
Effects of Varying Ratios of Glycyrrhiza uralensis and Donkey Hide Gelatin Water Extracts on Dinitrochlorobenzene-Induced Atopic Dermatitis in NC/Nga Mice

 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Animal
2.2. Atopic Dermatitis Induced by DNCB
2.3. Sample Preparation
2.4. Intervention
2.5. Evaluation of Atopic Dermatitis Skin Symptoms
2.6. Analysis of Dorsal Skin Tissue, Axillary Lymph Node (ALN), and Peripheral Blood Mononuclear Cells (PBMCs) Using Fluorescence-Activated Cell Sorting
2.7. Immunoglobulin E (IgE) Level Measurement
2.8. Detection of Th1/Th2 Cytokine Expression
2.9. Quantitative Polymerase Chain Reaction (qPCR) in Dorsal Skin Tissue
2.10. Histology Analysis
2.11. Immunofluorescence to Measure Claudin 1 and Sirtuin 1 (Sirt 1) Expression in the Epidermis
2.12. Cell Culture and MTT Assay
2.13. Measurement of Nitric Oxide and Cytokines in RAW264.7 Cells
2.14. Measurement of Chemokines and Cytokines in HaCaT Cells
2.15. NF-κB Binding Activity
2.16. Isolation of Nuclear and Cytoplasmic Fractions
2.17. Western Blot Analysis
2.18. UHPLC Analysis
2.19. Statistical Analysis
3. Results
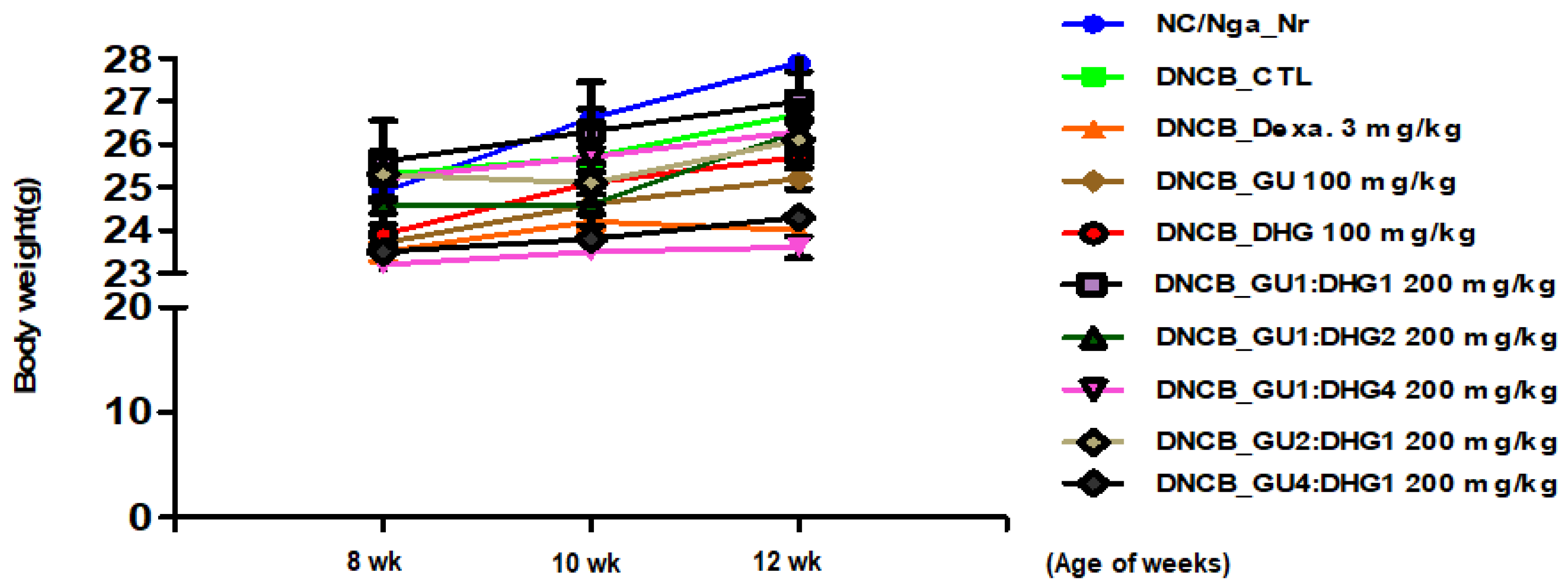
3.1. Changes in Body Weight
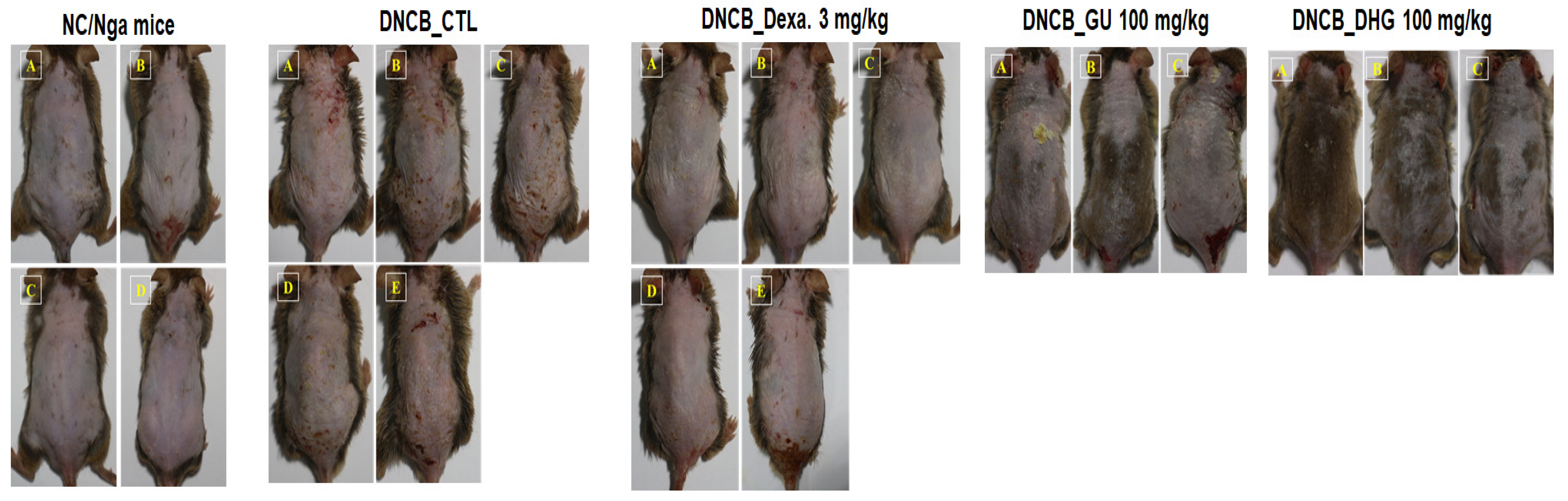
3.2. Changes in Skin Symptom Evaluation Score
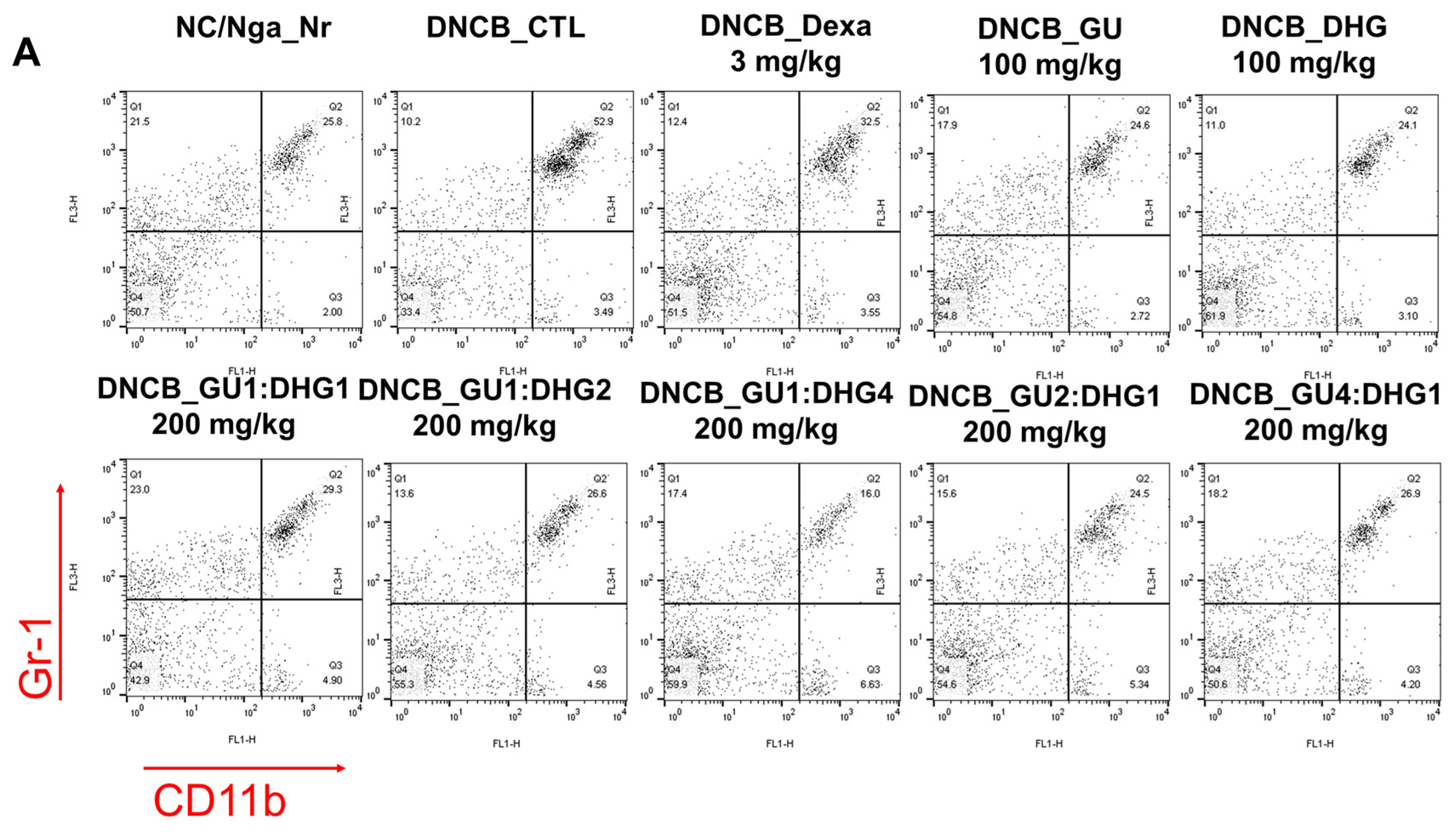
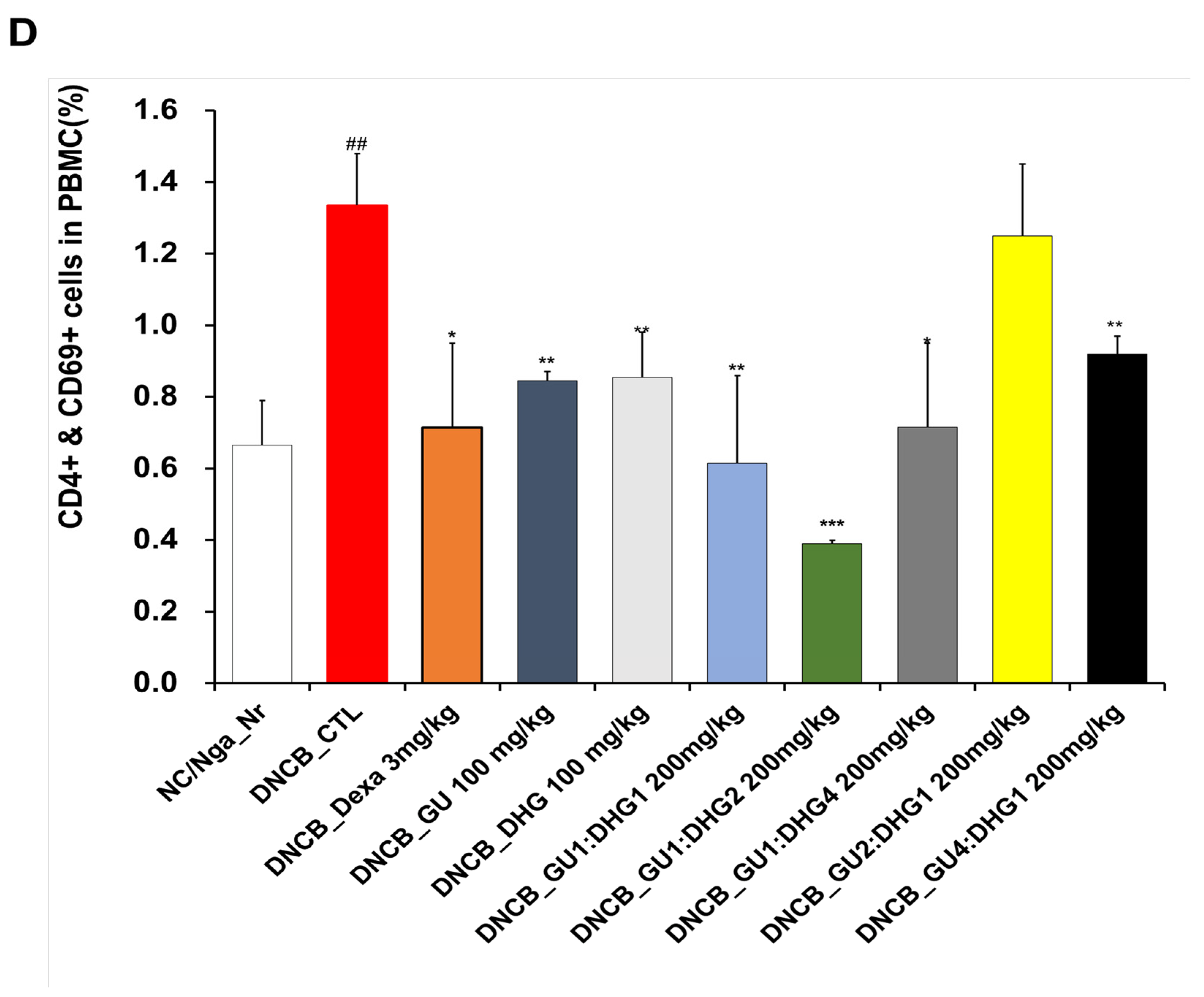
3.3. Changes in CD4+/CD69+ and Gr-1+/CD11b+ Cell Frequency through PBMC Immunofluorescence Staining
3.4. Changes in Absolute Cell Number of Dorsal Skin Tissue and ALN
3.5. Immunoglobulin E Level in Serum
3.6. IL-4, IL-5, IL-13, and IFN-γ Protein Production in the Culture Solution of Splenocytes
3.7. mRNA Expression
3.8. Histology Analysis of Dorsal Skin Tissue
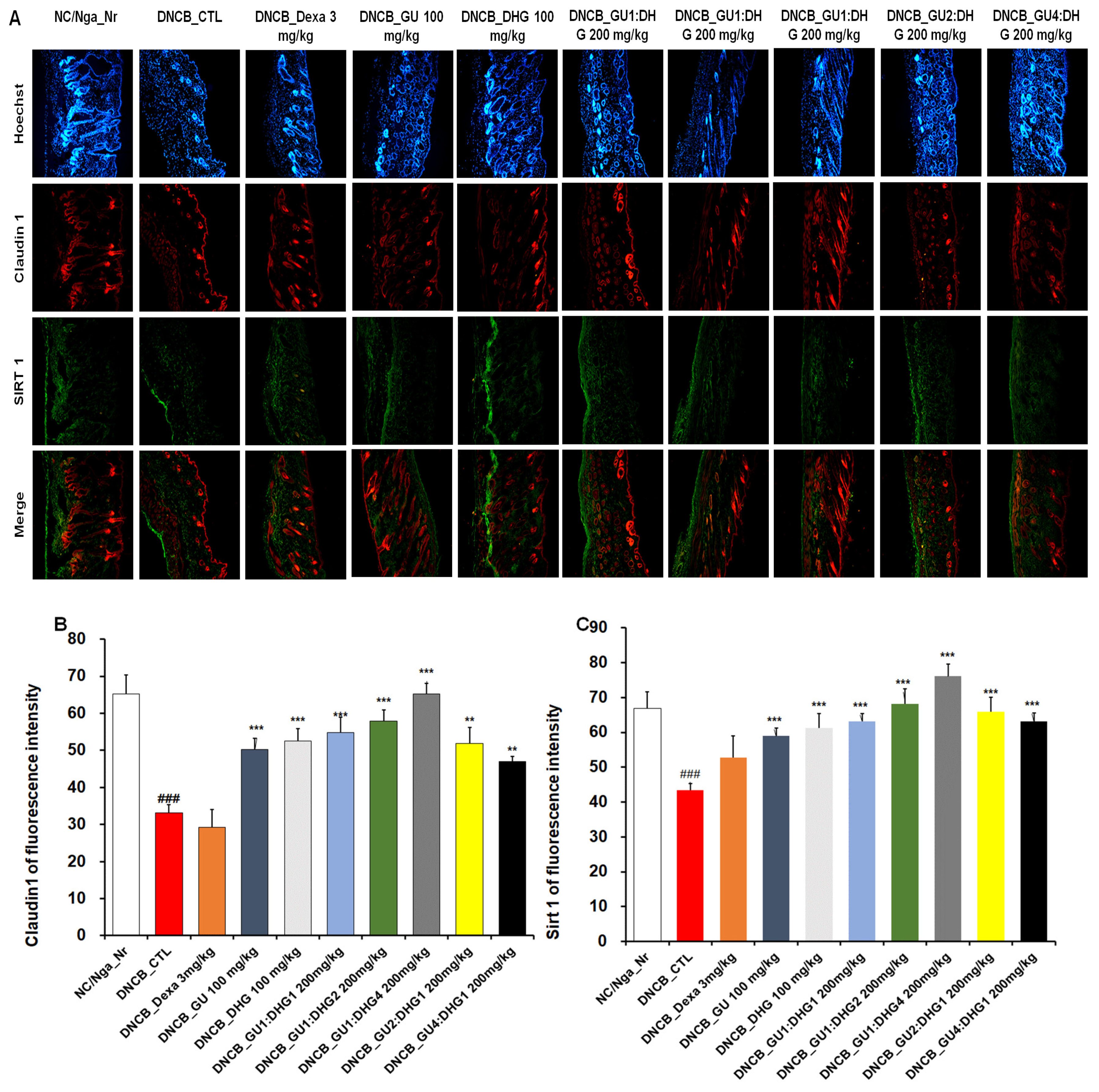
3.9. Immunofluorescence Staining to Evaluate Claudin 1 and Sirt 1 Expression in Dorsal Skin Tissue
3.10. Effects of GU, DHG, and GU + DHG Mixtures on the Viability of RAW264.7 Macrophages and HaCaT Keratinocytes
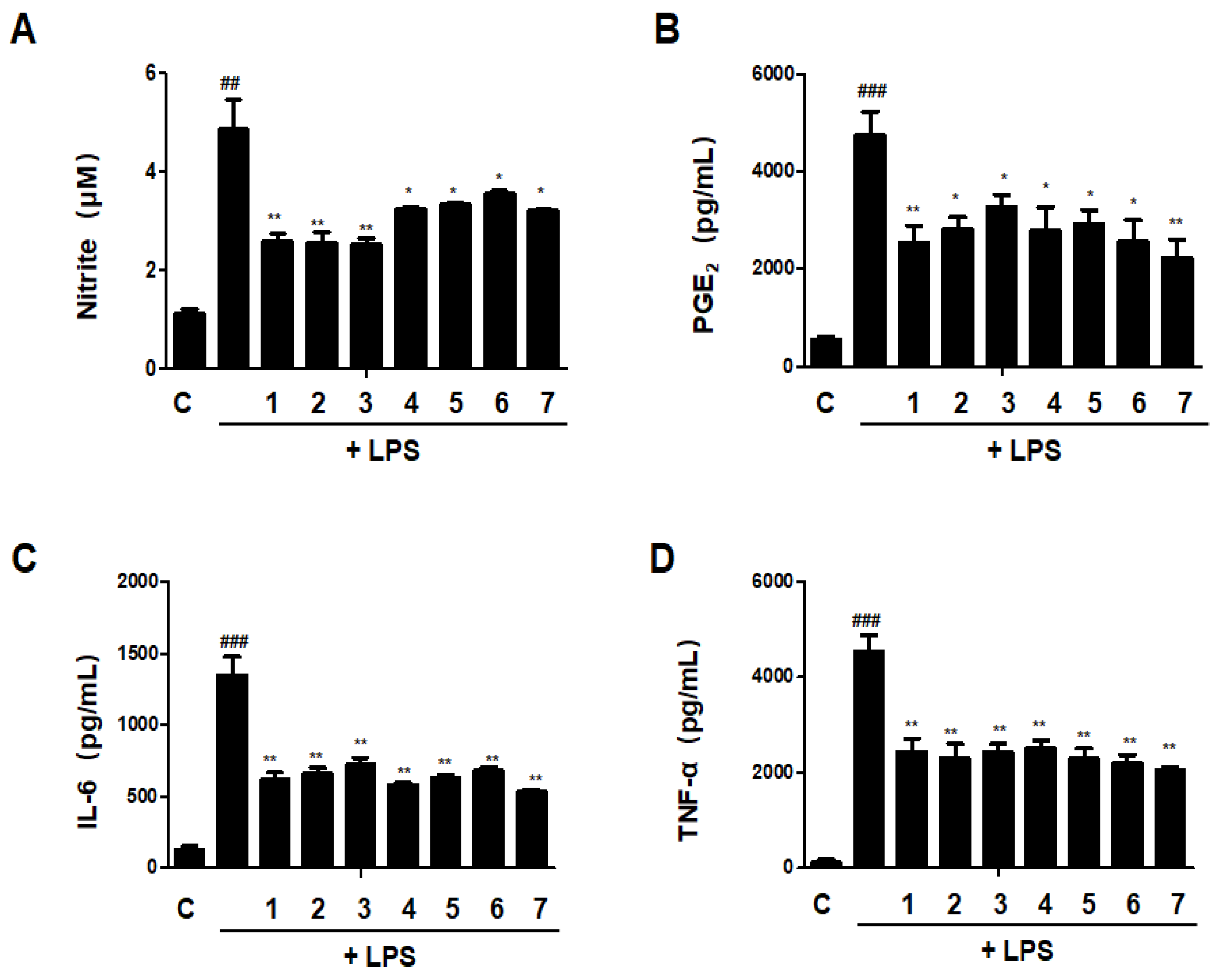
3.11. Effects of GU, DHG and GU + DHG Mixture on Inflammatory Mediators Nitrite, PGE2, TNF-α, and IL-6 in LPS-Induced RAW264.7 Macrophages
