

Pharmacological Activity of Flavonoid Quercetin and Its Therapeutic Potential in Testicular Injury

Abstract
1. Introduction
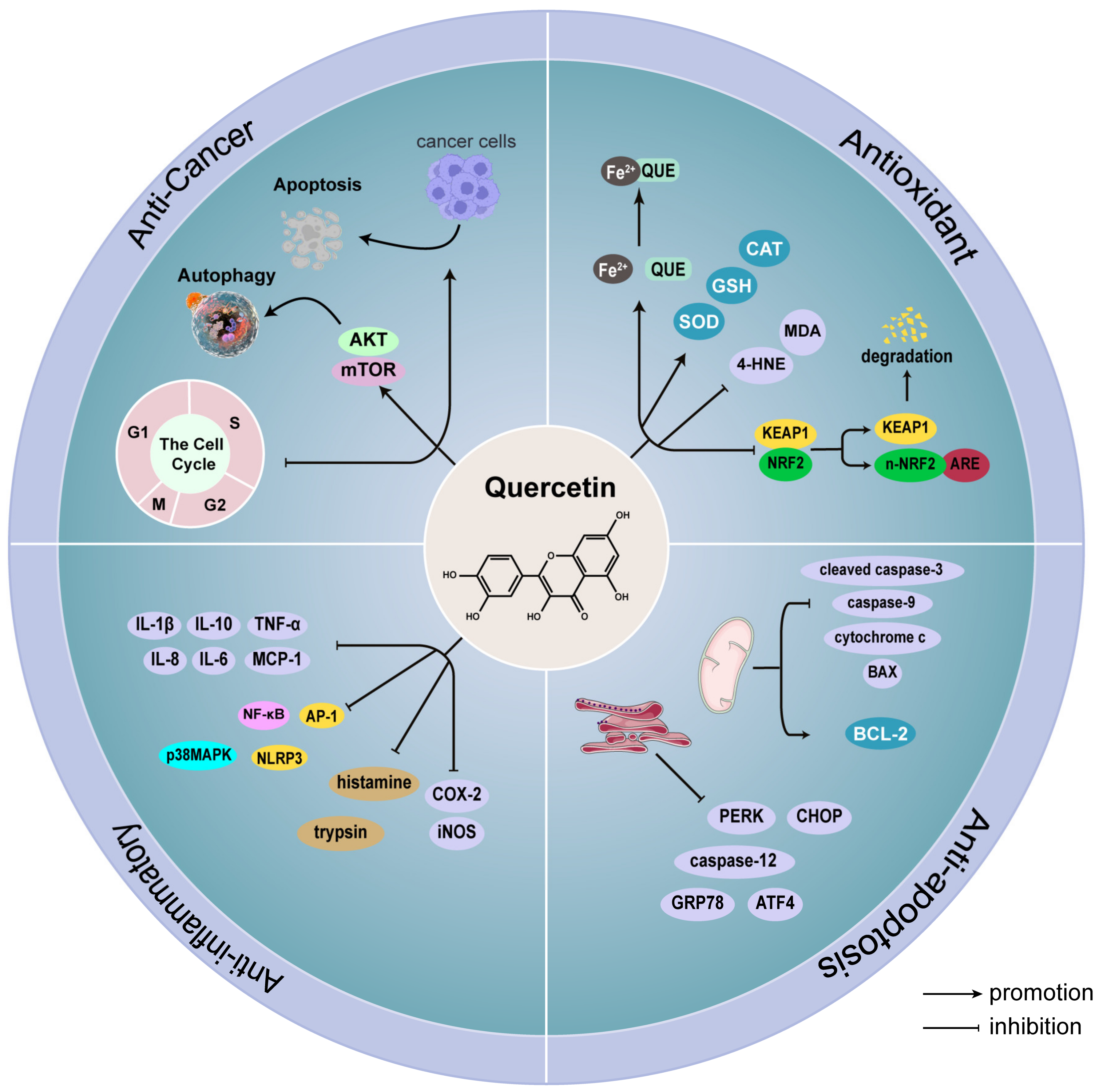
2. Quercetin
2.1. Antioxidant Effect of Quercetin
2.2. Anti-Apoptotic Effect of Quercetin
2.3. Anti-Inflammatory Effect of Quercetin
2.4. Anti-Cancer
3. Protective Effect of Quercetin on Testicular Injury
3.1. Diabetes
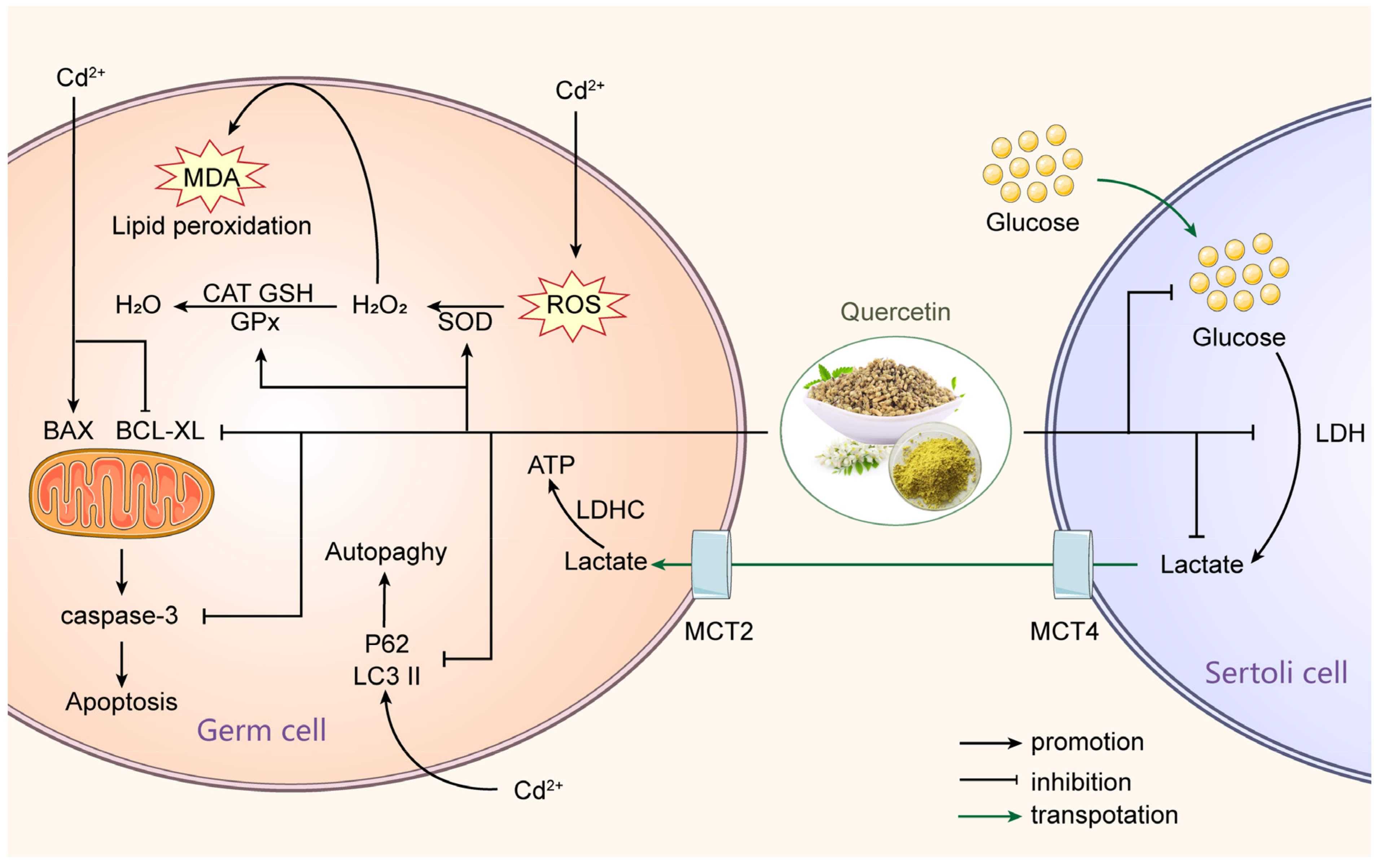
3.2. Environmental Pollutants
3.2.1. Heavy Metals Exposure
3.2.2. Other Environmental Pollutants Exposure
3.3. Drugs
3.4. Testicular Torsion/Detorsion
4. Clinical Studies, Application Prospects, and Limitations of Quercetin
4.1. Clinical Trials on Quercetin
4.2. The Application Prospects and Limitations of Quercetin
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stevenson, D.E.; Hurst, R.D. Polyphenolic phytochemicals—Just antioxidants or much more? Cell. Mol. Life Sci. CMLS 2007, 64, 2900–2916. [Google Scholar] [CrossRef] [PubMed]
- Somerville, V.; Bringans, C.; Braakhuis, A. Polyphenols and Performance: A Systematic Review and Meta-Analysis. Sports Med. 2017, 47, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.S.; Reboredo-Rodríguez, P.; Süntar, I.; Sureda, A.; Belwal, T.; Loizzo, M.R.; Tundis, R.; Sobarzo-Sanchez, E.; Rastrelli, L.; Forbes-Hernandez, T.Y.; et al. Evaluation of the status quo of polyphenols analysis: Part I-phytochemistry, bioactivity, interactions, and industrial uses. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3191–3218. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cao, H.; Huang, Q.; Xiao, J.; Teng, H. Absorption, metabolism and bioavailability of flavonoids: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 7730–7742. [Google Scholar] [CrossRef]
- Rufino, A.T.; Costa, V.M.; Carvalho, F.; Fernandes, E. Flavonoids as antiobesity agents: A review. Med. Res. Rev. 2021, 41, 556–585. [Google Scholar] [CrossRef]
- Han, S.; Luo, Y.; Liu, B.; Guo, T.; Qin, D.; Luo, F. Dietary flavonoids prevent diabetes through epigenetic regulation: Advance and challenge. Crit. Rev. Food Sci. Nutr. 2022, 1–17. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Y.; Song, J.; Wang, R.; Gao, L.; Zhang, L.; Fang, L.; Lu, Y.; Du, G. Total flavonoids from Anchusa italica Retz. Improve cardiac function and attenuate cardiac remodeling post myocardial infarction in mice. J. Ethnopharmacol. 2020, 257, 112887. [Google Scholar] [CrossRef]
- Khan, H.; Belwal, T.; Efferth, T.; Farooqi, A.A.; Sanches-Silva, A.; Vacca, R.A.; Nabavi, S.F.; Khan, F.; Prasad Devkota, H.; Barreca, D.; et al. Targeting epigenetics in cancer: Therapeutic potential of flavonoids. Crit. Rev. Food Sci. Nutr. 2021, 61, 1616–1639. [Google Scholar] [CrossRef]
- De Andrade Teles, R.B.; Diniz, T.C.; Costa Pinto, T.C.; de Oliveira Júnior, R.G.; Gama, E.; Silva, M.; de Lavor, É.M.; Fernandes, A.W.C.; de Oliveira, A.P.; de Almeida Ribeiro, F.P.R.; et al. Flavonoids as Therapeutic Agents in Alzheimer’s and Parkinson’s Diseases: A Systematic Review of Preclinical Evidences. Oxidative Med. Cell. Longev. 2018, 2018, 7043213. [Google Scholar] [CrossRef]
- Rengasamy, K.R.R.; Khan, H.; Gowrishankar, S.; Lagoa, R.J.L.; Mahomoodally, F.M.; Khan, Z.; Suroowan, S.; Tewari, D.; Zengin, G.; Hassan, S.T.S.; et al. The role of flavonoids in autoimmune diseases: Therapeutic updates. Pharmacol. Ther. 2019, 194, 107–131. [Google Scholar] [CrossRef]
- Oteiza, P.I.; Fraga, C.G.; Mills, D.A.; Taft, D.H. Flavonoids and the gastrointestinal tract: Local and systemic effects. Mol. Asp. Med. 2018, 61, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Peng, P.; Zou, J.; Zhong, B.; Zhang, G.; Zou, X.; Xie, T. Protective Effects and Mechanisms of Flavonoids in Renal Ischemia-Reperfusion Injury. Pharmacology 2023, 108, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, J.; Hou, J.; Hui, M.; Qi, H.; Lei, T.; Zhang, X.; Zhao, L.; Du, H. Induction of autophagy via the PI3K/Akt/mTOR signaling pathway by Pueraria flavonoids improves non-alcoholic fatty liver disease in obese mice. Biomed. Pharmacother. 2023, 157, 114005. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.-J.; Yang, J.-M.; Hai, D.-M.; Liu, N.; Ma, L.; Lan, X.-B.; Niu, J.-G.; Zheng, P.; Yu, J.-Q. Interplay between male reproductive system dysfunction and the therapeutic effect of flavonoids. Fitoterapia 2020, 147, 104756. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.-L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef]
- Palnitkar, G.; Phillips, C.L.; Hoyos, C.M.; Marren, A.J.; Bowman, M.C.; Yee, B.J. Linking sleep disturbance to idiopathic male infertility. Sleep Med. Rev. 2018, 42, 149–159. [Google Scholar] [CrossRef]
- Minhas, S.; Bettocchi, C.; Boeri, L.; Capogrosso, P.; Carvalho, J.; Cilesiz, N.C.; Cocci, A.; Corona, G.; Dimitropoulos, K.; Gül, M.; et al. European Association of Urology Guidelines on Male Sexual and Reproductive Health: 2021 Update on Male Infertility. Eur. Urol. 2021, 80, 603–620. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Hosseini, A.; Razavi, B.M.; Banach, M.; Hosseinzadeh, H. Quercetin and metabolic syndrome: A review. Phytother. Res. PTR 2021, 35, 5352–5364. [Google Scholar] [CrossRef]
- Shen, P.; Lin, W.; Deng, X.; Ba, X.; Han, L.; Chen, Z.; Qin, K.; Huang, Y.; Tu, S. Potential Implications of Quercetin in Autoimmune Diseases. Front. Immunol. 2021, 12, 689044. [Google Scholar] [CrossRef]
- Lai, W.-F.; Wong, W.-T. Design and optimization of quercetin-based functional foods. Crit. Rev. Food Sci. Nutr. 2022, 62, 7319–7335. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ashida, H.; Terao, J. Multitargeted cancer prevention by quercetin. Cancer Lett. 2008, 269, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Zhao, X.; Amevor, F.K.; Du, X.; Wang, Y.; Li, D.; Shu, G.; Tian, Y.; Zhao, X. Therapeutic application of quercetin in aging-related diseases: SIRT1 as a potential mechanism. Front. Immunol. 2022, 13, 943321. [Google Scholar] [CrossRef] [PubMed]
