating HIF-1α transcription in hepatocellular carcinoma. Oncogene 2018, 38, 1845–1859. [Google Scholar] [CrossRef]
- Cappuyns, S.; Llovet, J.M. Combination Therapies for Advanced Hepatocellular Carcinoma: Biomarkers and Unmet Needs. Clin. Cancer Res. 2022, 28, 3405–3407. [Google Scholar] [CrossRef]
- Yao, H.; Liu, N.; Lin, M.C.; Zheng, J. Positive feedback loop between cancer stem cells and angiogenesis in hepatocellular carcinoma. Cancer Lett. 2016, 379, 213–219. [Google Scholar] [CrossRef]
- Cheng, W.; Cheng, Z.; Weng, L.; Xing, D.; Zhang, M. Asparagus Polysaccharide inhibits the Hypoxia-induced migration, invasion and angiogenesis of Hepatocellular Carcinoma Cells partly through regulating HIF1α/VEGF expression via MAPK and PI3K signaling pathway. J. Cancer 2021, 12, 3920–3929. [Google Scholar] [CrossRef]
- Yucui, L.; Hongxing, Z. Research Overview on the Mechanism of Traditional Chinese Medicine in Preventing and Treating Liver Cancer. J. Pract. Tradit. Chin. Intern. Med. 2023, 1–8. [Google Scholar]
- Su, W.; Wu, L.; Liang, Q.; Lin, X.; Xu, X.; Yu, S.; Lin, Y.; Zhou, J.; Fu, Y.; Gao, X.; et al. Extraction Optimization, Structural Characterization, and Anti-Hepatoma Activity of Acidic Polysaccharides From Scutellaria barbata D. Don. Front. Pharmacol. 2022, 13, 827782. [Google Scholar] [CrossRef]
- Li, Q.-Z.; Chen, Y.-Y.; Liu, Q.-P.; Feng, Z.-H.; Zhang, L.; Zhang, H. Cucurbitacin B suppresses hepatocellular carcinoma progression through inducing deoxyribonucleic acid (DNA) damage-dependent cell cycle arrest. Phytomedicine 2024, 126, 155177. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Sage J. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Fernald, K.; Kurokawa, M. Evading apoptosis in cancer. Trends Cell Biol. 2013, 23, 620–633. [Google Scholar] [CrossRef]
- Nie, C.; Wang, B.; Wang, B.; Lv, N.; Yu, R.; Zhang, E. Protopine triggers apoptosis via the intrinsic pathway and regulation of ROS/PI3K/Akt signalling pathway in liver carcinoma. Cancer Cell Int. 2021, 21, 396. [Google Scholar] [CrossRef] [PubMed]
- Ichimiya, T.; Yamakawa, T.; Hirano, T.; Yokoyama, Y.; Hayashi, Y.; Hirayama, D.; Wagatsuma, K.; Itoi, T.; Nakase, H. Autophagy and autophagy-related diseases: A review. Int. J. Mol. Sci. 2020, 21, 8974. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Zhang, J.; Ji, L.; Li, M.; Sun, X.; Feng, H.; Yu, Z.; Gao, Y. Current Perspective of Traditional Chinese Medicines and Active Ingredients in the Therapy of Hepatocellular Carcinoma. J. Hepatocell. Carcinoma 2022, 9, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, W.; Shi, X.; Ding, J.; Liu, W.; He, H.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef]
- Chen, X.; He, W.-t.; Hu, L.; Li, J.; Fang, Y.; Wang, X.; Xu, X.; Wang, Z.; Huang, K.; Han, J. Pyroptosis is driven by non-selective gasdermin-D pore and its morphology is different from MLKL channel-mediated necroptosis. Cell Res. 2016, 26, 1007–1020. [Google Scholar] [CrossRef]
- Sun, Z.; Ma, C.; Zhan, X. Ajmalicine induces the pyroptosis of hepatoma cells to exert the antitumor effect. J. Biochem. Mol. Toxicol. 2023, 38, e23614. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, M.; Zhao, L.; Geng, Y.; Li, G.; Chen, L.; Yu, J.; Yuan, H.; Zhang, H.; Yun, H. HBx-induced HSPA8 stimulates HBV replication and suppresses ferroptosis to support liver cancer progression. Cancer Res. 2023, 83, 1048–1061. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhou, C.; Jing, Q.; Li, Y.; Yang, J.; Yang, C.; Wang, L.; Hu, J.; Li, H.; Wang, H. FTH promotes the proliferation and renders the HCC cells specifically resist to ferroptosis by maintaining iron homeostasis. Cancer Cell Int. 2021, 21, 709. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. J. Gen. Physiol. 1927, 8, 519. [Google Scholar] [CrossRef] [PubMed]
- Cascone, T.; McKenzie, J.A.; Mbofung, R.M.; Punt, S.; Wang, Z.; Xu, C.; Williams, L.J.; Wang, Z.; Bristow, C.A.; Carugo, A. Increased tumor glycolysis characterizes immune resistance to adoptive T cell therapy. Cell Metab. 2018, 27, 977–987.e974. [Google Scholar] [CrossRef] [PubMed]