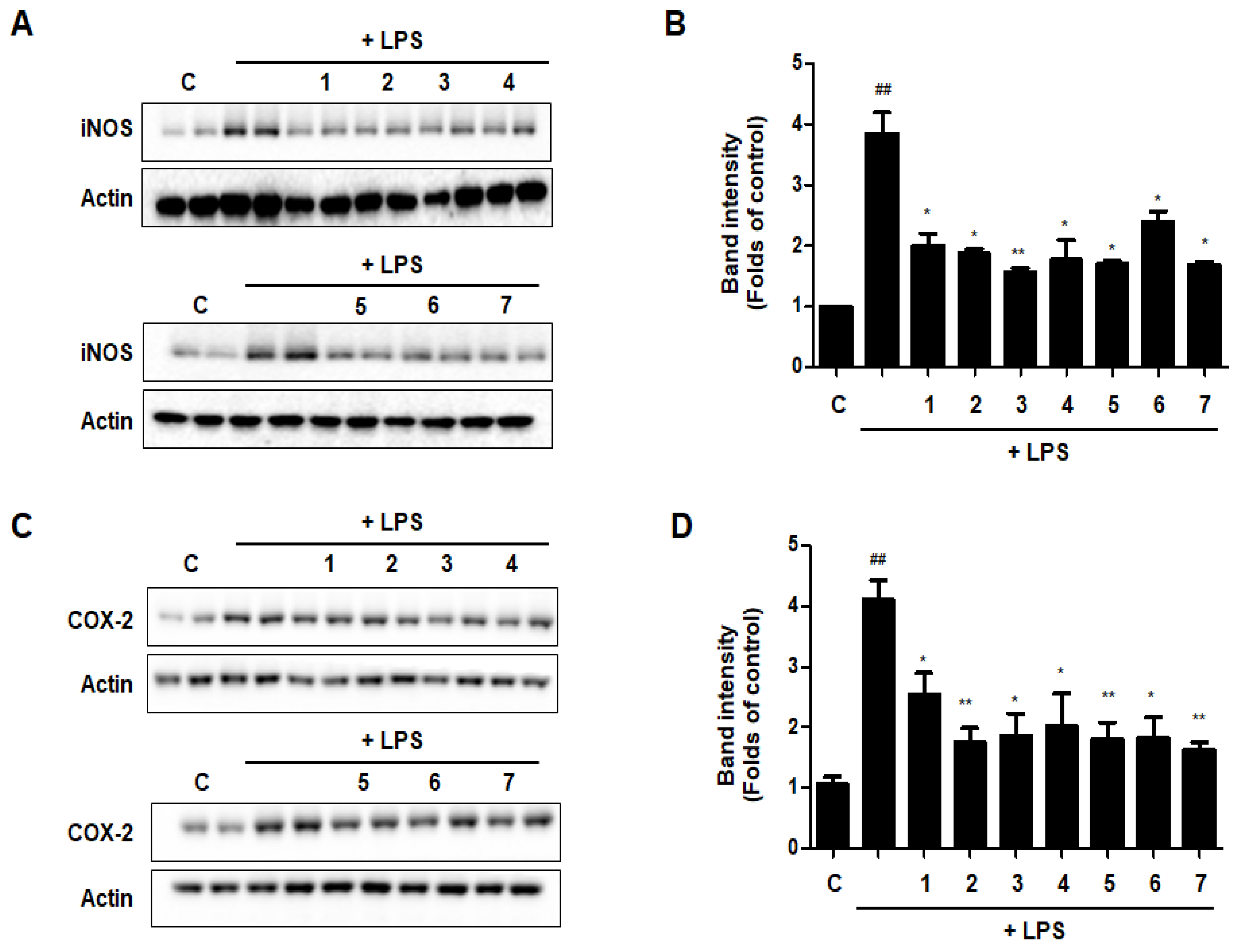
3.12. Effects of GU, DHG, and GU + DHG Mixtures on Protein Expression Levels of Nitric Oxide Synthase (iNOS) and Cyclooxygenase-2 (COX-2) in LPS-Induced RAW264.7 Macrophages
3.13. Effects of GU, DHG, and GU + DHG Mixtures on TNF-α/IFN-γ-Induced Pro-Inflammatory Cytokines and Chemokines in HaCaT Cells
3.14. Effects of GU, DHG, and GU + DHG Mixtures on TNF-α/IFN-γ-Induced Expression of ICAM-1 and COX-2 in HaCaT Cells
3.15. Effects of GU, DHG, and GU + DHG Mixtures on NF-κB Binding Activity in LPS-Induced RAW264.7 Cells/TNF-α/IFN-γ-Induced HaCaT Cells
3.16. UHPLC-TOF-HRMS Analysis of the GU1:DHG4 Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wollenberg, A.; Barbarot, S.; Bieber, T.; Christen-Zaech, S.; Deleuran, M.; Fink-Wagner, A.; Gieler, U.; Girolomoni, G.; Lau, S.; Muraro, A.; et al. Consensus-based European guidelines for treatment of atopic eczema (atopic dermatitis) in adults and children. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 657–682. [Google Scholar] [CrossRef]
- Nutten, S. Atopic dermatitis: Global epidemiology and risk factors. Ann. Nutr. Metab. 2015, 66, 8–16. [Google Scholar] [CrossRef]
- Zhang, B.; Luo, P.; Sun, J.; Li, D.; Liu, Z.; Liu, X.; Zhao, Z.; Xie, X.; Yang, J.; Shen, C. The Epidermal Barrier Structure and Function of Re-Harvested Skin from Non-Scalp Donor Sites. J. Investig. Surg. 2023, 36, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, N.; Brandner, J.M. Barriers and more: Functions of tight junction proteins in the skin. Ann. N. Y. Acad. Sci. 2012, 1257, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.L.; Balkrishnan, R.; Feldman, S.R.; Fleischer, A.B., Jr.; Manuel, J.C. The burden of atopic dermatitis: Impact on the patient, family, and society. Pediatr. Dermatol. 2005, 22, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Pescitelli, L.; Rosi, E.; Ricceri, F.; Pimpinelli, N.; Prignano, F. Novel Therapeutic Approaches and Targets for the Treatment of Atopic Dermatitis. Curr. Pharm. Biotechnol. 2021, 22, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Kasraie, S.T.; Werfel, T. Role of Macrophages in the Pathogenesis of Atopic Dermatitis. Mediat. Inflamm. 2013, 2013, 942375. [Google Scholar] [CrossRef]