- Shabir, I.; Kumar Pandey, V.; Shams, R.; Dar, A.H.; Dash, K.K.; Khan, S.A.; Bashir, I.; Jeevarathinam, G.; Rusu, A.V.; Esatbeyoglu, T.; et al. Promising bioactive properties of quercetin for potential food applications and health benefits: A review. Front. Nutr. 2022, 9, 999752. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.A.; Tatarskiy, V.; Kalinina, E. Synthetic Pathways and the Therapeutic Potential of Quercetin and Curcumin. Int. J. Mol. Sci. 2022, 23, 14413. [Google Scholar] [CrossRef] [PubMed]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2020, 60, 3290–3303. [Google Scholar] [CrossRef]
- Lv, Q.-Z.; Long, J.-T.; Gong, Z.-F.; Nong, K.-Y.; Liang, X.-M.; Qin, T.; Huang, W.; Yang, L. Current State of Knowledge on the Antioxidant Effects and Mechanisms of Action of Polyphenolic Compounds. Nat. Prod. Commun. 2021, 16, 1–13. [Google Scholar] [CrossRef]
- Deepika; Maurya, P. K. Health Benefits of Quercetin in Age-Related Diseases. Molecules 2022, 27, 2498. [Google Scholar] [CrossRef]
- Machha, A.; Achike, F.I.; Mustafa, A.M.; Mustafa, M.R. Quercetin, a flavonoid antioxidant, modulates endothelium-derived nitric oxide bioavailability in diabetic rat aortas. Nitric Oxide Biol. Chem. 2007, 16, 442–447. [Google Scholar] [CrossRef]
- Vásquez-Espinal, A.; Yañez, O.; Osorio, E.; Areche, C.; García-Beltrán, O.; Ruiz, L.M.; Cassels, B.K.; Tiznado, W. Theoretical Study of the Antioxidant Activity of Quercetin Oxidation Products. Front Chem. 2019, 7, 818. [Google Scholar] [CrossRef]
- Rashidi, Z.; Aleyasin, A.; Eslami, M.; Nekoonam, S.; Zendedel, A.; Bahramrezaie, M.; Amidi, F. Quercetin protects human granulosa cells against oxidative stress via thioredoxin system. Reprod. Biol. 2019, 19, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Zhang, Q.; Shen, L.; Sharma, G.; Jiang, H.; Wang, Z.; Shen, J. Quercetin Attenuates Quinocetone-Induced Cell Apoptosis In Vitro by Activating the P38/Nrf2/HO-1 Pathway and Inhibiting the ROS/Mitochondrial Apoptotic Pathway. Antioxidants 2022, 11, 1498. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-Y.; Lee, Y.-J.; Hsu, M.-Y.; Wang, M.; Tsou, S.-C.; Chen, C.-C.; Lin, J.-A.; Hsiao, Y.-P.; Lin, H.-W. Protective Effect of Quercetin on Sodium Iodate-Induced Retinal Apoptosis through the Reactive Oxygen Species-Mediated Mitochondrion-Dependent Pathway. Int. J. Mol. Sci. 2021, 22, 4056. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Miao, Z.; Wang, S.; Shi, X.; Xu, S. Quercetin antagonizes imidacloprid-induced mitochondrial apoptosis through PTEN/PI3K/AKT in grass carp hepatocytes. Environ. Pollut. 2021, 290, 118036. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ding, L.; Ye, Y.; Wang, K.; Yu, W.; Yan, B.; Liu, Z.; Wang, J. Protective effect of quercetin on cadmium-induced renal apoptosis through cyt-c/caspase-9/caspase-3 signaling pathway. Front. Pharmacol. 2022, 13, 990993. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tian, Z.; Li, W.; Wang, X.; Man, Z.; Sun, S. Inhibitory effect of quercetin on titanium particle-induced endoplasmic reticulum stress (ERS)-related apoptosis and in vivoosteolysis. Biosci. Rep. 2017, 37, BSR20170961. [Google Scholar] [CrossRef]
- Wang, J.; Ding, L.; Wang, K.; Huang, R.; Yu, W.; Yan, B.; Wang, H.; Zhang, C.; Yang, Z.; Liu, Z. Role of endoplasmic reticulum stress in cadmium-induced hepatocyte apoptosis and the protective effect of quercetin. Ecotoxicol. Environ. Saf. 2022, 241, 113772. [Google Scholar] [CrossRef]
- Liu, X.-X.; Tang, L.; Ge, R.; Li, J.-K.; Kang, Y.; Zhu, M.-X.; Li, Q.-S.; Hao, X.-L. iTRAQ-based quantitative proteomic analysis of the anti-apoptotic effect of hyperin, which is mediated by Mcl-1 and Bid, in H2O2-injured EA. hy926 cells. Int. J. Mol. Med. 2016, 37, 1083–1090. [Google Scholar] [CrossRef]
- Fawzy, M.A.; Nasr, G.; Ali, F.E.M.; Fathy, M. Quercetin potentiates the hepatoprotective effect of sildenafil and/or pentoxifylline against intrahepatic cholestasis: Role of Nrf2/ARE, TLR4/NF-κB, and NLRP3/IL-1β signaling pathways. Life Sci. 2023, 314, 121343. [Google Scholar] [CrossRef]
- Liu, J.; Li, X.; Yue, Y.; Li, J.; He, T.; He, Y. The inhibitory effect of quercetin on IL-6 production by LPS-stimulated neutrophils. Cell Mol. Immunol. 2005, 2, 455–460. [Google Scholar]
- Chansiw, N.; Champakam, S.; Chusri, P.; Pangjit, K.; Srichairatanakool, S. Quercetin-Rich Ethanolic Extract of Polygonum odoratum var Pakphai Leaves Decreased Gene Expression and Secretion of Pro-Inflammatory Mediators in Lipopolysaccharide-Induced Murine RAW264.7 Macrophages. Molecules 2022, 27, 3657. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Miao, M.; Zhang, Y.; Liu, R.; Li, X.; Cui, Y.; Qu, L. Quercetin ameliorates liver injury induced with Tripterygium glycosides by reducing oxidative stress and inflammation. Can. J. Physiol. Pharmacol. 2015, 93, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhang, X.; Zhang, L.; Bian, H.-X.; Xu, N.; Bao, B.; Liu, J. Quercetin reduces obesity-associated ATM infiltration and inflammation in mice: A mechanism including AMPKα1/SIRT1. J. Lipid Res. 2014, 55, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhang, X.; Zhu, G.; Liu, H.; Chen, J.; Wang, Y.; He, X. Quercetin inhibits TNF-α induced HUVECs apoptosis and inflammation via downregulating NF-kB and AP-1 signaling pathway in vitro. Medicine 2020, 99, e22241. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Pan, Y.; Zhang, Q.-Y.; Wang, F.-M.; Kong, L.-D. Quercetin and allopurinol ameliorate kidney injury in STZ-treated rats with regulation of renal NLRP3 inflammasome activation and lipid accumulation. PLoS ONE 2012, 7, e38285. [Google Scholar] [CrossRef]
- Min, Y.D.; Choi, C.H.; Bark, H.; Son, H.Y.; Park, H.H.; Lee, S.; Park, J.W.; Park, E.K.; Shin, H.I.; Kim, S.H. Quercetin inhibits expression of inflammatory cytokines through attenuation of NF-κB and p38 MAPK in HMC-1 human mast cell line. Inflamm. Res. 2007, 56, 210–215. [Google Scholar] [CrossRef]
