
Serum-Derived Bovine Immunoglobulin Promotes Barrier Integrity and Lowers Inflammation for 24 Human Adults Ex Vivo

 , ,
, ,  and
and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Test Compounds
2.2. Experimental Design, Timeline and Analysis
2.3. Host–Microbiome Interaction Assay
2.4. Key Fermentation Parameters
2.5. Taxonomic Microbiota Analysis by Quantitative 16S rRNA Gene Profiling
2.6. Untargeted Metabolite Profiling
2.7. Data Analysis
3. Results
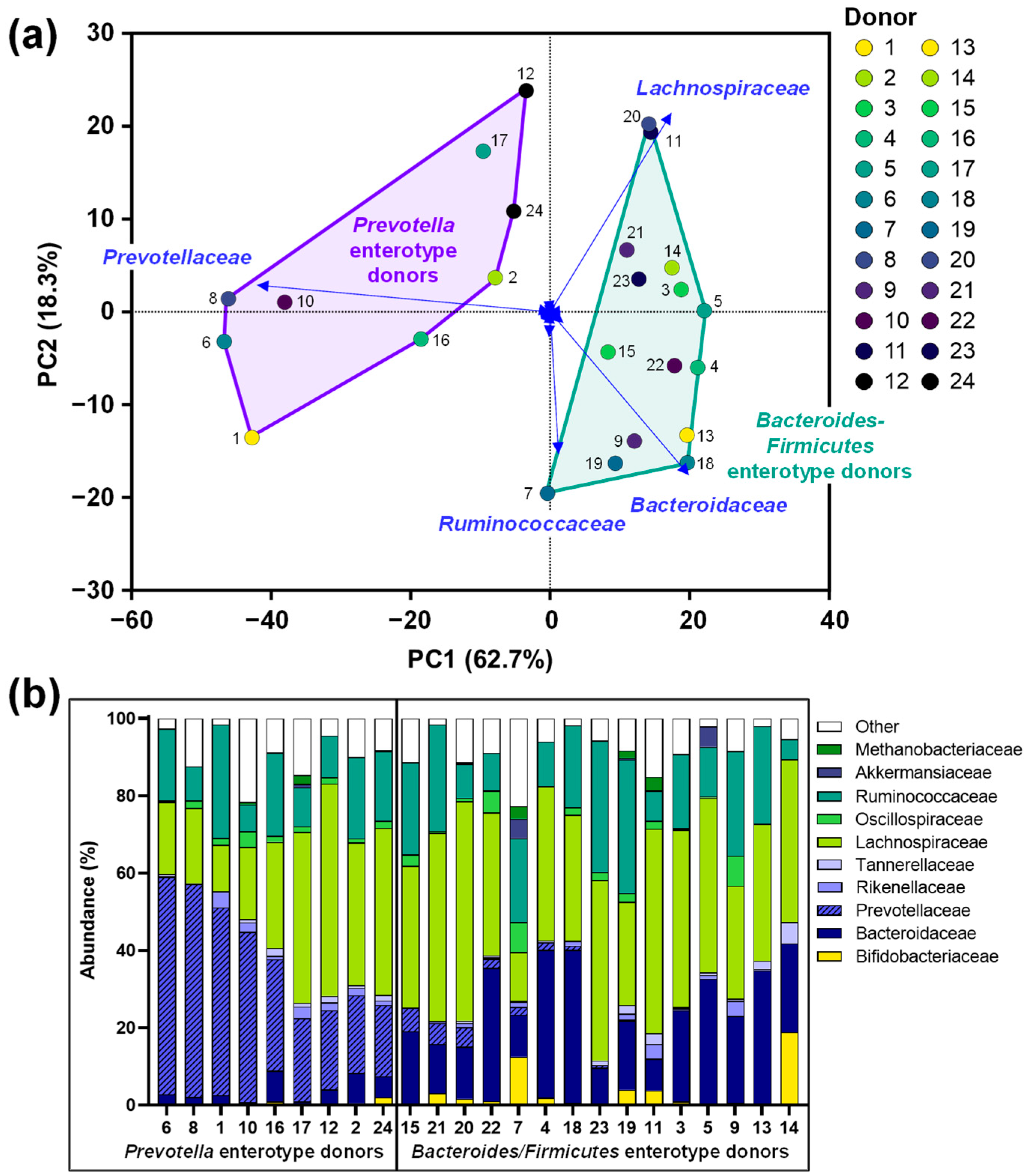
3.1. The Study Cohort Covered a Relevant Spectrum of Interpersonal Differences in Microbiota Composition
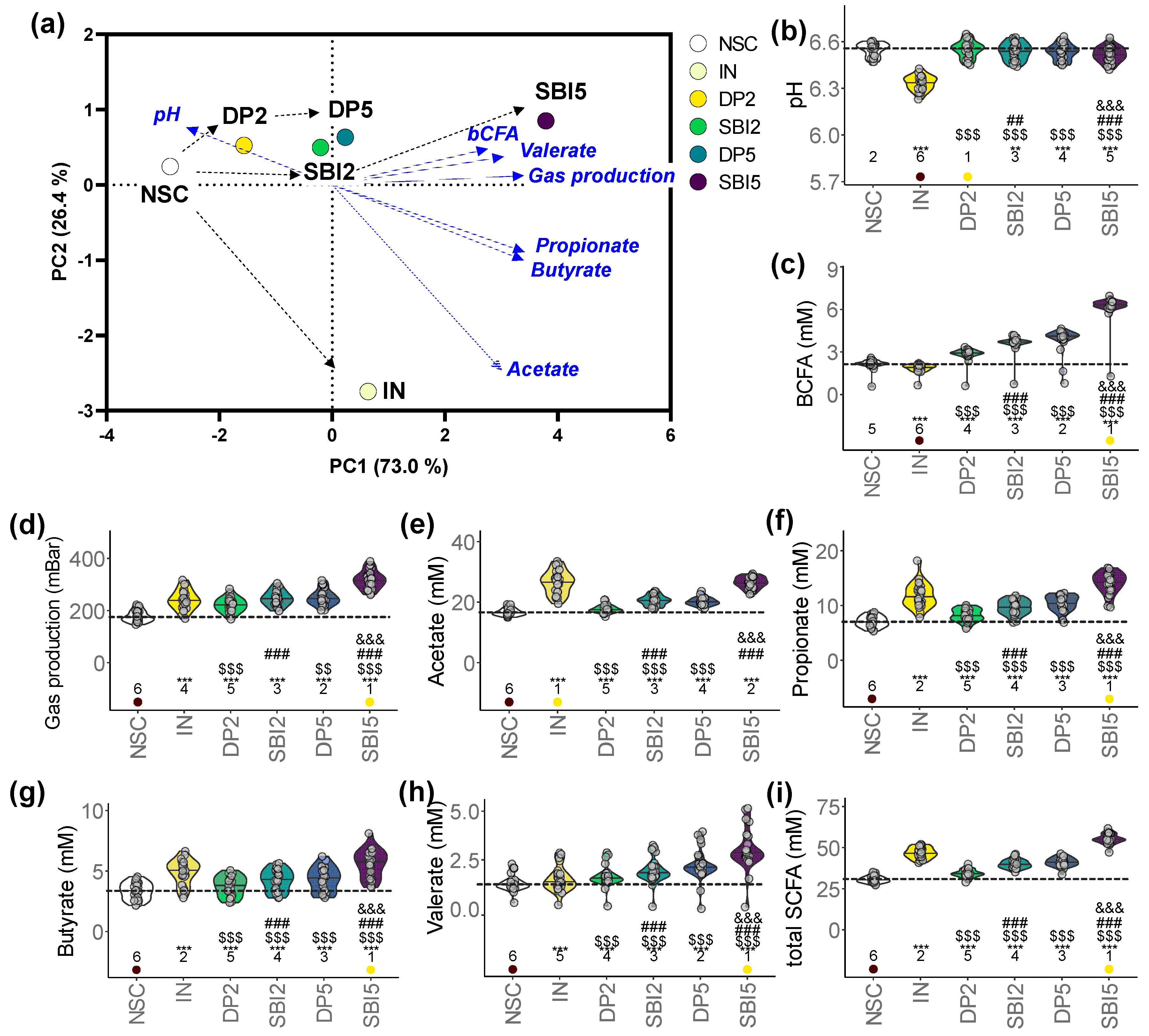
3.2. SBI Promoted Gut Barrier Integrity While Suppressing Pro-Inflammatory Markers
3.3. IN and SBI Stimulated Microbial Metabolite Production
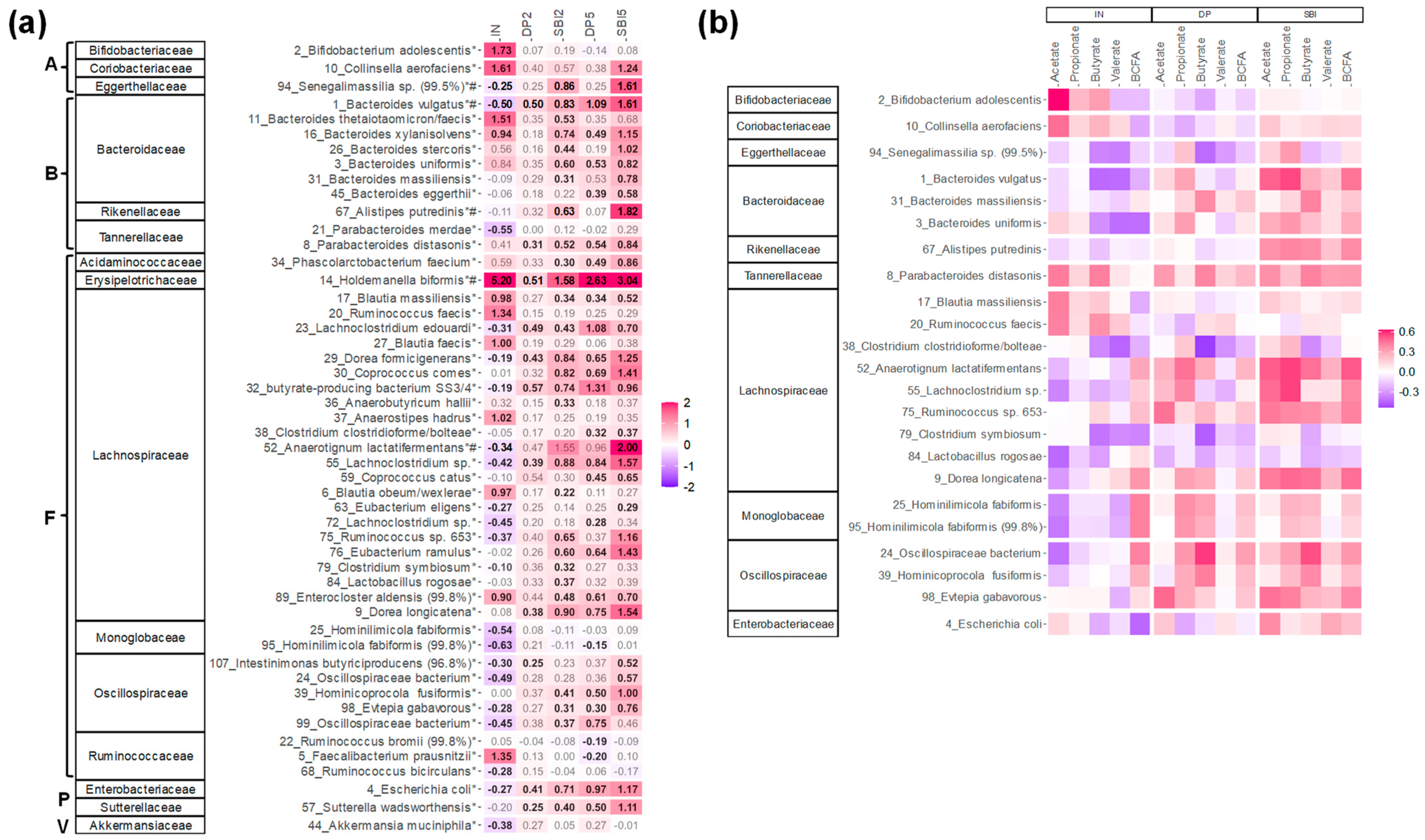
3.4. IN and SBI Each Stimulated a Specific Spectrum of Gut Microbes
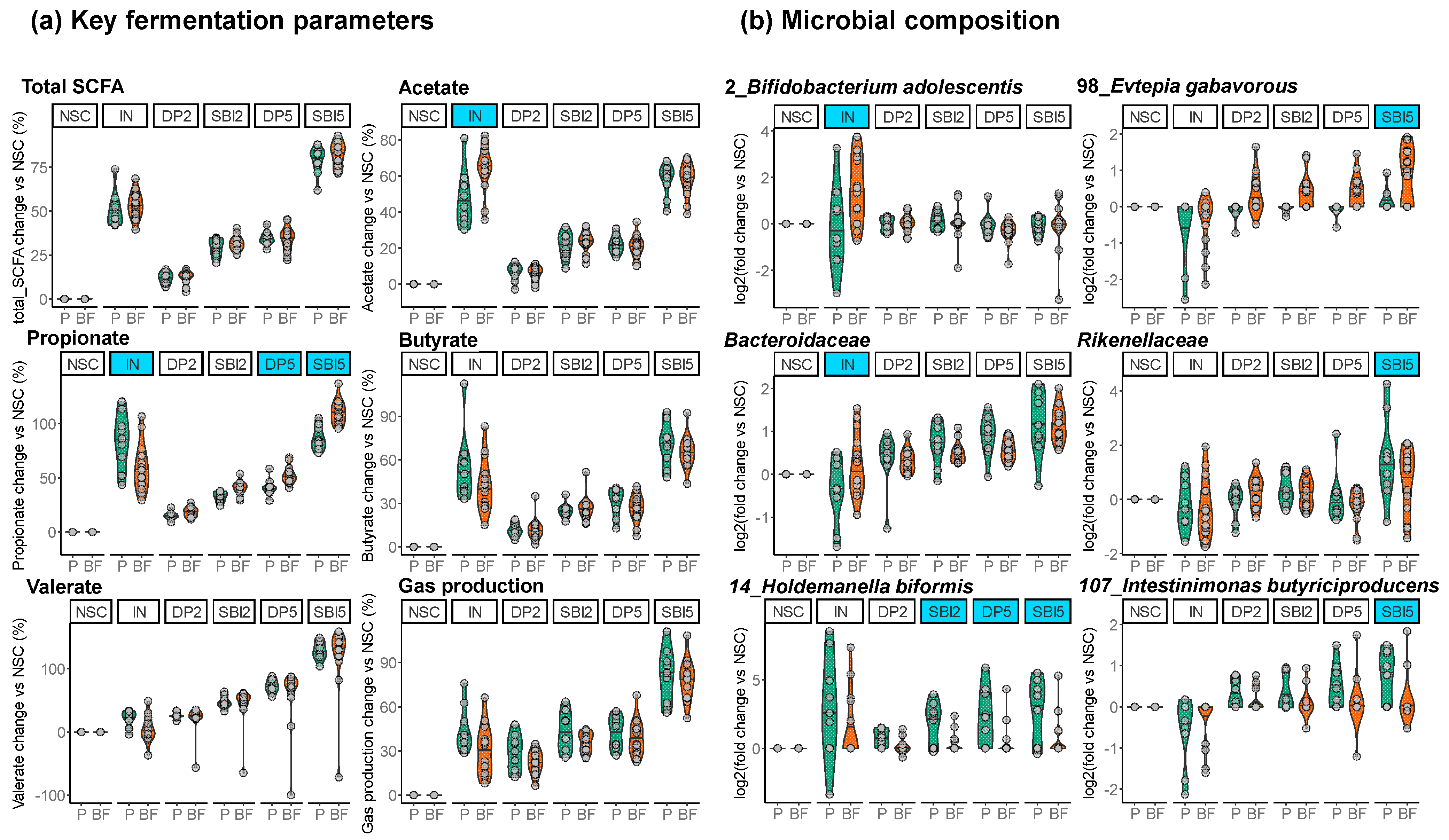
3.5. Stratification of Donor Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ulfman, L.H.; Leusen, J.H.W.; Savelkoul, H.F.J.; Warner, J.O.; van Neerven, R.J.J. Effects of Bovine Immunoglobulins on Immune Function, Allergy, and Infection. Front. Nutr. 2018, 5, 52. [Google Scholar] [CrossRef]
- Detzel, C.J.; Horgan, A.; Henderson, A.L.; Petschow, B.W.; Warner, C.D.; Maas, K.J.; Weaver, E.M. Bovine Immunoglobulin/Protein Isolate Binds pro-Inflammatory Bacterial Compounds and Prevents Immune Activation in an Intestinal Co-Culture Model. PLoS ONE 2015, 10, e0120278. [Google Scholar] [CrossRef]
- Henderson, A.L.; Brand, M.W.; Darling, R.J.; Maas, K.J.; Detzel, C.J.; Hostetter, J.; Wannemuehler, M.J.; Weaver, E.M. Attenuation of Colitis by Serum-Derived Bovine Immunoglobulin/Protein Isolate in a Defined Microbiota Mouse Model. Dig. Dis. Sci. 2015, 60, 3293–3303. [Google Scholar] [CrossRef]
- Utay, N.S.; Somasunderam, A.; Hinkle, J.E.; Petschow, B.W.; Detzel, C.J.; Somsouk, M.; Fichtenbaum, C.J.; Weaver, E.M.; Shaw, A.L.; Asmuth, D.M. Serum Bovine Immunoglobulins Improve Inflammation and Gut Barrier Function in Persons with HIV and Enteropathy on Suppressive ART. Pathog. Immun. 2019, 4, 124–146. [Google Scholar] [CrossRef]