- Greene, L.I.; Bruno, T.C.; Christenson, J.L.; D’Alessandro, A.; Culp-Hill, R.; Torkko, K.; Borges, V.F.; Slansky, J.E.; Richer, J.K. A role for tryptophan-2, 3-dioxygenase in CD8 T-cell suppression and evidence of tryptophan catabolism in breast cancer patient plasma. Mol. Cancer Res. 2019, 17, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Fang, X.; Wang, H.; Li, D.; Wang, X. Ovarian cancer-intrinsic fatty acid synthase prevents anti-tumor immunity by disrupting tumor-infiltrating dendritic cells. Front. Immunol. 2018, 9, 2927. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hong, Y.; Jiang, L.; Yi, X.; Chen, Y.; Liu, L.; Chen, Z.; Wu, Y.; Cai, Z. Global Metabolomic and Lipidomic Analysis Reveal the Synergistic Effect of Bufalin in Combination with Cinobufagin against HepG2 Cells. J. Proteome Res. 2020, 19, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Alam, Z.; Devalaraja, S.; Li, M.; To, T.K.J.; Folkert, I.W.; Mitchell-Velasquez, E.; Dang, M.T.; Young, P.; Wilbur, C.J.; Silverman, M.A.; et al. Counter Regulation of Spic by NF-κB and STAT Signaling Controls Inflammation and Iron Metabolism in Macrophages. Cell Rep. 2020, 31, 107825. [Google Scholar] [CrossRef]
- Xie, X.; Shen, W.; Zhou, Y.; Ma, L.; Xu, D.; Ding, J.; He, L.; Shen, B.; Zhou, C. Characterization of a polysaccharide from Eupolyphaga sinensis walker and its effective antitumor activity via lymphocyte activation. Int. J. Biol. Macromol. 2020, 162, 31–42. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, H.; Zheng, F.; Liu, H.; Qiu, F.; Chen, Y.; Liang, C.-L.; Dai, Z. Resveratrol exerts antitumor effects by downregulating CD8+CD122+ Tregs in murine hepatocellular carcinoma. OncoImmunology 2020, 9, 1829346. [Google Scholar] [CrossRef]
- Anfray, C.; Ummarino, A.; Andón, F.T.; Allavena, P. Current Strategies to Target Tumor-Associated-Macrophages to Improve Anti-Tumor Immune Responses. Cells 2019, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Degroote, H.; Van Dierendonck, A.; Geerts, A.; Van Vlierberghe, H.; Devisscher, L. Preclinical and Clinical Therapeutic Strategies Affecting Tumor-Associated Macrophages in Hepatocellular Carcinoma. J. Immunol. Res. 2018, 2018, 7819520. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, Y.; Li, S.; Wang, T.; Wang, M.; Wu, H.; Zhang, M.; Luo, S.; Zhao, C.; Li, Q.; et al. Cinobufacini injection delays hepatocellular carcinoma progression by regulating lipid metabolism via SREBP1 signaling pathway and affecting macrophage polarization. J. Ethnopharmacol. 2024, 321, 117472. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, H.; Wang, J.; Gao, W.; Guan, Y.; Yi, S.; Ren, G.; Bai, C.; Zhu, Y. Research progress on the mechanism of Traditional Chinese Medicine regulating intestinal functionin the treatment of hepatocellular carcinoma based on the gut liver axis. Chin. J. Integr. Tradit. West. Med. Dig. 2024, 32, 222–225. [Google Scholar] [CrossRef]
- Liu, S.; Dong, W.; Yang, L.; Zhou, J.; Liu, D. Improvement and Mechanism of Ganoderma lucidum Polysaccharides and Its Flora Metabolites on Insulin Resistance in HepG2 Cells. Sci. Technol. Food Ind. 2023, 44, 314–321. [Google Scholar] [CrossRef]
- Ren, Z.; Chen, X.; Hong, L.; Zhao, X.; Cui, G.; Li, A.; Liu, Y.; Zhou, L.; Sun, R.; Shen, S. Nanoparticle conjugation of ginsenoside Rg3 inhibits hepatocellular carcinoma development and metastasis. Small 2020, 16, 1905233. [Google Scholar] [CrossRef] [PubMed]
- Ponziani, F.R.; Bhoori, S.; Castelli, C.; Putignani, L.; Rivoltini, L.; Del Chierico, F.; Sanguinetti, M.; Morelli, D.; Paroni Sterbini, F.; Petito, V. Hepatocellular carcinoma is associated with gut microbiota profile and inflammation in nonalcoholic fatty liver disease. Hepatology 2019, 69, 107–120. [Google Scholar] [CrossRef]
- Xie, G.; Wang, X.; Liu, P.; Wei, R.; Chen, W.; Rajani, C.; Hernandez, B.Y.; Alegado, R.; Dong, B.; Li, D. Distinctly altered gut microbiota in the progression of liver disease. Oncotarget 2016, 7, 19355. [Google Scholar] [CrossRef] [PubMed]