- Abramovits, W. Atopic dermatitis. J. Am. Acad. 2005, 53, S86–S93. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Ulzii, D.; Vu, Y.H.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T. Pathogenesis of atopic dermatitis: Current paradigm. Iran. J. Immunol. 2019, 16, 97–107. [Google Scholar]
- Dipiro, J.T. Concepts in Clinical Pharmacokinetics, 5th ed.; ASHP: Bethesda, MD, USA, 2010. [Google Scholar]
- Reigner, D.; Williams, P.; Patel, I.H.; Steimer, J.L.; Peck, C.; van Brummelen, P. An Evaluation of the Integration of Pharmacokinetic and Pharmacodynamic Principles in Clinical Drug Development. Clin. Pharm. 1997, 33, 142–152. [Google Scholar] [CrossRef]
- Kim, H.U.; Ryu, J.Y.; Lee, J.O.; Lee, S.Y. A systems approach to traditional oriental medicine. Nat. Biotechnol. 2015, 33, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Guan, E.; Zhang, Y.; Shu, Z.; Wang, B.; Wu, X.; Chen, J.; Liu, J.; Fu, X.; Sun, W.; et al. Chemical profile and anti-inflammatory activity of total flavonoids from Glycyrrhiza uralensis Fisc . Iran. J. Pharm. Res. 2018, 17, 726–734. [Google Scholar] [PubMed]
- Chen, J.; Gu, X.L.; Chen, J.G.; Luo, Y.; Wang, M.Y.; Yang, H.Y.; Guo, X.; Zhu, X.Q. Immunomodulatory effects of Glycyrrhiza uralensis polysaccharide in glycinin-induced allergic mouse model. Food Agric. Immunol. 2016, 28, 179–188. [Google Scholar] [CrossRef]
- Wang, D.; Ru, W.; Xu, Y.; Zhang, J.; He, X.; Fan, G.; Mao, B.; Zhou, X.; Qin, Y. Chemical constituents and bioactivities of Colla corii asini . Drug Discov. Ther. 2014, 8, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.M.; Mao, G.N.; Kang, R.R. Effect of Colla corri asini effervescent granules on immune function in mice. Progr. Vet. Med. 2011, 32, 73–75. [Google Scholar]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef]
- Suto, H.; Matsuda, H.; Mitsuishi, K.; Hira, K.; Uchida, T.; Unno, T.; Ra, C. NC/Nga mice: A mouse model for atopic dermatitis. Int. Arch Allerg. Immunol. 1999, 120 (Suppl. 1), 70–75. [Google Scholar] [CrossRef]