- Horgan, A.; Maas, K.; Henderson, A.; Detzel, C.; Weaver, E. Serum-Derived Bovine Immunoglobulin/Protein Isolate Binds to Pathogen-Associated Molecular Patterns (836.6). FASEB J. 2014, 28, 836.6. [Google Scholar] [CrossRef]
- Petschow, B.W.; Blikslager, A.T.; Weaver, E.M.; Campbell, J.M.; Polo, J.; Shaw, A.L.; Burnett, B.P.; Klein, G.L.; Rhoads, J.M. Bovine Immunoglobulin Protein Isolates for the Nutritional Management of Enteropathy. World J. Gastroenterol. 2014, 20, 11713–11726. [Google Scholar] [CrossRef]
- Petschow, B.W.; Burnett, B.P.; Shaw, A.L.; Weaver, E.M.; Klein, G.L. Dietary Requirement for Serum-Derived Bovine Immunoglobulins in the Clinical Management of Patients with Enteropathy. Dig. Dis. Sci. 2015, 60, 13–23. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Detzel, C.; Rose, A.; Deyaert, S.; Baudot, A.; Warner, C. Serum-Derived Bovine Immunoglobulin Stimulates SCFA Production by Specific Microbes in the Ex Vivo SIFR® Technology. Microorganisms 2023, 11, 659. [Google Scholar] [CrossRef]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut Microbial Metabolites as Multi-Kingdom Intermediates. Nat. Rev. Microbiol. 2021, 19, 77–94. [Google Scholar] [CrossRef]
- Lavelle, A.; Lennon, G.; O’Sullivan, O.; Docherty, N.; Balfe, A.; Maguire, A.; Mulcahy, H.E.; Doherty, G.; O’Donoghue, D.; Hyland, J.; et al. Spatial Variation of the Colonic Microbiota in Patients with Ulcerative Colitis and Control Volunteers. Gut 2015, 64, 1553–1561. [Google Scholar] [CrossRef]
- Ruppin, H.; Bar-Meir, S.; Soergel, K.H.; Wood, C.M.; Schmitt, M.G. Absorption of Short-Chain Fatty Acids by the Colon. Gastroenterology 1980, 78, 1500–1507. [Google Scholar] [CrossRef]
- Delcour, J.A.; Aman, P.; Courtin, C.M.; Hamaker, B.R.; Verbeke, K. Prebiotics, Fermentable Dietary Fiber, and Health Claims. Adv. Nutr. 2016, 7, 1–4. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Deyaert, S.; Thabuis, C.; Perreau, C.; Bajic, D.; Wintergerst, E.; Joossens, M.; Firrman, J.; Walsh, D.; Baudot, A. Bridging Preclinical and Clinical Gut Microbiota Research Using the Ex Vivo SIFR® Technology. Front. Microbiol. 