
Plum-Derived Exosome-like Nanovesicles Induce Differentiation of Osteoblasts and Reduction of Osteoclast Activation

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
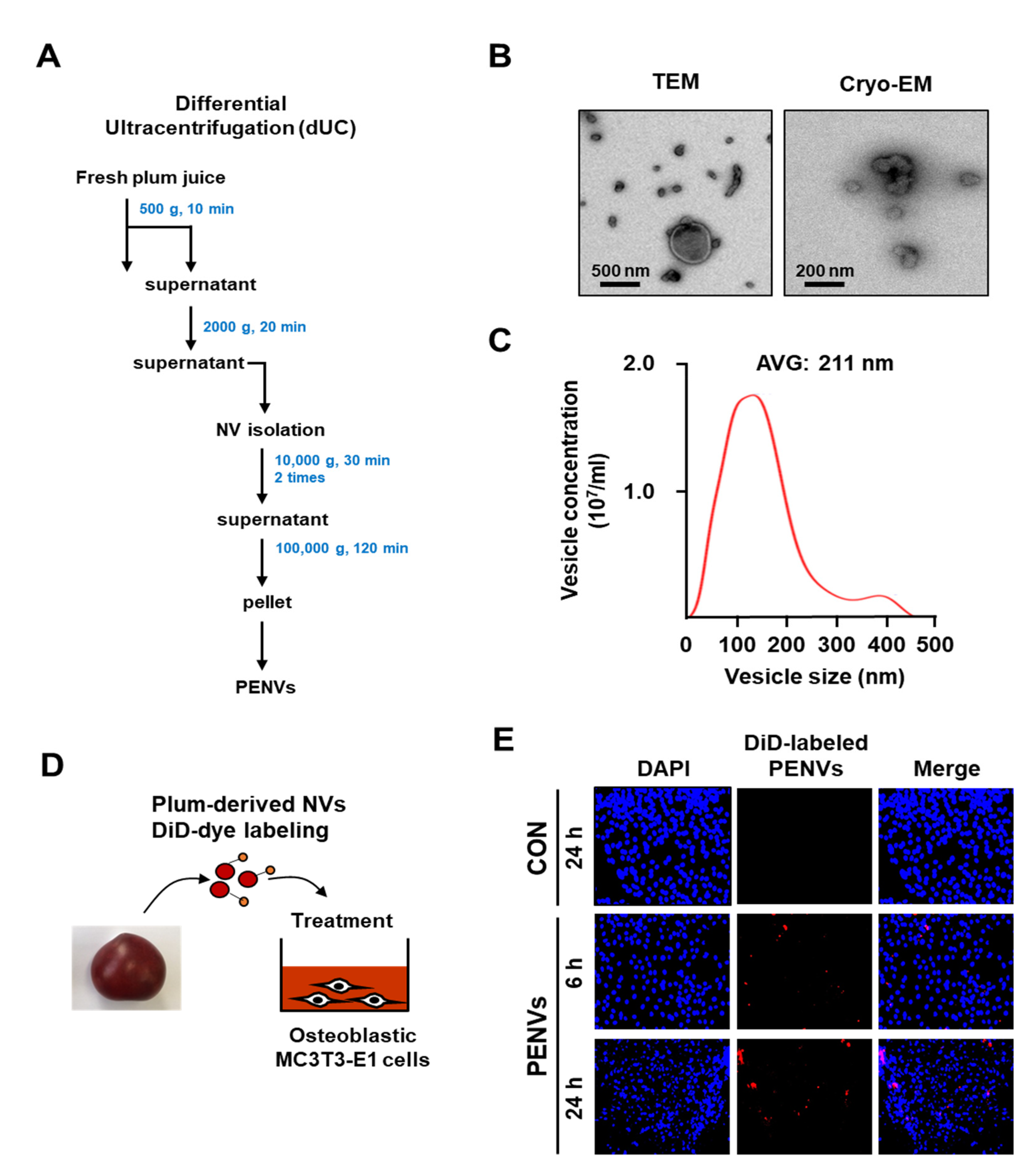
2.3. Purification and Isolation of Plum-Derived Exosome-like Nanovesicles
2.4. Nanoparticle Tracking Analysis (NTA)
2.5. Transmission Electron Microscopy (TEM) and Cryogenic Electron Microscopy (Cryo-EM)
2.6. Labeling of PENVs and Uptake of Labeled PENVs in Cell Cultures
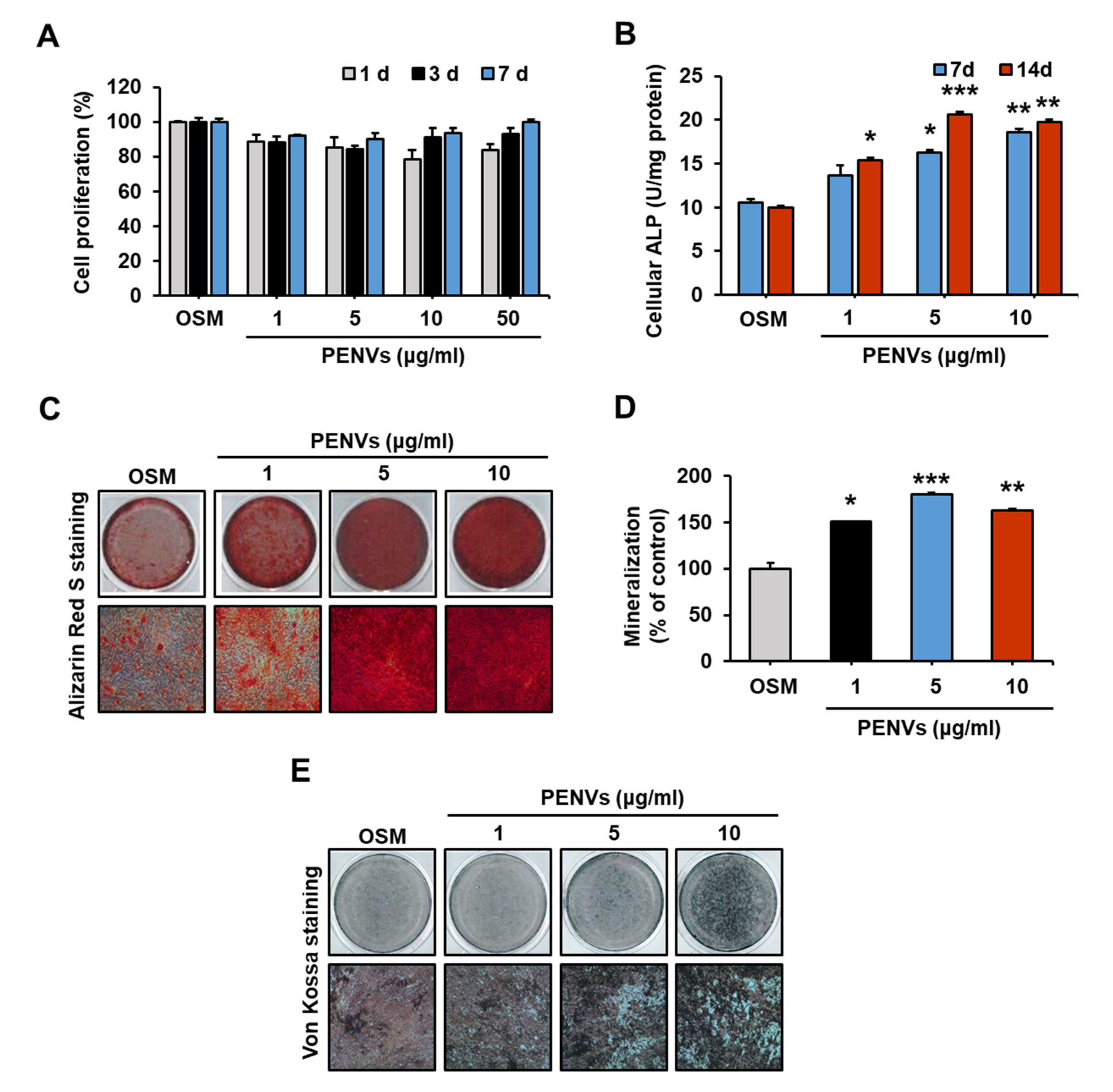
2.7. MTT Assay
2.8. Determination of Alkaline Phosphatase (ALP) Activity
2.9. Alizarin Red S and von Kossa Staining
2.10. Tartrate-Resistant Acid Phosphatase (TRAP) Staining and Activity Analysis
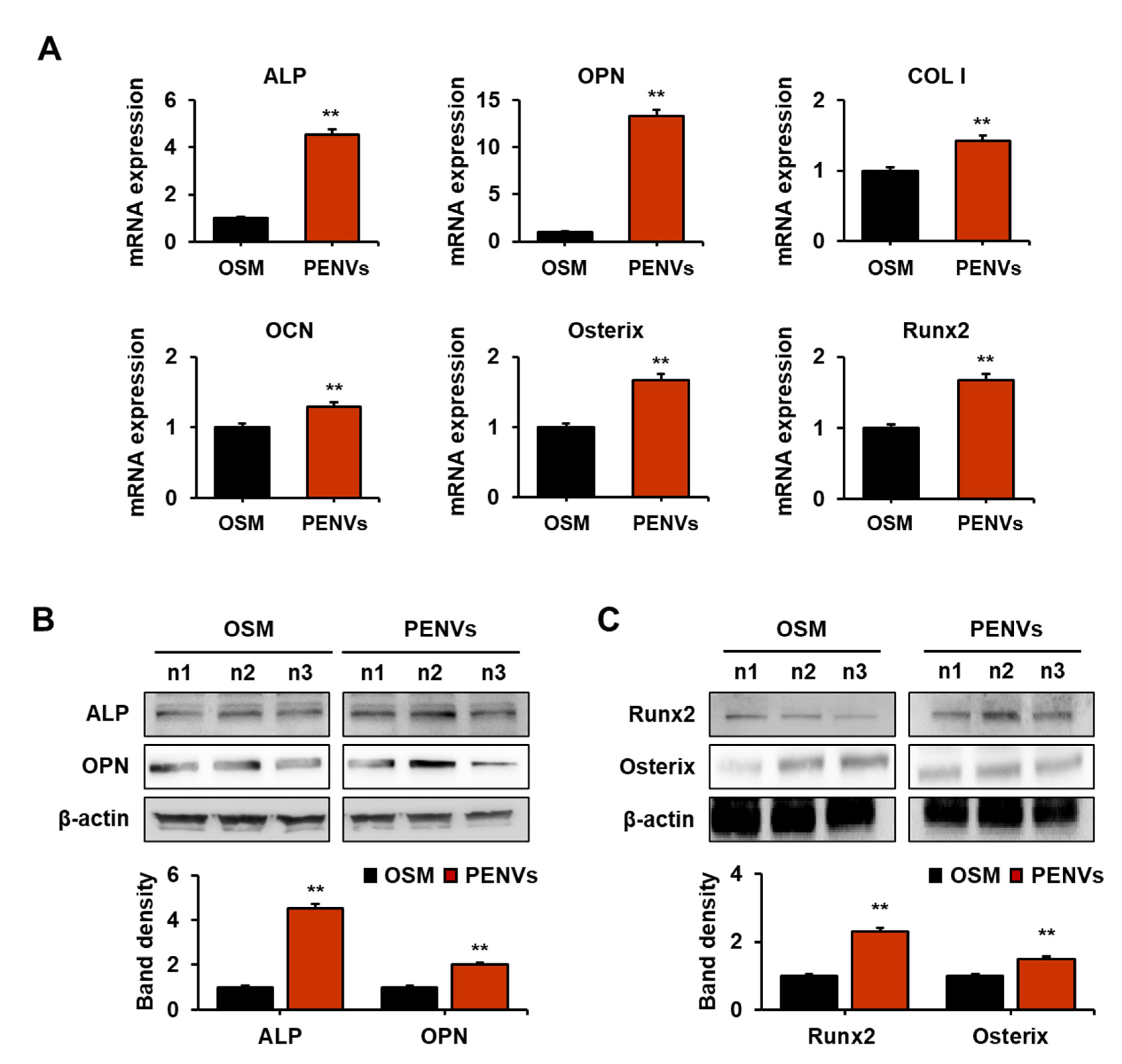
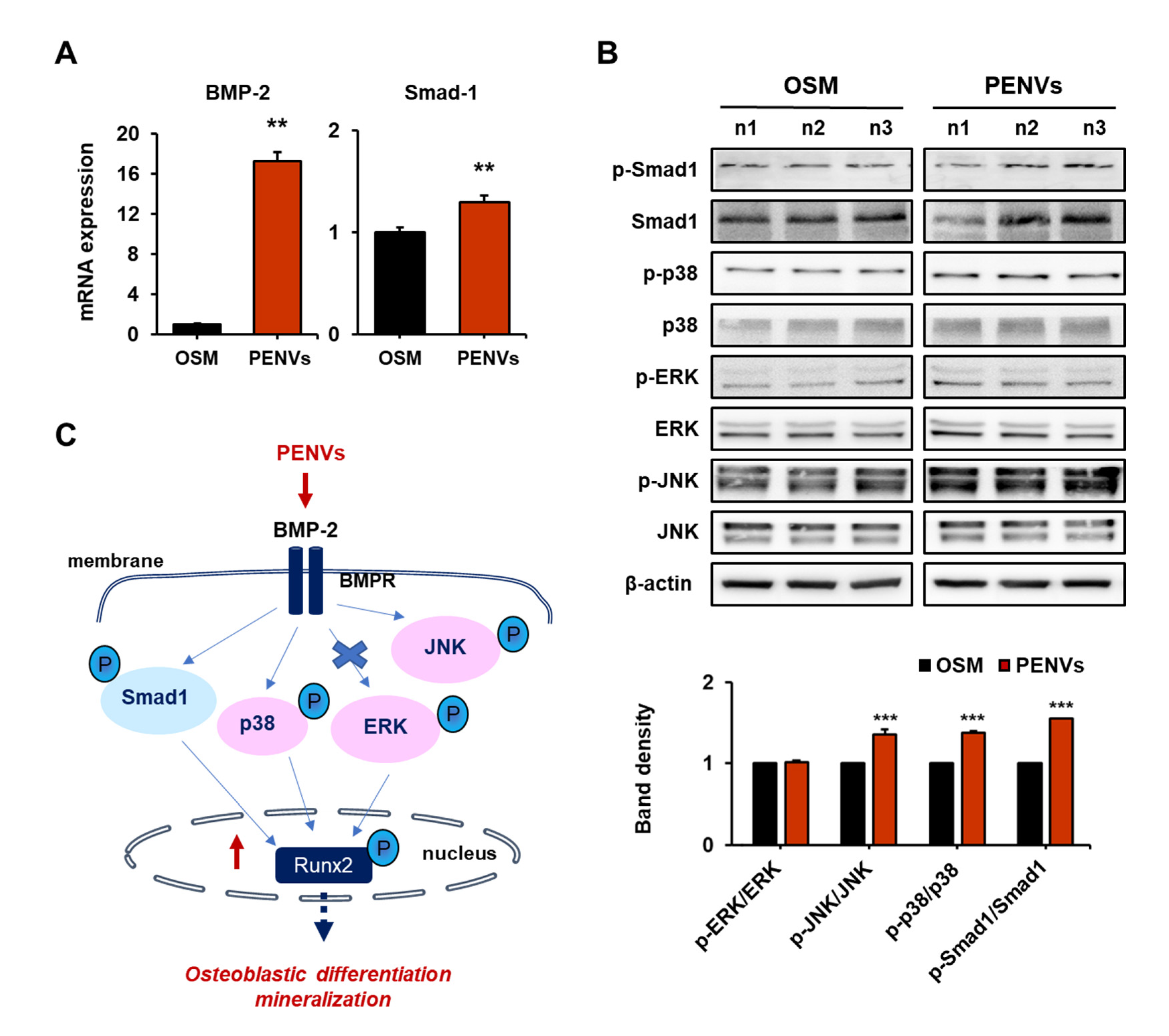
2.11. Real-Time Quantitative Polymerase Chain Reaction (RT qPCR)
2.12. Western Blot Analysis
2.13. Statistical Analysis
3. Results
3.1. Isolation and Characterization of PENVs

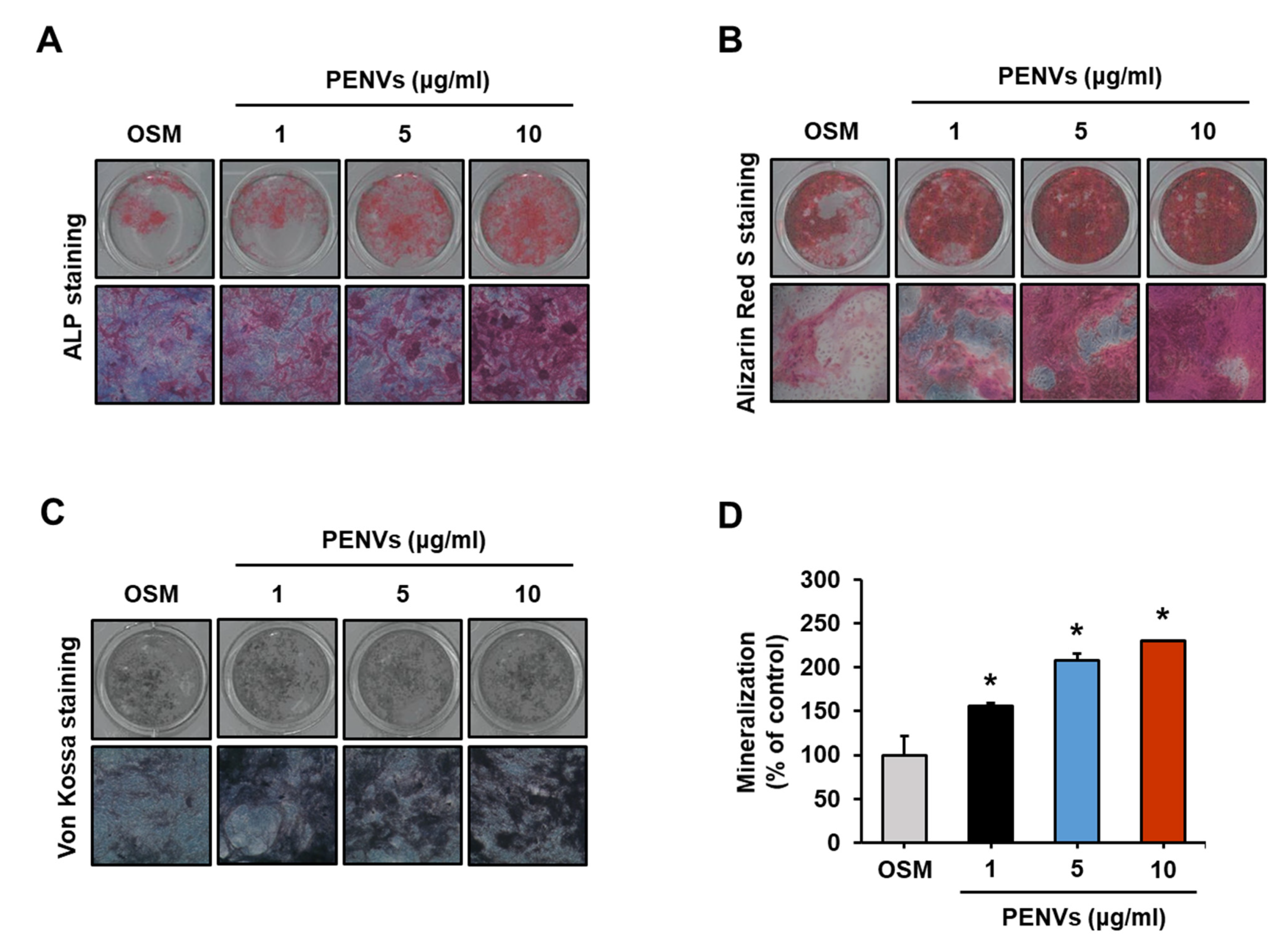
3.2. PENVs Activated the Osteoblast Differentiation and Mineralization
3.3. PENVs Enhanced Osteoblast Differentiation Marker Gene and Protein Expressions
3.4. PENVs Stimulated Osteoblast Differentiation through BMP-2/MAPK/Smad-1 Dependent Runx2 Pathway
3.5. PENVs Enhanced Osteoblast Differentiation and Mineralization of Mouse Primary Osteoblasts