- Huo, R.; Yang, W.-J.; Liu, Y.; Liu, T.; Li, T.; Wang, C.-Y.; Pan, B.-S.; Wang, B.-L.; Guo, W. Stigmasterol: Remodeling Gut Microbiota and Suppressing Tumor Growth through Treg and CD8+ T Cells in Hepatocellular Carcinoma. Phytomedicine 2023, 129, 155225. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; He, R.; Zhong, H.; Liu, S.; Liu, X.; Hussain, M.; Sun, P. A cold-water extracted polysaccharide-protein complex from Grifola frondosa exhibited anti-tumor activity via TLR4-NF-κB signaling activation and gut microbiota modification in H22 tumor-bearing mice. Int. J. Biol. Macromol. 2023, 239, 124291. [Google Scholar] [CrossRef]
- Dahmani, R.; Just, P.-A.; Perret, C. The Wnt/β-catenin pathway as a therapeutic target in human hepatocellular carcinoma. Clin. Res. Hepatol. Gastroenterol. 2011, 35, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, T.; Sun, Y.; Huang, T.; Li, C.; Fu, Y.; Li, Y.; Li, C. p53-Mediated PI3K/AKT/mTOR Pathway Played a Role in PtoxDpt-Induced EMT Inhibition in Liver Cancer Cell Lines. Oxidative Med. Cell. Longev. 2019, 2019, 2531493. [Google Scholar] [CrossRef]
- Lei, R.-E.; Shi, C.; Zhang, P.-L.; Hu, B.-L.; Jiang, H.-X.; Qin, S.-Y. IL-9 promotes proliferation and metastasis of hepatocellular cancer cells by activating JAK2/STAT3 pathway. Int. J. Clin. Exp. Pathol. 2017, 10, 7940. [Google Scholar] [PubMed]
- Cai, H.; Zhang, Y.; Meng, F.; Cui, C.; Li, H.; Sui, M.; Zhang, H.; Lu, S. Preoperative Serum IL6, IL8, and TNF-α May Predict the Recurrence of Hepatocellular Cancer. Gastroenterol. Res. Pract. 2019, 2019, 6160783. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Xu, K.; Jiang, R.; Li, D.-Y.; Guo, X.-X.; Zhou, P.; Tang, J.-F.; Li, L.-S.; Zeng, D.; Hu, L.; et al. Evodiamine inhibits proliferation and promotes apoptosis of hepatocellular carcinoma cells via the Hippo-Yes-Associated Protein signaling pathway. Life Sci. 2020, 251, 117424. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Q.; Xie, S.; Wu, X.; Fu, J. Mechanism of action of cordycepin in the treatment of hepatocellular carcinoma via regulation of the Hippo signaling pathway. Food Sci. Hum. Wellness 2024, 13, 1040–1054. [Google Scholar] [CrossRef]
- Wu, D.-L.; Liao, Z.-D.; Chen, F.-F.; Zhang, W.; Ren, Y.-S.; Wang, C.-C.; Chen, X.-X.; Peng, D.-Y.; Kong, L.-Y. Benzophenones from Anemarrhena asphodeloides Bge. Exhibit Anticancer Activity in HepG2 Cells via the NF-κB Signaling Pathway. Molecules 2019, 24, 2246. [Google Scholar] [CrossRef] [PubMed]
- Nüsslein-Volhard, C.; Wieschaus, E. Mutations affecting segment number and polarity in Drosophila. Nature 1980, 287, 795–801. [Google Scholar] [CrossRef]
- Wu, J.; Tan, H.-Y.; Chan, Y.-T.; Lu, Y.; Feng, Z.; Yuan, H.; Zhang, C.; Feng, Y.; Wang, N. PARD3 drives tumorigenesis through activating Sonic Hedgehog signalling in tumour-initiating cells in liver cancer. J. Exp. Clin. Cancer Res. 2024, 43, 42. [Google Scholar] [CrossRef]
- Ekiert, H.M.; Szopa, A. Biological Activities of Natural Products. Molecules 2020, 25, 5769. [Google Scholar] [CrossRef]
- Wang, M.; Peng, P.; Chen, Z.; Deng, X. Nanoparticle delivery of active traditional Chinese medicine ingredients: A new strategy for the treatment of liver cancer. Curr. Pharm. Biotechnol. 2023, 24, 1630–1644. [Google Scholar] [CrossRef]
- Zheng, Y.; Kong, F.; Liu, S.; Liu, X.; Pei, D.; Miao, X. Membrane protein-chimeric liposome-mediated delivery of triptolide for targeted hepatocellular carcinoma therapy. Drug Deliv. 2021, 28, 2033–2043. [Google Scholar] [CrossRef]
- Shelash Al-Hawary, S.I.; Abdalkareem Jasim, S.; M Kadhim, M.; Jaafar Saadoon, S.; Ahmad, I.; Romero Parra, R.M.; Hasan Hammoodi, S.; Abulkassim, R.; M Hameed, N.; K Alkhafaje, W.; et al. Curcumin in the treatment of liver cancer: From mechanisms of action to nanoformulations. Phytother. Res. 2023, 37, 1624–1639. [Google Scholar] [CrossRef]
- Chen, X.-P.; Li, Y.; Zhang, Y.; Li, G.-W. Formulation, Characterization And Evaluation Of Curcumin- Loaded PLGA- TPGS Nanoparticles For Liver Cancer Treatment. Drug Des. Dev. Ther. 2019, 13, 3569–3578. [Google Scholar] [CrossRef]
- Zhu, M.; Shi, X.; Gong, Z.; Su, Q.; Yu, R.; Wang, B.; Yang, T.; Dai, B.; Zhan, Y.; Zhang, D.; et al. Cantharidin treatment inhibits hepatocellular carcinoma development by regulating the JAK2/STAT3 and PI3K/Akt pathways in an EphB4-dependent manner. Pharmacol. Res. 2020, 158, 104868. [Google Scholar] [CrossRef]