2023, 14, 1131662. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Goggans, M.; Deyaert, S.; Baudot, A.; Van de Vliet, M.; Calatayud, M.; Lelah, M. Lacticaseibacillus Rhamnosus ATCC 53103 and Limosilactobacillus Reuteri ATCC 53608 Synergistically Boost Butyrate Levels upon Tributyrin Administration Ex Vivo. Int. J. Mol. Sci. 2023, 24, 5859. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Deyaert, S.; Albers, R.; Baudot, A.; Mercenier, A. Carrot RG-I Reduces Interindividual Differences between 24 Adults through Consistent Effects on Gut Microbiota Composition and Function Ex Vivo. Nutrients 2023, 15, 2090. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Poppe, J.; Deyaert, S.; Laurie, I.; Otto Gravert, T.K.; Abrahamsson, A.; Baudot, A.; Karnik, K.; Risso, D. Low-No-Calorie Sweeteners Exert Marked Compound-Specific Impact on the Human Gut Microbiota Ex Vivo. Int. J. Food Sci. Nutr. 2023, 74, 630–644. [Google Scholar] [CrossRef]
- Bajic, D.; Wiens, F.; Wintergerst, E.; Deyaert, S.; Baudot, A.; Van den Abbeele, P. HMOs Exert Marked Bifidogenic Effects on Children’s Gut Microbiota Ex Vivo, Due to Age-Related Bifidobacterium Species Composition. Nutrients 2023, 15, 1701. [Google Scholar] [CrossRef]
- Tintoré, M.; Cuñé, J.; Vu, L.D.; Poppe, J.; Van den Abbeele, P.; Baudot, A.; de Lecea, C. A Long-Chain Dextran Produced by Weissella Cibaria Boosts the Diversity of Health-Related Gut Microbes Ex Vivo. Biology 2024, 13, 51. [Google Scholar] [CrossRef]
- Le Bastard, Q.; Chapelet, G.; Javaudin, F.; Lepelletier, D.; Batard, E.; Montassier, E. The Effects of Inulin on Gut Microbial Composition: A Systematic Review of Evidence from Human Studies. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 403–413. [Google Scholar] [CrossRef]