- Kempuraj, D.; Madhappan, B.; Christodoulou, S.; Boucher, W.; Cao, J.; Papadopoulou, N.; Cetrulo, C.L.; Theoharides, T.C. Flavonols inhibit proinflammatory mediator release, intracellular calcium ion levels and protein kinase C theta phosphorylation in human mast cells. Br. J. Pharmacol. 2005, 145, 934–944. [Google Scholar] [CrossRef]
- Primikyri, A.; Chatziathanasiadou, M.V.; Karali, E.; Kostaras, E.; Mantzaris, M.D.; Hatzimichael, E.; Shin, J.-S.; Chi, S.-W.; Briasoulis, E.; Kolettas, E.; et al. Direct binding of Bcl-2 family proteins by quercetin triggers its pro-apoptotic activity. ACS Chem. Biol. 2014, 9, 2737–2741. [Google Scholar] [CrossRef]
- Luo, C.-L.; Liu, Y.-Q.; Wang, P.; Song, C.-H.; Wang, K.-J.; Dai, L.-P.; Zhang, J.-Y.; Ye, H. The effect of quercetin nanoparticle on cervical cancer progression by inducing apoptosis, autophagy and anti-proliferation via JAK2 suppression. Biomed. Pharmacother. 2016, 82, 595–605. [Google Scholar] [CrossRef]
- Zhu, Y.; Jiang, Y.; Shi, L.; Du, L.; Xu, X.; Wang, E.; Sun, Y.; Guo, X.; Zou, B.; Wang, H.; et al. 7-O-Geranylquercetin induces apoptosis in gastric cancer cells via ROS-MAPK mediated mitochondrial signaling pathway activation. Biomed. Pharmacother. 2017, 87, 527–538. [Google Scholar] [CrossRef]
- Teekaraman, D.; Elayapillai, S.P.; Viswanathan, M.P.; Jagadeesan, A. Quercetin inhibits human metastatic ovarian cancer cell growth and modulates components of the intrinsic apoptotic pathway in PA-1 cell line. Chem. Biol. Interact. 2019, 300, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, X.; Zhuang, J.; Zhang, L.; Lin, Y. Induction of death receptor 5 and suppression of survivin contribute to sensitization of TRAIL-induced cytotoxicity by quercetin in non-small cell lung cancer cells. Carcinogenesis 2007, 28, 2114–2121. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.S.; Yao, C.N.; Liu, H.C.; Yu, F.S.; Lin, J.J.; Lu, K.W.; Liao, C.L.; Chueh, F.S.; Chung, J.G. Quercetin induced apoptosis of human oral cancer SAS cells through mitochondria and endoplasmic reticulum mediated signaling pathways. Oncol. Lett. 2018, 15, 9663–9672. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-H.; An, J.Y.; Kwon, Y.T.; Rhee, J.G.; Lee, Y.J. Effects of low dose quercetin: Cancer cell-specific inhibition of cell cycle progression. J. Cell. Biochem. 2009, 106, 73–82. [Google Scholar] [CrossRef]
- Lee, T.-J.; Kim, O.H.; Kim, Y.H.; Lim, J.H.; Kim, S.; Park, J.-W.; Kwon, T.K. Quercetin arrests G2/M phase and induces caspase-dependent cell death in U937 cells. Cancer Lett. 2006, 240, 234–242. [Google Scholar] [CrossRef]
- Sudan, S.; Rupasinghe, H.P.V. Quercetin-3-O-glucoside induces human DNA topoisomerase II inhibition, cell cycle arrest and apoptosis in hepatocellular carcinoma cells. Anticancer Res. 2014, 34, 1691–1699. [Google Scholar]
- Lan, C.-Y.; Chen, S.-Y.; Kuo, C.-W.; Lu, C.-C.; Yen, G.-C. Quercetin facilitates cell death and chemosensitivity through RAGE/PI3K/AKT/mTOR axis in human pancreatic cancer cells. J. Food Drug Anal. 2019, 27, 887–896. [Google Scholar] [CrossRef]
- Jia, L.; Huang, S.; Yin, X.; Zan, Y.; Guo, Y.; Han, L. Quercetin suppresses the mobility of breast cancer by suppressing glycolysis through Akt-mTOR pathway mediated autophagy induction. Life Sci. 2018, 208, 123–130. [Google Scholar] [CrossRef]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L.; et al. Quercetin induces protective autophagy in gastric cancer cells: Involvement of Akt-mTOR- and hypoxia-induced factor 1α-mediated signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef]
- Mao, B.; Bu, T.; Mruk, D.; Li, C.; Sun, F.; Cheng, C.Y. Modulating the Blood–Testis Barrier Towards Increasing Drug Delivery. Trends Pharmacol. Sci. 2020, 41, 690–700. [Google Scholar] [CrossRef]
- Maresch, C.C.; Stute, D.C.; Alves, M.G.; Oliveira, P.F.; de Kretser, D.M.; Linn, T. Diabetes-induced hyperglycemia impairs male reproductive function: A systematic review. Hum. Reprod. Update 2017, 24, 86–105. [Google Scholar] [CrossRef] [PubMed]
- Nna, V.U.; Bakar, A.B.A.; Ahmad, A.; Mohamed, M. Down-regulation of steroidogenesis-related genes and its accompanying fertility decline in streptozotocin-induced diabetic male rats: Ameliorative effect of metformin. Andrology 2019, 7, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Khamis, T.; Abdelalim, A.F.; Abdallah, S.H.; Saeed, A.A.; Edress, N.M.; Arisha, A.H. Early intervention with breast milk mesenchymal stem cells attenuates the development of diabetic-induced testicular dysfunction via hypothalamic Kisspeptin/Kiss1r-GnRH/GnIH system in male rats. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165577. [Google Scholar] [CrossRef]
- Tvrdá, E.; Kováč, J.; Ferenczyová, K.; Kaločayová, B.; Ďuračka, M.; Benko, F.; Almášiová, V.; Barteková, M. Quercetin Ameliorates Testicular Damage in Zucker Diabetic Fatty Rats through Its Antioxidant, Anti-Inflammatory and Anti-Apoptotic Properties. Int. J. Mol. Sci. 2022, 23, 16056. [Google Scholar] [CrossRef]
- Kanter, M.; Aktas, C.; Erboga, M. Protective effects of quercetin against apoptosis and oxidative stress in streptozotocin-induced diabetic rat testis. Food Chem. Toxicol. 2012, 50, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Tiss, M.; Hamden, K. Globularia alypum Extracts Attenuate Hyperglycemia and Protect against Various Organ Toxicities in Alloxan-Induced Experimental Diabetic Rats. Evid. Based Complement. Altern. Med. 2022, 2022, 6816942. [Google Scholar] [CrossRef]
- Ali, W.; Ma, Y.; Zhu, J.; Zou, H.; Liu, Z. Mechanisms of Cadmium-Induced Testicular Injury: A Risk to Male Fertility. Cells 2022, 11, 3601. [Google Scholar] [CrossRef]
- Nna, V.U.; Ujah, G.A.; Mohamed, M.; Etim, K.B.; Igba, B.O.; Augustine, E.R.; Osim, E.E. Cadmium chloride–induced testicular toxicity in male wistar rats; prophylactic effect of quercetin, and assessment of testicular recovery following cadmium chloride withdrawal. Biomed. Pharmacother. 2017, 94, 109–123. [Google Scholar] [CrossRef]
