
Genome-Wide Admixture and Association Study of Serum Selenium Deficiency to Identify Genetic Variants Indirectly Linked to Selenium Regulation in Brazilian Adults
 , , ,
, , ,  ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethic Statement
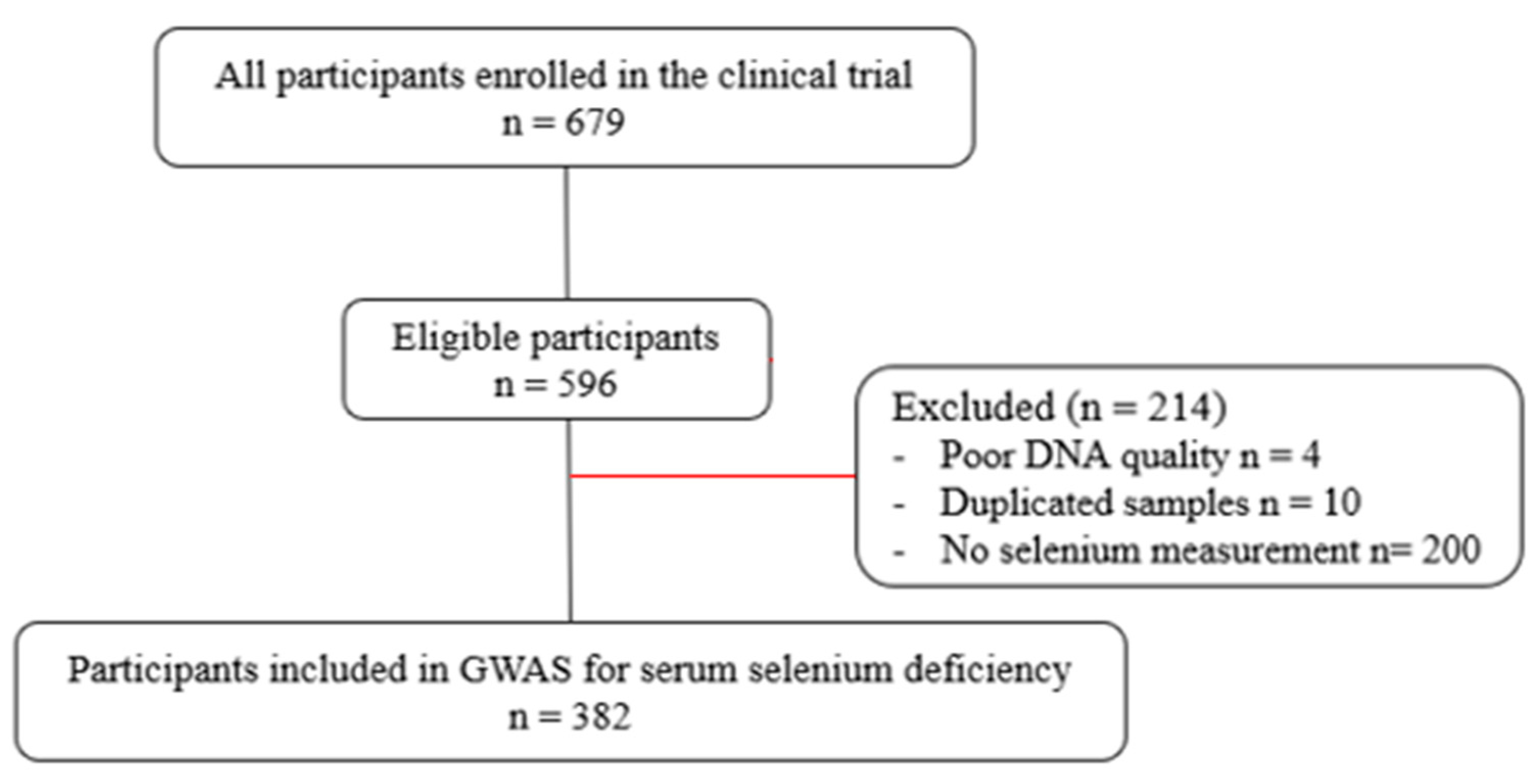
2.2. Study Population
2.3. Anthropometric Evaluation
2.4. Determination of Se Levels
2.5. Selenium Intake
2.6. Exploratory Analysis
2.7. Genotype Data
2.8. Genotype Data Processing and Association Analysis
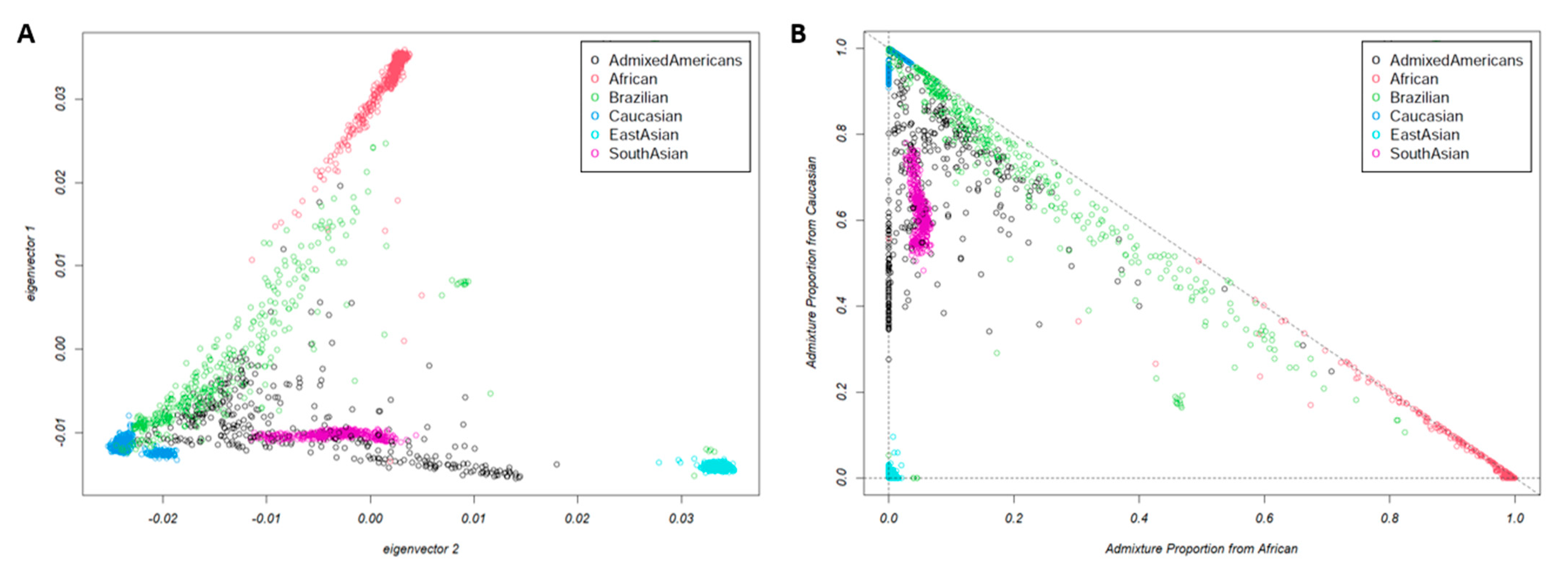
2.9. Admixture Analysis
3. Results
3.1. Population Characteristics and Structure
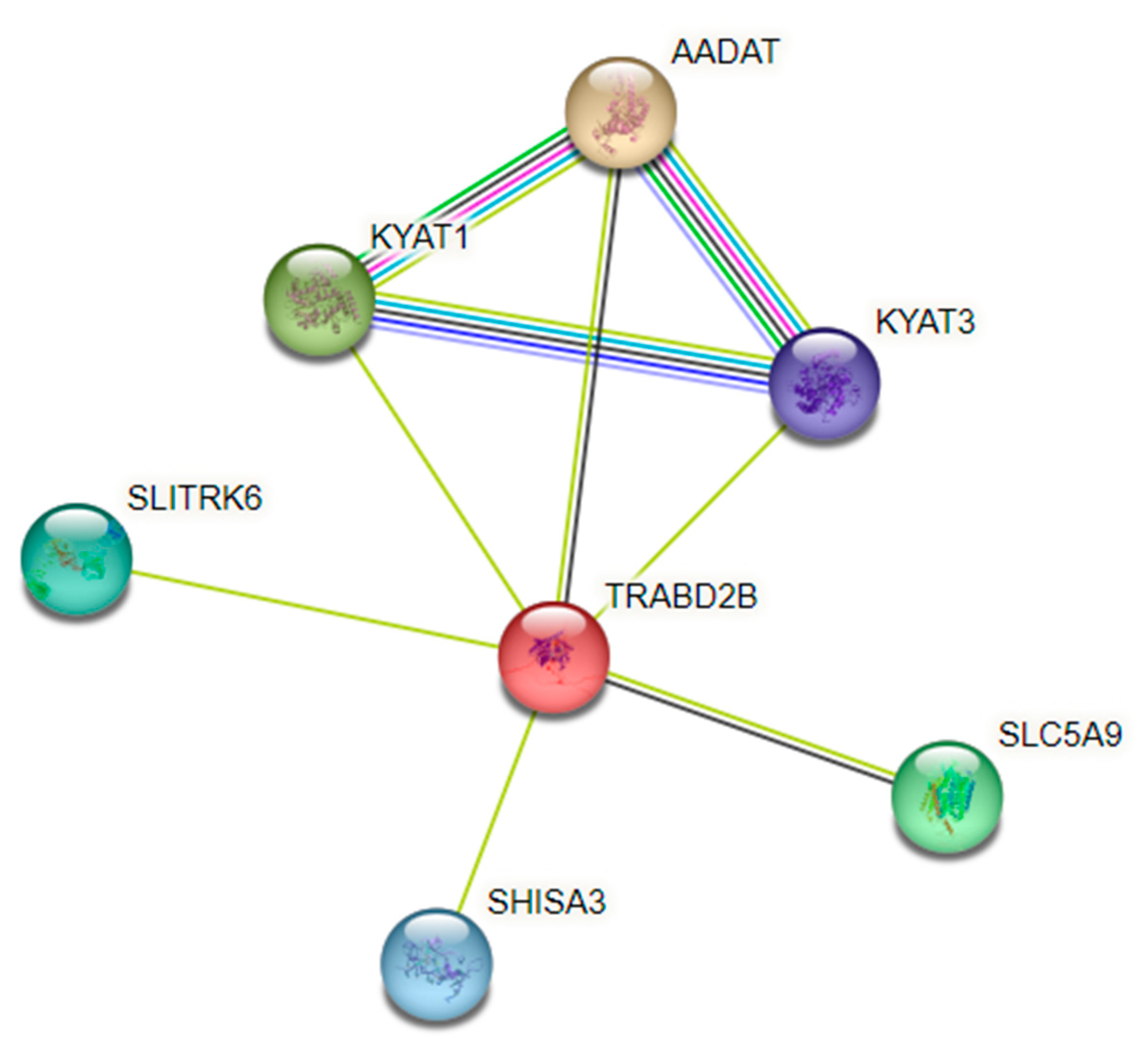
3.2. GWAS
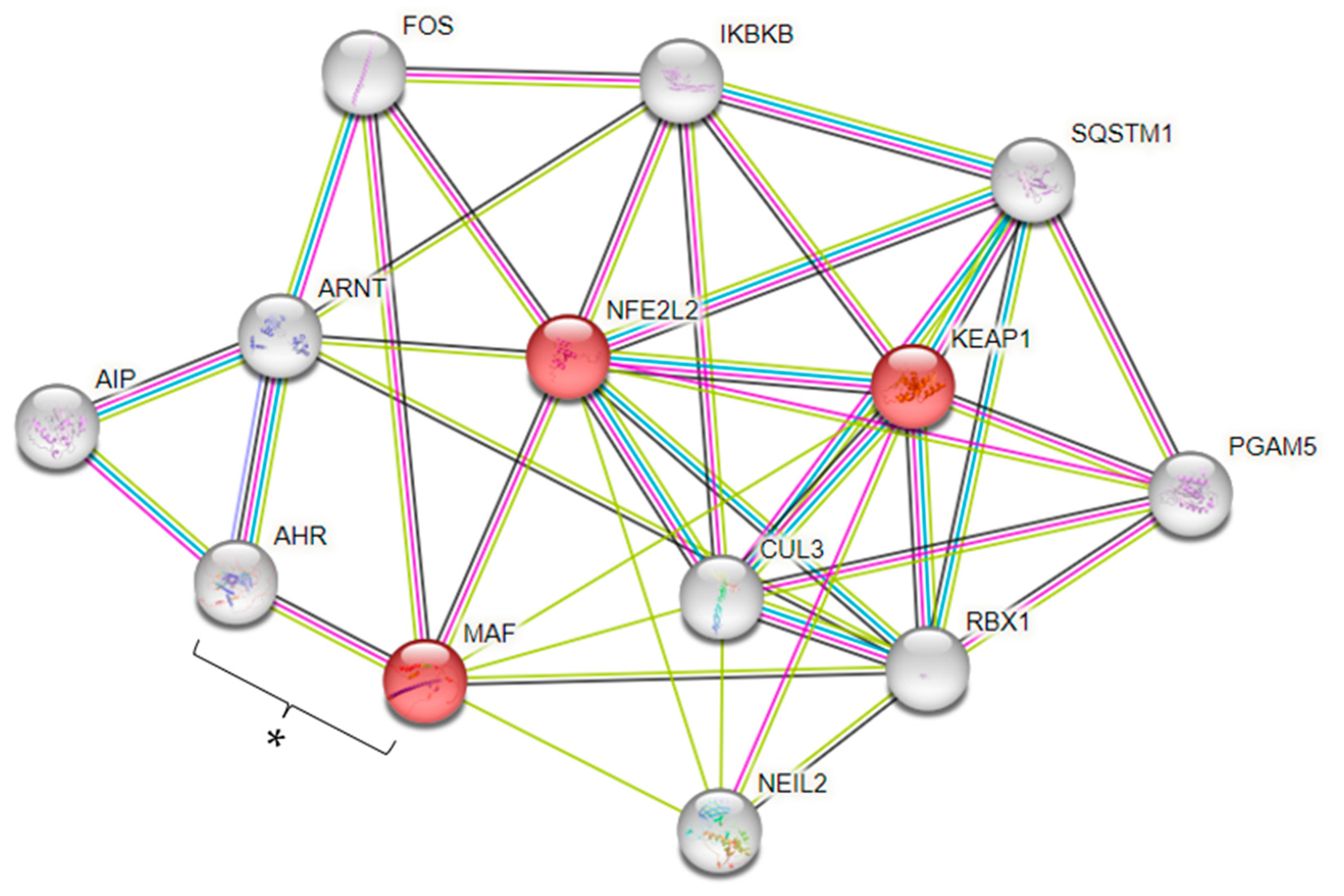
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kieliszek, M. Selenium–Fascinating Microelement, Properties and Sources in Food. Molecules 2019, 24, 1298. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Hornick, J.L.; Istasse, L.; Dufrasne, I. Selenium in the Environment, Metabolism and Involvement in Body Functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef]
- Reszka, E.; Jablonska, E.; Gromadzinska, J.; Wasowicz, W. Relevance of Selenoprotein Transcripts for Selenium Status in Humans. Genes Nutr. 2012, 7, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium Intake, Status, and Health: A Complex Relationship. Hormones 2020, 19, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Shimada, B.K.; Swanson, S.; Toh, P.; Seale, L.A. Metabolism of Selenium, Selenocysteine, and Selenoproteins in Ferroptosis in Solid Tumor Cancers. Biomolecules 2022, 12, 1581. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, L.M.; Navarro, A.M.; Seale, L.A. Intersection between Obesity, Dietary Selenium, and Statin Therapy in Brazil. Nutrients 2021, 13, 2027. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Hsu, L.; Harrison, T.; King, I.B.; Stürup, S.; Song, X.; Duggan, D.; Liu, Y.; Hutter, C.; Chanock, S.J.; et al. Genome-Wide Association Study of Serum Selenium Concentrations. Nutrients 2013, 5, 1706. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L. Selenium Deficiency Due to Diet, Pregnancy, Severe Illness, or COVID-19—A Preventable Trigger for Autoimmune Disease. Int. J. Mol. Sci. 2021, 22, 8532. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, M.C.; Fornage, M.; Foy, M.; Xun, P.; Gladyshev, V.N.; Morris, S.; Chasman, D.I.; Hu, F.B.; Rimm, E.B.; Kraft, P.; et al. Genome-Wide Association Study of Selenium Concentrations. Hum. Mol. Genet. 2015, 24, 1469–1477. [Google Scholar] [CrossRef]
- White, L.; Romagné, F.; Müller, E.; Erlebach, E.; Weihmann, A.; Parra, G.; Andrés, A.M.; Castellano, S. Genetic Adaptation to Levels of Dietary Selenium in Recent Human History. Mol. Biol. Evol. 2015, 32, 1507–1518. [Google Scholar] [CrossRef]