- Toffolon, A.; de Rocco-Ponce, M.; Vettore, M.; Iori, E.; Lante, A.; Tessari, P. Effect of Reversal of Whey-Protein to Casein Ratio of Cow Milk, on Insulin, Incretin, and Amino Acid Responses in Humans. Mol. Nutr. Food Res. 2021, 65, 2100069. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST Static In Vitro Simulation of Gastrointestinal Food Digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Satsu, H.; Ishimoto, Y.; Nakano, T.; Mochizuki, T.; Iwanaga, T.; Shimizu, M. Induction by Activated Macrophage-like THP-1 Cells of Apoptotic and Necrotic Cell Death in Intestinal Epithelial Caco-2 Monolayers via Tumor Necrosis Factor-Alpha. Exp. Cell Res. 2006, 312, 3909–3919. [Google Scholar] [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Doneanu, C.E.; Chen, W.; Mazzeo, J.R. UPLC/MS Monitoring of Water-Soluble Vitamin Bs in Cell Culture Media in Minutes. Water Appl. Note 2011, 2011, 1–7. [Google Scholar]
- Adams, K.J.; Pratt, B.; Bose, N.; Dubois, L.G.; John-Williams, L.; Perrott, K.M.; Ky, K.; Kapahi, P.; Sharma, V.; MacCoss, M.J. Skyline for Small Molecules: A Unifying Software Package for Quantitative Metabolomics. J. Proteome Res. 2020, 19, 1447–1458. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Cao, K.-A.L. mixOmics: An R Package for ‘omics Feature Selection and Multiple Data Integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic Inulin-Type Fructans Induce Specific Changes in the Human Gut Microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef]
- Costea, P.I.; Hildebrand, F.; Arumugam, M.; Bäckhed, F.; Blaser, M.J.; Bushman, F.D.; de Vos, W.M.; Ehrlich, S.D.; Fraser, C.M.; Hattori, M.; et al. Enterotypes in the Landscape of Gut Microbial Community Composition. Nat. Microbiol. 2018, 3, 8–16. [Google Scholar] [CrossRef]
- Shaw, A.L.; Mathews, D.W.; Hinkle, J.E.; Petschow, B.W.; Weaver, E.M.; Detzel, C.J.; Klein, G.L.; Bradshaw, T.P. Absorption and Safety of Serum-Derived Bovine Immunoglobulin/Protein Isolate in Healthy Adults. Clin. Exp. Gastroenterol. 2016, 9, 365–375. [Google Scholar] [CrossRef]
- Pham, V.T.; Seifert, N.; Richard, N.; Raederstorff, D.; Steinert, R.; Prudence, K.; Mohajeri, M.H. The Effects of Fermentation Products of Prebiotic Fibres on Gut Barrier and Immune Functions In Vitro. PeerJ 2018, 6, e5288. [Google Scholar] [CrossRef]