- Yao, H.; Zhao, J.; Wang, Z.; Lv, J.; Du, G.; Jin, Y.; Zhang, Y.; Song, S.; Han, G. Enhanced anticancer efficacy of cantharidin by mPEG-PLGA micellar encapsulation: An effective strategy for application of a poisonous traditional Chinese medicine. Colloids Surf. B Biointerfaces 2020, 196, 111285. [Google Scholar] [CrossRef]
- Wang, P.; Yang, J.; Zhu, Z.; Zhang, X. Marsdenia tenacissima: A Review of Traditional Uses, Phytochemistry and Pharmacology. Am. J. Chin. Med. 2018, 46, 1449–1480. [Google Scholar] [CrossRef]
- Wang, X.; Yan, Y.; Chen, X.; Zeng, S.; Qian, L.; Ren, X.; Wei, J.; Yang, X.; Zhou, Y.; Gong, Z.; et al. The Antitumor Activities of Marsdenia tenacissima. Front. Oncol. 2018, 8, 473. [Google Scholar] [CrossRef]
- Lin, S.; Sheng, Q.; Ma, X.; Li, S.; Xu, P.; Dai, C.; Wang, M.; Kang, H.; Dai, Z. Marsdenia tenacissima Extract Induces Autophagy and Apoptosis of Hepatocellular Cells via MIF/mToR Signaling. Evid.-Based Complement. Altern. Med. 2022, 2022, 7354700. [Google Scholar] [CrossRef]
- Li, L.; Zhang, W.; Desikan Seshadri, V.D.; Cao, G. Synthesis and characterization of gold nanoparticles from Marsdenia tenacissima and its anticancer activity of liver cancer HepG2 cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3029–3036. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, J.; Song, Y.; Zhang, T.; Wang, B. Combating liver cancer through GO-targeted biomaterials. Biomed. Mater. 2021, 16, 065003. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| No. | Name | Origin | In Vitro (a) or In Vivo (b) | Optimal Doses (/kg Body Weight) | Model | Potential Mechanism | References |
|---|---|---|---|---|---|---|---|
| 1 | Wild Cordyceps polysaccharides | Cordyceps sinensis | b | 100 mg | H22 tumor-bearing BALB/c mice | Modulates immunity. Modulation of IL-10/STAT3/Bcl2 and Cytoc/Caspase8/3 signaling pathways promotes apoptosis. | [33] |
| 2 | Low molecular weight polysaccharide from Grifola frondosa | Grifola frondosa | b | 200 mg | H22 tumor-bearing BALB/c mice | Modulates immune activity; promotes tumor cell apoptosis via the mitochondrial apoptotic pathway. | [37] |
| 3 | G. frondosa polysaccharide | G. frondosa | b | 200 mg | H22 tumor-bearing mice | Enhances immunity and induces cell cycle arrest at G0/G1 phase. | [37] |
| 4 | Pleurotus citrinopileatus polysaccharides | Pleurotus citrinopileatus | b | 300 mg | H22 tumor-bearing mice | Enhances immunity and induces cell cycle arrest in S phase. | [38] |
| 5 | Ganoderma lucidum polysaccharide | Ganoderma lucidum | a + b | 200 mg | Hepatic carcinoma Hepa1-6 allograft mice; RAW 264.7 and Hepa1-6 co-culture | Regulates macrophage polarization through activating MAPK/NF-κB signaling. | [39] |
| 6 | Bletilla striata polysaccharides | Bletilla striata | b | 200 mg | H22 tumor-bearing BALB/c mice | Modulates immunity and induces cell cycle arrest at G1 phase. | [40] |
| 7 | Acetylaminoglucan | / | b | 50 mg | H22 tumor-bearing BALB/c mice | Modulates immunity and induces cell cycle arrest in S phase. | [41] |
| 8 | Bupleurum chinense DC root polysaccharides | Bupleurum chinense DC root | b | 300 mg | H22 tumor-bearing Kunming mice | Induces S-phase block of the cell cycle and activates the mitochondrial pathway to induce apoptosis. | [42] |
| 9 | Angelica dahurica polysaccharide | Angelica dahurica | b | 300 mg | H22 tumor-bearing BALB/c mice | Induces apoptosis by cell cycle arrest in G0/G1 phase and reduction of cellular mitochondrial membrane potential. | [43] |
| 10 | Rhodiola rosea L. root polysaccharide | Rhodiola rosea L. root | b | 300 mg | H22 tumor-bearing mice | Induces cell cycle S-phase block by disrupting mitochondrial membrane potential and inducing apoptosis in tumor cells. | [44] |
| 11 | Dandelion polysaccharides | Dandelion | a + b | 400 mg | H22 tumor-bearing BALB/c mice; HepG2, Huh7, and Hepa1-6 cells | Down-regulates PI3K/AKT/mTOR pathway, inhibits cell proliferation and apoptosis, and enhances immune response. | [45] |
| 12 | Dandelion polysaccharide | Dandelion | a + b | 200 mg | H22 tumor-bearing BALB/c mice; HepG2, Huh7, Hepa1-6, and H22 cells | Inhibits IL-6-activated JAK-STAT signaling pathway; reduces hepcidin. | [46] |