- Abarikwu, S.O.; Pant, A.B.; Farombi, E.O. Dietary antioxidant, quercetin, protects sertoli-germ cell coculture from atrazine-induced oxidative damage. J. Biochem. Mol. Toxicol. 2012, 26, 477–485. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, X.; Ge, R.-S. Toxicological Effects of Cadmium on Mammalian Testis. Front. Genet. 2020, 11, 527. [Google Scholar] [CrossRef]
- Behairy, A.; Hashem, M.M.; Abo-El-Sooud, K.; El-Metwally, A.E.; Hassan, B.A.; Abd-Elhakim, Y.M. Quercetin Abates Aluminum Trioxide Nanoparticles and Lead Acetate Induced Altered Sperm Quality, Testicular Oxidative Damage, and Sexual Hormones Disruption in Male Rats. Antioxidants 2022, 11, 2133. [Google Scholar] [CrossRef] [PubMed]
- Farombi, E.O.; Adedara, I.A.; Akinrinde, S.A.; Ojo, O.O.; Eboh, A.S. Protective effects of kolaviron and quercetin on cadmium-induced testicular damage and endocrine pathology in rats. Andrologia 2012, 44, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.; Mi, Y.; Zeng, W.; Zhang, C. Protective effect of quercetin on cadmium-induced oxidative toxicity on germ cells in male mice. Anat. Rec. 2011, 294, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Benoff, S.; Jacob, A.; Hurley, I.R. Male infertility and environmental exposure to lead and cadmium. Hum. Reprod. Update 2000, 6, 107–121. [Google Scholar] [CrossRef]
- Khodabandeh, Z.; Dolati, P.; Zamiri, M.J.; Mehrabani, D.; Bordbar, H.; Alaee, S.; Jamhiri, I.; Azarpira, N. Protective Effect of Quercetin on Testis Structure and Apoptosis Against Lead Acetate Toxicity: An Stereological Study. Biol. Trace Elem. Res. 2021, 199, 3371–3381. [Google Scholar] [CrossRef] [PubMed]
- Dolati, P.; Khodabandeh, Z.; Zamiri, M.J.; Jamhiri, I.; Mehrabani, D. The Effect of Lead Acetate and Quercetin on the Tight and Gap Junctions in the Mouse Testis. Biol. Trace Elem. Res. 2020, 198, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, J.K.; Panchal, H. Quercetin mediated attenuation of cadmium-induced oxidative toxicity and apoptosis of spermatogenic cells in caprine testes in vitro. Environ. Mol. Mutagen 2021, 62, 374–384. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, H.; Wang, K.; Yang, Z.; Liu, Z. Protective effect of quercetin on rat testes against cadmium toxicity by alleviating oxidative stress and autophagy. Environ. Sci. Pollut. Res. 2020, 27, 25278–25286. [Google Scholar] [CrossRef]
- Ujah, G.A.; Nna, V.U.; Agah, M.I.; Omue, L.O.; Leku, C.B.; Osim, E.E. Effect of quercetin on cadmium chloride-induced impairments in sexual behaviour and steroidogenesis in male Wistar rats. Andrologia 2018, 50, e12866. [Google Scholar] [CrossRef]
- Adedara, I.A.; Subair, T.I.; Ego, V.C.; Oyediran, O.; Farombi, E.O. Chemoprotective role of quercetin in manganese-induced toxicity along the brain-pituitary-testicular axis in rats. Chem. Biol. Interact. 2017, 263, 88–98. [Google Scholar] [CrossRef]
- Hussein, M.M.A.; Ali, H.A.; Saadeldin, I.M.; Ahmed, M.M. Querectin Alleviates Zinc Oxide Nanoreprotoxicity in Male Albino Rats. J. Biochem. Mol. Toxicol. 2016, 30, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Khorsandi, L.; Orazizadeh, M.; Moradi-Gharibvand, N.; Hemadi, M.; Mansouri, E. Beneficial effects of quercetin on titanium dioxide nanoparticles induced spermatogenesis defects in mice. Environ. Sci. Pollut. Res. Int. 2017, 24, 5595–5606. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Pant, A.B.; Farombi, E.O. Effects of quercetin on mRNA expression of steroidogenesis genes in primary cultures of Leydig cells treated with atrazine. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2013, 27, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Pant, A.B.; Farombi, E.O. Quercetin decreases steroidogenic enzyme activity, NF-κB expression, and oxidative stress in cultured Leydig cells exposed to atrazine. Mol. Cell. Biochem. 2013, 373, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Abdel Aziz, R.L.; Abdel-Wahab, A.; Abo El-Ela, F.I.; Hassan, N.E.-H.Y.; El-Nahass, E.-S.; Ibrahim, M.A.; Khalil, A.-T.A.Y. Dose- dependent ameliorative effects of quercetin and l-Carnitine against atrazine- induced reproductive toxicity in adult male Albino rats. Biomed. Pharmacother. 2018, 102, 855–864. [Google Scholar] [CrossRef]
- Ndufeiya-Kumasi, L.C.; Abarikwu, S.O.; Ohanador, R.; Omoregie, E.S. Curcumin improves the protective effects of quercetin against atrazine-induced testicular injury in adult Wistar rats. Andrologia 2022, 54, e14445. [Google Scholar] [CrossRef]
- Baltaci, B.B.; Uygur, R.; Caglar, V.; Aktas, C.; Aydin, M.; Ozen, O.A. Protective effects of quercetin against arsenic-induced testicular damage in rats. Andrologia 2016, 48, 1202–1213. [Google Scholar] [CrossRef]
- Jahan, S.; Iftikhar, N.; Ullah, H.; Rukh, G.; Hussain, I. Alleviative effect of quercetin on rat testis against arsenic: A histological and biochemical study. Syst. Biol. Reprod. Med. 2015, 61, 89–95. [Google Scholar] [CrossRef]
- Samova, S.; Patel, C.N.; Doctor, H.; Pandya, H.A.; Verma, R.J. The effect of bisphenol A on testicular steroidogenesis and its amelioration by quercetin: An in vivo and in silico approach. Toxicol. Res. 2018, 7, 22–31. [Google Scholar] [CrossRef]
- Khazaeel, K.; Daaj, S.A.Z.; Sadeghi, A.; Tabandeh, M.R.; Basir, Z. Potential protective effect of quercetin on the male reproductive system against exposure of Wistar rats to crude oil vapor: Genetic, biochemical, and histopathological evidence. Reprod. Toxicol. 2022, 113, 10–17. [Google Scholar] [CrossRef]
- Izawa, H.; Kohara, M.; Aizawa, K.; Suganuma, H.; Inakuma, T.; Watanabe, G.; Taya, K.; Sagai, M. Alleviative effects of quercetin and onion on male reproductive toxicity induced by diesel exhaust particles. Biosci. Biotechnol. Biochem. 2008, 72, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Lovato, F.L.; de Oliveira, C.R.; Adedara, I.A.; Barbisan, F.; Moreira, K.L.S.; Dalberto, M.; da Rocha, M.I.U.M.; Marroni, N.P.; da Cruz, I.B.; Costabeber, I.B. Quercetin ameliorates polychlorinated biphenyls-induced testicular DNA damage in rats. Andrologia 2016, 48, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Sönmez, M.; Türk, G.; Çeribaşı, S.; Çiftçi, M.; Yüce, A.; Güvenç, M.; Özer Kaya, S.; Çay, M.; Aksakal, M. Quercetin attenuates carbon tetrachloride-induced testicular damage in rats. Andrologia 2014, 46, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Tu, L.; Wang, H.; Zeng, W.; Zhang, C. Supplementation with quercetin attenuates 4-nitrophenol-induced testicular toxicity in adult male mice. Anat. Rec. 2013, 296, 1650–1657. [Google Scholar] [CrossRef]