- Kono, G.; Yoshida, K.; Kokubo, E.; Ikeda, M.; Matsubara, T.; Koyama, T.; Iwamoto, H.; Miyaji, K. Fermentation Supernatant of Elderly Feces with Inulin and Partially Hydrolyzed Guar Gum Maintains the Barrier of Inflammation-Induced Caco-2/HT29-MTX-E12 Co-Cultured Cells. J. Agric. Food Chem. 2023, 71, 1510–1517. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms Regulating Intestinal Barrier Integrity and Its Pathological Implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Friedrich, M.; Pohin, M.; Powrie, F. Cytokine Networks in the Pathophysiology of Inflammatory Bowel Disease. Immunity 2019, 50, 992–1006. [Google Scholar] [CrossRef]
- Vazirinejad, R.; Ahmadi, Z.; Arababadi, M.K.; Hassanshahi, G.; Kennedy, D. The Biological Functions, Structure and Sources of CXCL10 and Its Outstanding Part in the Pathophysiology of Multiple Sclerosis. Neuroimmunomodulation 2014, 21, 322–330. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef]
- Romaní-Pérez, M.; López-Almela, I.; Bullich-Vilarrubias, C.; Rueda-Ruzafa, L.; Gómez Del Pulgar, E.M.; Benítez-Páez, A.; Liebisch, G.; Lamas, J.A.; Sanz, Y. Holdemanella biformis Improves Glucose Tolerance and Regulates GLP-1 Signaling in Obese Mice. FASEB J. 2021, 35, e21734. [Google Scholar] [CrossRef]
- Cuffaro, B.; Assohoun, A.L.W.; Boutillier, D.; Súkeníková, L.; Desramaut, J.; Boudebbouze, S.; Salomé-Desnoulez, S.; Hrdý, J.; Waligora-Dupriet, A.-J.; Maguin, E.; et al. In Vitro Characterization of Gut Microbiota-Derived Commensal Strains: Selection of Parabacteroides Distasonis Strains Alleviating TNBS-Induced Colitis in Mice. Cells 2020, 9, 2104. [Google Scholar] [CrossRef]
- Sun, H.; Guo, Y.; Wang, H.; Yin, A.; Hu, J.; Yuan, T.; Zhou, S.; Xu, W.; Wei, P.; Yin, S.; et al. Gut Commensal Parabacteroides distasonis Alleviates Inflammatory Arthritis. Gut 2023, 72, 1664–1677. [Google Scholar] [CrossRef] [PubMed]
- Dot, T.; Osawa, R.; Stackebrandt, E. Phascolarctobacterium faecium Gen. Nov, Spec. Nov., a Novel Taxon of the Sporomusa Group of Bacteria. Syst. Appl. Microbiol. 1993, 16, 380–384. [Google Scholar] [CrossRef]