| 13 | Eucommia folium polysaccharide | Eucommia folium | b | 300 mg | H22 tumor-bearing Kunming mice | Induces S-phase block of the cell cycle and apoptosis via the mitochondrial pathway. | [47] |
| 14 | Pseudostellaria heterophylla polysaccharide | Pseudostellaria heterophylla | a + b | 50 mg | H22 tumor-bearing C57BL/6 mice; RAW264.7 and Huh-7 cells | Regulates macrophage polarization through activating MAPK/NF-κB signaling. | [31] |
| 15 | Black fungus polysaccharide | Black fungus | a + b | 5 mg | HCCLM3 tumor-bearing BALB/c mice; HepG2, HCCLM3, and SK-Hep1 cells | Suppresses tumor cell proliferation by promoting DNA damage, attenuating DNA damage repair, and inhibiting DNA synthesis. | [35] |
| 16 | Fucoidan | Brown algae | a + b | 15 mg | MHCI297H tumor-bearing BALB/c mice; MHCC-97H and Hep3B cells | Causes lncRNA Linc00261 overexpression. | [48] |
| 17 | Ulva lactuca polysaccharide | Ulva lactuca | a + b | 300 mg | H22 liver cancer tumor-bearing mice; HepG2 and H22 cells | Inhibits tumor growth through modulation of gut microbial community and metabolism and modulation of miR-98-5p/ROS signaling pathway. | [34] |
| 18 | Sipunculus nudus polysaccharide | Sipunculus nudus | b | 200 mg | HepG2 tumor-bearing athymic nu/nu mice | Enhances immunity and induces apoptosis in tumor cells via the mitochondrial apoptotic pathway. | [49] |
| No. | Name | Origin | In Vitro (a) or In Vivo (b) | Optimal Doses (/kg Body Weight) | Model | Potential Mechanism | References |
|---|---|---|---|---|---|---|---|
| 1 | (R)-7,3′-dihydroxy-4′-methoxy-8-methylflavane | Resina Draconis | a + b | 20 mg | HepG2 tumor-bearing BALB/c mice and H22 tumor-bearing Kunming mice; HepG2 and SK-HEP-1 cells | Induction of apoptosis and G2/M phase block by DNA damage-driven upregulation of p21 expression in human hepatocellular carcinoma cells. | [50] |
| 2 | Neobavaisoflavone | Psoralea | a + b | 30 mg | HCCLM3 tumor-bearing BALB/c mice; HepG2 and HCCLM3 cells | Induction of cellular pyroptosis via the tom20/bax/caspase3/GSDME pathway. | [54] |
| 3 | Isorhamnetin | / | b | 100 mg | DEN + CCl4-induced HCC mice | Suppression of inflammation; regulates Akt and MAPKs to inhibit Nrf2 signaling; activates PPAR-γ and autophagy. | [55] |
| 4 | Sinensetin | citrus fruits. | b | 40 mg | HepG2/C3A tumor-bearing BALB/c nude mice | Inhibition of angiogenesis in hepatocellular carcinoma by regulating VEGF/VEGFR2/AKT signaling. | [51] |
| 5 | Icaritin | plants of the Epimedium genus | b | 70 mg | H22 tumor-bearing BALB/c mice | Prevention of MDSC generation via the attenuation of EMH. | [52] |
| 6 | Apigenin | parsley and chamomile | a + b | 50 mg | Huh7 tumor-bearing BALB/c mice; Hep3B cells | Downregulation of H19 induces inactivation of the Wnt/β-catenin signaling pathway. | [56] |
| 7 | Lysionotin | Lysionotus pauciflorus Maxim | a + b | 20 mg | HepG2 nude mice; HepG2 and SMMC-7721 cells | Induction of apoptosis in hepatocellular carcinoma cells via caspase-3-mediated mitochondrial pathway. | [57] |
| 8 | Kaempferide | Mountain apple root | a + b | 25 mg | N1S1 orthotopically injected SD rats; HepG2, Huh7, and N1S1 cells | Induction of apoptosis. | [58] |
| 9 | Wogonin | Scutellaria baicalensis | a + b | 50 mg | Orthotopically HCC-implantation mice; HepG2 cells | Inhibition of cell cycle progression by activating the glycogen synthase kinase-3 beta. | [59] |
| 10 | Total flavonoids | Oldenlandia diffusa | a + b | 0.4 mg | H22 tumor-bearing BALB/c mice; HepG2, Hep3B, and HCCLM3 H22 cells | Induction of apoptosis and autophagy of HCC cells by inducing endoplasmic reticulum (ER) stress response and activating PERK-eIF2α-ATF4 signaling pathway. | [60] |
| 11 | Isoliquiritigenin | roots of plants belonging to licorice | a + b | 50 mg | SMMC7721 tumor-bearing BALB/c mice; MHCC97-H and SMMC7721 cells | Downregulation of PI3K/AKT/mTOR pathway induces apoptosis and autophagy in hepatocellular carcinoma cells. | [61] |
| 12 | Prunetin | / | b | 100 μM | DEN-induced HCC mice | Regulation of the NF-κB/PI3K/AKT signaling pathway. | [62] |