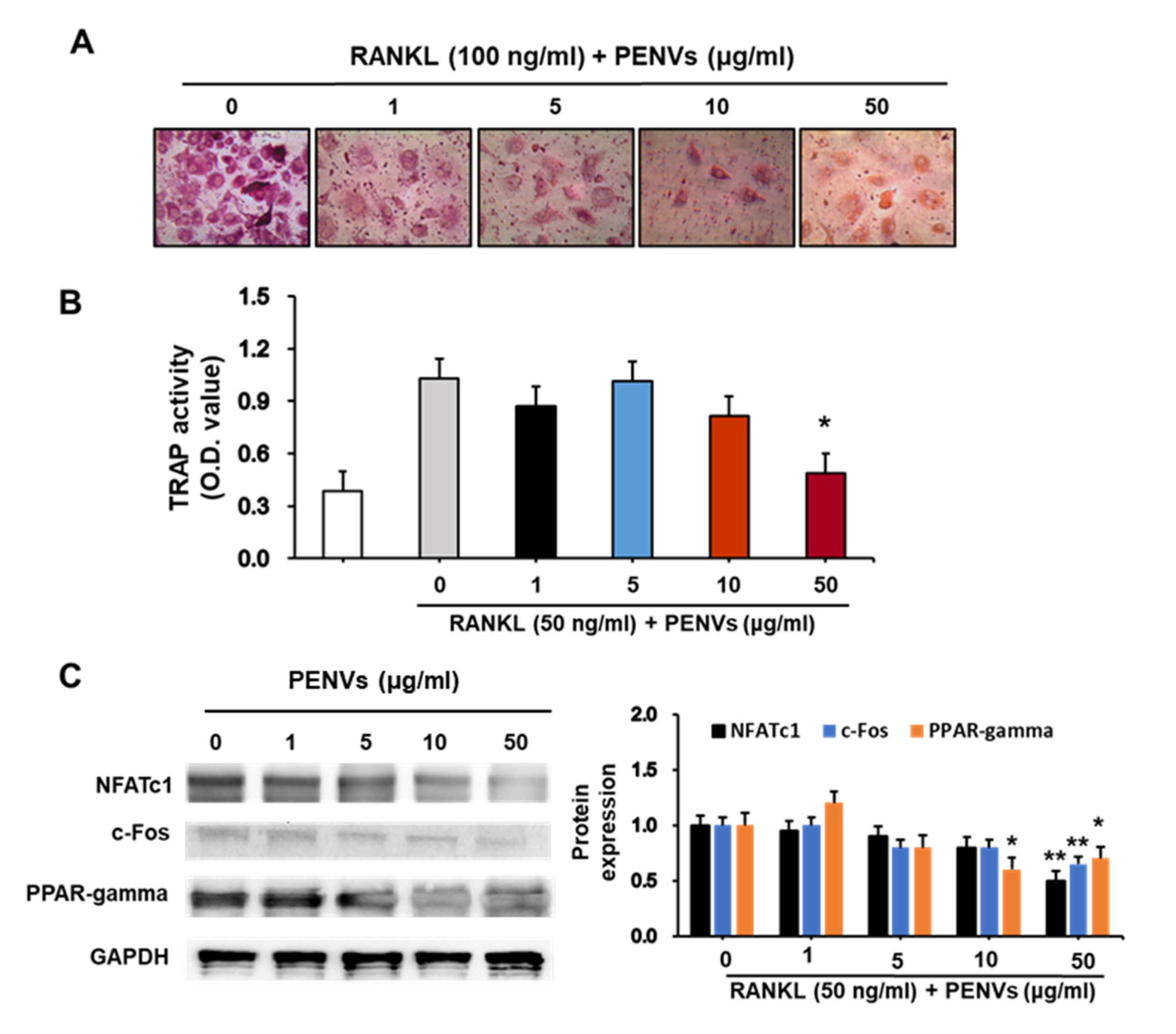
3.6. PENVs Inhibited Osteoclast Differentiation of Mouse Primary Osteoclasts
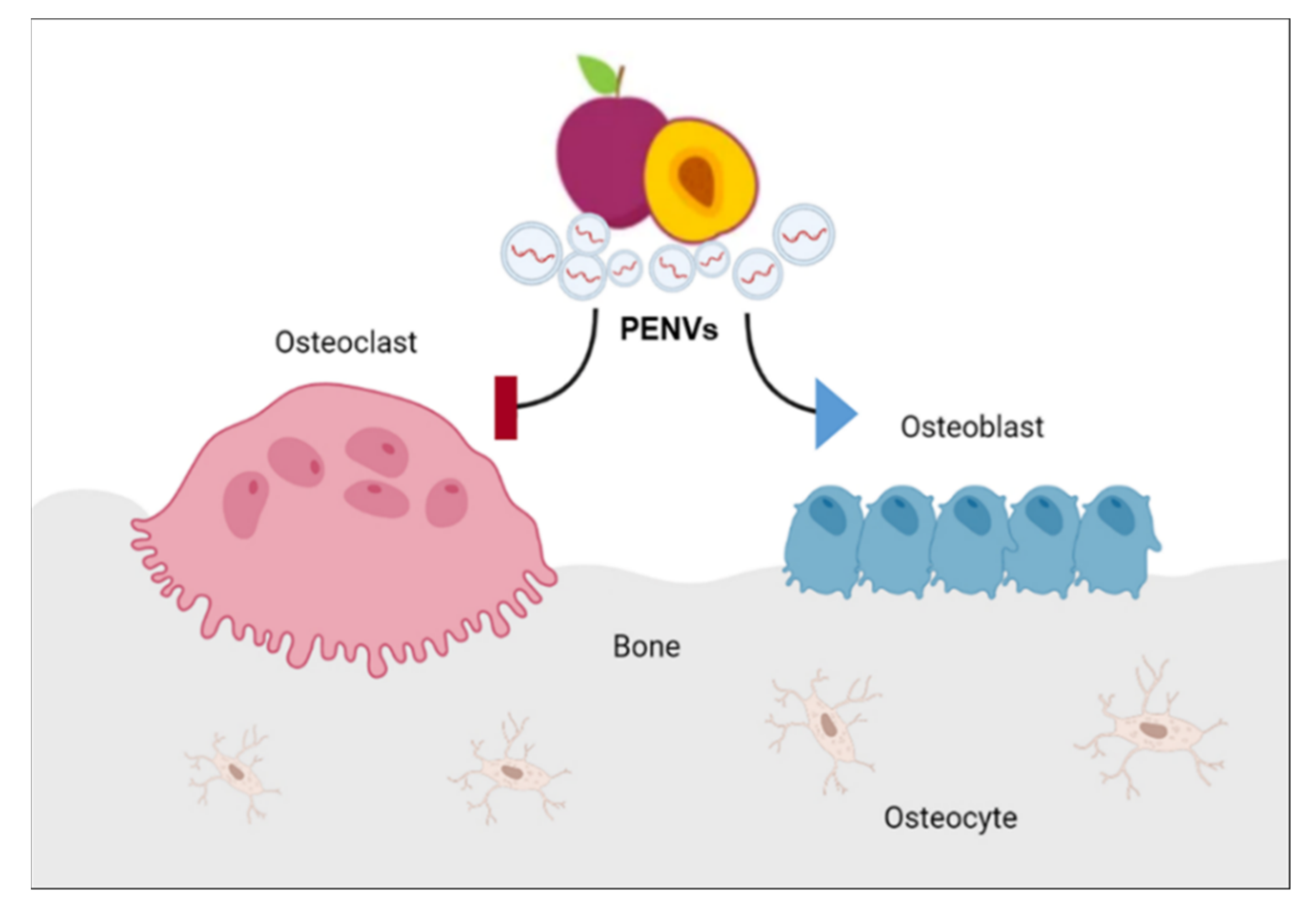
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; You, X.; Zhang, L.; Zhang, C.; Zou, W. Mechanical regulation of bone remodeling. Bone Res. 2022, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, M. Skeletal remodeling in health and disease. Nat. Med. 2007, 13, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Feng, X.; Teitelbaum, S.L. Osteoclasts: New Insights. Bone Res. 2013, 1, 11–26. [Google Scholar] [CrossRef]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef]
- Long, F. Building strong bones: Molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell Biol. 2011, 13, 27–38. [Google Scholar] [CrossRef]
- Harada, S.; Rodan, G.A. Control of osteoblast function and regulation of bone mass. Nature 2003, 423, 349–355. [Google Scholar] [CrossRef]
- Lin, X.; Patil, S.; Gao, Y.G.; Qian, A. The Bone Extracellular Matrix in Bone Formation and Regeneration. Front. Pharmacol. 2020, 11, 757. [Google Scholar] [CrossRef]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Tuan, R.S.; Robinson, L.J.; Schlesinger, P.H.; Nelson, D.J. Osteoblast Differentiation and Bone Matrix Formation In Vivo and In Vitro. Tissue Eng. Part B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef]