- Batai, K.; Trejo, M.J.; Chen, Y.; Kohler, L.N.; Lance, P.; Ellis, N.A.; Cornelis, M.C.; Chow, H.H.S.; Hsu, C.H.; Jacobs, E.T. Genome-Wide Association Study of Response to Selenium Supplementation and Circulating Selenium Concentrations in Adults of European Descent. J. Nutr. 2021, 151, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.M.; Zheng, J.P.; Yang, D.; Liu, Z.F.; Li, Z.; Hu, Z.Z.; Li, Z.N. Recessive/Dominant Model: Alternative Choice in Case-Control-Based Genome-Wide Association Studies. PLoS ONE 2021, 16, e0254947. [Google Scholar] [CrossRef] [PubMed]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-Wide Association Studies. Nat. Rev. Methods Prim. 2021, 1, 59. [Google Scholar] [CrossRef]
- Evans, D.M.; Zhu, G.; Dy, V.; Heath, A.C.; Madden, P.A.F.; Kemp, J.P.; McMahon, G.; Pourcain, B.S.; Timpson, N.J.; Golding, J.; et al. Genome-Wide Association Study Identifies Loci Affecting Blood Copper, Selenium, and Zinc. Hum. Mol. Genet. 2013, 22, 3998–4006. [Google Scholar] [CrossRef] [PubMed]
- Escher, L.M.; Naslavsky, M.S.; Scliar, M.O.; Duarte, Y.A.O.; Zatz, M.; Nunes, K.; Oliveira, S.F. Challenges in Selecting Admixture Models and Marker Sets to Infer Genetic Ancestry in a Brazilian Admixed Population. Sci. Rep. 2022, 12, 21240. [Google Scholar] [CrossRef]