- Lee, H.; Liu, Z.M.; Dong, L.S.; Cheong, S.H.; Lee, D.S. Lycopus maackianus Makino MeOH Extract Exhibits Antioxidant and Anti-Neuroinflammatory Effects in Neuronal Cells and Zebrafish Model. Antioxidants 2022, 11, 690. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Herrera-Bravo, J.; Belén, L.H.; Kaur, R.; Kregiel, D.; Suleria, H.A.R. Glycyrrhiza genus: Enlightening phytochemical components for pharmacological and health-promoting abilities. Oxid. Med. Cell Longev. 2021, 2021, 1–20. [Google Scholar] [CrossRef]
- Oyoshi, M.K.; He, R.; Kumar, L.; Yoon, J.; Geha, R.S. Cellular and molecular mechanisms in atopic dermatitis. Adv. Immunol. 2009, 102, 135–226. [Google Scholar]
- Mosmann, T.R.; Sad, S. The expanding universe of T-cell subsets: Th1, Th2 and more. Immunol. Today 1996, 17, 138–146. [Google Scholar] [CrossRef]
- Grewe, M.; Bruijnzeel-Koomen, C.; Schpf, E.; Thepen, T.; Langeveld-Wildschut, A.G.; Ruzicka, T.; Krutmann, J. A role for Th1 and Th2 cells in the immunopathogenesis of atopic dermatitis. Immunol. Today 1998, 19, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Benedetto, A.D.; Rafaels, N.M.; Mcgirt, L.Y.; Ivanov, A.I.; Georas, S.N.; Cheadle, C.; Berger, A.E.; Zhang, K.; Vidyasagar, S.; Yoshida, T.; et al. Tight junction defects in patients with atopic dermatitis. J. Allerg. Clin. Immunol. 2010, 127, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Park, K.I.; Lee, M.R.; Oh, T.W.; Kim, K.Y.; Ma, J.Y. Antibacterial activity and effects of Colla corii asini on Salmonella typhimurium invasion in vitro and in vivo. BMC Complement. Altern. Med. 2017, 17, 520. [Google Scholar]
- Zhang, S.; Xu, L.; Liu, Y.X.; Fu, H.Y.; Xiao, Z.B.; She, Y.B. Characterization of aroma-active components and antioxidant activity analysis of E-jiao (Colla Corii Asini) from different geographical origins Nat . Prod. Bioprospect. 2018, 8, 71–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, M.; Cao, J.; Cheng, Y.; Zhuo, C.; Xu, H.; Tian, S.; Zhang, Y.; Zhang, J.; Wang, F. 2012. Effect of Colla corii asini (E’iao) on D-galactose induced aging mice. Biol. Pharm. Bull. 2012, 35, 2128–2132. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, X.; Liu, H.; Xiang, H.; Xing, J.; Zhanga, L.; Qin, X. The hematinic effect of Colla corii asini (Ejiao) using 1H-NMR metabolomics coupled with correlation analysis in APH-induced anemic rats. RSC Adv. 2017, 7, 8952–8962. [Google Scholar] [CrossRef]
- Wang, W.; Hu, X.; Zhao, Z.; Liu, P.; Hu, Y.; Zhou, J.; Zhou, D.; Wang, Z.; Guo, D.; Guo, H. Antidepressant-like effects of liquiritin and isoliquiritin from Glycyrrhiza uralensis in the forced swimming test and tail suspension test in mice. Prog. Neuropsychopharmacol. Biol. Psychiatr. 2008, 32, 1179–1184. [Google Scholar] [CrossRef]
- Koan, S.W. Antioxidative Activity of Heat Treated Licorice (Glycyrrhiza uralensis Fisch) Extracts. Korean J. Food Sci. Technol. 2006, 38, 355–360. [Google Scholar]
- Adianti, M.; Aoki, C.; Komoto, M.; Deng, L.; Shoji, I.; Wahyuni, T.S.; Lusida, M.I.; Soetjipto, F.H.; Kawahara, N.; Hotta, H. Anti-hepatitis C virus compounds obtained from Glycyrrhiza uralensis and other Glycyrrhizaspecies. Microbiol. Immunol. 2014, 58, 180–187. [Google Scholar] [CrossRef]
- Kao, T.C.; Shyu, M.H.; Yen, G.C. Glycyrrhizic acid and 18beta-glycyrrhetinic acid inhibit inflammation via PI3K/Akt/GSK3beta signaling and glucocorticoid receptor activation. J. Agric. Food Chem. 2010, 15, 8623–8629. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Park, S.H.; Yoon, H.K.; Han, M.J.; Kim, D.H. Anti-allergic activity of 18beta-glycyrrhetinic acid-3-O-beta-D-glucuronide. Arch Pharm. Res. 2004, 27, 57. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Zhang, T.T.; Song, M.Y. Effects of Colla corii asini on lung function and pathological injury in rats with chronic obstructive pulmonary disease. Basic Clin. Med. 2021, 17, 970. [Google Scholar]
- Kang, Y.M.; Kim, H.M.; Lee, J.S. Colla Corii Asini suppresses the atopic dermatitis-like skin lesions in NC/Nga mice and HaCaT keratinocytes. Pharmacological. Res. Mod. Chin. Med. 2022, 2, 100039. [Google Scholar] [CrossRef]