- Schlesinger, P.H.; Blair, H.C.; Beer Stolz, D.; Riazanski, V.; Ray, E.C.; Tourkova, I.L.; Nelson, D.J. Cellular and extracellular matrix of bone, with principles of synthesis and dependency of mineral deposition on cell membrane transport. Am. J. Physiol. Cell Physiol. 2020, 318, C111–C124. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yang, P.; Chu, X.; Huang, Y.; Liu, T.; Zhang, Q.; Li, Q.; Hu, L.; Waqas, Y.; Ahmed, N.; et al. Cellular evidence for nano-scale exosome secretion and interactions with spermatozoa in the epididymis of the Chinese soft-shelled turtle, Pelodiscus sinensis. Oncotarget 2016, 7, 19242–19250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Viennois, E.; Xu, C.; Merlin, D. Plant derived edible nanoparticles as a new therapeutic approach against diseases. Tissue Barriers 2016, 4, e1134415. [Google Scholar] [CrossRef]
- Bu, S.Y.; Hunt, T.S.; Smith, B.J. Dried plum polyphenols attenuate the detrimental effects of TNF-alpha on osteoblast function coincident with up-regulation of Runx2, Osterix and IGF-I. J. Nutr. Biochem. 2009, 20, 35–44. [Google Scholar] [CrossRef]
- Bu, S.Y.; Lerner, M.; Stoecker, B.J.; Boldrin, E.; Brackett, D.J.; Lucas, E.A.; Smith, B.J. Dried plum polyphenols inhibit osteoclastogenesis by downregulating NFATc1 and inflammatory mediators. Calcif. Tissue Int. 2008, 82, 475–488. [Google Scholar] [CrossRef]