- Ciftci, O.; Aydin, M.; Ozdemir, I.; Vardi, N. Quercetin prevents 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced testicular damage in rats. Andrologia 2012, 44, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Oluwabayo, T.O.; Akinmoladun, A.C.; Akindahunsi, A.A. Arrested Acetylene-Induced Pulmonary and Testicular Toxicity in Rats Through Treatment with Polyphenols. Drug Chem. Toxicol. 2022, 46, 1–12. [Google Scholar] [CrossRef]
- Xia, L.-Z.; Jiang, M.-Z.; Liu, L.-L.; Wu, Y.; Zhang, Y.-L.; Yang, L.-X.; Shen, X.-Y.; Zhang, Q.-Y.; Lin, M.; Gao, H.-T. Quercetin inhibits testicular toxicity induced by the mixture of three commonly used phthalates in rats. J. Sci. Food Agric. 2023, 103, 1541–1549. [Google Scholar] [CrossRef]
- Abd-Ellah, M.F.; Aly, H.A.A.; Mokhlis, H.A.M.; Abdel-Aziz, A.H. Quercetin attenuates di-(2-ethylhexyl) phthalate-induced testicular toxicity in adult rats. Hum. Exp. Toxicol. 2016, 35, 232–243. [Google Scholar] [CrossRef]
- Ahmed, Z.A.; Abtar, A.N.; Othman, H.H.; Aziz, T.A. Effects of quercetin, sitagliptin alone or in combination in testicular toxicity induced by doxorubicin in rats. Drug Des. Dev. Ther. 2019, 13, 3321–3329. [Google Scholar] [CrossRef]
- Ebokaiwe, A.P.; Obasi, D.O.; Njoku, R.C.; Osawe, S. Cyclophosphamide-induced testicular oxidative-inflammatory injury is accompanied by altered immunosuppressive indoleamine 2, 3-dioxygenase in Wister rats: Influence of dietary quercetin. Andrologia 2022, 54, e14341. [Google Scholar] [CrossRef]
- Bostancıeri, N.; Taşlidere, A.; Elbe, H.; Taşlıdere, E. Protective effects of quercetin against testis damage caused by cisplatin. Biotech. Histochem. Off. Publ. Biol. Stain Comm. 2022, 97, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Aldemir, M.; Okulu, E.; Kösemehmetoğlu, K.; Ener, K.; Topal, F.; Evirgen, O.; Gürleyik, E.; Avcı, A. Evaluation of the protective effect of quercetin against cisplatin-induced renal and testis tissue damage and sperm parameters in rats. Andrologia 2014, 46, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Altintas, R.; Ciftci, O.; Aydin, M.; Akpolat, N.; Oguz, F.; Beytur, A. Quercetin prevents docetaxel-induced testicular damage in rats. Andrologia 2015, 47, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Selim, M.E.; Aleisa, N.A.; Daghestani, M.H. Evaluation of the Possible Protective Role of Quercetin on Letrozole-induced Testicular Injury in Male Albino Rats. Ultrastruct. Pathol. 2013, 37, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yu, Q.; Zhao, F.; Ji, J.; Jiang, Z.; Chen, X.; Gao, P.; Ren, Y.; Shao, S.; Zhang, L.; et al. Protection of Quercetin against Triptolide-induced apoptosis by suppressing oxidative stress in rat Leydig cells. Chem. Biol. Interact. 2015, 240, 38–46. [Google Scholar] [CrossRef]
- Osawe, S.O.; Farombi, E.O. Quercetin and rutin ameliorates sulphasalazine-induced spermiotoxicity, alterations in reproductive hormones and steroidogenic enzyme imbalance in rats. Andrologia 2018, 50, e12981. [Google Scholar] [CrossRef]
- Chi, K.-K.; Zhang, W.-H.; Chen, Z.; Cui, Y.; He, W.; Wang, S.-G.; Zhang, C.; Chen, J.; Wang, G.-C. Comparison of quercetin and resveratrol in the prevention of injury due to testicular torsion/detorsion in rats. Asian J. Androl. 2016, 18, 908–912. [Google Scholar] [CrossRef]
- Aktoz, T.; Kanter, M.; Aktas, C. Protective effects of quercetin on testicular torsion/detorsion-induced ischaemia-reperfusion injury in rats. Andrologia 2010, 42, 376–383. [Google Scholar] [CrossRef]
- Aldemir, M.; Özgün, G.; Önen, E.; Okulu, E.; Kayıgil, Ö. Quercetin has a protective role on histopathological findings on testicular ischaemia-reperfusion injury in rats. Andrologia 2012, 44 (Suppl. S1), 479–483. [Google Scholar] [CrossRef]
- Brüll, V.; Burak, C.; Stoffel-Wagner, B.; Wolffram, S.; Nickenig, G.; Müller, C.; Langguth, P.; Alteheld, B.; Fimmers, R.; Naaf, S.; et al. Effects of a quercetin-rich onion skin extract on 24 h ambulatory blood pressure and endothelial function in overweight-to-obese patients with (pre-)hypertension: A randomised double-blinded placebo-controlled cross-over trial. Br. J. Nutr. 2015, 114, 1263–1277. [Google Scholar] [CrossRef]
- Edwards, R.L.; Lyon, T.; Litwin, S.E.; Rabovsky, A.; Symons, J.D.; Jalili, T. Quercetin Reduces Blood Pressure in Hypertensive Subjects. J. Nutr. 2007, 137, 2405–2411. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Boesch-Saadatmandi, C.; Wolffram, S.; Rimbach, G.; Müller, M.J. Serum lipid and blood pressure responses to quercetin vary in overweight patients by apolipoprotein E genotype. J. Nutr. 2010, 140, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Bosy-Westphal, A.; Seiberl, J.; Kürbitz, C.; Settler, U.; Plachta-Danielzik, S.; Wagner, A.E.; Frank, J.; Schrezenmeir, J.; Rimbach, G.; et al. Quercetin reduces systolic blood pressure and plasma oxidised low-density lipoprotein concentrations in overweight subjects with a high-cardiovascular disease risk phenotype: A double-blinded, placebo-controlled cross-over study. Br. J. Nutr. 2009, 102, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-Y.; Lee, H.; Woo, J.S.; Jang, H.H.; Hwang, S.J.; Kim, H.S.; Kim, W.-S.; Kim, Y.-S.; Choue, R.; Cha, Y.-J.; et al. Effect of onion peel extract on endothelial function and endothelial progenitor cells in overweight and obese individuals. Nutrition 2015, 31, 1131–1135. [Google Scholar] [CrossRef]
- Brüll, V.; Burak, C.; Stoffel-Wagner, B.; Wolffram, S.; Nickenig, G.; Müller, C.; Langguth, P.; Alteheld, B.; Fimmers, R.; Stehle, P.; et al. Acute intake of quercetin from onion skin extract does not influence postprandial blood pressure and endothelial function in overweight-to-obese adults with hypertension: A randomized, double-blind, placebo-controlled, crossover trial. Eur. J. Nutr. 2017, 56, 1347–1357. [Google Scholar] [CrossRef]