- Giolo, S.R.; Soler, J.M.P.; Greenway, S.C.; Almeida, M.A.A.; De Andrade, M.; Seidman, J.G.; Seidman, C.E.; Krieger, J.E.; Pereira, A.C. Brazilian Urban Population Genetic Structure Reveals a High Degree of Admixture. Eur. J. Hum. Genet. 2012, 20, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Batista, B.L.; Rodrigues, J.L.; Nunes, J.A.; de Oliveira Souza, V.C.; Barbosa, F. Exploiting Dynamic Reaction Cell Inductively Coupled Plasma Mass Spectrometry (DRC-ICP-MS) for Sequential Determination of Trace Elements in Blood Using a Dilute-and-Shoot Procedure. Anal. Chim. Acta 2009, 639, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Troyanskaya, O.; Cantor, M.; Sherlock, G.; Brown, P.; Hastie, T.; Tibshirani, R.; Botstein, D.; Altman, R.B. Missing Value Estimation Methods for DNA Microarrays. Bioinformatics 2001, 17, 520–525. [Google Scholar] [CrossRef]
- Granato, I.S.C.; Galli, G.; de Oliveira Couto, E.G.; e Souza, M.B.; Mendonça, L.F.; Fritsche-Neto, R. SnpReady: A Tool to Assist Breeders in Genomic Analysis. Mol. Breed. 2018, 38, 102. [Google Scholar] [CrossRef]
- Wigginton, J.E.; Cutler, D.J.; Abecasis, G.R. A Note on Exact Tests of Hardy-Weinberg Equilibrium. Am. J. Hum. Genet. 2005, 76, 887–893. [Google Scholar] [CrossRef]
- González, J.R.; Armengol, L.; Solé, X.; Guinó, E.; Mercader, J.M.; Estivill, X.; Moreno, V. SNPassoc: An R Package to Perform Whole Genome Association Studies. Bioinformatics 2007, 23, 654–655. [Google Scholar] [CrossRef]