- Cheng, F.; Zhou, Y.; Wang, M.; Guo, C.; Cao, Z.; Zhang, R.; Peng, C. A review of pharmacological and pharmacokinetic properties of stachydrine. Pharmacol. Res. 2020, 155, 104755. [Google Scholar] [CrossRef]
- Noh, H.M.; Park, S.G.; Kim, W.; Jo, E.H.; Ki, H.H.; Kim, D.K.; Park, M.C. Anti-allergic effects of Jagamcho-tang in ovalbumin-induced atopic dermatitis mouse model. J. Physiol. Pathol. Korean Med. 2017, 31, 277–283. [Google Scholar] [CrossRef]
- Lin, C.Y.; Lin, Y.C.; Paul, C.R.; Hsieh, D.J.Y.; Day, C.H.; Chen, R.J.; Huang, C.Y. Isoliquiritigenin ameliorates advanced glycation end-products toxicity on renal proximal tubular epithelial cells. Environ. Toxicol. 2022, 37, 2096–2102. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, T.; Lu, Z.; He, X.; Quan, J.; Liu, S.; Yu, L. Liquiritin exhibits anti-acute lung injury activities through suppressing the JNK/Nur77/c-Jun pathway. Chin. Med. 2023, 18, 35. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Concentration |
|---|---|
| NC/Nga_Nr | |
| DNCB_CTL | |
| DNCB_Dexa | 3 mg/kg |
| DNCB_GU | 100 mg/kg |
| DNCB_DHG | 100 mg/kg |
| DNCB_GU1:DHG1 | 200 mg/kg |
| DNCB_GU1:DHG2 | 200 mg/kg |
| DNCB_GU1:DHG4 | 200 mg/kg |
| DNCB_GU2:DHG1 | 200 mg/kg |
| DNCB_GU4:DHG1 | 200 mg/kg |
| Gene | Primer | Sequence |
|---|---|---|
| GADPH | VIC-probe | 5′-CATCCTGCACCACCAACTGCTTAGCC-3′ |
| Il-31R | Forward | 5′-ATGCCCAACAAAGCAGAGAC-3′ |
| Reverse | 5′-TGAGAGAACCAGGGAGCTGT-3′ | |
| IL-13 | Forward | 5′-ATGCCCAACAAAGCAGAGAC-3′ |
| Reverse | 5′-TGAGAGAACCAGGGAGCTGT-3′ | |
| TNF-α | Forward | 5′-TGGGAGGCACTTGCATTGA-3′ |
| Forward | 5′-GGCTTTCCGAATTCACTGGAGCCCT-3′ | |
| Reverse | 5′-CCCCGGCCTTCCAAATAAATACATTCATTCATA-3′ |
| Group Number | Treatment | Concentration |
|---|---|---|
| control | ||
| TNF-α/IFN-γ or LPS | ||
| 1 | DHG | 200 μg/mL |
| 2 | GU1:DHG4 | 100 μg/mL |
| 3 | GU1:DHG2 | 100 μg/mL |
| 4 | GU1:DHG1 | 100 μg/mL |
| 5 | GU2:DHG1 | 100 μg/mL |
| 6 | GU4:DHG1 | 100 μg/mL |
| 7 | GU | 100 μg/mL |