- Menni, C.; Hernandez, M.M.; Vital, M.; Mohney, R.P.; Spector, T.D.; Valdes, A.M. Circulating Levels of the Anti-Oxidant Indoleproprionic Acid Are Associated with Higher Gut Microbiome Diversity. Gut Microbes 2019, 10, 688–695. [Google Scholar] [CrossRef]
- Zhao, Z.-H.; Xin, F.-Z.; Xue, Y.; Hu, Z.; Han, Y.; Ma, F.; Zhou, D.; Liu, X.-L.; Cui, A.; Liu, Z.; et al. Indole-3-Propionic Acid Inhibits Gut Dysbiosis and Endotoxin Leakage to Attenuate Steatohepatitis in Rats. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Yang, L.; You, K.; Chen, T.; Su, Z.; Cui, Z.; Wang, M.; Zhang, W.; Liu, B.; Zhou, K.; et al. Indole-3-Acetic Acid Alters Intestinal Microbiota and Alleviates Ankylosing Spondylitis in Mice. Front. Immunol. 2022, 13, 762580. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial Tryptophan Metabolites Regulate Gut Barrier Function via the Aryl Hydrocarbon Receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Turroni, F.; Ventura, M.; van Sinderen, D. Insights into Endogenous Bifidobacterium Species in the Human Gut Microbiota during Adulthood. Trends Microbiol. 2022, 30, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Duranti, S.; Longhi, G.; Ventura, M.; van Sinderen, D.; Turroni, F. Exploring the Ecology of Bifidobacteria and Their Genetic Adaptation to the Mammalian Gut. Microorganisms 2020, 9, 8. [Google Scholar] [CrossRef]
- Cantu-Jungles, T.M.; Hamaker, B.R. New View on Dietary Fiber Selection for Predictable Shifts in Gut Microbiota. mBio 2020, 11, e02179-19. [Google Scholar] [CrossRef]
- Cantu-Jungles, T.M.; Bulut, N.; Chambry, E.; Ruthes, A.; Iacomini, M.; Keshavarzian, A.; Johnson, T.A.; Hamaker, B.R. Dietary Fiber Hierarchical Specificity: The Missing Link for Predictable and Strong Shifts in Gut Bacterial Communities. mBio 2021, 12, e01028-21. [Google Scholar] [CrossRef] [PubMed]