- González, J.R.; Carrasco, J.L.; Dudbridge, F.; Armengol, L.; Estivill, X.; Moreno, V. Maximizing Association Statistics over Genetic Models. Genet. Epidemiol. 2008, 32, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; Von Mering, C.; et al. STRING v9.1: Protein-Protein Interaction Networks, with Increased Coverage and Integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Gogarten, S.M.; Lawrence, M.; Stilp, A.; Conomos, M.P.; Weir, B.S.; Laurie, C.; Levine, D. SeqArray-a Storage-Efficient High-Performance Data Format for WGS Variant Calls. Bioinformatics 2017, 33, 2251–2257. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Levine, D.; Shen, J.; Gogarten, S.M.; Laurie, C.; Weir, B.S. A High-Performance Computing Toolset for Relatedness and Principal Component Analysis of SNP Data. Bioinformatics 2012, 28, 3326–3328. [Google Scholar] [CrossRef] [PubMed]
- Auton, A.; Abecasis, G.R.; Altshuler, D.M.; Durbin, R.M.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Donnelly, P.; Eichler, E.E.; Flicek, P.; et al. A Global Reference for Human Genetic Variation. Nature 2015, 526, 68. [Google Scholar] [PubMed]
- Meyer, H. RStudio “plinkQC”—Genotype Quality Control with PlinkQC. RStudio. 2018. Available online: https://cran.r-project.org/web/packages/plinkQC/index.html (accessed on 22 October 2023).