| 13 | Baicalein and baicalin | Scutellaria baicalensis Georgi | a + b | 50 mg 80 mg | H22 tumor-bearing BALB/c mice; SMMC-7721 and HepG2 cells | Promotion of anti-tumor immunity by inhibiting PD-L1 expression. | [63] |
| 14 | Hesperidin | Citrus | b | 200 mg | DEN-induced HCC mice | Downregulation of the PI3K/Akt signaling pathway. | [64] |
| 15 | Cardamonin | cardamom | b | 50 mg | HepG2 tumor-bearing nude mice | Inhibition of NF-κB signaling pathway. | [65] |
| 16 | Safflower yellow | Carthamus tinctorius | a + b | 5 mg | DEN-induced HCC mice; Hepa1-6 cells | Inhibition of inflammatory response; promotes collagen degradation and modulates gut microbiota to improve immune microenvironment. | [66] |
| 17 | Daidzin | soybean | a + b | 100 mg | Hep3B tumor-bearing nude mice; HCCLM3 and Hep3B cells | Interference with hepatocellular carcinoma survival and migration via TPI1 and gluconeogenic pathways. | [67] |
| 18 | Salvigenin | Scutellariae barbatae Herba | a + b | 10 μg | Huh7 tumor-bearing BALB/c mice; Human HCC cell lines | Blocking of aerobic glycolysis in HCC cells by inhibiting the PI3K/AKT/GSK-3β pathway. | [68] |
| No. | Name | Origin | In Vitro (a) or In Vivo (b) | Optimal Doses (/kg Body Weight) | Model | Potential Mechanism | References |
|---|---|---|---|---|---|---|---|
| 1 | Ginsenoside Rh4 | ginseng | a + b | / | HCC tumor-bearing BALB/c mice; HUH7 and LM3 cells | Inhibition of inflammation-associated HCC progression by targeting HDAC4/IL-6/STAT3 signaling. | [74] |
| 2 | Octyl ester derivative of ginsenoside Rh2 (Rh2-O) | ginseng | b | 10 mg | H22 tumor-bearing Tlr4−/− mice | Anti-hepatocellular carcinoma through TLR4-mediated immunomodulation of lymphocytes. | [78] |
| 3 | Ginsenoside Compound K | ginseng | a + b | 5 mg | DEN-induced SD mice; Bel-7404 and Huh7 cells | Regulation of HIF-1α-mediated glycolysis by Bclaf1 inhibits cell proliferation. | [75] |
| 4 | Ginsenoside Rk3 | ginseng | a + b | 100 mg | HCC-LM3 tumor-bearing BALB/c mice; HepG2 and HCC-LM3 cells | Inhibition of cell proliferation and induction of cell cycle arrest. Promotion of cell autophagy and apoptosis via PI3K/AKT. | [76] |
| 5 | Ginsenoside Rh2 | ginseng | a + b | 30 μmol | H22 tumor-bearing C57BL/6 mice; Huh-7 and H22 cells | Suppression of hepatocellular carcinoma invasion and metastasis by inhibition of c-Jun/COX2/PGE2 pathway-mediated EMT. | [77] |
| 6 | Ginsenoside CK | ginseng | a + b | 60 mg | HCC-LM3 tumor-bearing BALB/c mice; HepG2, SMMC-7721, HCC-LM3, and Huh-7 cells | Inhibition of hypoxia-induced epithelial mesenchymal transition in hepatocellular carcinoma through the HIF-1α/NF-κB feedback pathway. | [81] |
| 7 | Saikosaponin-b2 (SS-b2) | Radix bupleuri | a + b | 20 mg | H22 tumor-bearing C57BL/6 mice; HepG2 and HUVECs cells | Inhibition of angiogenesis through inhibition of the VEGF/ERK/HIF-1α signaling pathway. | [82] |
| 8 | Saikosaponin-b2 (SS-b2) | Radix bupleuri | a + b | 6 mg | DEN-induced BALB/c mice; RAW 264.7 macrophages | Upregulation of STK4 inhibits IRAK1/NF-κB signaling axis effectively suppresses PLCs. | [83] |
| 9 | Astragaloside IV (AS-IV) | Astragalus membranaceus | a + b | 40 mg; 20 μmol/L | HT, genotype: pSmad3C+/−; HO, genotype: Nrf2−/−; lentivirus-transfected HepG2 cells | Amelioration of hepatocellular carcinoma by Nrf2-mediated transformation of pSmad3C/3L. | [84] |
| 10 | Dihydrotanshinon | Salvia miltiorrhiza Bunge | a + b | 15 mg; 4 μg/mL | SMMC7721 tumor-bearing Balb/c mice; HCCLM3, SMMC7721, Hep3B. and HepG2 cells | Promotion of apoptosis by blocking the JAK2/STAT3 pathway. | [85] |
| 11 | Betulinic acid | bark of several plants | a + b | 40 mg | HUH7 tumor-bearing BALB/c mice; HUH7 and HCCLM3 cells | Inhibition of hepatocellular carcinoma cell growth through activation of NCOA4-mediated ferritin phagocytosis pathway. | [79] |
| 12 | 2α, 3α, 23-trihydroxy-urs-12-en-28-oic acid, | Ganoderma lucidum | b | 400 mg | S180 and H22 tumor-bearing Kunming mice | Amelioration of lipid peroxidation. Down-regulation of Bcl-2 and up-regulation of Bax. Increase in intestinal flora richness and structure. | [80] |
| 13 | Pseudolaric acid B | Pseudolarix kaempferi | a + b | 20 mg | Hepa1–6 tumor-bearing C57BL/6 mice; Hepa1–6 cells | Triggering apoptosis through activation of the AMPK/JNK/dr P1/mitochondrial fission pathway. | [86] |