- Liu, X.; Liu, M.; Turner, R.; Iwaniec, U.; Kim, H.; Halloran, B. Dried plum mitigates spinal cord injury-induced bone loss in mice. JOR Spine 2020, 3, e1113. [Google Scholar] [CrossRef] [PubMed]
- Graef, J.L.; Ouyang, P.; Wang, Y.; Rendina-Ruedy, E.; Lerner, M.R.; Marlow, D.; Lucas, E.A.; Smith, B.J. Dried Plum Polyphenolic Extract Combined with Vitamin K and Potassium Restores Trabecular and Cortical Bone in Osteopenic Model of Postmenopausal Bone Loss. J. Funct. Foods 2018, 42, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Eom, J.Y.; Choi, S.H.; Seo, H.J.; Kwun, I.S.; Chun, I.J.; Sung, J.; Lim, J.H.; Kim, J.; Song, B.J.; et al. Plum Prevents Intestinal and Hepatic Inflammation in the Acute and Chronic Models of Dextran Sulfate Sodium-Induced Mouse Colitis. Mol. Nutr. Food Res. 2022, 66, e2101049. [Google Scholar] [CrossRef]
- Hwang, J.H.; Park, Y.S.; Kim, H.S.; Kim, D.H.; Lee, S.H.; Lee, C.H.; Lee, S.H.; Kim, J.E.; Lee, S.; Kim, H.M.; et al. Yam-derived exosome-like nanovesicles stimulate osteoblast formation and prevent osteoporosis in mice. J. Control. Release 2023, 6, 184–198. [Google Scholar] [CrossRef]