- Rasmussen, L.B.; Hollenbach, B.; Laurberg, P.; Carlé, A.; Hög, A.; Jørgensen, T.; Vejbjerg, P.; Ovesen, L.; Schomburg, L. Serum Selenium and Selenoprotein P Status in Adult Danes—8-Year Followup. J. Trace Elem. Med. Biol. 2009, 23, 265–271. [Google Scholar] [CrossRef]
- Pritchett, N.R.; Burgert, S.L.; Murphy, G.A.; Brockman, J.D.; White, R.E.; Lando, J.; Chepkwony, R.; Topazian, M.D.; Abnet, C.C.; Dawsey, S.M.; et al. Cross-Sectional Study of Serum Selenium Concentration and Esophageal Squamous Dysplasia in Western Kenya. BMC Cancer 2017, 17, 835. [Google Scholar] [CrossRef]
- Fadista, J.; Manning, A.K.; Florez, J.C.; Groop, L. The (in)Famous GWAS P-Value Threshold Revisited and Updated for Low-Frequency Variants. Eur. J. Hum. Genet. 2016, 24, 1202–1205. [Google Scholar] [CrossRef]
- Kaler, A.S.; Purcell, L.C. Estimation of a Significance Threshold for Genome-Wide Association Studies. BMC Genom. 2019, 20, 618. [Google Scholar] [CrossRef]
- Chen, Z.; Boehnke, M.; Wen, X.; Mukherjee, B. Revisiting the Genome-Wide Significance Threshold for Common Variant GWAS. G3 Genes Genomes Genet. 2021, 11, jkaa056. [Google Scholar] [CrossRef]
- Kaibara, F.S.; de Araujo, T.K.; Araujo, P.A.O.R.A.; Alvim, M.K.M.; Yasuda, C.L.; Cendes, F.; Lopes-Cendes, I.; Secolin, R. Association Analysis of Candidate Variants in Admixed Brazilian Patients With Genetic Generalized Epilepsies. Front. Genet. 2021, 12, 672304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; MacDonald, B.T.; Gao, H.; Shamashkin, M.; Coyle, A.J.; Martinez, R.V.; He, X. Characterization of Tiki, a New Family of Wnt-Specific Metalloproteases. J. Biol. Chem. 2016, 291, 2435–2443. [Google Scholar] [CrossRef]
- Kipp, A.; Banning, A.; van Schothorst, E.M.; Méplan, C.; Schomburg, L.; Evelo, C.; Coort, S.; Gaj, S.; Keijer, J.; Hesketh, J.; et al. Four Selenoproteins, Protein Biosynthesis, and Wnt Signalling Are Particularly Sensitive to Limited Selenium Intake in Mouse Colon. Mol. Nutr. Food Res. 2009, 53, 1561–1572. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Kipp, A.P. Selenium in the Redox Regulation of the Nrf2 and the Wnt Pathway. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2013; Volume 527. [Google Scholar]
- Korbut, E.; Ptak-Belowska, A.; Brzozowski, T. Inhibitory Effect of Selenomethionine on Carcinogenesis in the Model of Human Colorectal Cancer in Vitro and Its Link to the Wnt/β-Catenin Pathway. Acta Biochim. Pol. 2018, 65, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jin, Z.; Chen, M.; Duan, D.; Lammi, M.J.; Guo, X.; Chang, Y. Inhibiting the Aberrant Activation of Wnt/β-Catenin Signaling by Selenium Supplementation Ameliorates Deoxynivalenol-Induced Toxicity and Catabolism in Chondrocytes. J. Cell Physiol. 2020, 235, 4434–4442. [Google Scholar] [CrossRef]
- Grainger, S.; Willert, K. Mechanisms of Wnt Signaling and Control. Wiley Interdiscip. Rev. Syst. Biol. Med. 2018, 10, e1422. [Google Scholar] [CrossRef]
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/β-Catenin Signalling: Function, Biological Mechanisms, and Therapeutic Opportunities. Signal Transduct. Target. Ther. 2022, 7, 3. [Google Scholar] [CrossRef]
- Pinto, J.T.; Krasnikov, B.F.; Alcutt, S.; Jones, M.E.; Dorai, T.; Villar, M.T.; Artigues, A.; Li, J.; Cooper, A.J.L. Kynurenine Aminotransferase III and Glutamine Transaminase L Are Identical Enzymes That Have Cysteine S-Conjugate β-Lyase Activity and Can Transaminate L-Selenomethionine. J. Biol. Chem. 2014, 289, 30950–30961. [Google Scholar] [CrossRef]