- Chaib, S.; Tchkonia, T.; Kirkland, J.L. Cellular senescence and senolytics: The path to the clinic. Nat. Med. 2022, 28, 1556–1568. [Google Scholar] [CrossRef]
- Justice, J.N.; Nambiar, A.M.; Tchkonia, T.; LeBrasseur, N.K.; Pascual, R.; Hashmi, S.K.; Prata, L.; Masternak, M.M.; Kritchevsky, S.B.; Musi, N.; et al. Senolytics in idiopathic pulmonary fibrosis: Results from a first-in-human, open-label, pilot study. EBioMedicine 2019, 40, 554–563. [Google Scholar] [CrossRef]
- Gonzales, M.M.; Garbarino, V.R.; Marques Zilli, E.; Petersen, R.C.; Kirkland, J.L.; Tchkonia, T.; Musi, N.; Seshadri, S.; Craft, S.; Orr, M.E. Senolytic Therapy to Modulate the Progression of Alzheimer’s Disease (SToMP-AD): A Pilot Clinical Trial. J. Prev. Alzheimer’s Dis. 2022, 9, 22–29. [Google Scholar] [CrossRef]
- Hickson, L.J.; Langhi Prata, L.G.P.; Bobart, S.A.; Evans, T.K.; Giorgadze, N.; Hashmi, S.K.; Herrmann, S.M.; Jensen, M.D.; Jia, Q.; Jordan, K.L.; et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine 2019, 47, 446–456. [Google Scholar] [CrossRef]
- Javadi, F.; Ahmadzadeh, A.; Eghtesadi, S.; Aryaeian, N.; Zabihiyeganeh, M.; Rahimi Foroushani, A.; Jazayeri, S. The Effect of Quercetin on Inflammatory Factors and Clinical Symptoms in Women with Rheumatoid Arthritis: A Double-Blind, Randomized Controlled Trial. J. Am. Coll. Nutr. 2017, 36, 9–15. [Google Scholar] [CrossRef]
- Dehghani, F.; Sezavar Seyedi Jandaghi, S.H.; Janani, L.; Sarebanhassanabadi, M.; Emamat, H.; Vafa, M. Effects of quercetin supplementation on inflammatory factors and quality of life in post-myocardial infarction patients: A double blind, placebo-controlled, randomized clinical trial. Phytother. Res. 2021, 35, 2085–2098. [Google Scholar] [CrossRef] [PubMed]
- Khorshidi, M.; Moini, A.; Alipoor, E.; Rezvan, N.; Gorgani-Firuzjaee, S.; Yaseri, M.; Hosseinzadeh-Attar, M.J. The effects of quercetin supplementation on metabolic and hormonal parameters as well as plasma concentration and gene expression of resistin in overweight or obese women with polycystic ovary syndrome. Phytother. Res. 2018, 32, 2282–2289. [Google Scholar] [CrossRef] [PubMed]
- Rezvan, N.; Moini, A.; Janani, L.; Mohammad, K.; Saedisomeolia, A.; Nourbakhsh, M.; Gorgani-Firuzjaee, S.; Mazaherioun, M.; Hosseinzadeh-Attar, M.J. Effects of Quercetin on Adiponectin-Mediated Insulin Sensitivity in Polycystic Ovary Syndrome: A Randomized Placebo-Controlled Double-Blind Clinical Trial. Horm. Metab. Res. 2017, 49, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Sajadi Hezaveh, Z.; Azarkeivan, A.; Janani, L.; Hosseini, S.; Shidfar, F. The effect of quercetin on iron overload and inflammation in β-thalassemia major patients: A double-blind randomized clinical trial. Complement. Ther. Med. 2019, 46, 24–28. [Google Scholar] [CrossRef]
- Torella, M.; Del Deo, F.; Grimaldi, A.; Iervolino, S.A.; Pezzella, M.; Tammaro, C.; Gallo, P.; Rappa, C.; De Franciscis, P.; Colacurci, N. Efficacy of an orally administered combination of hyaluronic acid, chondroitin sulfate, curcumin and quercetin for the prevention of recurrent urinary tract infections in postmenopausal women. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 207, 125–128. [Google Scholar] [CrossRef]
- Corsale, I.; Carrieri, P.; Martellucci, J.; Piccolomini, A.; Verre, L.; Rigutini, M.; Panicucci, S. Flavonoid mixture (diosmin, troxerutin, rutin, hesperidin, quercetin) in the treatment of I-IIIdegree hemorroidal disease: A double-blind multicenter prospective comparative study. Int. J. Color. Dis. 2018, 33, 1595–1600. [Google Scholar] [CrossRef]
- Boots, A.W.; Drent, M.; de Boer, V.C.J.; Bast, A.; Haenen, G.R.M.M. Quercetin reduces markers of oxidative stress and inflammation in sarcoidosis. Clin. Nutr. 2011, 30, 506–512. [Google Scholar] [CrossRef]
- Gallelli, G.; Cione, E.; Serra, R.; Leo, A.; Citraro, R.; Matricardi, P.; Di Meo, C.; Bisceglia, F.; Caroleo, M.C.; Basile, S.; et al. Nano-hydrogel embedded with quercetin and oleic acid as a new formulation in the treatment of diabetic foot ulcer: A pilot study. Int. Wound J. 2020, 17, 485–490. [Google Scholar] [CrossRef]
- Lu, N.T.; Crespi, C.M.; Liu, N.M.; Vu, J.Q.; Ahmadieh, Y.; Wu, S.; Lin, S.; McClune, A.; Durazo, F.; Saab, S.; et al. A Phase I Dose Escalation Study Demonstrates Quercetin Safety and Explores Potential for Bioflavonoid Antivirals in Patients with Chronic Hepatitis C. Phytother. Res. PTR 2016, 30, 160–168. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Y.; Miao, C.; Yang, L.; Wang, R.; Chen, B.; Zhang, Q. Quercetin alleviates cyclophosphamide-induced premature ovarian insufficiency in mice by reducing mitochondrial oxidative stress and pyroptosis in granulosa cells. J. Ovarian Res. 2022, 15, 138. [Google Scholar] [CrossRef]
- Elkady, M.A.; Shalaby, S.; Fathi, F.; El-Mandouh, S. Effects of quercetin and rosuvastatin each alone or in combination on cyclophosphamide-induced premature ovarian failure in female albino mice. Hum. Exp. Toxicol. 2019, 38, 1283–1295. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.P.; Kiparissis, Y.; Hwang, G.S.; Niimi, A.J.; Janz, D.M.; Metcalfe, C.D. Increased cellular apoptosis after chronic aqueous exposure to nonylphenol and quercetin in adult medaka (Oryzias latipes). Comp. Biochem. Physiol. Toxicol. Pharmacol. 2002, 131, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Ranawat, P.; Kaushik, G.; Saikia, U.N.; Pathak, C.M.; Khanduja, K.L. Quercetin impairs the reproductive potential of male mice. Andrologia 2013, 45, 56–65. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Z. Exploring the role of gut microbiome in male reproduction. Andrology 2022, 10, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-B.; Chen, K.; Li, Z.-F.; Wang, Z.-Y.; Wang, L. Glyphosate-induced gut microbiota dysbiosis facilitates male reproductive toxicity in rats. Sci. Total. Environ. 2022, 805, 150368. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Cai, D.; Li, X.; Liu, B.; Chen, J.; Jiang, X.; Li, H.; Li, Z.; Teerds, K.; Sun, J.; et al. Effects of Bisphenol A on reproductive toxicity and gut microbiota dysbiosis in male rats. Ecotoxicol. Environ. Saf. 2022, 239, 113623. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Liu, Y. Microorganisms in the reproductive system and probiotic’s regulatory effects on reproductive health. Comput. Struct. Biotechnol. J. 2022, 20, 1541–1553. [Google Scholar] [CrossRef]