| 14 | Mallotucin D | Croton crassifolius | a | 20 μM | HepG2 cells | Induction of cellular pyroptosis. Inhibition of the PI3K/AKT/mTOR pathway activates mitochondrial autophagy. | [87] |
| 15 | Heteronemin | Hippospongia sp. | a | 20 μM | HA22T and HA59T cells | Induction of apoptosis via the caspase pathway. Induction of ferroptosis by down-regulation of GPX4. | [88] |
| No. | Name | Origin | In Vitro (a) or In Vivo (b) | Optimal Dose (/kg Body Weight) or Concentration | Model | Potential Mechanism | References |
|---|---|---|---|---|---|---|---|
| 1 | Matrine | Sophora flavescens Aition | b | 100 mg/kg matrine and 2 mg/kg cisplatin. | HepG2 tumor-bearing BALB/c mice | Promotion of apoptosis via suppression of survivin and activation of the caspase pathway. | [94] |
| 2 | Cepharanthine | Stephania cephalantha Hayata | a + b | 20 mg; 60 µM | Huh7 tumor-bearing BALB/c mice; Huh7 and HepG2 cells | Inhibition of the Wnt/β-catenin/Hedgehog signaling pathway. | [95] |
| 3 | Veratramine | Veratrum nigrum L. | a + b | 2 mg; 10, 20 and 40 µM | HepG2 tumor-bearing BALB/c mice; HepG2 cells | Activation of autophagy-mediated apoptosis through inhibition of the PI3K/Akt/mTOR signaling pathway. | [96] |
| 4 | Anisodamine | Anisodus | b | 200 mg | HepG2 tumor-bearing BALB/c mice | Inhibition of NLRP3 inflammatory vesicles; induction of apoptosis. | [91] |
| 5 | Chaetocochin J | Chaetomium sp. | a + b | 0.5 mg | HepG2 tumor-bearing nude mice; HepG2 and Hep3B cells | Inhibition of the PI3K/Akt/mTOR/p70S6K/4EBP1 pathway. Disruption of HIF-1α/p300 binding. | [97] |
| 6 | Sophoridine | Sophora alopecuroides L. | a + b | 50 mg | HepG2 LR tumor-bearing BALB/c mice; HepG2 and Huh7 cells | Inhibition of the RAS/MEK/ERK axis by decreasing VEGFR2 expression. | [98] |
| 7 | Stachydrine hydrochloride | Panzeria alaschanica Kupr | a + b | 30 mg | Patient-derived xenograft model; SMMC-7721 and HepG2 cells | Regulation of LIF/AMPK induced autophagy and senescence. | [92] |
| 8 | Cyclovirobuxine D | Buxus microphylla | a + b | 10 mg | HepG2 tumor-bearing BALB/c mice; HepG2 and HCCLM3 cells | Suppression of cell proliferation, migration, and invasion through inhibition of the EGFR-FAK-AKT/ERK1/2-Slug signaling pathway. | [99] |
| 9 | Piperlongumine | Piper longum L. | a + b | 10 mg/kg PL and 5 mg/kg sorafenib | HCCLM3 tumor-bearing BALB/c mice; HCCLM3 and SMMC7721 cells | Mediation of ROS-AMPK activation and targeting of CPSF7. | [100] |
| 10 | Bufothionine | cinobufacini injection | a + b | 343.35 μg/kg | H22-tumor-bearing Kunming mice; SMMC7721 cells | Induction of autophagy in HCC by inhibiting JAK2/STAT3 pathway. | [101] |
| 11 | Abrine | Abrus cantonments | a + b | 15 mg | HepG2 tumor-bearing BALB/c mice; HepG2 and Huh7 cells | Regulation of hepatocellular carcinoma cell growth and apoptosis through the KAT5/PD-L1 axis and regulation of T cell proliferation and activation. | [102] |
| 12 | Cepharanthine | Stephania cepharantha Hayata | a + b | 20 mg | Hep3B tumor-bearing nude mice; Hep3B and HCCLM3 cells | Inhibition of HCC cell proliferation by regulating amino acid metabolism and cholesterol metabolism; promotion of apoptosis and necrosis. | [93] |
| 13 | Ventilagolin | Ventilago leiocarpa Benth | b | 12 mg | SMMC-7721 tumor-bearing BALB/c-nu nude mice | Inhibition of HCC cell growth, migration, and invasion by regulating Pim-1 expression and EMT markers. | [103] |
| No. | Name | Origin | In Vitro (a) or In Vivo (b) | Optimal Dose (/kg Body Weight) or Concentration | Model | Potential Mechanism | References |
|---|---|---|---|---|---|---|---|
| 1 | Tannins | Terminalia bellirica | b | 2 g | HepG2 tumor-bearing ICR mice | Regulation of the EGFR signaling pathway and modulation of the immunosuppressive tumor microenvironment. | [106] |
| 2 | Proanthocyanidin-B2 | peanut skin | a + b | 300 mg | DEN + CCl4-induced HCC mice; Huh7 and Smmc-7721 cells | Inhibition of AKT leads to cell cycle arrest and tumor cell metabolic suppression. | [111] |
| 3 | Procyanidin B1 | grape seed | b | 15 mg | HepG2 tumor-bearing Balb/c mice | Inhibition of tumor growth in mice by inhibiting Kv10.1 current. | [112] |