- Selvam, A.K.; Björnstedt, M. A Novel Assay Method to Determine the β-Elimination of Se-Methylselenocysteine to Monomethylselenol by Kynurenine Aminotransferase 1. Antioxidants 2020, 9, 139. [Google Scholar] [CrossRef]
- Teumer, A.; Chaker, L.; Groeneweg, S.; Li, Y.; Di Munno, C.; Barbieri, C.; Schultheiss, U.T.; Traglia, M.; Ahluwalia, T.S.; Akiyama, M.; et al. Genome-Wide Analyses Identify a Role for SLC17A4 and AADAT in Thyroid Hormone Regulation. Nat. Commun. 2018, 9, 4455. [Google Scholar] [CrossRef] [PubMed]
- Gorini, F.; Sabatino, L.; Pingitore, A.; Vassalle, C. Selenium: An Element of Life Essential for Thyroid Function. Molecules 2021, 26, 7084. [Google Scholar] [CrossRef] [PubMed]
- Rawal, R.; Teumer, A.; Völzke, H.; Wallaschofski, H.; Ittermann, T.; Åsvold, B.O.; Bjøro, T.; Greiser, K.H.; Tiller, D.; Werdan, K.; et al. Meta-Analysis of Two Genome-Wide Association Studies Identifies Four Genetic Loci Associated with Thyroid Function. Hum. Mol. Genet. 2012, 21, 3275–3282. [Google Scholar] [CrossRef] [PubMed]
- Figlioli, G.; Köhler, A.; Chen, B.; Elisei, R.; Romei, C.; Cipollini, M.; Cristaudo, A.; Bambi, F.; Paolicchi, E.; Hoffmann, P.; et al. Novel Genome-Wide Association Study-Based Candidate Loci for Differentiated Thyroid Cancer Risk. J. Clin. Endocrinol. Metab. 2014, 99, E2084–E2092. [Google Scholar] [CrossRef] [PubMed]
- Hwangbo, Y.; Park, Y.J. Genome-Wide Association Studies of Autoimmune Thyroid Diseases, Thyroid Function, and Thyroid Cancer. Endocrinol. Metab. 2018, 33, 175. [Google Scholar] [CrossRef] [PubMed]
- Mittag, J.; Behrends, T.; Hoefig, C.S.; Vennström, B.; Schomburg, L. Thyroid Hormones Regulate Selenoprotein Expression and Selenium Status in Mice. PLoS ONE 2010, 5, e12931. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Lossow, K.; Kopp, J.F.; Schwerdtle, T.; Kipp, A.P. Crosstalk of Nrf2 with the Trace Elements Selenium, Iron, Zinc, and Copper. Nutrients 2019, 11, 2112. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Wang, R.; Long, T.; Xu, Y.; Zhang, Y.; Peng, R.; Zhang, X.; Guo, H.; Yang, H.; Wu, T.; et al. NRF2 Genetic Polymorphism Modifies the Association of Plasma Selenium Levels with Incident Coronary Heart Disease among Individuals with Type 2 Diabetes. Diabetes 2022, 71, 2009–2019. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Peng, V.; Sudan, R.; Ulezko Antonova, A.; Di Luccia, B.; Ohara, T.E.; Fachi, J.L.; Grajales-Reyes, G.E.; Jaeger, N.; Trsan, T.; et al. Repression of the Aryl-Hydrocarbon Receptor Prevents Oxidative Stress and Ferroptosis of Intestinal Intraepithelial Lymphocytes. Immunity 2023, 56, 797–812. [Google Scholar] [CrossRef]
- Shen, H.; Li, M.; Wang, B.; Lai, I.K.; Robertson, L.W.; Ludewig, G. Dietary Antioxidants (Selenium and N-Acetylcysteine) Modulate Paraoxonase 1 (PON1) in PCB 126-Exposed Rats. Environ. Sci. Pollut. Res. 2014, 21, 6384–6399. [Google Scholar] [CrossRef]
- Ge, J.; Guo, K.; Huang, Y.; Morse, P.D.; Zhang, C.; Lv, M.W.; Li, J.L. Comparison of Antagonistic Effects of Nanoparticle-Selenium, Selenium-Enriched Yeast and Sodium Selenite against Cadmium-Induced Cardiotoxicity via AHR/CAR/PXR/Nrf2 Pathways Activation. J. Nutr. Biochem. 2022, 105, 108992. [Google Scholar] [CrossRef] [PubMed]