- Ryu, J.K.; Kim, S.J.; Rah, S.H.; Kang, J.I.; Jung, H.E.; Lee, D.; Lee, H.K.; Lee, J.O.; Park, B.S.; Yoon, T.Y.; et al. Reconstruction of LPS Transfer Cascade Reveals Structural Determinants within LBP, CD14, and TLR4-MD2 for Efficient LPS Recognition and Transfer. Immunity 2017, 46, 38–50. [Google Scholar] [CrossRef]
- Whyte, M.P. Hypophosphatasia: Nature’s window on alkaline phosphatase function in humans. In Principles of Bone Biology, 4th ed.; Bilezikian, J.P., Martin, T.J., Clemens, T., Rosen, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1569–1599. [Google Scholar]
- Si, J.; Wang, C.; Zhang, D.; Wang, B.; Zhou, Y. Osteopontin in Bone Metabolism and Bone Diseases. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e919159. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Zhang, R.; Geoffroy, V.; Ridall, A.L.; Karsenty, G. Osf2/Cbfa1: A transcriptional activator of osteoblast differentiation. Cell 1997, 89, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, T.; Kawaguchi, H.; Jinno, S.; Hoshi, K.; Itaka, K.; Takato, T.; Nakamura, K.; Okayama, H. Bone morphogenetic protein 2-induced osteoblast differentiation requires Smad-mediated down-regulation of Cdk6. Mol. Cell. Biol. 2004, 24, 6560–6568. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Deng, C.; Li, Y.P. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Koniusz, S.; Andrzejewska, A.; Muraca, M.; Srivastava, A.K.; Janowski, M.; Lukomska, B. Extracellular Vesicles in Physiology, Pathology, and Therapy of the Immune and Central Nervous System, with Focus on Extracellular Vesicles Derived from Mesenchymal Stem Cells as Therapeutic Tools. Front. Cell. Neurosci. 