- Goetze, O.; Fruehauf, H.; Pohl, D.; Giarrè, M.; Rochat, F.; Ornstein, K.; Menne, D.; Fried, M.; Thumshirn, M. Effect of a Prebiotic Mixture on Intestinal Comfort and General Wellbeing in Health. Br. J. Nutr. 2008, 100, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Chow, E.; Liu, S.; Du, Y.; Sandy, K.-C. The Caco-2 Cell Monolayer: Usefulness and Limitations. Expert Opin. Drug Metab. Toxicol. 2008, 4, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, I.J.; Raub, T.J.; Borchardt, R.T. Characterization of the Human Colon Carcinoma Cell Line (Caco-2) as a Model System for Intestinal Epithelial Permeability. Gastroenterology 1989, 96, 736–749. [Google Scholar] [CrossRef] [PubMed]
- Yee, S. In Vitro Permeability across Caco-2 Cells (Colonic) Can Predict In Vivo (Small Intestinal) Absorption in Man—Fact or Myth. Pharm. Res. 1997, 14, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and Characterization of a Human Acute Monocytic Leukemia Cell Line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.; Collins, S.; Trujillo, J.; McCredie, K.; Ahearn, M.; Tsai, S.; Metzgar, R.; Aulakh, G.; Ting, R.; Ruscetti, F.; et al. Characterization of the Continuous, Differentiating Myeloid Cell Line (HL-60) from a Patient with Acute Promyelocytic Leukemia. Blood 1979, 54, 713–733. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 Cell Line: An In Vitro Cell Model for Immune Modulation Approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.; Evans, M.; Weaver, E.; Shaw, A.L.; Klein, G.L. Evaluation of Serum-Derived Bovine Immunoglobulin Protein Isolate in Subjects with Diarrhea-Predominant Irritable Bowel Syndrome. Clin. Med. Insights Gastroenterol. 2013, 6, 49–60. [Google Scholar] [CrossRef]
- Shafran, I.; Burgunder, P.; Wei, D.; Young, H.E.; Klein, G.; Burnett, B.P. Management of Inflammatory Bowel Disease with Oral Serum-Derived Bovine Immunoglobulin. Ther. Adv. Gastroenterol. 2015, 8, 331–339. [Google Scholar] [CrossRef]
- Duffy, M.A.; Chen, Q.; Zhang, J.; Halbur, P.G.; Opriessnig, T. Impact of Dietary Spray-Dried Bovine Plasma Addition on Pigs Infected with Porcine Epidemic Diarrhea Virus. Transl. Anim. Sci. 2018, 2, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Maijó, M.; Miró, L.; Polo, J.; Campbell, J.; Russell, L.; Crenshaw, J.; Weaver, E.; Moretó, M.; Pérez-Bosque, A. Dietary Plasma Proteins Modulate the Adaptive Immune Response in Mice with Acute