- Tian, X.; Yu, Z.; Feng, P.; Ye, Z.; Li, R.; Liu, J.; Hu, J.; Kakade, A.; Liu, P.; Li, X. Lactobacillus plantarum TW1-1 Alleviates Diethylhexylphthalate-Induced Testicular Damage in Mice by Modulating Gut Microbiota and Decreasing Inflammation. Front. Cell. Infect. Microbiol. 2019, 9, 221. [Google Scholar] [CrossRef]
- Zhu, Y.; Du, Q.; Jiao, N.; Shu, A.; Gao, Y.; Chen, J.; Lv, G.; Lu, J.; Chen, Y.; Xu, H. Catalpol ameliorates diabetes-induced testicular injury and modulates gut microbiota. Life Sci. 2021, 267, 118881. [Google Scholar] [CrossRef]
- Chen, G.; Wang, G.; Zhu, C.; Jiang, X.; Sun, J.; Tian, L.; Bai, W. Effects of cyanidin-3-O-glucoside on 3-chloro-1,2-propanediol induced intestinal microbiota dysbiosis in rats. Food Chem. Toxicol. 2019, 133, 110767. [Google Scholar] [CrossRef]
- Rakhshandeh, H.; Rajabi Khasevan, H.; Saviano, A.; Mahdinezhad, M.R.; Baradaran Rahimi, V.; Ehtiati, S.; Etemad, L.; Ebrahimzadeh-Bideskan, A.; Maione, F.; Askari, V.R. Protective Effect of Portulaca oleracea on Streptozotocin-Induced Type I Diabetes-Associated Reproductive System Dysfunction and Inflammation. Molecules 2022, 27, 6075. [Google Scholar] [CrossRef] [PubMed]
- Rotimi, D.E.; Adeyemi, O.S. Plantain-based diet decreases oxidative stress and inflammatory markers in the testes of rats exposed to atrazine. Mol. Cell. Biochem. 2023, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.M.; Hussein, M.M.A.; Saber, T.; Abd-Elhakim, Y.M. Palliative Effect of Resveratrol against Nanosized Iron Oxide-Induced Oxidative Stress and Steroidogenesis-Related Genes Dysregulation in Testicular Tissue of Adult Male Rats. Int. J. Env. Res. Public Health 2022, 19, 8171. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Song, W.; Wang, Z.; Wang, Z.; Jin, X.; Xu, J.; Bai, L.; Li, Y.; Cui, J.; Cai, L. Resveratrol attenuates testicular apoptosis in type 1 diabetic mice: Role of Akt-mediated Nrf2 activation and p62-dependent Keap1 degradation. Redox Biol. 2018, 14, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, M.F.; Hussain, A.; Sameen, A.; Sahar, A.; Khan, S.; Siddique, R.; Aadil, R.M.; Xu, B. Novel extraction, rapid assessment and bioavailability improvement of quercetin: A review. Ultrason. Sonochemistry 2021, 78, 105686. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Types of Pollutants | Exposure Time and Dose | Animal Model | Quercetin Dose and Duration | The Effect of Quercetin | Reference | |
|---|---|---|---|---|---|---|
| Heavy metals | Pb | PbAc: 50 mg/kg bw/d, oral for 60 days Al2O3NPs: 100 mg/kg bw/d, oral for 60 days | SD rats | 20 mg/kg bw/d, oral for 60 days | reversed adverse effects on testis weight, improved sperm parameters, increased serum testosterone level, increased antioxidant enzymes and decreased malondialdehyde (MDA), improved histology, decreased pro-inflammatory cytokines. | [71] |
| 150 mg/kg bw/d, gavage for 34.5 days (one cycle of the seminiferous epithelium) | NMRI strain male mice | 75 mg/kg bw/d, gavage for 34.5 days | restored morphological changes, increased number of testicular cells, reduced apoptosis level. | [75] | ||
| 150 mg/kg bw/d, oral gavage for at least 35 days | NMRI strain male mice | 75 mg/kg bw/d, oral gavage for at least 35 days | increased the expression of connexin 43 (C × 43). | [76] | ||
| Cd | 5 mg/kg bw/d, oral for 4 weeks | Wistar rats | 20 mg/kg bw/d, oral for 4 weeks | improved testis weight, recovered semen quality and serum reproductive hormones, decreased glucose, lactate, and lactate dehydrogenase (LDH) in testis, increased enzymatic and non-enzymatic antioxidants level. | [68] | |
| 15 mg/kg bw/d, oral for 5 days | Wistar rats | 10 mg/kg bw/d, oral for 8 days (including 3 days of pretreatment) | improved the sperm characteristics, increased plasma hormone level, reversed superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), and glutathione S-transferase (GST) activities and restored the H2O2 and MDA levels, attenuated histological damage. | [72] | ||
| 4 mg/kg bw/d, oral for 2 weeks | ICR mice | 75 mg/kg bw/d, oral for 2 weeks | enhanced antioxidant capacity, inhibited cell apoptosis. | [73] | ||
| 2 mg/kg bw/d, intraperitoneal injection for 4 weeks | SD rats | 50 mg/kg bw/d, intragastrical administration for 4 weeks | reversed bad effects on testis weight and body weight, relieved oxidative stress status, alleviated histopathological disorder, ameliorated the expression of P62 and LC3-II. | [78] | ||
| 5 mg/kg bw/d, oral for 4 weeks | Wistar rats | 20 mg/kg bw/d, oral for 4 weeks | reversed adverse effect to sexual behavior, downregulated serum NO concentration and testicular cholesterol concentration, recovered 3β-hydroxysteroid dehydrogenase (3β-HSD) and 17β-HSD activity, increased testicular testosterone concentration. | [79] | ||
| Mn | 15 mg/kg bw/d, oral for 45 days | Wistar rats | 10 or 20 mg/kg bw/d, oral for 45 days | increased antioxidant enzyme activities, decreased inflammatory biomarkers and cleaved caspase-3 in the brain, testes, and epididymis, modulated circulatory hormones concentration and marker enzymes of testicular function, augmented sperm functional parameters, prevented histological changes. | [80] | |
| Zn | 100 or 400 mg/kg bw/d, intragastric intubation for 12 weeks | Albino rats | 100 mg/kg bw/d, intragastric intubation for 12 weeks | improved sperm parameters, ameliorated oxidative stress, restored testosterone level and steroidogenesis, improved histology. | [81] | |