2016, 10, 109. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Garaeva, L.; Kamyshinsky, R.; Kil, Y.; Varfolomeeva, E.; Verlov, N.; Komarova, E.; Garmay, Y.; Landa, S.; Burdakov, V.; Myasnikov, A.; et al. Delivery of functional exogenous proteins by plant-derived vesicles to human cells in vitro. Sci. Rep. 2021, 11, 6489. [Google Scholar] [CrossRef]
- Eom, J.Y.; Choi, S.H.; Kim, H.J.; Kim, D.H.; Bae, J.H.; Kwon, G.S.; Lee, D.H.; Hwang, J.H.; Kim, D.K.; Baek, M.C.; et al. Hemp-Derived Nanovesicles Protect Leaky Gut and Liver Injury in Dextran Sodium Sulfate-Induced Colitis. Int. J. Mol. Sci. 2022, 23, 9955. [Google Scholar] [CrossRef] [PubMed]
- Sim, Y.; Seo, H.J.; Kim, D.H.; Lee, S.H.; Kwon, J.; Kwun, I.S.; Jung, C.; Kim, J.I.; Lim, J.H.; Kim, D.K.; et al. The effect of apple-derived nanovesicles on the osteoblastogenesis of osteoblastic MC3T3-E1 Cells. J. Med. Food 2023, 26, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Rendina, E.; Lim, Y.F.; Marlow, D.; Wang, Y.; Clarke, S.L.; Kuvibidila, S.; Lucas, E.A.; Smith, B.J. Dietary supplementation with dried plum prevents ovariectomy-induced bone loss while modulating the immune response in C57BL/6J mice. J. Nutr. Biochem. 2012, 23, 60–68. [Google Scholar] [CrossRef]