| 4 | Chlorogenic acid | Eucommia ulmoides Oliver | a | 0, 30 and 300 µM | HepG2 and Huh-7cells | Inhibition of the NF-κB signaling pathway and triggering of mitochondrial apoptosis. | [107] |
| 5 | Curcumae | turmeric | b | 10 mg | DEN-induced albino Wistar rats | Regulation of oxidative stress, inflammatory response, and gut microbiota. | [109] |
| 6 | Curcumin | turmeric | a | 0, 5, 10 μM | SMMC-7721 and HepG2 cells | Down-regulation of DJ-1 inhibits cell proliferation. | [108] |
| 7 | Resveratrol | a + b | 100 mg | HepG2 tumor-bearing BALB/c nude mice; HepG2 and Hep3B cells | Down-regulation of MARCH1 regulates PTEN/AKT signaling. | [110] |
| No. | Name | Origin | In Vitro (a) or In Vivo (b) | Optimal Dose (/kg Body Weight) or Concentration | Model | Potential Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| 1 | Emodin | Rheum palmatum L. | a | 0, 25, 50, 75, and 100 μM | HepG2 cells | Induction of autophagy and inhibited EMT through inhibition of PI3K/AKT/mTOR and Wnt/β-catenin pathways. | [114] |
| 2 | Emodin succinyl ester | Rheum palmatum | b | 200 mg | DEN-induced HCC mice | Inhibition of cell proliferation and migration through inhibition of AR interaction with EZH2. | [115] |
| 3 | Thymoquinone | Nigella sativa | b | 5 mg | DEN-induced HCC mice | Inhibition of angiogenesis by targeting miR-1-3p. | [116] |
| 4 | Plumbagin | Plumbagin zeylanica L | b | 2 mg | HepG2 tumor-bearing Balb/c mice | Inhibition of USP31 activity leads to GPX4 protein degradation and apoptosis. | [117] |
| No. | Name | Origin | In Vitro (a) or In Vivo (b) | Optimal Dose (/kg Body Weight) or Concentration | Model | Potential Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| 1 | Artemisia argyi essential oil | Artemisia argyi | a + b | 230 mg | HepG2-Luc tumor-bearing BALB/c mice; HepG2 and SMMC-7721 cells | Inhibition of the Wnt/β-catenin signaling pathway suppresses metastasis of hepatocellular carcinoma. | [118] |
| 2 | D. pycnastera leaf essential oil | Duguetia pycnastera Sandwith | a + b | 40 mg | HepG2 tumor-bearing C.B-17 SCID mice; HepG2 cell | Inhibition of tumor growth. | [119] |
| 3 | Conobea scoparioides essential oil | Conobea scoparioides | a + b | 80 mg | HepG2 tumor-bearing C.B-17 SCID mice; HepG2 cell | Induction of apoptosis and inhibition of tumor growth. | [120] |
| 4 | Aniba parviflora essential oil | Aniba parviflora | a + b | 80 mg | HepG2 tumor-bearing C.B-17 SCID mice | Inhibition of tumor cell proliferation and inhibition of tumor growth. | [121] |
| 5 | C. articulatus rhizome essential oil | Cyperus articulatus L | b | 80 mg | HepG2 tumor-bearing C.B-17 SCID mice | Blocking the cell cycle and inducing apoptosis. | [122] |
| 6 | Essential oil of lemon myrtle | Lemon myrtle | a | 40.90 µM | HepG2 cells | Blocking the cell cycle and inducing apoptosis | [123] |
| 7 | Pogostemon cablin essential oils | Pogostemon cablin | a + b | 200 mg | HepG2 tumor-bearing Balb/c nude mice; HCC, SVEC, MDCK, and BNL CL.2 cells | Blocking the cell cycle and inducing apoptosis. | [124] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Yan, W.; Duan, H.; Wang, D.; Zhou, Y.; Feng, D.; Zheng, Y.; Zhou, S.; Liu, G.; Qin, X. Therapeutic Effects of Natural Products on Liver Cancer and Their Potential Mechanisms. Nutrients 2024, 16, 1642. https://doi.org/10.3390/nu16111642
Guo J, Yan W, Duan H, Wang D, Zhou Y, Feng D, Zheng Y, Zhou S, Liu G, Qin X. Therapeutic Effects of Natural Products on Liver Cancer and Their Potential Mechanisms. Nutrients. 2024; 16(11):1642. https://doi.org/10.3390/nu16111642
Chicago/Turabian StyleGuo, Jinhong, Wenjie Yan, Hao Duan, Diandian Wang, Yaxi Zhou, Duo Feng, Yue Zheng, Shiqi Zhou, Gaigai Liu, and Xia Qin. 2024. "Therapeutic Effects of Natural Products on Liver Cancer and Their Potential Mechanisms" Nutrients 16, no. 11: 1642. https://doi.org/10.3390/nu16111642
APA StyleGuo, J., Yan, W., Duan, H., Wang, D., Zhou, Y., Feng, D., Zheng, Y., Zhou, S., Liu, G., & Qin, X. (2024). Therapeutic Effects of Natural Products on Liver Cancer and Their Potential Mechanisms. Nutrients, 16(11), 1642. https://doi.org/10.3390/nu16111642