- Aderao, G.N.; Jadhav, S.E.; Pattanaik, A.K.; Gupta, S.K.; Ramakrishnan, S.; Lokesha, E.; Chaudhary, P.; Vaswani, S.; Singh, A.; Panigrahi, M.; et al. Dietary Selenium Levels Modulates Antioxidant, Cytokine and Immune Response and Selenoproteins MRNA Expression in Rats under Heat Stress Condition. J. Trace Elem. Med. Biol. 2023, 75, 127105. [Google Scholar] [CrossRef] [PubMed]
- Kose, S.; Furuta, M.; Imamoto, N. Hikeshi, a Nuclear Import Carrier for Hsp70s, Protects Cells from Heat Shock-Induced Nuclear Damage. Cell 2012, 149, 578–589. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | Chr: Position | Gene | HWE * (p-Value) | MAF (%) | Allele | Mixed-Model # (p-Value) | Max-Stat (p-Value) |
|---|---|---|---|---|---|---|---|
| rs1561573 | 1:48227943 | TRABD2B | 0.1183 | 48.8 | C/T | 6.60 × 10−5 | 3.08 × 10−4 |
| rs806792 | 6:26198916 | H3C4 | 0.0892 | 10.2 | A/G | 1.72 × 10−4 | 4.57 × 10−5 |
| rs9383382 | 6:10067722 | OFCC1 | 0.3704 | 22.1 | T/C | 7.56 × 10−6 | 6.25 × 10−5 |
| rs4478892 | 10:55634794 | PCDH15 | 0.9051 | 31.1 | G/A | 1.18 × 10−4 | 3.72 × 10−4 |
| rs6592284 | 11:86055901 | HIKESHI | 0.4897 | 18.3 | T/C | 3.42 × 10−4 | 2.35 × 10−5 |
| rs2201161 | 12:99631219 | ANKS1B | 0.2116 | 45.1 | A/G | 6.11 × 10−6 | 1.70 × 10−6 |
| rs10444656 | 13:24671601 | SPATA13 | 0.6674 | 39.6 | T/C | 1.27 × 10−4 | 1.31 × 10−4 |
| rs1241636 | 14:25354440 | STXBP6 | 1.0000 | 44.4 | C/T | 2.26 × 10−4 | 1.95 × 10−4 |
| rs10483637 | 14:55157430 | SAMD4A | 0.5579 | 15.6 | C/T | 1.01 × 10−4 | 6.67 × 10−5 |
| rs8017018 | 14:79505543 | NRXN3 | 1.0000 | 45.6 | T/C | 8.53 × 10−5 | 7.11 × 10−5 |
| rs425664 | 16:79282227 | MAF | 0.6023 | 44.7 | A/C | 1.67 × 10−4 | 8.04 × 10−5 |
| rs6509701 | 19:53384185 | ZNF320 | 0.4966 | 34.9 | C/T | 4.27 × 10−5 | 2.11 × 10−5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moriguchi Watanabe, L.; Sousa, L.; Couto, F.M.; Noronha, N.Y.; de Souza Pinhel, M.A.; da Silva Carvalho, G.F.; da Silva Rodrigues, G.; Bueno Júnior, C.R.; Kulikowski, L.D.; Barbosa Júnior, F.; et al. Genome-Wide Admixture and Association Study of Serum Selenium Deficiency to Identify Genetic Variants Indirectly Linked to Selenium Regulation in Brazilian Adults. Nutrients 2024, 16, 1627. https://doi.org/10.3390/nu16111627
Moriguchi Watanabe L, Sousa L, Couto FM, Noronha NY, de Souza Pinhel MA, da Silva Carvalho GF, da Silva Rodrigues G, Bueno Júnior CR, Kulikowski LD, Barbosa Júnior F, et al. Genome-Wide Admixture and Association Study of Serum Selenium Deficiency to Identify Genetic Variants Indirectly Linked to Selenium Regulation in Brazilian Adults. Nutrients. 2024; 16(11):1627. https://doi.org/10.3390/nu16111627
Chicago/Turabian StyleMoriguchi Watanabe, Ligia, Lisete Sousa, Francisco M. Couto, Natália Yumi Noronha, Marcela Augusta de Souza Pinhel, Gleyson Francisco da Silva Carvalho, Guilherme da Silva Rodrigues, Carlos Roberto Bueno Júnior, Leslie Domenici Kulikowski, Fernando Barbosa Júnior, and et al. 2024. "Genome-Wide Admixture and Association Study of Serum Selenium Deficiency to Identify Genetic Variants Indirectly Linked to Selenium Regulation in Brazilian Adults" Nutrients 16, no. 11: 1627. https://doi.org/10.3390/nu16111627
APA StyleMoriguchi Watanabe, L., Sousa, L., Couto, F. M., Noronha, N. Y., de Souza Pinhel, M. A., da Silva Carvalho, G. F., da Silva Rodrigues, G., Bueno Júnior, C. R., Kulikowski, L. D., Barbosa Júnior, F., & Nonino, C. B. (2024). Genome-Wide Admixture and Association Study of Serum Selenium Deficiency to Identify Genetic Variants Indirectly Linked to Selenium Regulation in Brazilian Adults. Nutrients, 16(11), 1627. https://doi.org/10.3390/nu16111627