Lung Inflammation. J. Nutr. 2012, 142, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Arthington, J.D.; Jaynes, C.A.; Tyler, H.D.; Kapil, S.; Quigley, J.D. The Use of Bovine Serum Protein as an Oral Support Therapy Following Coronavirus Challenge in Calves. J. Dairy Sci. 2002, 85, 1249–1254. [Google Scholar] [CrossRef] [PubMed]
- Rosell-Cardona, C.; Amat, C.; Griñán-Ferré, C.; Polo, J.; Pallàs, M.; Pérez-Bosque, A.; Moretó, M.; Miró, L. The Neuroprotective Effects of Spray-Dried Porcine Plasma Supplementation Involve the Microbiota−Gut−Brain Axis. Nutrients 2022, 14, 2211. [Google Scholar] [CrossRef]
- Rosell-Cardona, C.; Griñan-Ferré, C.; Pérez-Bosque, A.; Polo, J.; Pallàs, M.; Amat, C.; Moretó, M.; Miró, L. Dietary Spray-Dried Porcine Plasma Reduces Neuropathological Alzheimer’s Disease Hallmarks in SAMP8 Mice. Nutrients 2021, 13, 2369. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van den Abbeele, P.; Kunkler, C.N.; Poppe, J.; Rose, A.; van Hengel, I.A.J.; Baudot, A.; Warner, C.D. Serum-Derived Bovine Immunoglobulin Promotes Barrier Integrity and Lowers Inflammation for 24 Human Adults Ex Vivo. Nutrients 2024, 16, 1585. https://doi.org/10.3390/nu16111585
Van den Abbeele P, Kunkler CN, Poppe J, Rose A, van Hengel IAJ, Baudot A, Warner CD. Serum-Derived Bovine Immunoglobulin Promotes Barrier Integrity and Lowers Inflammation for 24 Human Adults Ex Vivo. Nutrients. 2024; 16(11):1585. https://doi.org/10.3390/nu16111585
Chicago/Turabian StyleVan den Abbeele, Pieter, Charlotte N. Kunkler, Jonas Poppe, Alexis Rose, Ingmar A. J. van Hengel, Aurélien Baudot, and Christopher D. Warner. 2024. "Serum-Derived Bovine Immunoglobulin Promotes Barrier Integrity and Lowers Inflammation for 24 Human Adults Ex Vivo" Nutrients 16, no. 11: 1585. https://doi.org/10.3390/nu16111585
APA StyleVan den Abbeele, P., Kunkler, C. N., Poppe, J., Rose, A., van Hengel, I. A. J., Baudot, A., & Warner, C. D. (2024). Serum-Derived Bovine Immunoglobulin Promotes Barrier Integrity and Lowers Inflammation for 24 Human Adults Ex Vivo. Nutrients, 16(11), 1585. https://doi.org/10.3390/nu16111585