- Arjmandi, B.H.; Johnson, S.A.; Pourafshar, S.; Navaei, N.; George, K.S.; Hooshmand, S.; Chai, S.C.; Akhavan, N.S. Bone-protective effects of dried plum in postmenopausal women: Efficacy and possible mechanisms. Nutrients 2017, 14, 496. [Google Scholar] [CrossRef]
- Aubin, J.E. Regulation of osteoblast formation and function. Rev. Endocr. Metab. Disord. 2001, 2, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J. Osteoblast dysfunctions in bone diseases: From cellular and molecular mechanisms to therapeutic strategies. Cell. Mol. Life Sci. CMLS 2015, 72, 1347–1361. [Google Scholar] [CrossRef]
- Yari, Z. Review of isoflavones and their potential cinical impacts on cardiovascular and bonemetabolism markers in peritoneal dialysis patients. Prev. Nutr. Food Sci. 2022, 27, 347–353. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence (5′ → 3′) |
|---|---|
| ALP | F: CAAGGATGCTGGGAAGTCCG R: CGGATAACGAGATGCCACCA |
| OPN | F: CTGGCAGCTCAGAGGAGAAG R: CAGCATTCTGTGGCGCAAG |
| COL 1 | F: ACGTCCTGGTGAAGTTGGTC R: CAGGGAAGCCTCTTTCTCCT |
| OCN | F: GCAATAAGGTAGTGAACAGACTC R: GTTTGTAGGCGGTCTTCAAGC |
| Osterix | F: GTCAAGAGTCTTAGCCAAACTC R: AAATGATGTGAGGCCAGATGG |
| BMP-2 | F: CGCACGCGATGCAACACCAC R: ACTGCATGTCCCCGGGCTCA |