| Cell PHENOTYPES in ALN & D-Skin | DNCB-Induced Atopic Dermatitis Murine Model (Absolute no.) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| NC/Nga-Nr | DNCB_CTL | DNCB_Dexa. 3 mg/kg | DNCB_GU 100 mg/kg | DNCB_DHG 100 mg/kg | DNCB_GU1:DHG1 200 mg/kg | DNCB_GU1:DHG2 200 mg/kg | DNCB_GU1:DHG4 200 mg/kg | DNCB_GU2:DHG1 200 mg/kg | DNCB_GU4:DHG1 200 mg/kg | ||
| CD19+ (×105 cells) | ALN | 9.18 ± 6.45 | 89.78 ± 21.15 ## | 26.19 ± 7.49 ** | 61.95 ± 10.95 | 69.98 ± 5.86 | 39.49 ± 3.97 * | 48.16 ± 1.79 | 49.12 ± 15.46 | 78.95 ± 19.06 | 77.96 ± 34.29 |
| CD4+ (×105 cells) | 18.46 ± 13.35 | 103.37 ± 23.91 ## | 50.08 ± 15.99 * | 72.57 ± 7.63 | 104.71 ± 3.59 | 54.84 ± 2.41 * | 46.91 ± 13.87 * | 66.37 ± 11.46 | 78.91 ± 14.89 | 79.60 ± 17.91 | |
| CD8+ (×105 cells) | 11.6 ± 9.11 | 105.40± 19.49 ### | 52.94 ± 21.50 * | 58.44 ± 4.85 * | 95.49 ± 4.85 | 62.00 ± 4.76 * | 47.07 ± 9.95 ** | 70.01 ± 13.06 | 84.78 ± 12.52 | 86.75 ± 19.60 | |
| CD4+/CD69+ (×105 cells) | 2.06 ± 1.50 | 9.20 ± 0.25 ### | 4.33 ± 1.44 ** | 7.97 ± 2.24 | 13.51 ± 2.45 | 7.89 ± 1.21 | 5.40 ± 2.33 * | 8.40 ± 1.26 | 9.50 ± 0.41 | 11.12 ± 2.46 | |
| CD23+/B220+ (×105 cells) | 10.67 ± 8.12 | 101.05 ± 35.01 ## | 23.19 ± 7.21 *** | 52.34 ± 8.41 * | 79.71 ± 5.61 | 56.74 ± 4.53 * | 56.54 ± 4.45 * | 67.40 ± 11.81 | 79.08 ± 15.07 | 76.33 ± 30.40 | |
| CD4+ (×105 cells) | D-skin | 1.02 ± 0.15 | 14.32 ± 4.56 ## | 5.16 ± 1.30 * | 6.60 ± 0.23 | 9.70 ± 0.20 | 7.01 ± 2.23 | 4.39 ± 0.58 * | 2.89 ± 1.13 ** | 5.96 ± 0.98 | 5.25 ± 0.74 |
| CD8+ (×105 cells) | 0.55 ± 0.04 | 5.36 ± 0.12 ### | 0.57 ± 0.25 *** | 1.71 ± 0.10 *** | 2.21 ± 0.16 *** | 1.68 ± 0.62 *** | 1.16 ± 0.61 *** | 0.58 ± 0.08 *** | 2.09 ± 0.61 *** | 1.80 ± 0.38 *** | |
| Gr-1+/CD11b+ (×105 cells) | 0.37 ± 0.13 | 3.27 ± 0.73 ## | 0.96 ± 0.03 ** | 1.43 ± 0.01 ** | 1.66 ± 0.30 * | 1.65 ± 0.73 | 1.14 ± 0.01 ** | 0.52 ± 0.01 ** | 1.47 ± 1.00 | 1.68 ± 0.87 | |
| Compound | M | RT (Min) | m/z Traces (+) | m/z Traces (–) | MS2 |
|---|---|---|---|---|---|
| Sucrose | 342.1 | 1.30 | 343.1 | – | MS2 (+) [343.1]: 69.4, 85.4, 96.9, 127.6, 145.8, 163.2 |
| Stachydrine | 143.1 | 1.51 | 144.1 | – | MS2 (+) [144.1]: 84.1, 144.4 |
| Hydroxyferulic acid | 210.1 | 8.35 | – | 209.1 | MS2 (–) [209.1]: 59.3, 93.2, 121.1, 165.2 |