| Ti | 300 mg/kg bw/d, oral gavage for 35 days | NMRI mice | 75 mg/kg bw/d, oral gavage for 42 days | increased testis weight, increased serum testosterone level and testosterone concentration in testis, increased sperm count and motility, attenuated histopathological damage, decreased apoptotic index, improved SOD and CAT activities, attenuated MDA activities. | [82] | |
| Other environmental pollutants | Atrazine | 232 μM for 6 h | Wistar ratsLeydig cells | 50 μM for 6 h | normalized the expressions of steroidogenesis genes. | [83] |
| 50 μg/mL for 6 or 24 h | Wistar ratsLeydig cells | 50 μM 6 or 24 h | recovered cell viability, improved oxidative stress and lipid peroxidation, prevented the activities of steroidogenesis enzymes, restored nuclear factor kappa-B (NF-κB) mRNA and protein levels. | [84] | ||
| 232 μM for 6 or 24 h | Wistar rats Sertoli cells | 50 μM for 6 or 24 h | improved cell viability, attenuated oxidative damage; upregulated SOD-1, GPx, glutathione reductase (GR), and GST expressions. | [69] | ||
| 120 mg/kg bw/d, oral gavage for 21 days | Albino rats | 10–50 mg/kg bw/d, oral gavage for 21 days | increased the body weight, improved the antioxidant capacity, restored serum testosterone and sperm morphology, increased the serum IgA, inhibited the percentage of DNA fragmentation, disturbed the cytochrome P450 family 17 subfamily A member 1 (Cyp17a1) mRNA expression, slightly improved histology. | [85] | ||
| 50 mg/kg bw/d, oral every other day for 60 days. | Wistar rats | 5–10 mg/kg bw/d, oral every other day for 60 days. | reduced GSH and GST concentrations, decreased MDA concentrations, enhanced anti-inflammatory effects, reversed serum hormones level, increased morphometric parameters and sperm quality parameters, improved histopathology. | [86] | ||
| As | 10 mg/kg bw/d, oral gavage for 15 days | SD rats | 50 mg/kg bw/d, oral gavage for 15 days | lessened terminal dUTP nick end-labeling (TUNEL)-positive germ cells, improved proliferating cell nuclear antigen (PCNA)-positive cells, amplified antioxidant effect. | [87] | |
| 50 ppm in drinking water for 49 days | SD rats | 50 mg/kg bw/d, oral gavage for 49 days | ameliorated histology and morphometric, increased antioxidant enzymes and testosterone level in testis and plasma. | [88] | ||
| BPA | 80–240 mg/kg bw/d, oral gavage for 45 days | Swiss strain albino mice | 30–90 mg/kg bw/d, oral gavage for 45 days | increased body weight and testis weight, increased serum testosterone level and the activity of steroidogenic enzymes. | [89] | |
| COV | inhalation for 5 h daily in pollution chamber for 30 days | Wistar rats | 50 mg/kg bw/d, oral for 30 days | improved testicular weight, gonadosomatic index and sperm parameters, alleviated histopathological damage, reduced apoptotic, increased antioxidant capacity. | [90] | |
| DEPs | 220 μg/mouse, subcutaneous injection for 10 times during 5 weeks | BALB/cmice | Feed with 0.3%, 0.1%, 0.03% w/w quercetin, oral for 30 days in CE-2 commercial diets | restored spermatogenesis and sperm morphological abnormalities, increased the numbers of Sertoli cells. | [91] | |
| PCBs | 2 mg/kg bw/d, intraperitoneal injection for 25 days | Wistar rats | 50 mg/kg bw/d, oral for 25 days | restored oxidative stress indices, reversed DNA fragmentation, reversed histological damage. | [92] | |
| CCl4 | 0.25 mL/kg bw/w, oral gavage for 10 weeks | Wistar rats | 150 mg/kg bw/d, oral gavage for 10 weeks | decreased MDA level, improved abnormal sperm rate, reduced histopathological lesions and apoptosis in testis. | [93] | |
| PNP | 50 mg/kg bw/d, intraperitoneal injection for 6 weeks | ICR mice | 75 mg/kg bw/d, intraperitoneal injection for 6 weeks | attenuated histopathological damage; changed antioxidant status caspase-3 activity and number of TUNEL-positive cells; altered BCL-2 associated X protein (BAX), B-cell lymphoma-extra large (BCL-XL), X Box binding protein (XBP-1), and heme oxygenase-1 (HO-1) expression. | [94] | |
| TCDD | 2 μg/kg bw/w, oral gavage for 60 days | Wistar rats | 20 mg/kg bw/d, oral gavage for 60 days | increased antioxidant capacity, increased sperm parameters and testis weight, reversed histopathological changes, increased serum testosterone levels. | [95] | |
| acetylene | inhalation 20 min daily for 30 days in pollution chamber with 58,000 ppm acetylene | Wistar rats | 30 mg/kg bw/d for 30 days | improved biochemical indexes and histopathological damage. | [96] | |
| PEs | 900 mg/kg bw/d, oral for 30 days | SD rats | 10, 30 and 90 mg/kg bw/d, oral for 30 days | increased testis weight and epididymis weight, increased serum testosterone, luteinizing hormone (LH), follicle-stimulating hormone (FSH), and estradiol level, inhibited testicular injuries, downregulated steroidogenic proteins expression, restored spermatogenesis. | [97] | |
| 300, 600, or 900 mg/kg bw/d, oral gavage for 15 days | Wistar rats | 90 mg/kg bw/d, 24 h before DEHP treatment | improved relative testes weight and sperm parameters, increased serum testosterone and prostatic acid phosphatase, reverted antioxidant enzyme activities, ameliorated histologic alterations. | [98] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Tang, Y.; Lu, G.; Gu, J. Pharmacological Activity of Flavonoid Quercetin and Its Therapeutic Potential in Testicular Injury. Nutrients 2023, 15, 2231. https://doi.org/10.3390/nu15092231
Zhang X, Tang Y, Lu G, Gu J. Pharmacological Activity of Flavonoid Quercetin and Its Therapeutic Potential in Testicular Injury. Nutrients. 2023; 15(9):2231. https://doi.org/10.3390/nu15092231
Chicago/Turabian StyleZhang, Xiaohui, Yufeng Tang, Guangping Lu, and Junlian Gu. 2023. "Pharmacological Activity of Flavonoid Quercetin and Its Therapeutic Potential in Testicular Injury" Nutrients 15, no. 9: 2231. https://doi.org/10.3390/nu15092231
APA StyleZhang, X., Tang, Y., Lu, G., & Gu, J. (2023). Pharmacological Activity of Flavonoid Quercetin and Its Therapeutic Potential in Testicular Injury. Nutrients, 15(9), 2231. https://doi.org/10.3390/nu15092231