| Smad-1 | F: AAGGTGGGGAAAGTGAAAC R: CTGCTTGGAACCAAATGGGAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.-S.; Kim, H.-W.; Hwang, J.-H.; Eom, J.-Y.; Kim, D.-H.; Park, J.; Tae, H.-J.; Lee, S.; Yoo, J.-G.; Kim, J.-I.; et al. Plum-Derived Exosome-like Nanovesicles Induce Differentiation of Osteoblasts and Reduction of Osteoclast Activation. Nutrients 2023, 15, 2107. https://doi.org/10.3390/nu15092107
Park Y-S, Kim H-W, Hwang J-H, Eom J-Y, Kim D-H, Park J, Tae H-J, Lee S, Yoo J-G, Kim J-I, et al. Plum-Derived Exosome-like Nanovesicles Induce Differentiation of Osteoblasts and Reduction of Osteoclast Activation. Nutrients. 2023; 15(9):2107. https://doi.org/10.3390/nu15092107
Chicago/Turabian StylePark, Yu-Seong, Hyun-Woo Kim, Jin-Hyeon Hwang, Jung-Young Eom, Dong-Ha Kim, Jinho Park, Hyun-Jin Tae, Seunghoon Lee, Jae-Gyu Yoo, Jee-In Kim, and et al. 2023. "Plum-Derived Exosome-like Nanovesicles Induce Differentiation of Osteoblasts and Reduction of Osteoclast Activation" Nutrients 15, no. 9: 2107. https://doi.org/10.3390/nu15092107
APA StylePark, Y.-S., Kim, H.-W., Hwang, J.-H., Eom, J.-Y., Kim, D.-H., Park, J., Tae, H.-J., Lee, S., Yoo, J.-G., Kim, J.-I., Lim, J.-H., Kwun, I.-S., Baek, M.-C., Cho, Y.-E., & Kim, D.-K. (2023). Plum-Derived Exosome-like Nanovesicles Induce Differentiation of Osteoblasts and Reduction of Osteoclast Activation. Nutrients, 15(9), 2107. https://doi.org/10.3390/nu15092107