| Isoliquiritigenin | 256.1 | 19.27 | 257.1 | – | MS2 (+) [257.1]: 137.1, 147.1, 257.1 |
| Liquiritin | 294.2 | 18.88 | – | 417.1 | MS2 (–) [417.1]: 119.1, 135.1, 255.1 |
| Ononin | 430.1 | 46.22 | 279.2 | – | MS2 (+) [279.2]: 213.1, 237.1, 253.1, 269.1 |
| Enoxolone | 470.3 | 30.36 | 453.3 | – | MS2 (+) [453.3]: 189.1, 235.1, 453.1 |
| Neokestose | 504.2 | 22.89 | – | 549.2 | MS2 (–) [549.2]: 119.4, 135.4, 255.6, 549.2 |
| 2-[4-[3-[3,4-dihydroxy-4-(hydroxymethyl)oxolan-2-yl]oxy- 4,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxyphenyl]-7-hydroxy-2,3-dihydrochromen-4-one | 550.2 | 19.37 | – | 549.2 | MS2 (–) [549.2]: 119.2, 135.3, 255.2, 549.2 |
| 7-[3-[(2R,3R,4R)-3,4-dihydroxy-4-(hydroxymethyl)oxolan-2-yl]oxy-4,5-dihydroxy-6-(hydroxymethyl)oxan-2-yl]oxy-3-(4-methoxyphenyl)chromen-4-one | 562.2 | 22.51 | 563.2 | – | MS2 (+) [563.2]: 254.1, 269.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Lee, J.H.; Jo, E.H.; Lee, J.-S.; Kim, S.-H.; Lee, D.-S.; Park, M.C. Effects of Varying Ratios of Glycyrrhiza uralensis and Donkey Hide Gelatin Water Extracts on Dinitrochlorobenzene-Induced Atopic Dermatitis in NC/Nga Mice. Nutrients 2023, 15, 2094. https://doi.org/10.3390/nu15092094
Dong L, Lee JH, Jo EH, Lee J-S, Kim S-H, Lee D-S, Park MC. Effects of Varying Ratios of Glycyrrhiza uralensis and Donkey Hide Gelatin Water Extracts on Dinitrochlorobenzene-Induced Atopic Dermatitis in NC/Nga Mice. Nutrients. 2023; 15(9):2094. https://doi.org/10.3390/nu15092094
Chicago/Turabian StyleDong, Linsha, Ju Hyun Lee, Eun Heui Jo, Jin-Sil Lee, Seung-Hyung Kim, Dong-Sung Lee, and Min Cheol Park. 2023. "Effects of Varying Ratios of Glycyrrhiza uralensis and Donkey Hide Gelatin Water Extracts on Dinitrochlorobenzene-Induced Atopic Dermatitis in NC/Nga Mice" Nutrients 15, no. 9: 2094. https://doi.org/10.3390/nu15092094
APA StyleDong, L., Lee, J. H., Jo, E. H., Lee, J.-S., Kim, S.-H., Lee, D.-S., & Park, M. C. (2023). Effects of Varying Ratios of Glycyrrhiza uralensis and Donkey Hide Gelatin Water Extracts on Dinitrochlorobenzene-Induced Atopic Dermatitis in NC/Nga Mice. Nutrients, 15(9), 2094. https://doi.org/10.3390/nu15092094